
High Tumor-Infiltrating Lymphocyte Count Is Associated with Distinct Gene Expression Profile and Longer Patient Survival in Advanced Ovarian Cancer

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
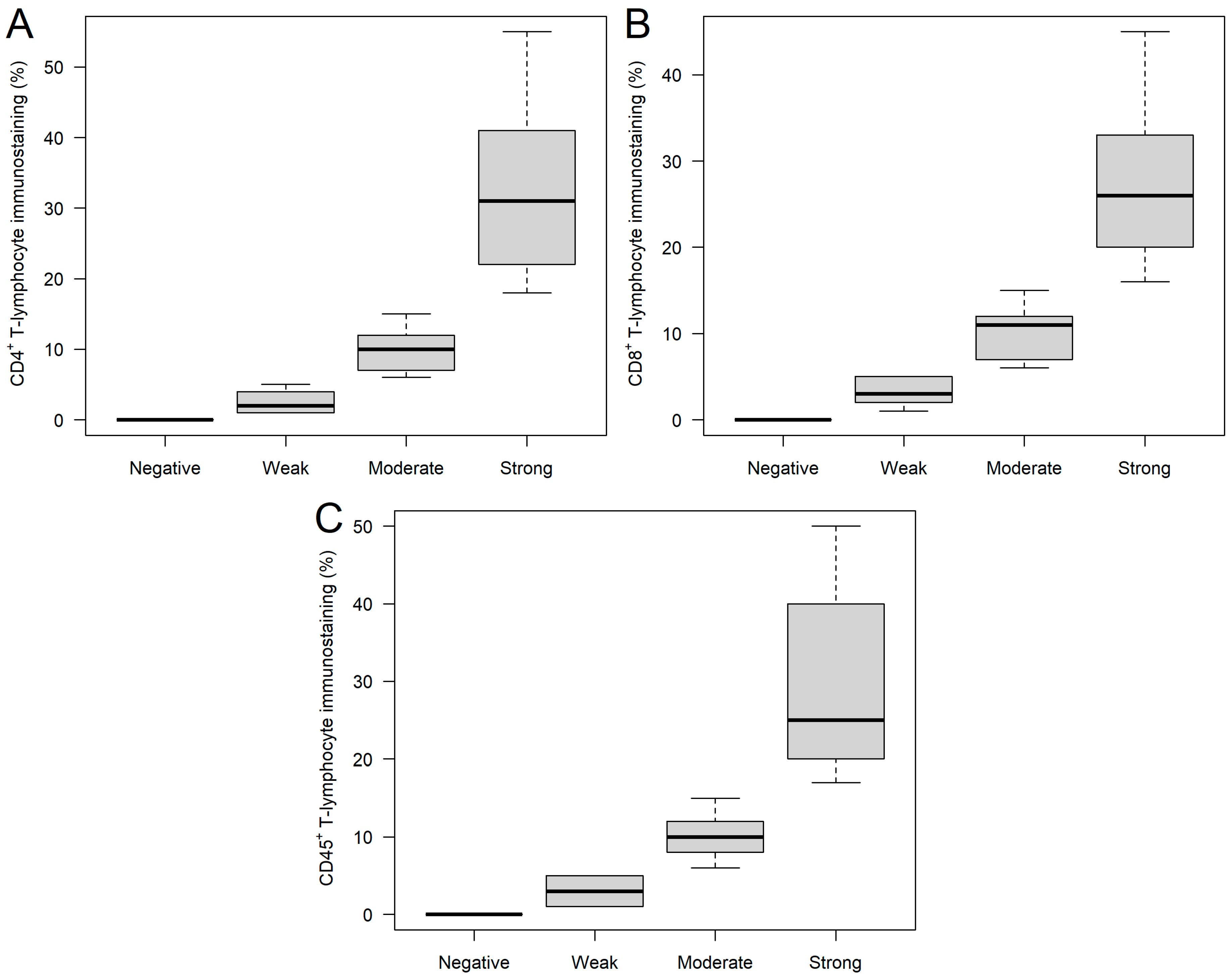
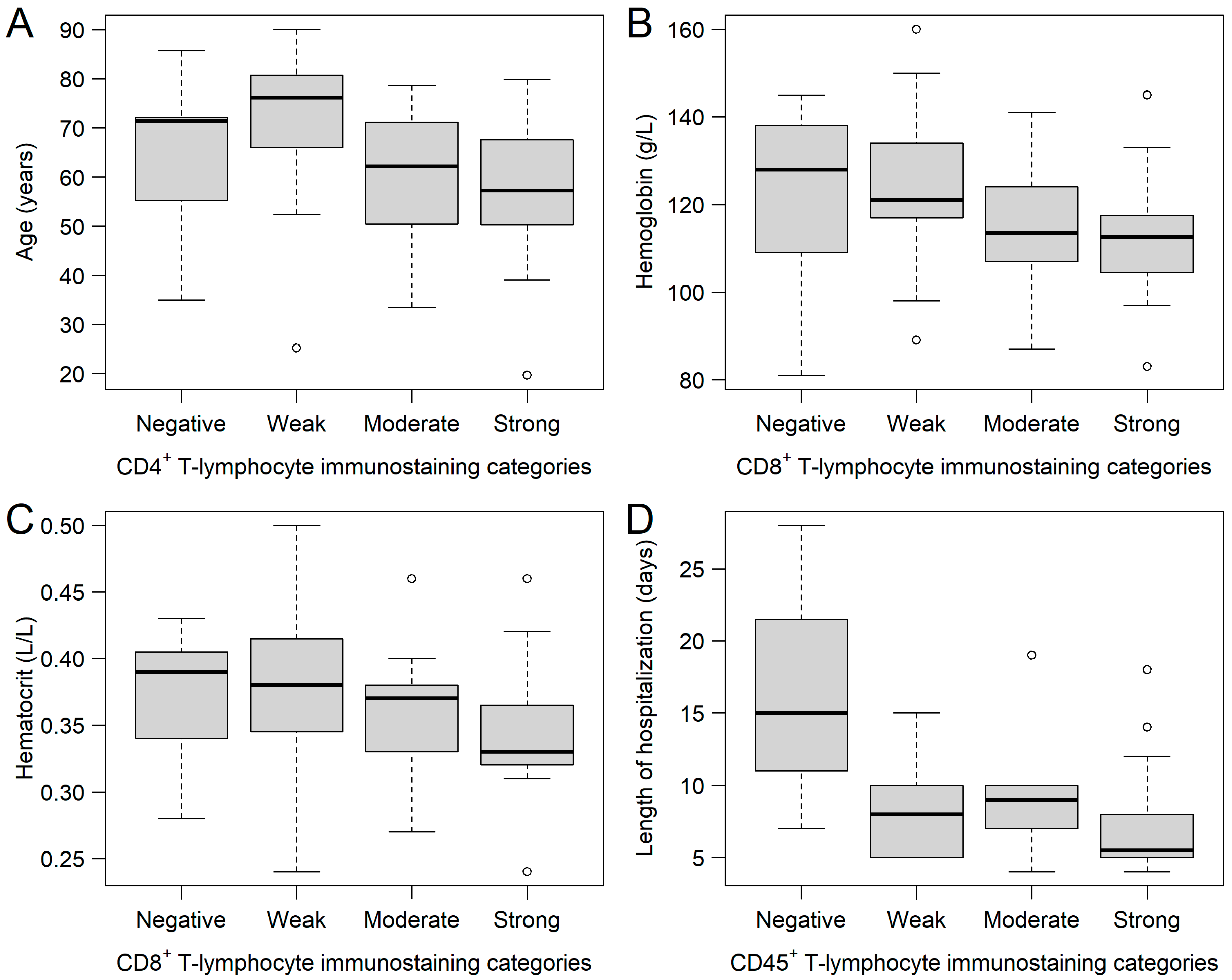
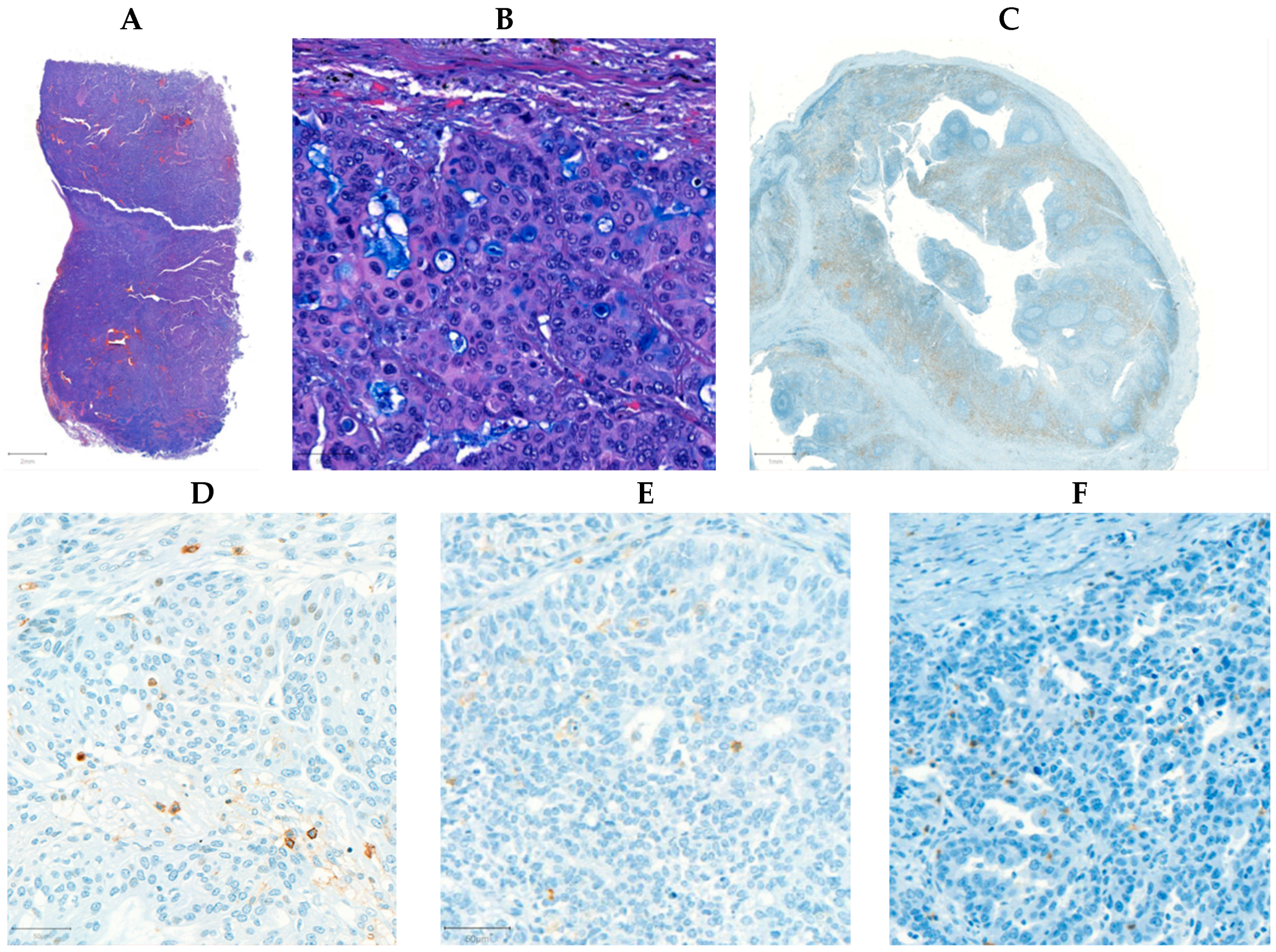
2.1. CD4+, CD8+, and CD45+ Immunohistochemical Analysis of Ovarian Tumor Samples
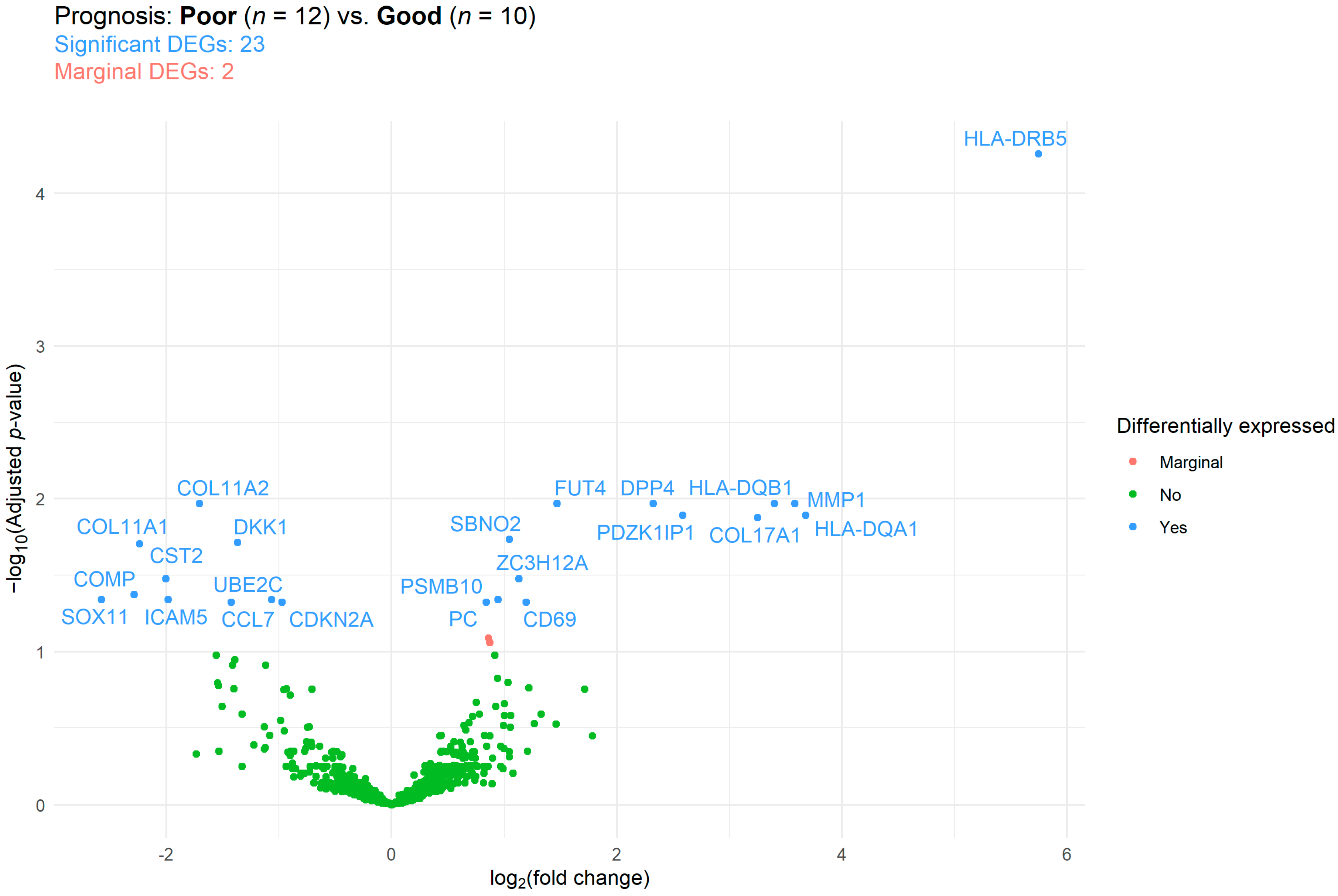
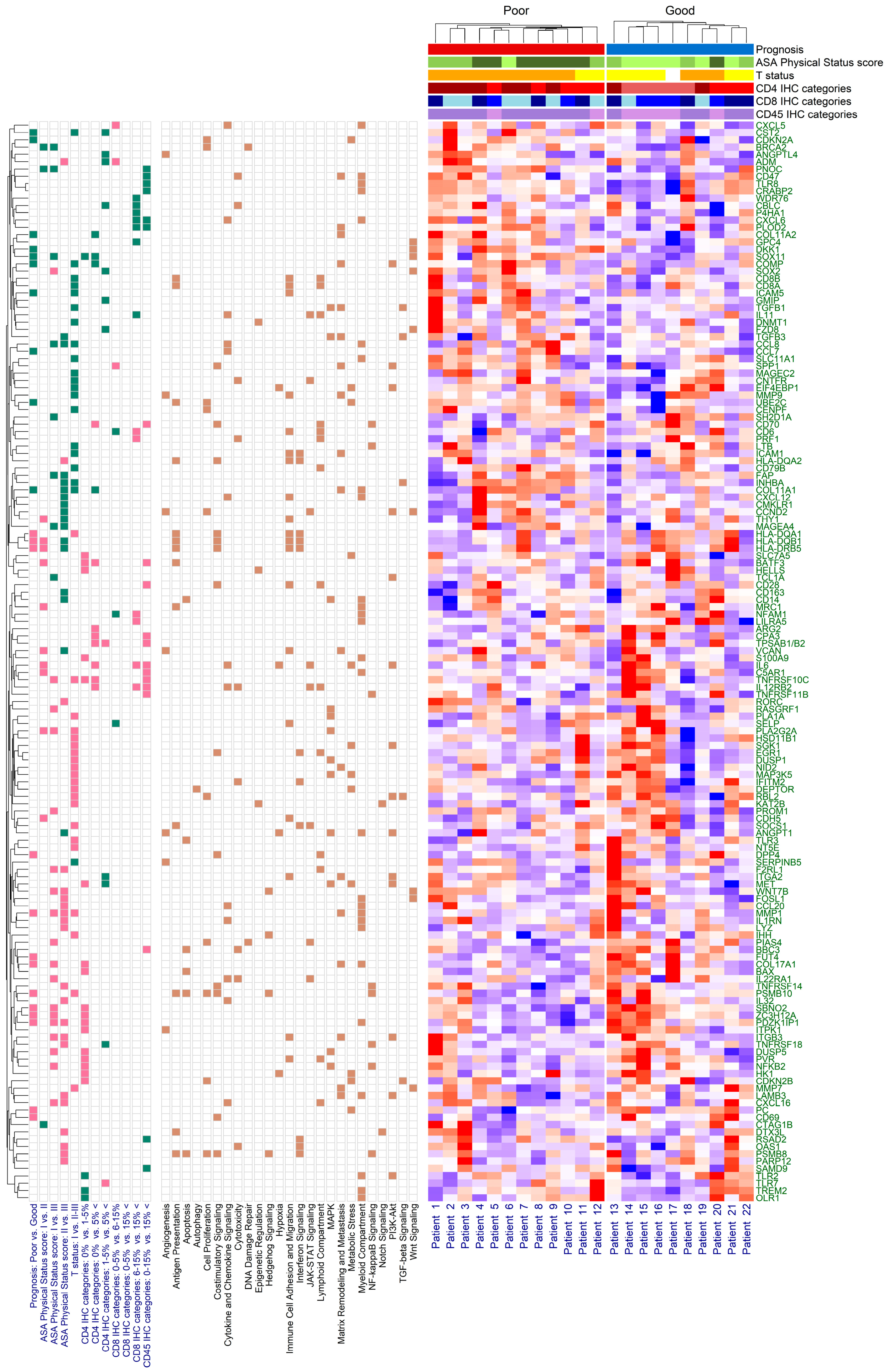
2.2. Immune Panel Gene Expression Analysis
2.3. Comparison of NanoString Data with the “Kaplan–Meier Plotter” Web Application
3. Discussion
Limitations
4. Materials and Methods
4.1. Patients and Study Design
4.2. Immunohistochemical Staining of OC Samples
4.3. NanoString nCounter PanCancer IO 360 Panel
4.4. Description of Clinicopathological Data
4.5. Data Collection from the Kaplan–Meier Plotter Web Application
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Burges, A.; Schmalfeldt, B. Ovarian cancer: Diagnosis and treatment. Dtsch. Ärzteblatt Int. 2011, 108, 635–641. [Google Scholar] [CrossRef]
- Stewart, C.; Ralyea, C.; Lockwood, S. Ovarian Cancer: An Integrated Review. Semin. Oncol. Nurs. 2019, 35, 151–156. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Coussens, L.M.; Zitvogel, L.; Palucka, A.K. Neutralizing tumor-promoting chronic inflammation: A magic bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, C.; Zhou, S. Targeting tumor microenvironment in ovarian cancer: Premise and promise. Biochim. Biophys. Acta BBA-Rev. Cancer 2020, 1873, 188361. [Google Scholar] [CrossRef]
- Roxburgh, C.S.; McMillan, D.C. The role of the in situ local inflammatory response in predicting recurrence and survival in patients with primary operable colorectal cancer. Cancer Treat. Rev. 2012, 38, 451–466. [Google Scholar] [CrossRef]
- Gavalas, N.G.; Karadimou, A.; Dimopoulos, M.A.; Bamias, A. Immune response in ovarian cancer: How is the immune system involved in prognosis and therapy: Potential for treatment utilization. Clin. Dev. Immunol. 2010, 2010, 791603. [Google Scholar] [CrossRef]
- Singh, M.; Loftus, T.; Webb, E.; Benencia, F. Minireview: Regulatory T Cells and Ovarian Cancer. Immunol. Investig. 2016, 45, 712–720. [Google Scholar] [CrossRef]
- Zou, R.; Jiang, Q.; Jin, T.; Chen, M.; Yao, L.; Ding, H. Pan-cancer analyses and molecular subtypes based on the cancer-associated fibroblast landscape and tumor microenvironment infiltration characterization reveal clinical outcome and immunotherapy response in epithelial ovarian cancer. Front. Immunol. 2022, 13, 956224. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Yang, J.; Wang, Z.; Zhang, Z.; Peng, J.; Wang, Y.; Hong, L. A novel tumor mutational burden-based risk model predicts prognosis and correlates with immune infiltration in ovarian cancer. Front. Immunol. 2022, 13, 943389. [Google Scholar] [CrossRef]
- Jordan, K.R.; Sikora, M.J.; Slansky, J.E.; Minic, A.; Richer, J.K.; Moroney, M.R.; Hu, J.; Wolsky, R.J.; Watson, Z.L.; Yamamoto, T.M.; et al. The Capacity of the Ovarian Cancer Tumor Microenvironment to Integrate Inflammation Signaling Conveys a Shorter Disease-free Interval. Clin. Cancer Res. 2020, 26, 6362–6373. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, M.C.W.; Milne, K.; Pedersen, M.; Hasselager, T.; Olsen, L.R.; Anglesio, M.S.; Borch, T.H.; Kennedy, M.; Briggs, G.; Ledoux, S.; et al. Changes in the Tumor Immune Microenvironment during Disease Progression in Patients with Ovarian Cancer. Cancers 2020, 12, 3828. [Google Scholar] [CrossRef] [PubMed]
- Khalique, S.; Nash, S.; Mansfield, D.; Wampfler, J.; Attygale, A.; Vroobel, K.; Kemp, H.; Buus, R.; Cottom, H.; Roxanis, I.; et al. Quantitative Assessment and Prognostic Associations of the Immune Landscape in Ovarian Clear Cell Carcinoma. Cancers 2021, 13, 3854. [Google Scholar] [CrossRef] [PubMed]
- James, N.E.; Woodman, M.; De La Cruz, P.; Eurich, K.; Ozsoy, M.A.; Schorl, C.; Hanley, L.C.; Ribeiro, J.R. Adaptive transcriptomic and immune infiltrate responses in the tumor immune microenvironment following neoadjuvant chemotherapy in high grade serous ovarian cancer reveal novel prognostic associations and activation of pro-tumorigenic pathways. Front. Immunol. 2022, 13, 965331. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Li, X.; Zhang, W.; Fan, J.; Zhou, Y.; Li, W.; Yin, J.; Yang, X.; Guo, E.; Li, X.; et al. Spatial heterogeneity of infiltrating T cells in high-grade serous ovarian cancer revealed by multi-omics analysis. Cell Rep. Med. 2022, 3, 100856. [Google Scholar] [CrossRef]
- Meagher, N.S.; Hamilton, P.; Milne, K.; Thornton, S.; Harris, B.; Weir, A.; Alsop, J.; Bisinoto, C.; Brenton, J.D.; Brooks-Wilson, A.; et al. Profiling the immune landscape in mucinous ovarian carcinoma. Gynecol. Oncol. 2023, 168, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Daabiss, M. American Society of Anaesthesiologists physical status classification. Indian J. Anaesth. 2011, 55, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B. Discovery and ranking of the most robust prognostic biomarkers in serous ovarian cancer. Geroscience 2023, 45, 1889–1898. [Google Scholar] [CrossRef]
- Kazemi, M.H.; Sadri, M.; Najafi, A.; Rahimi, A.; Baghernejadan, Z.; Khorramdelazad, H.; Falak, R. Tumor-infiltrating lymphocytes for treatment of solid tumors: It takes two to tango? Front. Immunol. 2022, 13, 1018962. [Google Scholar] [CrossRef]
- Lin, B.; Du, L.; Li, H.; Zhu, X.; Cui, L.; Li, X. Tumor-infiltrating lymphocytes: Warriors fight against tumors powerfully. Biomed. Pharmacother. 2020, 132, 110873. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, L.; Xu, B.; Xiong, Y.; Yang, M.; Rui, X.; Shi, L.; Wu, C.; Jiang, J.; Lu, B. Higher Numbers of T-Bet+ Tumor-Infiltrating Lymphocytes Associate with Better Survival in Human Epithelial Ovarian Cancer. Cell. Physiol. Biochem. 2017, 41, 475–483. [Google Scholar] [CrossRef]
- Liu, J.; Tan, Z.; He, J.; Jin, T.; Han, Y.; Hu, L.; Song, J.; Huang, S. Identification of three molecular subtypes based on immune infiltration in ovarian cancer and its prognostic value. Biosci. Rep. 2020, 40, BSR20201431. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Hua, D.; Li, J.; Zhang, X.; Zhang, Z.; Zhang, B.; Bei, T.; Cui, L.; Chen, S.; Wang, S.; et al. Tumor immune microenvironment changes are associated with response to neoadjuvant chemotherapy and long-term survival benefits in advanced epithelial ovarian cancer: A pilot study. Front. Immunol. 2023, 14, 1022942. [Google Scholar] [CrossRef]
- Ovarian Tumor Tissue Analysis Consortium; Goode, E.L.; Block, M.S.; Kalli, K.R.; Vierkant, R.A.; Chen, W.; Fogarty, Z.C.; Gentry-Maharaj, A.; Toloczko, A.; Hein, A.; et al. Dose-Response Association of CD8+ Tumor-Infiltrating Lymphocytes and Survival Time in High-Grade Serous Ovarian Cancer. JAMA Oncol. 2017, 3, e173290. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.C.; Bean, S.M.; Whitaker, R.S.; Kondoh, E.; Baba, T.; Fujii, S.; Marks, J.R.; Dressman, H.K.; Murphy, S.K.; Berchuck, A. Ovarian cancer tumor infiltrating T-regulatory (T(reg)) cells are associated with a metastatic phenotype. Gynecol. Oncol. 2010, 116, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, Q.; Zhu, Y.; Huang, Y.; Qin, J.; Wu, X.; Zhang, S. Lymphocyte and macrophage infiltration in omental metastases indicates poor prognosis in advance stage epithelial ovarian cancer. J. Int. Med. Res. 2021, 49, 3000605211066245. [Google Scholar] [CrossRef]
- Yang, L.; Wang, S.; Zhang, Q.; Pan, Y.; Lv, Y.; Chen, X.; Zuo, Y.; Hao, D. Clinical significance of the immune microenvironment in ovarian cancer patients. Mol. Omics 2018, 14, 341–351. [Google Scholar] [CrossRef]
- Wang, Q.; Lou, W.; Di, W.; Wu, X. Prognostic value of tumor PD-L1 expression combined with CD8(+) tumor infiltrating lymphocytes in high grade serous ovarian cancer. Int. Immunopharmacol. 2017, 52, 7–14. [Google Scholar] [CrossRef]
- Devlin, M.J.; Miller, R.; Laforets, F.; Kotantaki, P.; Garsed, D.W.; Kristeleit, R.; Bowtell, D.D.; McDermott, J.; Maniati, E.; Balkwill, F.R. The Tumor Microenvironment of Clear-Cell Ovarian Cancer. Cancer Immunol. Res. 2022, 10, 1326–1339. [Google Scholar] [CrossRef]
- Gallego, A.; Berjon, A.; Mendiola, M.; Diez, J.; Castelo, B.; Hernandez, A.; Hardisson, D.; Batlle, J.F.; Garcia, M.J.; Redondo, A. Tumor infiltrating lymphocytes (TILs) in endometrioid and clear cell ovarian carcinoma: Characterization, association with mismatch repair system deficiency and endometriosis, and prognostic implications. J. Clin. Oncol. 2021, 39, e17549. [Google Scholar] [CrossRef]
- Santoiemma, P.P.; Reyes, C.; Wang, L.P.; McLane, M.W.; Feldman, M.D.; Tanyi, J.L.; Powell, D.J., Jr. Systematic evaluation of multiple immune markers reveals prognostic factors in ovarian cancer. Gynecol. Oncol. 2016, 143, 120–127. [Google Scholar] [CrossRef]
- Milne, K.; Kobel, M.; Kalloger, S.E.; Barnes, R.O.; Gao, D.; Gilks, C.B.; Watson, P.H.; Nelson, B.H. Systematic analysis of immune infiltrates in high-grade serous ovarian cancer reveals CD20, FoxP3 and TIA-1 as positive prognostic factors. PLoS ONE 2009, 4, e6412. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.P.; Balmaceda, C.; Bravo, M.L.; Kato, S.; Villarroel, A.; Owen, G.I.; Roa, J.C.; Cuello, M.A.; Ibanez, C. Patient inflammatory status and CD4+/CD8+ intraepithelial tumor lymphocyte infiltration are predictors of outcomes in high-grade serous ovarian cancer. Gynecol. Oncol. 2018, 151, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.; Olson, S.H.; Ahn, J.; Bundy, B.; Nishikawa, H.; Qian, F.; Jungbluth, A.A.; Frosina, D.; Gnjatic, S.; Ambrosone, C.; et al. Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 18538–18543. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Chen, R.; Bai, Y.; Lu, X. The prognostic value of tumor-infiltrating T lymphocytes in ovarian cancer. Oncotarget 2017, 8, 15621–15631. [Google Scholar] [CrossRef] [PubMed]
- Arman Karakaya, Y.; Atigan, A.; Guler, O.T.; Demiray, A.G.; Bir, F. The relation of CD3, CD4, CD8 and PD-1 expression with tumor type and prognosis in epithelial ovarian cancers. Ginekol. Pol. 2021, 92, 344–351. [Google Scholar] [CrossRef]
- Yildirim, N.; Akman, L.; Acar, K.; Demir, S.; Ozkan, S.; Alan, N.; Zekioglu, O.; Terek, M.C.; Ozdemir, N.; Ozsaran, A. Do tumor-infiltrating lymphocytes really indicate favorable prognosis in epithelial ovarian cancer? Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 215, 55–61. [Google Scholar] [CrossRef]
- Preston, C.C.; Maurer, M.J.; Oberg, A.L.; Visscher, D.W.; Kalli, K.R.; Hartmann, L.C.; Goode, E.L.; Knutson, K.L. The ratios of CD8+ T cells to CD4+CD25+ FOXP3+ and FOXP3- T cells correlate with poor clinical outcome in human serous ovarian cancer. PLoS ONE 2013, 8, e80063. [Google Scholar] [CrossRef]
- Emens, L.; Molinero, L.; Loi, S.; Rugo, H.S.; Schneeweiss, A.; Diéras, V.; Iwata, H.; Barrios, C.H.; Nechaeva, M.; Duc, A.N.; et al. Atezolizumab and nab-Paclitaxel in Advanced Triple-Negative Breast Cancer: Biomarker Evaluation of the IMpassion130 Study. J. Natl. Cancer Inst. 2021, 113, 1005–1016. [Google Scholar] [CrossRef]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T Cells, Recurrence, and Survival in Epithelial Ovarian Cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Bound, N.T.; Vandenberg, C.J.; Kartikasari, A.E.R.; Plebanski, M.; Scott, C.L. Improving PARP inhibitor efficacy in high-grade serous ovarian carcinoma: A focus on the immune system. Front. Genet. 2022, 13, 886170. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.A.; Allison, K.H.; Garcia, R.L.; Gray, H.J.; Goff, B.A.; Swisher, E.M. Intratumoral T cells, tumor-associated macrophages, and regulatory T cells: Association with p53 mutations, circulating tumor DNA and survival in women with ovarian cancer. Gynecol. Oncol. 2008, 109, 215–219. [Google Scholar] [CrossRef]
- Montfort, A.; Barker-Clarke, R.J.; Piskorz, A.M.; Supernat, A.; Moore, L.; Al-Khalidi, S.; Böhm, S.; Pharoah, P.; McDermott, J.; Balkwill, F.R.; et al. Combining measures of immune infiltration shows additive effect on survival prediction in high-grade serous ovarian carcinoma. Br. J. Cancer 2020, 122, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, J.; Zhang, C.; Yang, H.; Qiu, H.; Li, J.; Liu, Y.; Qin, L.; Wang, L.; Hao, S.; et al. Infiltration of dendritic cells and T lymphocytes predicts favorable outcome in epithelial ovarian cancer. Cancer Gene Ther. 2015, 22, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Xu, Q.; Zhang, X.; Yang, M.; Wang, Y.; He, M.; Lu, J.; Liu, H. Spatial cytotoxic and memory T cells in tumor predict superior survival outcomes in patients with high-grade serous ovarian cancer. Cancer Med. 2021, 10, 3905–3918. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Chen, M.; Ma, X. The Immune Subtype Contributes to Distinct Overall Survival for Ovarian Cancer Patients With Platinum-Based Adjuvant Therapy. Front. Immunol. 2022, 13, 872991. [Google Scholar] [CrossRef]
- Gill, C.M.; D’andrea, M.R.; Tomita, S.; Suhner, J.; Umphlett, M.; Zakashansky, K.; Blank, S.V.; Tsankova, N.; Shrivastava, R.K.; Fowkes, M.; et al. Tumor immune microenvironment in brain metastases from gynecologic malignancies. Cancer Immunol. Immunother. 2021, 70, 2951–2960. [Google Scholar] [CrossRef]
- Arend, R.C.; Londoño, A.I.; Montgomery, A.M.; Smith, H.J.; Dobbin, Z.C.; Katre, A.A.; Martinez, A.; Yang, E.S.; Alvarez, R.D.; Huh, W.K.; et al. Molecular Response to Neoadjuvant Chemotherapy in High-Grade Serous Ovarian Carcinoma. Mol. Cancer Res. 2018, 16, 813–824. [Google Scholar] [CrossRef]
- Heong, V.; Tan, T.Z.; Miwa, M.; Ye, J.; Lim, D.; Herrington, C.S.; Iida, Y.; Yano, M.; Yasuda, M.; Ngoi, N.Y.; et al. A multi-ethnic analysis of immune-related gene expression signatures in patients with ovarian clear cell carcinoma. J. Pathol. 2021, 255, 285–295. [Google Scholar] [CrossRef]
- Rocconi, R.P.; Stanbery, L.; da Silva, L.M.; Barrington, R.A.; Aaron, P.; Manning, L.; Horvath, S.; Wallraven, G.; Bognar, E.; Walter, A.; et al. Long-Term Follow-Up of Gemogenovatucel-T (Vigil) Survival and Molecular Signals of Immune Response in Recurrent Ovarian Cancer. Vaccines 2021, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Weberpals, J.I.; Pugh, T.J.; Marco-Casanova, P.; Goss, G.D.; Wright, N.A.; Rath, P.; Torchia, J.; Fortuna, A.; Jones, G.N.; Roudier, M.P.; et al. Tumor genomic, transcriptomic, and immune profiling characterizes differential response to first-line platinum chemotherapy in high grade serous ovarian cancer. Cancer Med. 2021, 10, 3045–3058. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Lu, C.; Zhou, H.; Liu, Q.; Yang, J. Differential molecular pathway expression according to chemotherapeutic response in ovarian clear cell carcinoma. BMC Womens Health 2023, 23, 298. [Google Scholar] [CrossRef]
- Mairinger, F.; Bankfalvi, A.; Schmid, K.W.; Mairinger, E.; Mach, P.; Walter, R.F.; Borchert, S.; Kasimir-Bauer, S.; Kimmig, R.; Buderath, P. Digital Immune-Related Gene Expression Signatures In High-Grade Serous Ovarian Carcinoma: Developing Prediction Models For Platinum Response. Cancer Manag. Res. 2019, 11, 9571–9583. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.U.; Kim, A.; Kim, J.Y.; Kim, K.H.; Hwang, C.; Lee, S.J.; Park, W.Y.; Jung, S.; Choi, H.J.; Kim, K. Differences in immune-related gene expressions and tumor-infiltrating lymphocytes according to chemotherapeutic response in ovarian high-grade serous carcinoma. J. Ovarian Res. 2020, 13, 65. [Google Scholar] [CrossRef]
- Jordan, S.E.; Saad, H.; Covarrubias, A.S.; Siemon, J.; Pearson, J.M.; Slomovitz, B.M.; Huang, M.; Pinto, A.; Schlumbrecht, M.; George, S.H. mRNA expression in low grade serous ovarian cancer: Results of a nanoString assay in a diverse population. Gynecol. Oncol. 2020, 159, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Hamilton, A.M.; Furberg, H.; Pietzak, E.; Purdue, M.P.; Troester, M.A.; Hoadley, K.A.; Love, M.I. An approach for normalization and quality control for NanoString RNA expression data. Brief. Bioinform. 2021, 22, bbaa163. [Google Scholar] [CrossRef]
- Risso, D.; Ngai, J.; Speed, T.P.; Dudoit, S. Normalization of RNA-seq data using factor analysis of control genes or samples. Nat. Biotechnol. 2014, 32, 896–902. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Navarro, D.; Pedersen, T.L. ggplot2: Elegant Graphics for Data Analysis (3e). Available online: https://ggplot2-book.org/ (accessed on 1 June 2023).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | All Study Participants (n = 57) |
|---|---|
| Age (year) | 63.02 ± 15.67 |
| Weight (kg) | 73.14 ± 13.92 |
| No. of births | 1.86 ± 0.93 |
| No. of abortions | 0.44 ± 0.87 |
| Preoperative laboratory results | |
| 118.98 ± 17.51 |
| 0.37 ± 0.06 |
| 334.95 ± 125.73 |
| 103.47 ± 441.21 |
| 1118.26 ± 1567.83 |
| Symptoms | |
| 4.39 ± 7.21 |
| 31 (54.39%) |
| 14 (24.56%) |
| 6 (10.53%) |
| 15 (26.32%) |
| 6 (10.53%) |
| 7 (12.28%) |
| Surgery details | |
| 41 (71.93%) |
| 46 (80.70%) |
| 7 (12.28%) |
| 4 (7.02%) |
| 46 (80.70%) |
| 8.07 ± 4.50 |
| 0.68 ± 1.68 |
| ASA performance score (I:II:III:IV) [18] | 14:20:20:3 (24.56%:35.09%:35.09%:5.26%) |
| Histology | |
| 6 (10.53%) |
| 29 (50.88%) |
| 10 (17.54%) |
| 4 (7.02%) |
| 8 (14.04%) |
| Medical history | |
| 28 (49.12%) |
| 15 (26.32%) |
| 7 (12.28%) |
| 4 (7.02%) |
| 7 (12.28%) |
| Parameter | Univariate HR (95% CI) | Univariate p-Value | Multivariate HR (95% CI) | Multivariate p-Value |
|---|---|---|---|---|
| CD4+ T-lymphocytes (%) | 0.9459 (0.9121–0.9810) | 0.0028 | 0.9403 (0.8869–0.9970) | 0.0392 |
| CD8+ T-lymphocytes (%) | 1.0050 (0.9754–1.0350) | 0.7480 | 0.9967 (0.9548–1.0400) | 0.8801 |
| CD45+ leukocytes (%) | 0.9685 (0.9422–0.9954) | 0.0221 | 1.0073 (0.9544–1.0630) | 0.7913 |
| Immunostaining Sub-Cohorts | CD4+ | CD8+ | CD45+ |
|---|---|---|---|
| Negative (ref.) vs. weak | 0.8689 p = 0.7550 | 0.3206 p = 0.0389 | 0.3105 p = 0.0852 |
| Negative (ref.) vs. moderate | 0.2541 p = 0.0072 | 0.6884 p = 0.5021 | 0.3474 p = 0.1317 |
| Negative (ref.) vs. strong | 0.1543 p = 0.0013 | 0.4932 p = 0.2253 | 0.1132 p = 0.0010 |
| Weak (ref.) vs. moderate | 0.2924 p = 0.0074 | 2.1473 p = 0.0830 | 1.1188 p = 0.8254 |
| Weak (ref.) vs. strong | 0.1776 p = 0.0013 | 1.5385 p = 0.3550 | 0.3647 p = 0.0194 |
| Moderate (ref.) vs. strong | 0.6072 p = 0.3833 | 0.7165 p = 0.4850 | 0.3260 p = 0.0174 |
| Comparison | BATF3 | COL11A1 | IL6 | MMP1 | PDZK1IP1 | SOX11 | TNFRSF10C |
|---|---|---|---|---|---|---|---|
| Prognosis: Good (ref.) vs. Poor | – | + | + | – | |||
| ASA score: I (ref.) vs. II | + | + | |||||
| ASA score: I (ref.) vs. III | + | + | + | + | |||
| ASA score: II (ref.) vs. III | – | + | + | ||||
| T status: I (ref.) vs. II-III | – | + | + | ||||
| CD4+: 0% (ref.) vs. 1–5% | + | + | – | + | |||
| CD4+: 0% (ref.) vs. 5%< | + | – | + | – | + | ||
| CD8+: 6–15% (ref.) vs. 15%< | + | ||||||
| CD45+: 0–15% (ref.) vs. 15%< | + | + | + |
| Gene | Hazard Rate | 95% Confidence Interval | p-Value | Median Survival Time (Months) | |
|---|---|---|---|---|---|
| Lower Expression Group | Higher Expression Group | ||||
| A: Data obtained from the measurements of the current study | |||||
| COL11A1 | 1.1588 | 0.3092–4.3430 | 0.8268 | 8.26 | 4.01 |
| COL17A1 | 0.4503 | 0.1111–1.8257 | 0.2522 | 4.37 | 11.40 |
| COMP | 2.4723 | 0.6520–9.3754 | 0.1696 | 75.52 | 4.73 |
| CTAG1B | 0.7193 | 0.1923–2.6908 | 0.6229 | 4.99 | 5.13 |
| HLA-DQA1 | 0.5508 | 0.1365–2.2225 | 0.3954 | 4.93 | 11.40 |
| HLA-DQB1 | 0.5508 | 0.1365–2.2225 | 0.3954 | 4.93 | 5.26 |
| HLA-DRB5 | 0.3528 | 0.0873–1.4263 | 0.1272 | 4.22 | 5.26 |
| IL6 | 0.9211 | 0.2449–3.4637 | 0.9031 | 4.99 | 5.13 |
| ITGB3 | 2.0631 | 0.5501–7.7382 | 0.2728 | 75.58 | 3.32 |
| LYZ | 0.4036 | 0.1003–1.6245 | 0.1869 | 3.53 | 5.26 |
| MAGEC2 | 0.7766 | 0.2068–2.9159 | 0.7072 | 4.99 | 5.13 |
| MMP1 | 0.9489 | 0.2519–3.5747 | 0.9382 | 4.63 | 5.13 |
| PDZK1IP1 | 0.3304 | 0.0792–1.3782 | 0.1129 | 4.37 | 11.40 |
| PLA2G2A | 0.5690 | 0.1510–2.1444 | 0.3990 | 4.22 | 5.26 |
| SOX2 | 1.5048 | 0.4025–5.6254 | 0.5408 | 59.53 | 4.73 |
| SOX11 | 8.4100 | 1.6237–43.5603 | 0.0032 | 78.65 | 3.32 |
| TNFRSF18 | 0.2826 | 0.0665–1.2010 | 0.0716 | 3.66 | 11.40 |
| B: Data obtained from the Kaplan–Meier Plotter web application [19] | |||||
| COL11A1 | 1.30 | 1.14–1.48 | <0.0001 | 49.47 | 40.54 |
| COL17A1 | 0.91 | 0.80–1.03 | 0.1400 | 44.80 | 45.63 |
| COMP | 1.18 | 1.04–1.34 | 0.0110 | 48.00 | 41.00 |
| CTAG1B | 1.02 | 0.90–1.16 | 0.7200 | 45.63 | 44.77 |
| HLA-DQA1 | 0.96 | 0.84–1.09 | 0.4900 | 45.13 | 45.53 |
| HLA-DQB1 | 0.95 | 0.84–1.08 | 0.4500 | 45.23 | 45.17 |
| HLA-DRB5 | 0.89 | 0.78–1.01 | 0.0820 | 43.00 | 48.00 |
| IL6 | 1.07 | 0.94–1.22 | 0.3100 | 45.63 | 44.80 |
| ITGB3 | 0.93 | 0.82–1.06 | 0.2600 | 44.70 | 45.77 |
| LYZ | 0.91 | 0.80–1.04 | 0.1500 | 43.00 | 46.52 |
| MAGEC2 | 0.96 | 0.84–1.09 | 0.5000 | 44.53 | 45.77 |
| MMP1 | 1.08 | 0.95–1.23 | 0.2500 | 45.53 | 45.00 |
| PDZK1IP1 | 0.86 | 0.75–0.97 | 0.0180 | 41.89 | 48.00 |
| PLA2G2A | 1.04 | 0.91–1.18 | 0.5400 | 45.40 | 45.00 |
| SOX2 | 1.01 | 0.82–1.24 | 0.9300 | 45.00 | 41.89 |
| SOX11 | 1.25 | 1.10–1.42 | 0.0008 | 48.27 | 41.00 |
| TNFRSF18 | 0.93 | 0.76–1.14 | 0.4800 | 43.00 | 45.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barna, A.J.; Herold, Z.; Acs, M.; Bazsa, S.; Gajdacsi, J.; Garay, T.M.; Herold, M.; Madaras, L.; Muhl, D.; Nagy, A.; et al. High Tumor-Infiltrating Lymphocyte Count Is Associated with Distinct Gene Expression Profile and Longer Patient Survival in Advanced Ovarian Cancer. Int. J. Mol. Sci. 2023, 24, 13684. https://doi.org/10.3390/ijms241813684
Barna AJ, Herold Z, Acs M, Bazsa S, Gajdacsi J, Garay TM, Herold M, Madaras L, Muhl D, Nagy A, et al. High Tumor-Infiltrating Lymphocyte Count Is Associated with Distinct Gene Expression Profile and Longer Patient Survival in Advanced Ovarian Cancer. International Journal of Molecular Sciences. 2023; 24(18):13684. https://doi.org/10.3390/ijms241813684
Chicago/Turabian StyleBarna, Andras Jozsef, Zoltan Herold, Miklos Acs, Sandor Bazsa, Jozsef Gajdacsi, Tamas Marton Garay, Magdolna Herold, Lilla Madaras, Dorottya Muhl, Akos Nagy, and et al. 2023. "High Tumor-Infiltrating Lymphocyte Count Is Associated with Distinct Gene Expression Profile and Longer Patient Survival in Advanced Ovarian Cancer" International Journal of Molecular Sciences 24, no. 18: 13684. https://doi.org/10.3390/ijms241813684
APA StyleBarna, A. J., Herold, Z., Acs, M., Bazsa, S., Gajdacsi, J., Garay, T. M., Herold, M., Madaras, L., Muhl, D., Nagy, A., Szasz, A. M., & Dank, M. (2023). High Tumor-Infiltrating Lymphocyte Count Is Associated with Distinct Gene Expression Profile and Longer Patient Survival in Advanced Ovarian Cancer. International Journal of Molecular Sciences, 24(18), 13684. https://doi.org/10.3390/ijms241813684