

Anti-Inflammatory Effects of Miyako Bidens pilosa in a Mouse Model of Amyotrophic Lateral Sclerosis and Lipopolysaccharide-Stimulated BV-2 Microglia

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Results
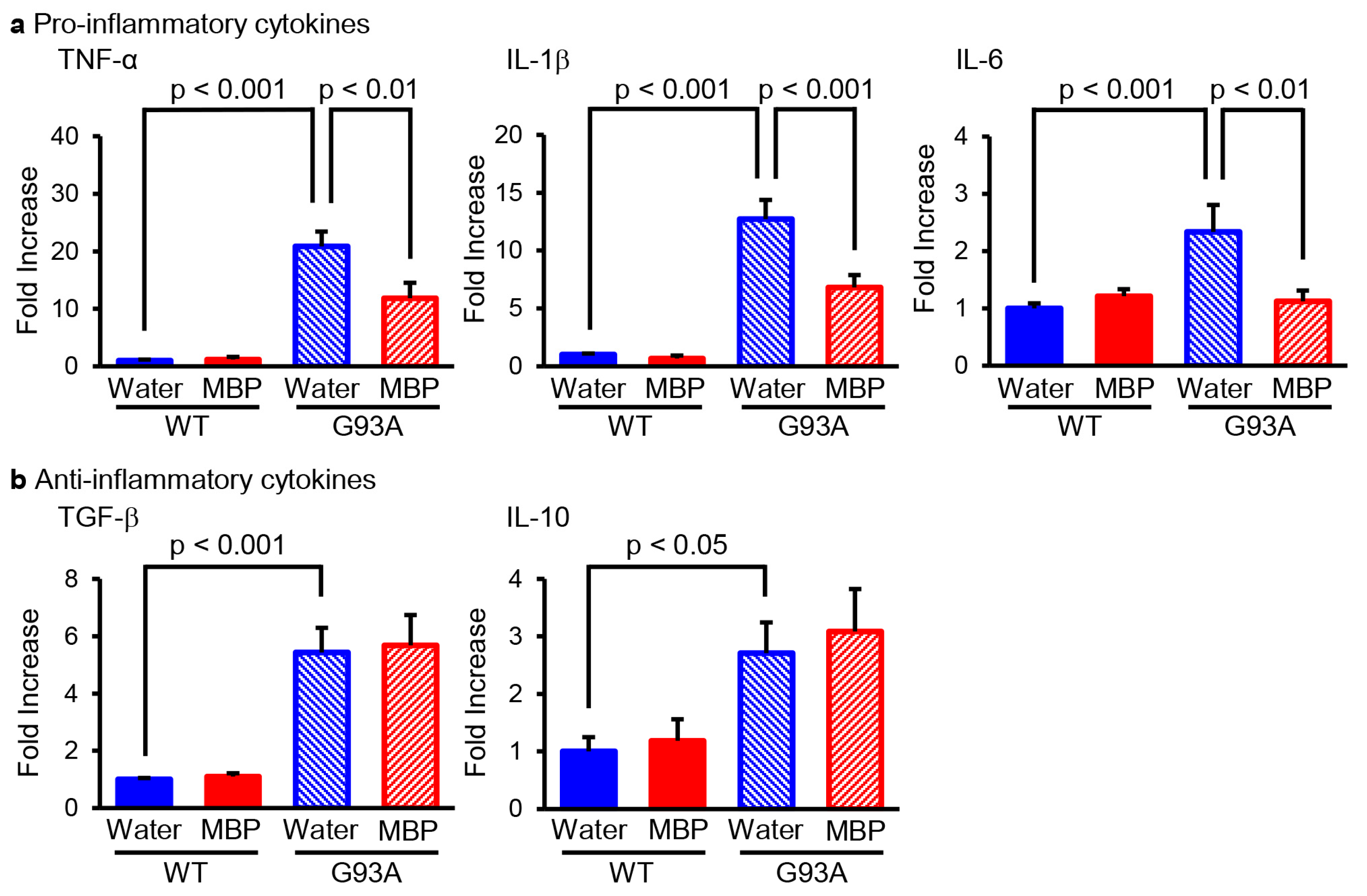
2.1. MBP Selectively Reduced the Production of M1-Related Pro-Inflammatory Cytokines in the Lumbar Spinal Cord of G93A Mice
2.2. MBP Selectively Inhibited M1-Microglia/Macrophage Polarization in the Lumbar Spinal Cord of G93A Mice
2.3. MBP Suppressed the Cell Proliferation of BV-2 Microglial Cells
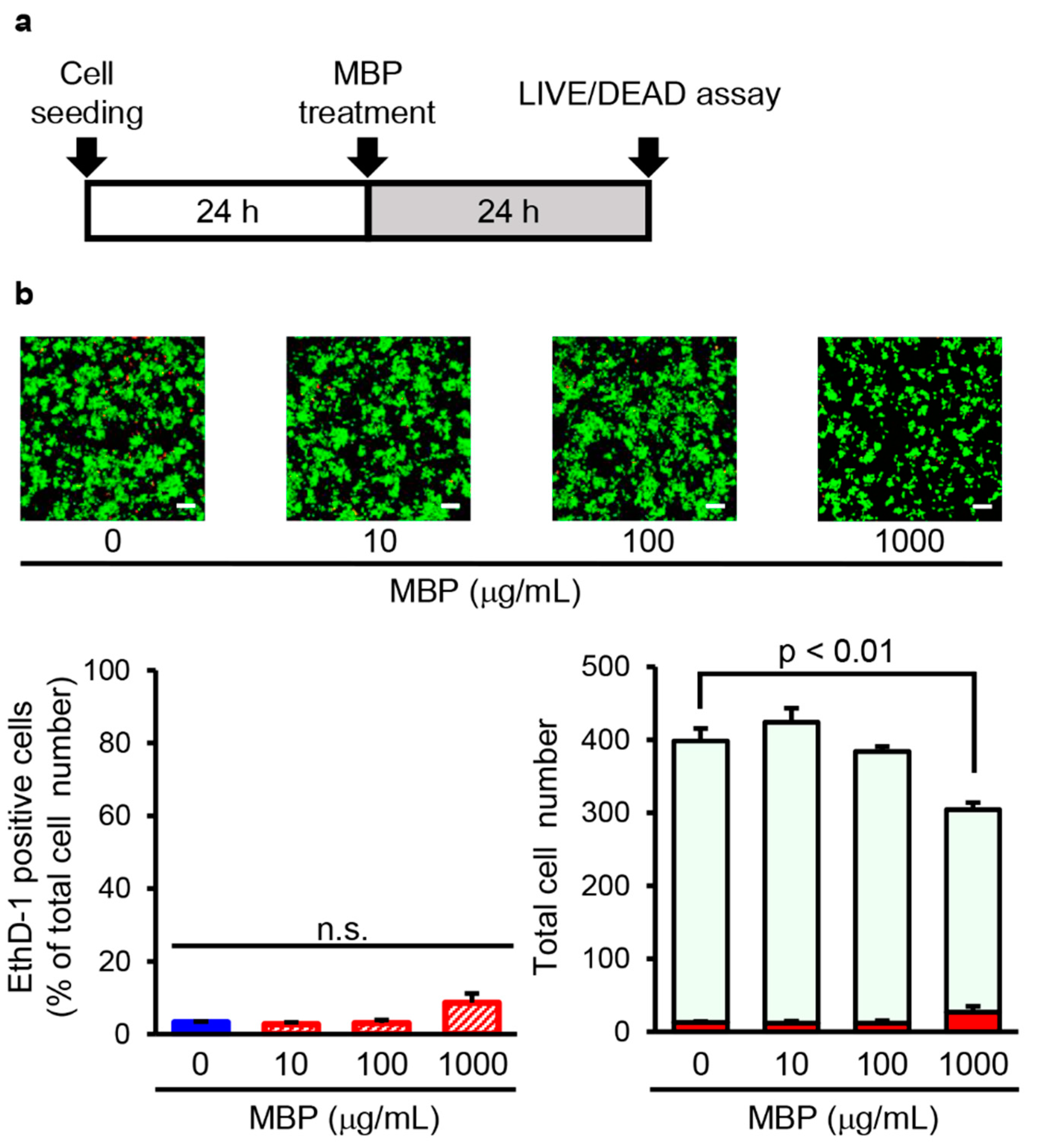
2.4. MBP Did Not Induce Cell Death in BV-2 Cells
2.5. MBP Inhibited LPS-Induced Pro-Inflammatory Cytokine Production in BV-2 Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. MBP Treatment
4.4. Real-Time PCR
4.5. MTT Reduction Assay and BrdU Incorporation Assay
4.6. LIVE/DEAD Assay
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosen, D.R.; Siddique, T.; Patterson, D.; Figlewicz, D.A.; Sapp, P.; Hentati, A.; Donaldson, D.; Goto, J.; O’Regan, J.P.; Deng, H.X.; et al. Mutations in Cu/Zn Superoxide Dismutase Gene Are Associated with Familial Amyotrophic Lateral Sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef]
- Gurney, M.E.; Pu, H.; Chiu, A.Y.; Dal Canto, M.C.; Polchow, C.Y.; Alexander, D.D.; Caliendo, J.; Hentati, A.; Kwon, Y.W.; Deng, H.-X.; et al. Motor Neuron Degeneration in Mice That Express a Human Cu, Zn Superoxide Dismutase Mutation. Science 1994, 264, 1772–1775. [Google Scholar] [CrossRef] [PubMed]
- Miana-Mena, F.J.; González-Mingot, C.; Larrodé, P.; Muñoz, M.J.; Oliván, S.; Fuentes-Broto, L.; Martínez-Ballarín, E.; Reiter, R.J.; Osta, R.; García, J.J. Monitoring Systemic Oxidative Stress in an Animal Model of Amyotrophic Lateral Sclerosis. J. Neurol. 2011, 258, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Kalmar, B.; Novoselov, S.; Gray, A.; Cheetham, M.E.; Margulis, B.; Greensmith, L. Late Stage Treatment with Arimoclomol Delays Disease Progression and Prevents Protein Aggregation in the SOD1G93A Mouse Model of ALS. J. Neurochem. 2008, 107, 339–350. [Google Scholar] [CrossRef]
- Cai, M.; Choi, S.-M.; Song, B.K.; Son, I.; Kim, S.; Yang, E.J. Scolopendra Subspinipes Mutilans Attenuates Neuroinflammation in Symptomatic hSOD1G93A mice. J. Neuroinflamm. 2013, 10, 131. [Google Scholar]
- Lee, J.; Hyeon, S.J.; Im, H.; Ryu, H.; Kim, Y.; Ryu, H. Astrocytes and Microglia as Non-Cell Autonomous Players in the Pathogenesis of ALS. Exp. Neurobiol. 2016, 25, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Boillée, S.; Yamanaka, K.; Lobsiger, C.S.; Copeland, N.G.; Jenkins, N.A.; Kassiotis, G.; Kollias, G.; Cleveland, D.W. Onset and Progression in Inherited ALS Determined by Motor Neurons and Microglia. Science 2006, 312, 1389–1392. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Chun, S.J.; Boillee, S.; Fujimori-Tonou, N.; Yamashita, H.; Gutmann, D.H.; Takahashi, R.; Misawa, H.; Cleveland, D.W. Astrocytes as Determinants of Disease Progression in Inherited Amyotrophic Lateral Sclerosis. Nat. Neurosci. 2008, 11, 251–253. [Google Scholar]
- Hooten, K.G.; Beers, D.R.; Zhao, W.; Appel, S.H. Protective and Toxic Neuroinflammation in Amyotrophic Lateral Sclerosis. Neurotherapeutics 2015, 12, 364. [Google Scholar] [CrossRef]
- Turner, M.R.; Cagnin, A.; Turkheimer, F.E.; Miller, C.C.J.; Shaw, C.E.; Brooks, D.J.; Leigh, P.N.; Banati, R.B. Evidence of Widespread Cerebral Microglial Activation in Amyotrophic Lateral Sclerosis: An [11C](R)-PK11195 Positron Emission Tomography Study. Neurobiol. Dis. 2004, 15, 601–609. [Google Scholar] [CrossRef]
- Corcia, P.; Tauber, C.; Vercoullie, J.; Arlicot, N.; Prunier, C.; Praline, J.; Nicolas, G.; Venel, Y.; Hommet, C.; Baulieu, J.L.; et al. Molecular Imaging of Microglial Activation in Amyotrophic Lateral Sclerosis. PLoS ONE 2012, 7, e52941. [Google Scholar] [CrossRef] [PubMed]
- Henkel, J.S.; Engelhardt, J.I.; Siklós, L.; Simpson, E.P.; Kim, S.H.; Pan, T.; Goodman, J.C.; Siddique, T.; Beers, D.R.; Appel, S.H. Presence of Dendritic Cells, MCP-1, and Activated Microglia/Macrophages in Amyotrophic Lateral Sclerosis Spinal Cord Tissue. Ann. Neurol. 2004, 55, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Beers, D.R.; Henkel, J.S.; Zhang, W.; Urushitani, M.; Julien, J.P.; Appel, S.H. Extracellular Mutant SOD1 Induces Microglial-Mediated Motoneuron Injury. Glia 2010, 58, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, J.; Li, S.; Al-Nusaif, M.; Zhou, Q.; Chen, S.; Le, W. Bone Marrow Stromal Cell Antigen 2: Is a Potential Neuroinflammation Biomarker of SOD1G93A Mouse Model of Amyotrophic Lateral Sclerosis in Pre-Symptomatic Stage. Front. Neurosci. 2022, 15, 788730. [Google Scholar] [CrossRef]
- Zhu, S.; Stavrovskaya, I.G.; Drozda, M.; Kim, B.Y.S.; Ona, V.; Li, M.; Sarang, S.; Liu, A.S.; Hartley, D.M.; Wu, D.C.; et al. Minocycline Inhibits Cytochrome c Release and Delays Progression of Amyotrophic Lateral Sclerosis in Mice. Nature 2002, 417, 74–78. [Google Scholar] [CrossRef]
- Kobayashi, K.; Imagama, S.; Ohgomori, T.; Hirano, K.; Uchimura, K.; Sakamoto, K.; Hirakawa, A.; Takeuchi, H.; Suzumura, A.; Ishiguro, N.; et al. Minocycline Selectively Inhibits M1 Polarization of Microglia. Cell Death Dis. 2013, 4, e525. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.Y.; Zhang, J.J.; Fu, Z.F.; Liu, Y.F.; Li, S.; Chen, S.; Le, W.D. Therapeutic Effects of Hirsutella Sinensis on the Disease Onset and Progression of Amyotrophic Lateral Sclerosis in SOD1G93A Transgenic Mouse Model. CNS Neurosci. Ther. 2020, 26, 90–100. [Google Scholar] [CrossRef]
- Beers, D.R.; Henkel, J.S.; Xiao, Q.; Zhao, W.; Wang, J.; Yen, A.A.; Siklos, L.; McKercher, S.R.; Appel, S.H. Wild-Type Microglia Extend Survival in PU.1 Knockout Mice with Familial Amyotrophic Lateral Sclerosis. Proc. Natl. Acad. Sci. USA 2006, 103, 16021–16026. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 75. [Google Scholar] [CrossRef]
- Lee, W.C.; Peng, C.C.; Chang, C.H.; Huang, S.H.; Chyau, C.C. Extraction of Antioxidant Components from Bidens pilosa Flowers and Their Uptake by Human Intestinal Caco-2 Cells. Molecules 2013, 18, 1582–1601. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, M.; Seyama, Y. Antiinflammatory and Antiallergic Activity of Bidens pilosa L. Var. Radiata SCHERFF. J. Health Sci. 2006, 52, 711–717. [Google Scholar] [CrossRef][Green Version]
- Horiuchi, M.; Wachi, H.; Seyama, Y. Effects of Bidens pilosa L. var. Radiata Scherff on Experimental Gastric Lesion. J. Nat. Med. 2010, 64, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, Y.; Kaneko, E.; Nango, H.; Miyagishi, H.; Ishige, K.; Ito, Y. Bidens pilosa Extract Administered after Symptom Onset Attenuates Glial Activation, Improves Motor Performance, and Prolongs Survival in a Mouse Model of Amyotrophic Lateral Sclerosis. Oxid. Med. Cell. Longev. 2020, 2020, 1020673. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.D.; Yang, E.J. Gamisoyo-San Ameliorates Neuroinflammation in the Spinal Cord of hSOD1G93A Transgenic Mice. Mediators Inflamm. 2018, 2018, 5897817. [Google Scholar] [CrossRef]
- Miyagishi, H.; Kosuge, Y.; Ishige, K.; Ito, Y. Expression of Microsomal Prostaglandin E Synthase-1 in the Spinal Cord in a Transgenic Mouse Model of Amyotrophic Lateral Sclerosis. J. Pharmacol. Sci. 2012, 118, 225–236. [Google Scholar] [CrossRef]
- Olsen, M.K.; Roberds, S.L.; Ellerbrock, B.R.; Fleck, T.J.; McKinley, D.K.; Gurney, M.E. Disease Mechanisms Revealed by Transcription Profiling in SOD1-G93A Transgenic Mouse Spinal Cord. Ann. Neurol. 2001, 50, 730–740. [Google Scholar] [CrossRef]
- Tada, S.; Okuno, T.; Yasui, T.; Nakatsuji, Y.; Sugimoto, T.; Kikutani, H.; Sakoda, S. Deleterious Effects of Lymphocytes at the Early Stage of Neurodegeneration in an Animal Model of Amyotrophic Lateral Sclerosis. J. Neuroinflamm. 2011, 8, 19. [Google Scholar] [CrossRef]
- Tada, S.; Okuno, T.; Hitoshi, Y.; Yasui, T.; Honorat, J.A.; Takata, K.; Koda, T.; Shimagami, H.; Chi-Jing, C.; Namba, A.; et al. Partial Suppression of M1 Microglia by Janus Kinase 2 Inhibitor Does Not Protect against Neurodegeneration in Animal Models of Amyotrophic Lateral Sclerosis. J. Neuroinflamm. 2014, 11, 179. [Google Scholar] [CrossRef]
- Cunha, C.; Santos, C.; Gomes, C.; Fernandes, A.; Correia, A.M.; Sebastião, A.M.; Vaz, A.R.; Brites, D. Downregulated Glia Interplay and Increased miRNA-155 as Promising Markers to Track ALS at an Early Stage. Mol. Neurobiol. 2018, 55, 4207–4224. [Google Scholar] [CrossRef]
- Xu, M.; Wang, J.; Zhang, X.; Yan, T.; Wu, B.; Bi, K.; Jia, Y. Polysaccharide from Schisandra Chinensis Acts via LRP-1 to Reverse Microglia Activation through Suppression of the NF-κB and MAPK Signaling. J. Ethnopharmacol. 2020, 256, 112798. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.; Ozmen, L.; Novelli, F.; Geuna, M.; Palestro, G.; Forni, G.; Garotta, G. Distribution of Interferon-γ Receptor in Human Tissues. Eur. J. Immunol. 1992, 22, 2403–2412. [Google Scholar] [CrossRef]
- Khurana Hershey, G.K. IL-13 Receptors and Signaling Pathways: An Evolving Web. J. Allergy Clin. Immunol. 2003, 111, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Deane, R.; Ali, Z.; Parisi, M.; Shapovalov, Y.; O’Banion, M.K.; Stojanovic, K.; Sagare, A.; Boillee, S.; Cleveland, D.W.; et al. ALS-Causing SOD1 Mutants Generate Vascular Changes Prior to Motor Neuron Degeneration. Nat. Neurosci. 2008, 11, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.A.; Sengillo, J.D.; Sullivan, J.S.; Henkel, J.S.; Appel, S.H.; Zlokovic, B.V. Blood–Spinal Cord Barrier Breakdown and Pericyte Reductions in Amyotrophic Lateral Sclerosis. Acta Neuropathol. 2013, 125, 111. [Google Scholar] [CrossRef]
- Epperly, M.W.; Fisher, R.; Rigatti, L.; Watkins, S.; Zhang, X.; Hou, W.; Shields, D.; Franicola, D.; Bayir, H.; Wang, H.; et al. Amelioration of Amyotrophic Lateral Sclerosis in SOD1G93A Mice by M2 Microglia from Transplanted Marrow. In Vivo 2019, 33, 675–688. [Google Scholar] [CrossRef]
- Noh, M.Y.; Cho, K.A.; Kim, H.; Kim, S.M.; Kim, S.H. Erythropoietin Modulates the Immune-Inflammatory Response of a SOD1G93A Transgenic Mouse Model of Amyotrophic Lateral Sclerosis (ALS). Neurosci. Lett. 2014, 574, 53–58. [Google Scholar] [CrossRef]
- Audet, J.N.; Gowing, G.; Paradis, R.; Soucy, G.; Julien, J.P. Ablation of Proliferating Cells in the CNS Exacerbates Motor Neuron Disease Caused by Mutant Superoxide Dismutase. PLoS ONE 2012, 7, e34932. [Google Scholar] [CrossRef]
- Chiarotto, G.B.; Cartarozzi, L.P.; Perez, M.; Biscola, N.P.; Spejo, A.B.; Gubert, F.; Francą Junior, M.; Mendez-Otero, R.; De Oliveira, A.L.R. Tempol Improves Neuroinflammation and Delays Motor Dysfunction in a Mouse Model (SOD1G93A) of ALS. J. Neuroinflamm. 2019, 16, 218. [Google Scholar] [CrossRef]
- Koh, S.H.; Kim, Y.; Kim, H.Y.; Won Cho, G.; Kim, K.S.; Kim, S.H. Recombinant Human Erythropoietin Suppresses Symptom Onset and Progression of G93A-SOD1 Mouse Model of ALS by Preventing Motor Neuron Death and Inflammation. Eur. J. Neurosci. 2007, 25, 1923–1930. [Google Scholar] [CrossRef]
- Zou, Y.H.; Guan, P.P.; Zhang, S.Q.; Guo, Y.S.; Wang, P. Rofecoxib Attenuates the Pathogenesis of Amyotrophic Lateral Sclerosis by Alleviating Cyclooxygenase-2-Mediated Mechanisms. Front. Neurosci. 2020, 14, 817. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, K.R.; Stoica, B.A.; Fricke, S.; Di Giovanni, S.; Faden, A.I. Cell Cycle Activation Contributes to Post-Mitotic Cell Death and Secondary Damage after Spinal Cord Injury. Brain 2007, 130, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.; Gao, Y.; Liu, S.; Ye, E.; Li, Y.; Jin, X.; Yang, H.; Yang, Z. Cyclin-Dependent Kinase Inhibitor Flavopiridol Promotes Remyelination in a Cuprizone Induced Demyelination Model. Cell Cycle 2016, 15, 2780–2791. [Google Scholar] [CrossRef]
- Kim, J.E.; Park, H.; Choi, S.H.; Kong, M.J.; Kang, T.C. Roscovitine Attenuates Microglia Activation and Monocyte Infiltration via P38 MAPK Inhibition in the Rat Frontoparietal Cortex Following Status Epilepticus. Cells 2019, 8, 746. [Google Scholar] [CrossRef] [PubMed]
- Akiko, K.; Yoshiyuki, S.; Eiji, U.; Takanori, K.; Makio, S.; Daisuke, T.; Genjiro, K. Studies on the Antioxidant Active Constituents of the Dried Powder from Bidens pilosa L. var. radiata SCH. Nat. Med. 2003, 57, 100–104. [Google Scholar]
- Matsumoto, T.; Horiuchi, M.; Kamata, K.; Seyama, Y. Effects of Bidens pilosa L. Var. radiata SCHERFF Treated with Enzyme on Histamine-Induced Contraction of Guinea Pig Ileum and on Histamine Release from Mast Cells. J. Smooth Muscle Res. 2009, 45, 75–86. [Google Scholar] [CrossRef]
- Nakatani, Y.; Yaguchi, M.; Ogino, K.; Noguchi, R.; Yamamoto, N.; Amano, T. Duloxetine Ameliorates Lipopolysaccharide-Induced Microglial Activation by Suppressing INOS Expression in BV-2 Microglial Cells. Psychopharmacology 2022, 239, 3133–3143. [Google Scholar] [CrossRef]
- Kosuge, Y.; Miyagishi, H.; Shinomiya, T.; Nishiyama, K.; Suzuki, S.; Osada, N.; Ishige, K.; Okubo, M.; Kawaguchi, M.; Ito, Y. Characterization of Motor Neuron Prostaglandin E2 EP3 Receptor Isoform in a Mouse Model of Amyotrophic Lateral Sclerosis. Biol. Pharm. Bull. 2015, 38, 1964–1968. [Google Scholar] [CrossRef]
- Nango, H.; Kosuge, Y.; Miyagishi, H.; Sugawa, K.; Ito, Y.; Ishige, K. Prostaglandin E2 Facilitates Neurite Outgrowth in a Motor Neuron-like Cell Line, NSC-34. J. Pharmacol. Sci. 2017, 135, 64–71. [Google Scholar] [CrossRef]
- Kosuge, Y.; Nango, H.; Kasai, H.; Yanagi, T.; Mawatari, T.; Nishiyama, K.; Miyagishi, H.; Ishige, K.; Ito, Y. Generation of Cellular Reactive Oxygen Species by Activation of the EP2 Receptor Contributes to Prostaglandin E2-Induced Cytotoxicity in Motor Neuron-Like NSC-34 Cells. Oxid. Med. Cell. Longev. 2020, 2020, 6101838. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | |
|---|---|---|
| IFN-γR | Forward | TGACGGGAGCACCTGTTACAC |
| Reverse | TTTCGACCGTATGTTTCGTATGTAG | |
| IL-13R | Forward | CAGTCTTGCAGCATGGGAACA |
| Reverse | TGAGTCCCTAAGGCCTGGAGATTAC | |
| TNF-α | Forward | AGCCCACGTCGTAGCAAACCAC |
| Reverse | AGGTACAACCCATCGGCTGGCA | |
| IL-1β | Forward | GCAACTGTTCCTGAACTCAACT |
| Reverse | ATCTTTTGGGGTCCGTCAACT | |
| IL-6 | Forward | GAGGATACCACTCCCAACAGACC |
| Reverse | AAGTGCATCATCGTTGTTCATACA | |
| TGF-β | Forward | TGACGTCACTGGAGTTGTACGG |
| Reverse | GGTTCATGTCATGGATGGTGC | |
| IL-10 | Forward | GCATGGCCCAGAAATCAAGG |
| Reverse | GAGAAATCGATGACAGCGCC | |
| GAPDH | Forward | TGTGTCCGTCGTGGATCTGA |
| Reverse | TTGCTGTTGAAGTCGCAGGAG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuruta, K.; Shidara, T.; Miyagishi, H.; Nango, H.; Nakatani, Y.; Suzuki, N.; Amano, T.; Suzuki, T.; Kosuge, Y. Anti-Inflammatory Effects of Miyako Bidens pilosa in a Mouse Model of Amyotrophic Lateral Sclerosis and Lipopolysaccharide-Stimulated BV-2 Microglia. Int. J. Mol. Sci. 2023, 24, 13698. https://doi.org/10.3390/ijms241813698
Tsuruta K, Shidara T, Miyagishi H, Nango H, Nakatani Y, Suzuki N, Amano T, Suzuki T, Kosuge Y. Anti-Inflammatory Effects of Miyako Bidens pilosa in a Mouse Model of Amyotrophic Lateral Sclerosis and Lipopolysaccharide-Stimulated BV-2 Microglia. International Journal of Molecular Sciences. 2023; 24(18):13698. https://doi.org/10.3390/ijms241813698
Chicago/Turabian StyleTsuruta, Komugi, Takato Shidara, Hiroko Miyagishi, Hiroshi Nango, Yoshihiko Nakatani, Naoto Suzuki, Taku Amano, Toyofumi Suzuki, and Yasuhiro Kosuge. 2023. "Anti-Inflammatory Effects of Miyako Bidens pilosa in a Mouse Model of Amyotrophic Lateral Sclerosis and Lipopolysaccharide-Stimulated BV-2 Microglia" International Journal of Molecular Sciences 24, no. 18: 13698. https://doi.org/10.3390/ijms241813698
APA StyleTsuruta, K., Shidara, T., Miyagishi, H., Nango, H., Nakatani, Y., Suzuki, N., Amano, T., Suzuki, T., & Kosuge, Y. (2023). Anti-Inflammatory Effects of Miyako Bidens pilosa in a Mouse Model of Amyotrophic Lateral Sclerosis and Lipopolysaccharide-Stimulated BV-2 Microglia. International Journal of Molecular Sciences, 24(18), 13698. https://doi.org/10.3390/ijms241813698