
Molecular Relay Stations in Membrane Nanotubes: IRSp53 Involved in Actin-Based Force Generation


 ,
,  ,
,
Abstract
1. Introduction
2. Results
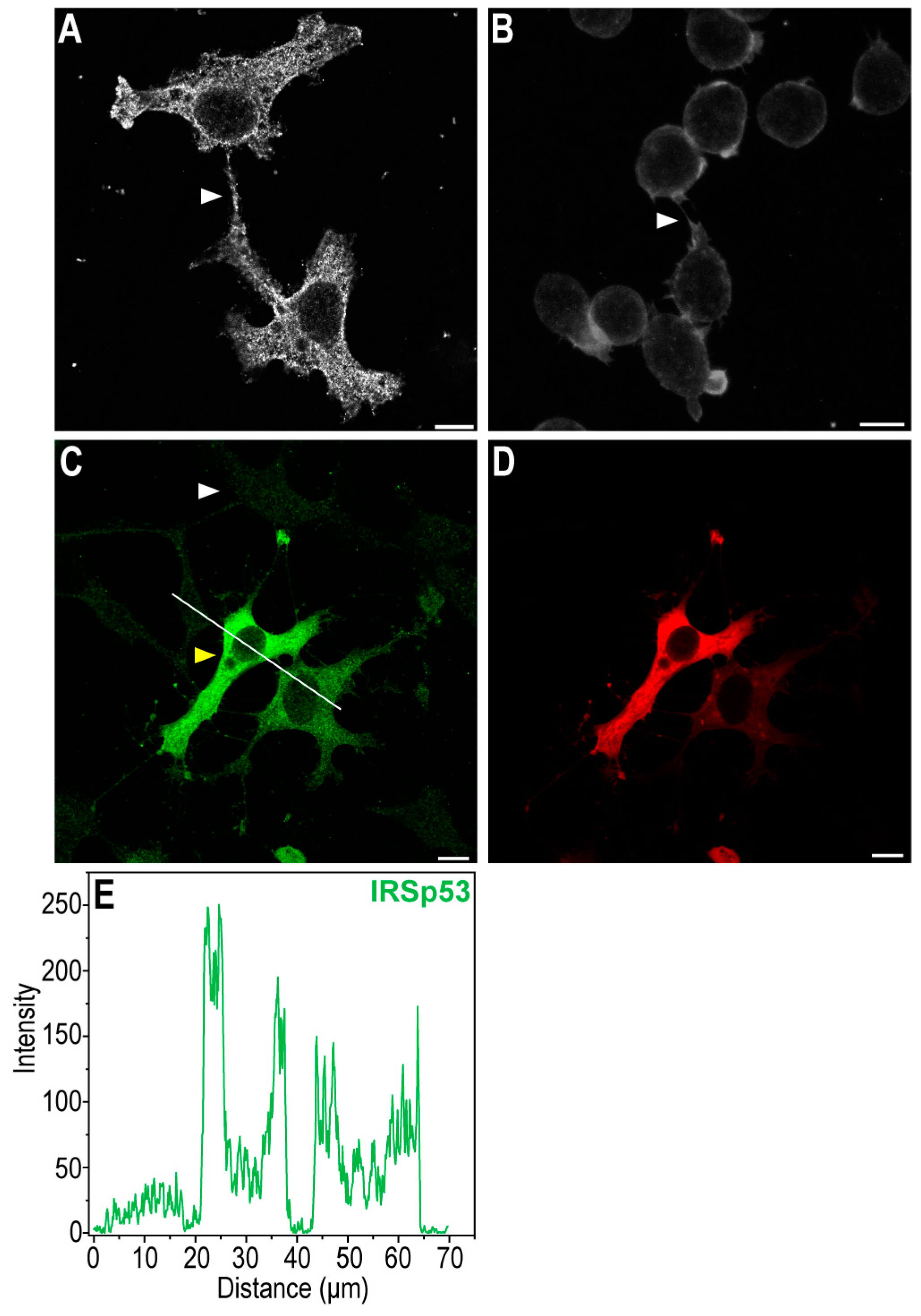
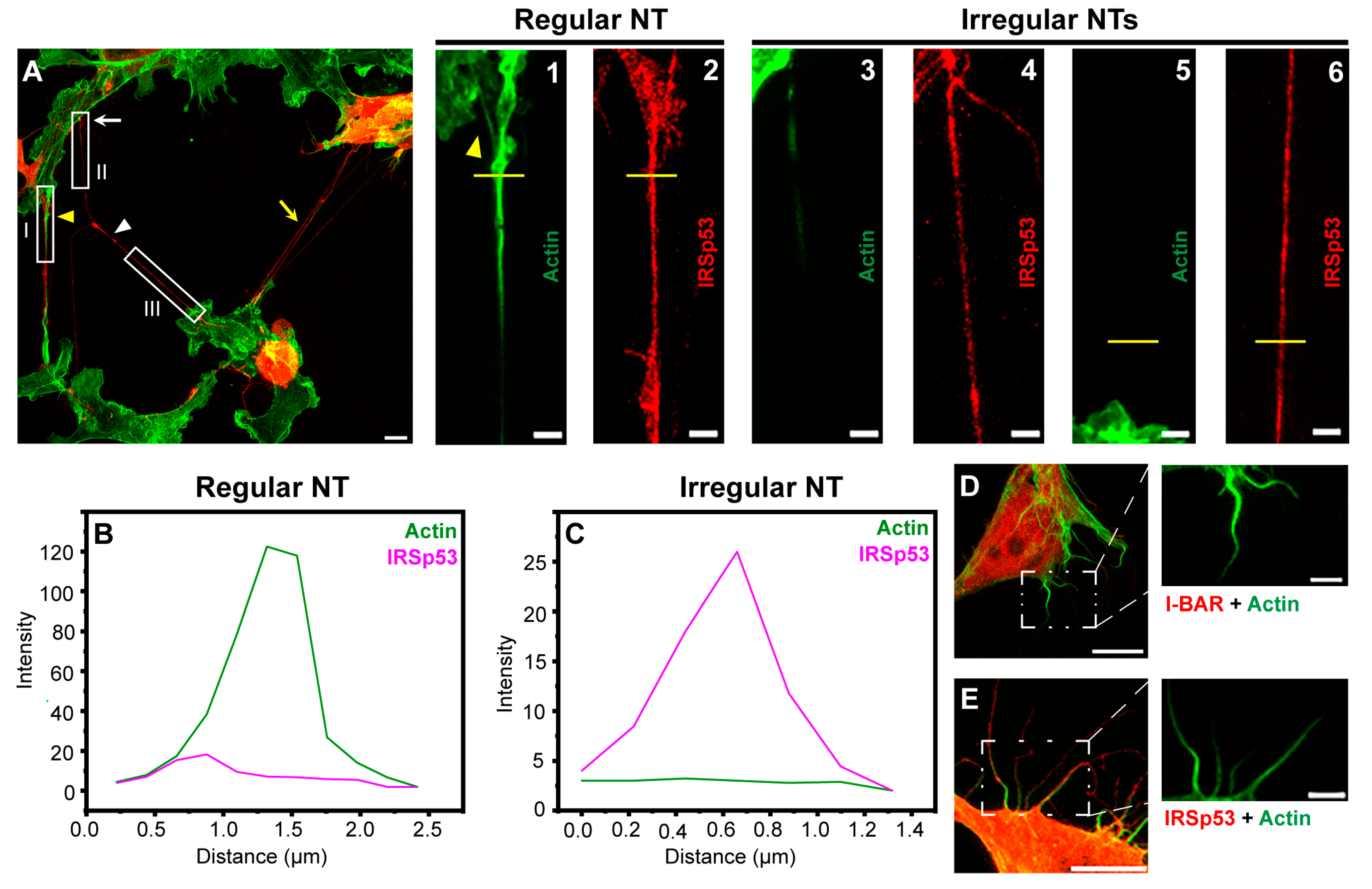
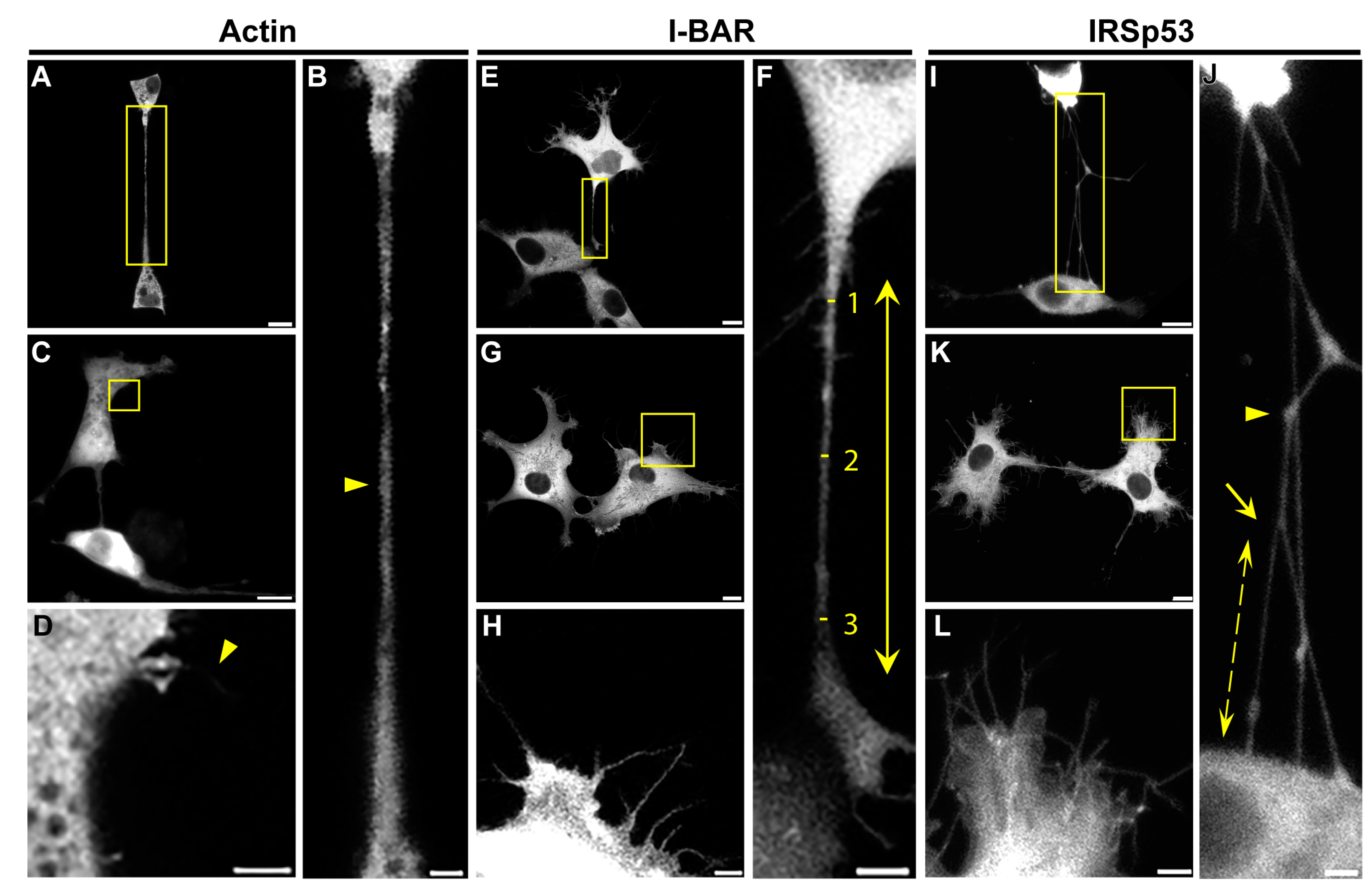
2.1. IRSp53 Is Expressed in the NTs of COS-7 Kidney and A20 B Lymphoma Cells
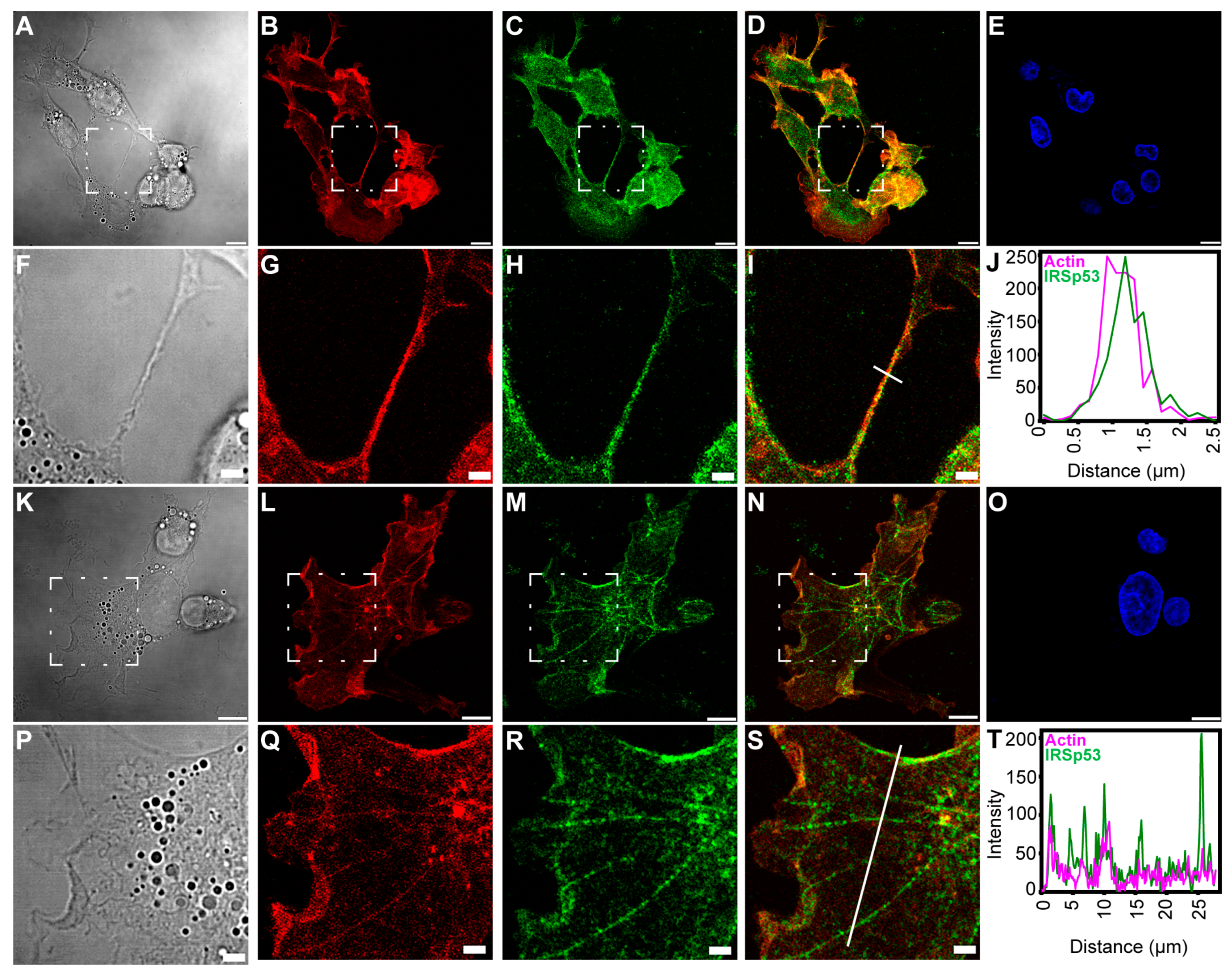
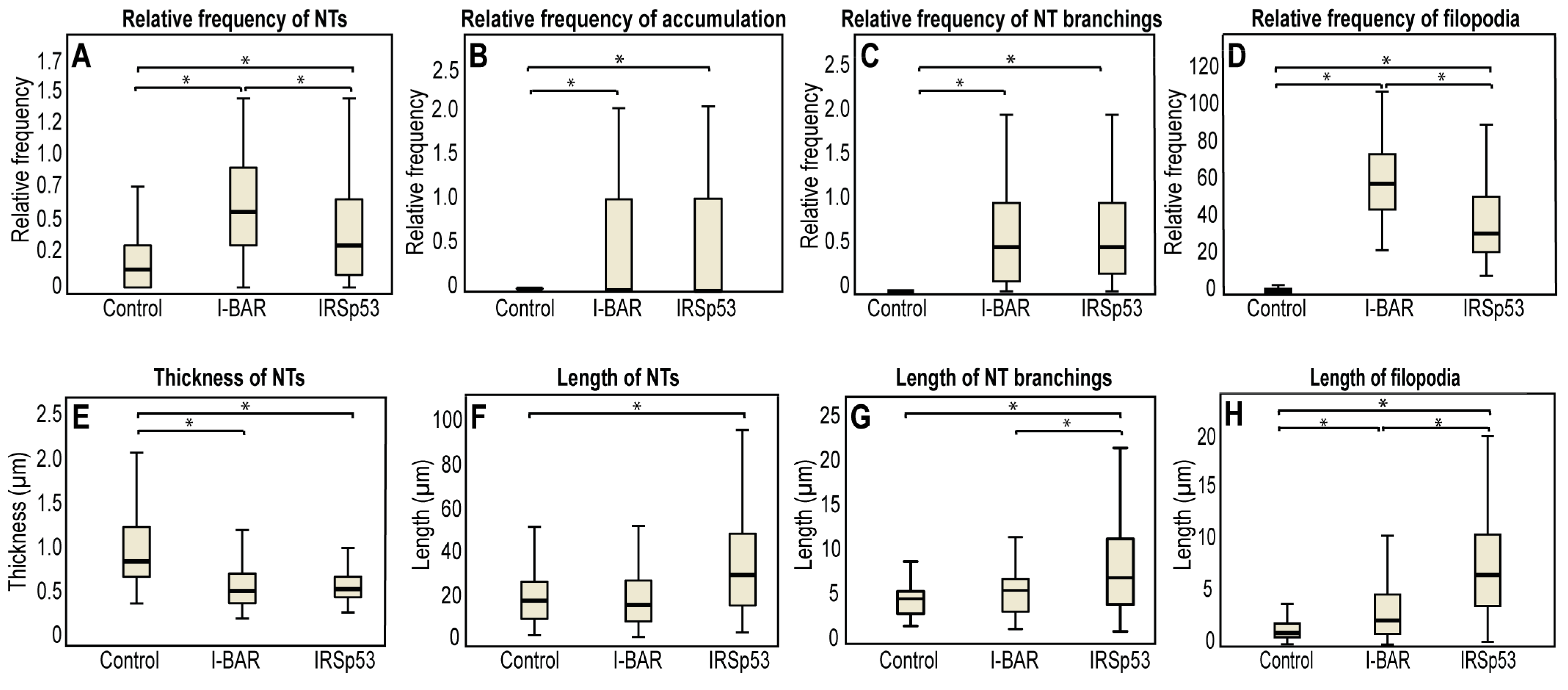
2.2. Overexpression of IRSp53 Protein and Its I-BAR Domain in COS-7 Kidney Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | COS-7 Control | I-BAR Overexpression | IRSp53 Overexpression |
|---|---|---|---|
| Number of filopodia per cell | 2 | 62 ± 4 | 38 ± 3 |
| Average length of filopodia (µm) | 1.80 ± 0.13 | 3.90 ± 0.19 | 8.15 ± 0.33 |
| Relative frequency of NTs | 0.29 ± 0.02 | 0.75 ± 0.03 | 0.54 ± 0.03 |
| Average diameter of the NTs (µm) | 1.06 ± 0.03 | 0.56 ± 0.01 | 0.67 ± 0.02 |
| Length of the NTs (µm) | 21.50 ± 1.13 | 20.25 ± 0.77 | 36.13 ± 1.55 |
| Relative branching | 0.33 ± 0.11 | 0.77 ± 0.12 | 0.78 ± 0.06 |
| Length of the branches (µm) | 4.80 ± 0.42 | 5.82 ± 0.34 | 8.77 ± 0.50 |
| Accumulation level in the NTs | 0.22 ± 0.05 | 0.50 ± 0.07 | 0.50 ± 0.08 |
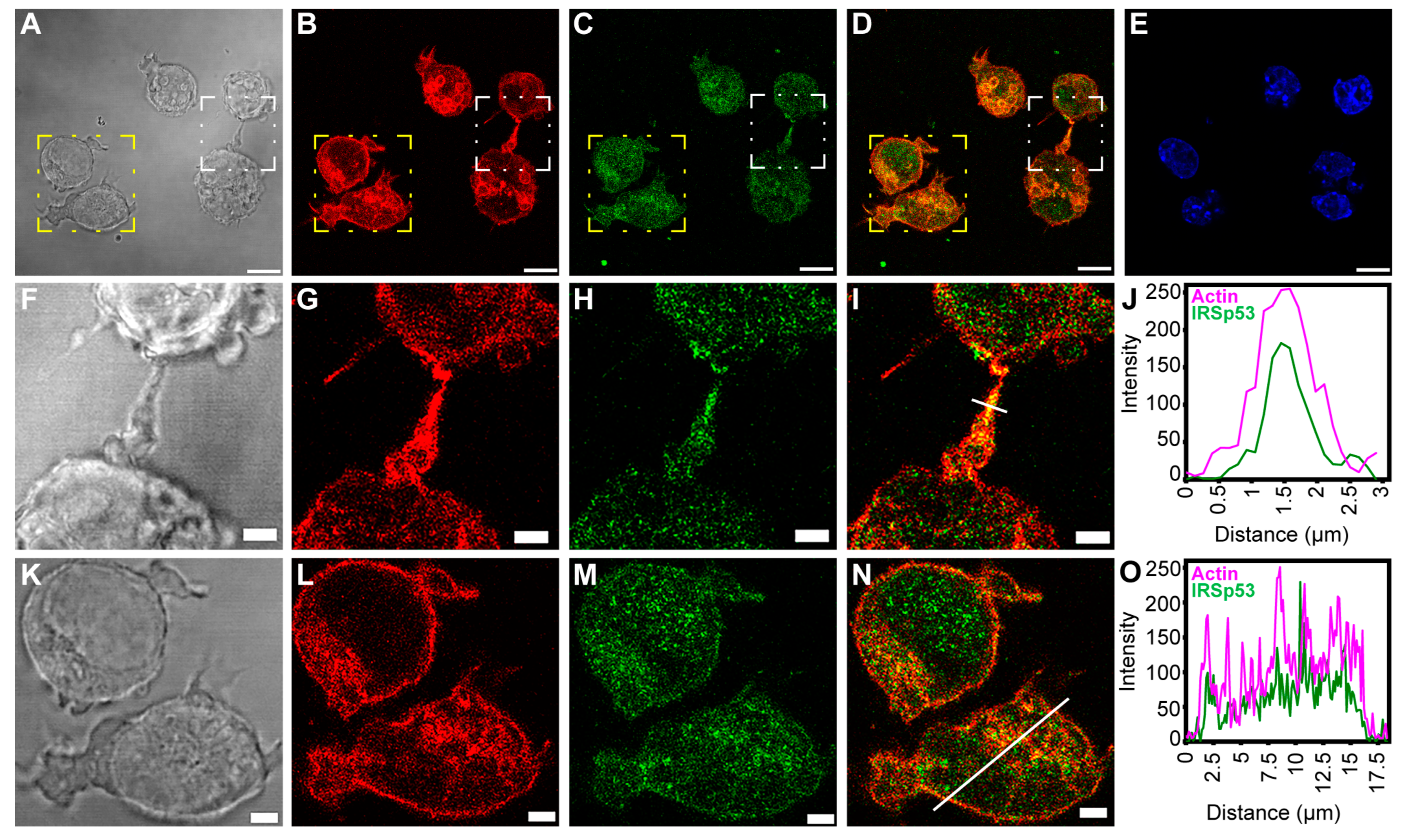
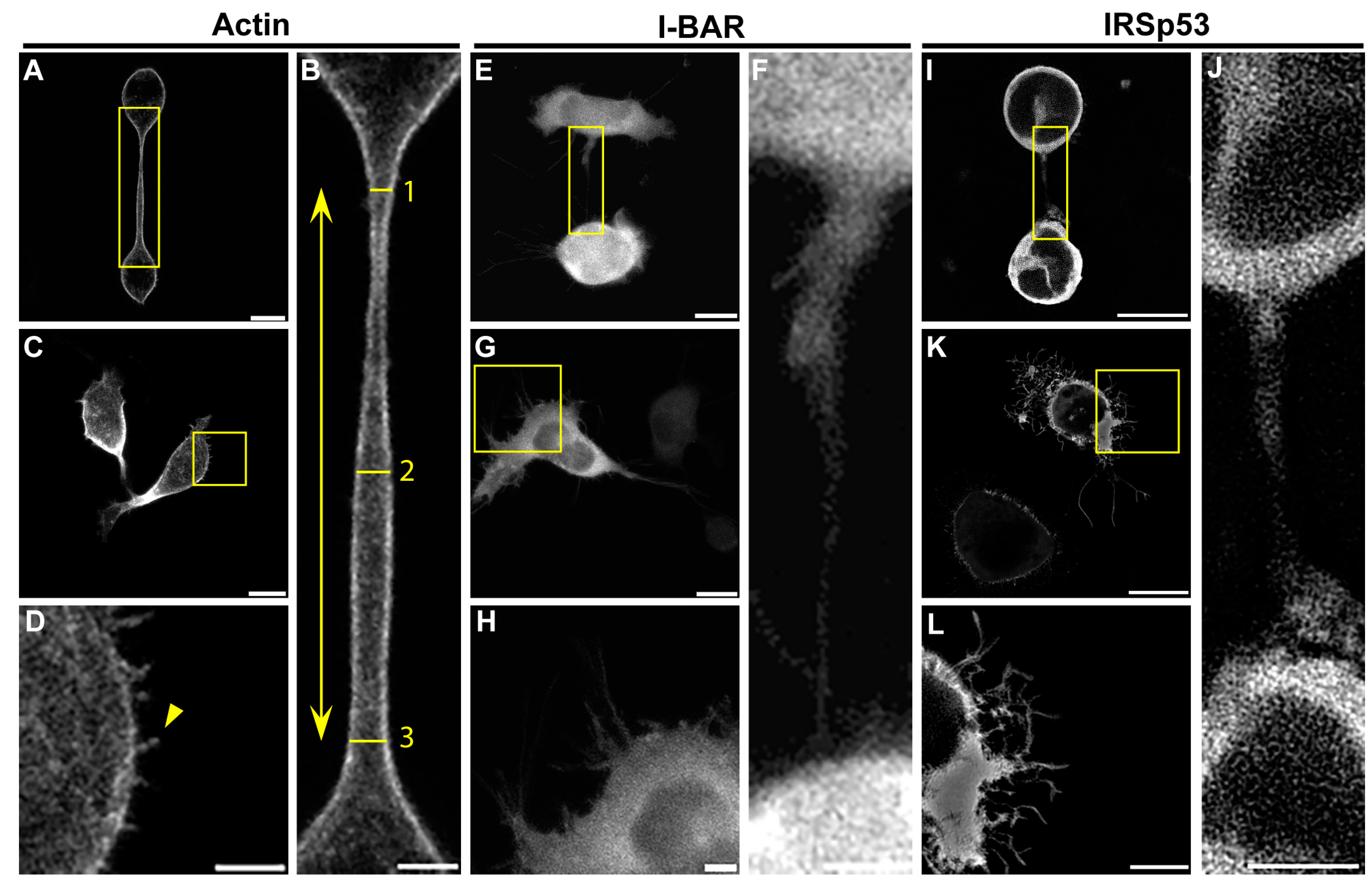
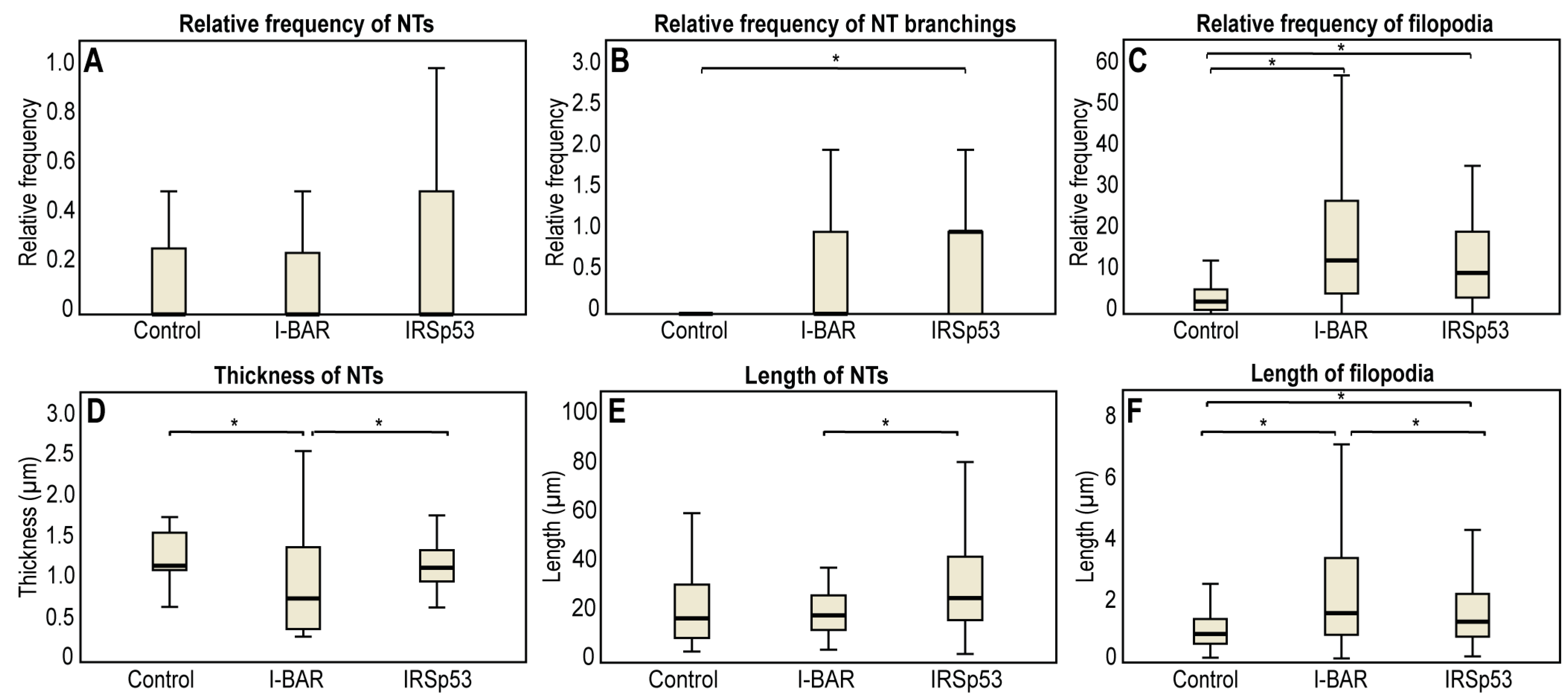
2.3. Overexpression of IRSp53 Protein and Its I-BAR Domain in A20 B Lymphoma Cells
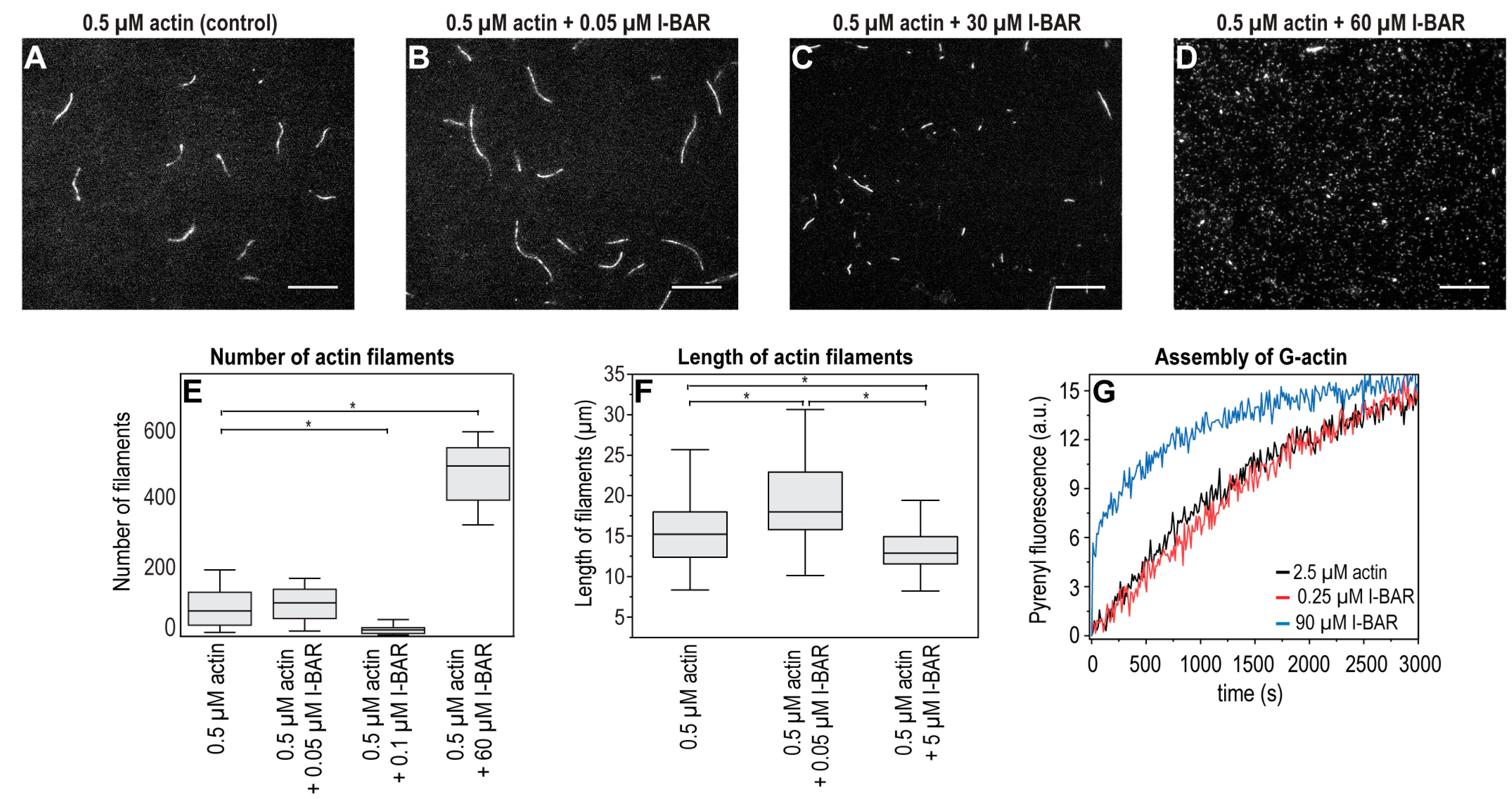
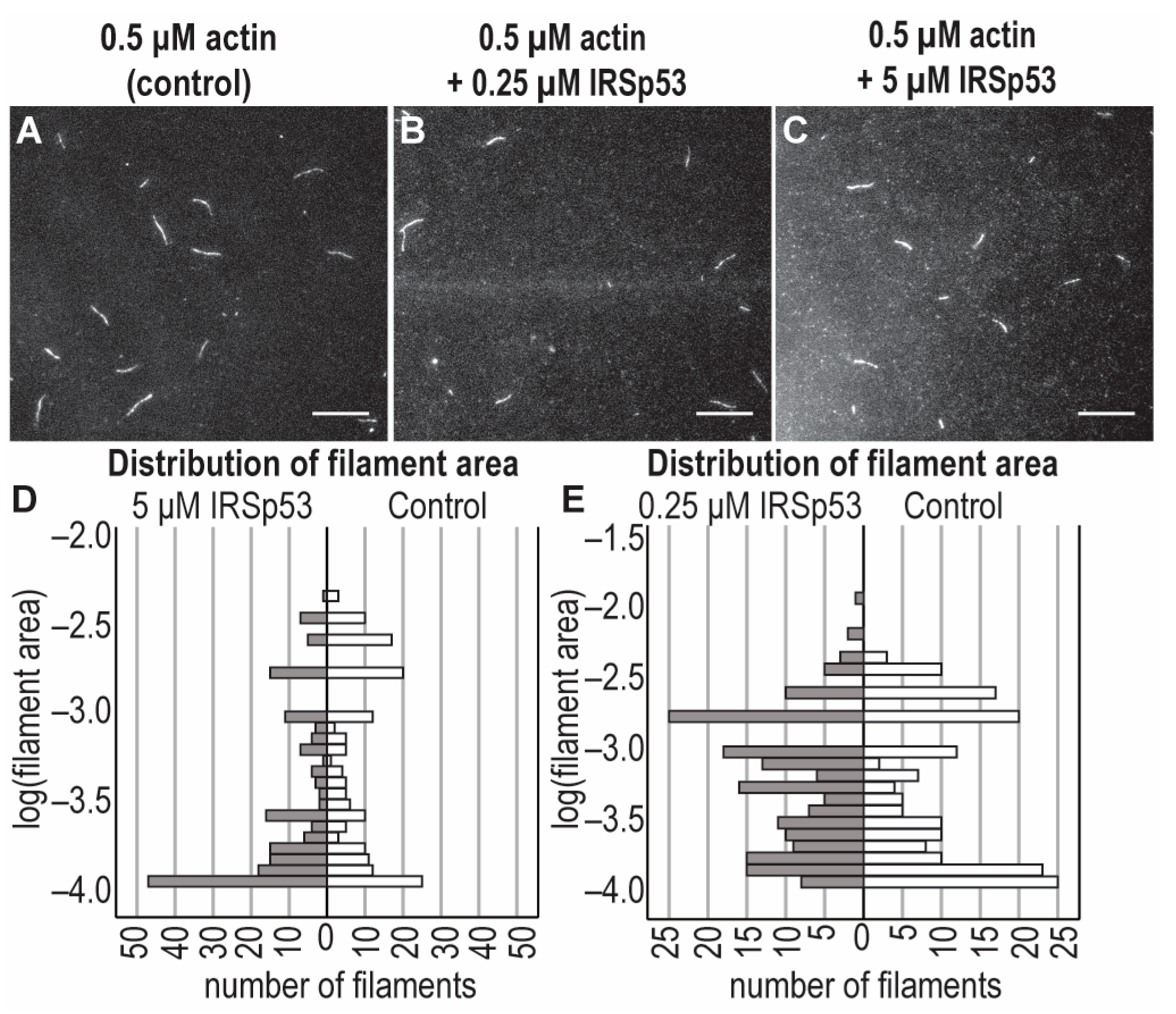
2.4. The Effects of IRSp53 and Its I-BAR Domain on Actin Polymerisation
3. Discussion
3.1. Background and Aim
3.2. Key Observations
3.3. General Concept
3.4. The Role of the IRSp53
3.5. Cell Line Differences
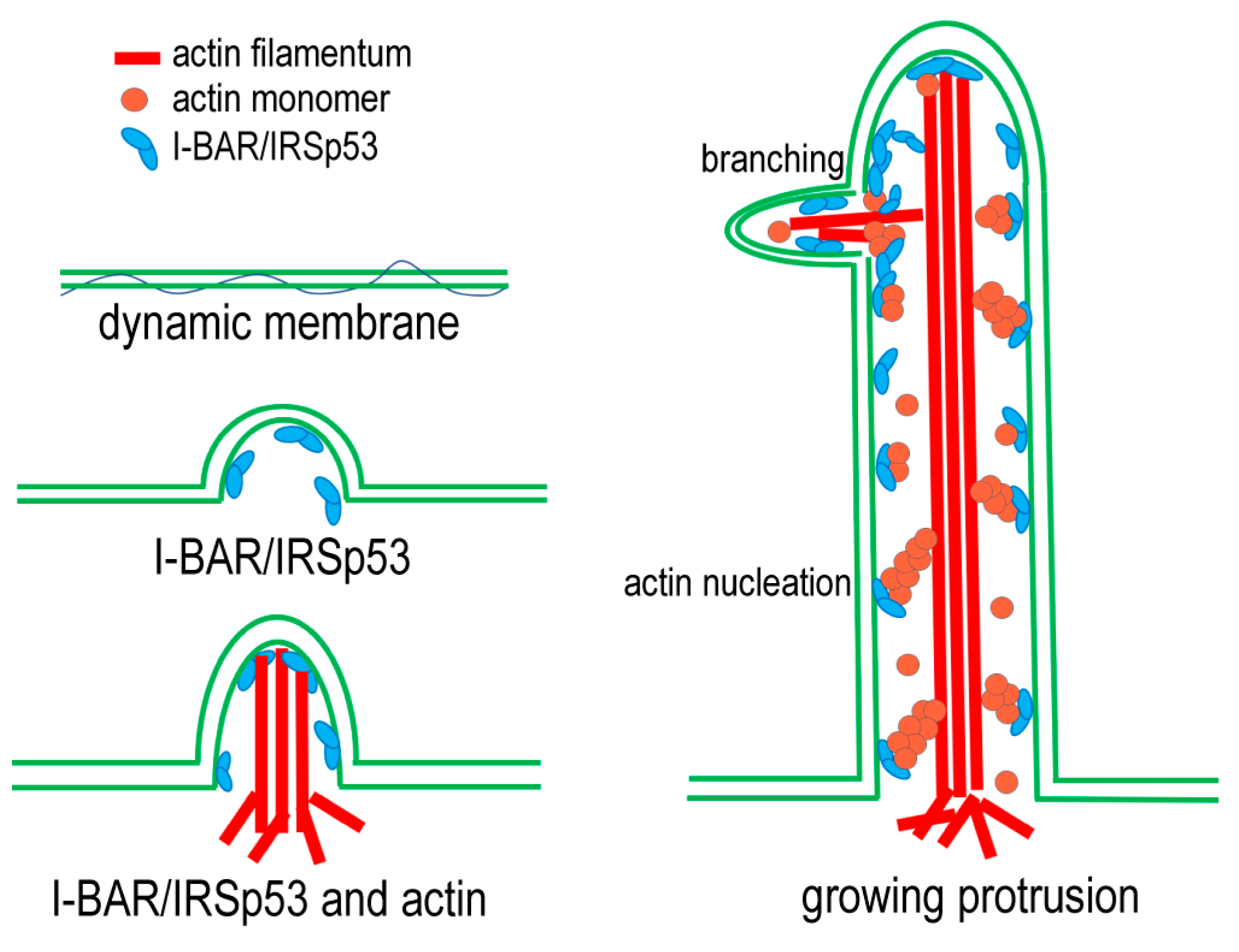
3.6. Model
4. Materials and Methods
4.1. Protein Purification and Labelling
4.2. Cells, Culture, Cell Transfection
4.3. Immunocytochemistry and Cell Staining
4.4. Microscopic Imaging
4.5. Statistical Analysis
4.6. Total Internal Reflection Fluorescence Microscopy
4.7. Pyrenyl Polymerisation Assay
4.8. General Experimental Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baumgart, T.; Capraro, B.R.; Zhu, C.; Das, S.L. Thermodynamics and Mechanics of Membrane Curvature Generation and Sensing by Proteins and Lipids. Annu. Rev. Phys. Chem. 2011, 62, 483–506. [Google Scholar] [CrossRef]
- Disanza, A.; Mantoani, S.; Hertzog, M.; Gerboth, S.; Frittoli, E.; Steffen, A.; Berhoerster, K.; Kreienkamp, H.-J.; Milanesi, F.; Di Fiore, P.P.; et al. Regulation of Cell Shape by Cdc42 Is Mediated by the Synergic Actin-Bundling Activity of the Eps8–IRSp53 Complex. Nat. Cell Biol. 2006, 8, 1337–1347. [Google Scholar] [CrossRef]
- Fošnarič, M.; Penič, S.; Iglič, A.; Kralj-Iglič, V.; Drab, M.; Gov, N.S. Theoretical Study of Vesicle Shapes Driven by Coupling Curved Proteins and Active Cytoskeletal Forces. Soft Matter 2019, 15, 5319–5330. [Google Scholar] [CrossRef]
- Millard, T.H.; Bompard, G.; Heung, M.Y.; Dafforn, T.R.; Scott, D.J.; Machesky, L.M.; Fütterer, K. Structural Basis of Filopodia Formation Induced by the IRSp53/MIM Homology Domain of Human IRSp53. EMBO J. 2005, 24, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, S.; Murayama, K.; Sakamoto, A.; Hanawa-Suetsugu, K.; Seto, A.; Oikawa, T.; Mishima, C.; Shirouzu, M.; Takenawa, T.; Yokoyama, S. The RAC Binding Domain/IRSp53-MIM Homology Domain of IRSp53 Induces RAC-Dependent Membrane Deformation. J. Biol. Chem. 2006, 281, 35347–35358. [Google Scholar] [CrossRef]
- Lee, S.H.; Kerff, F.; Chereau, D.; Ferron, F.; Klug, A.; Dominguez, R. Structural Basis for the Actin-Binding Function of Missing-in-Metastasis. Structure 2007, 15, 145–155. [Google Scholar] [CrossRef]
- Mattila, P.K.; Pykäläinen, A.; Saarikangas, J.; Paavilainen, V.O.; Vihinen, H.; Jokitalo, E.; Lappalainen, P. Missing-in-Metastasis and IRSp53 Deform PI(4,5)P2-Rich Membranes by an Inverse BAR Domain–like Mechanism. J. Cell Biol. 2007, 176, 953–964. [Google Scholar] [CrossRef]
- Saarikangas, J.; Zhao, H.; Pykäläinen, A.; Laurinmäki, P.; Mattila, P.K.; Kinnunen, P.K.J.; Butcher, S.J.; Lappalainen, P. Molecular Mechanisms of Membrane Deformation by I-BAR Domain Proteins. Curr. Biol. 2009, 19, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, R. The WH2 Domain and Actin Nucleation: Necessary but Insufficient. Trends Biochem. Sci. 2016, 41, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Krugmann, S.; Jordens, I.; Gevaert, K.; Driessens, M.; Vandekerckhove, J.; Hall, A. Cdc42 Induces Filopodia by Promoting the Formation of an IRSp53: Mena Complex. Curr. Biol. 2001, 11, 1645–1655. [Google Scholar] [CrossRef]
- Kaksonen, M.; Toret, C.P.; Drubin, D.G. Harnessing Actin Dynamics for Clathrin-Mediated Endocytosis. Nat. Rev. Mol. Cell Biol. 2006, 7, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Funato, Y.; Terabayashi, T.; Suenaga, N.; Seiki, M.; Takenawa, T.; Miki, H. IRSp53/Eps8 Complex Is Important for Positive Regulation of Rac and Cancer Cell Motility/Invasiveness. Cancer Res. 2004, 64, 5237–5244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Pykäläinen, A.; Lappalainen, P. I-BAR Domain Proteins: Linking Actin and Plasma Membrane Dynamics. Curr. Opin. Cell Biol. 2011, 23, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.-H. Nanotubular Highways for Intercellular Organelle Transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Delage, E.; Cervantes, D.C.; Pénard, E.; Schmitt, C.; Syan, S.; Disanza, A.; Scita, G.; Zurzolo, C. Differential Identity of Filopodia and Tunneling Nanotubes Revealed by the Opposite Functions of Actin Regulatory Complexes. Sci. Rep. 2016, 6, 39632. [Google Scholar] [CrossRef] [PubMed]
- Halász, H.; Ghadaksaz, A.R.; Madarász, T.; Huber, K.; Harami, G.; Tóth, E.A.; Osteikoetxea-Molnár, A.; Kovács, M.; Balogi, Z.; Nyitrai, M.; et al. Live Cell Superresolution-Structured Illumination Microscopy Imaging Analysis of the Intercellular Transport of Microvesicles and Costimulatory Proteins via Nanotubes between Immune Cells. Methods Appl. Fluoresc. 2018, 6, 045005. [Google Scholar] [CrossRef]
- Osteikoetxea-Molnár, A.; Szabó-Meleg, E.; Tóth, E.A.; Oszvald, Á.; Izsépi, E.; Kremlitzka, M.; Biri, B.; Nyitray, L.; Bozó, T.; Németh, P.; et al. The Growth Determinants and Transport Properties of Tunneling Nanotube Networks between B Lymphocytes. Cell. Mol. Life Sci. 2016, 73, 4531–4545. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, J.; Sun, X.; Zhang, Y. Tunneling-Nanotube Development in Astrocytes Depends on P53 Activation. Cell Death Differ. 2011, 18, 732–742. [Google Scholar] [CrossRef]
- Abounit, S.; Wu, J.W.; Duff, K.; Victoria, G.S.; Zurzolo, C. Tunneling Nanotubes: A Possible Highway in the Spreading of Tau and Other Prion-like Proteins in Neurodegenerative Diseases. Prion 2016, 10, 344–351. [Google Scholar] [CrossRef]
- Veranič, P.; Lokar, M.; Schütz, G.J.; Weghuber, J.; Wieser, S.; Hägerstrand, H.; Kralj-Iglič, V.; Iglič, A. Different Types of Cell-to-Cell Connections Mediated by Nanotubular Structures. Biophys. J. 2008, 95, 4416–4425. [Google Scholar] [CrossRef]
- Pepe, A.; Pietropaoli, S.; Vos, M.; Barba-Spaeth, G.; Zurzolo, C. Tunneling Nanotubes Provide a Route for SARS-CoV-2 Spreading. Sci. Adv. 2022, 8, eabo0171. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cao, J. Astrocyte-to-Neuron Transportation of Enhanced Green Fluorescent Protein in Cerebral Cortex Requires F-Actin Dependent Tunneling Nanotubes. Sci. Rep. 2021, 11, 16798. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, H.-H.; Bukoreshtliev, N.V.; Barroso, J.F.V. Tunneling Nanotubes: A New Route for the Exchange of Components between Animal Cells. FEBS Lett. 2007, 581, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, H.-H.; Rustom, A.; Wang, X. Tunneling Nanotubes, an Emerging Intercellular Communication Route in Development. Mech. Dev. 2013, 130, 381–387. [Google Scholar] [CrossRef]
- Gerdes, H.-H.; Carvalho, R.N. Intercellular Transfer Mediated by Tunneling Nanotubes. Curr. Opin. Cell Biol. 2008, 20, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Sisakhtnezhad, S.; Khosravi, L. Emerging Physiological and Pathological Implications of Tunneling Nanotubes Formation between Cells. Eur. J. Cell Biol. 2015, 94, 429–443. [Google Scholar] [CrossRef]
- Rustom, A. The Missing Link: Does Tunnelling Nanotube-Based Supercellularity Provide a New Understanding of Chronic and Lifestyle Diseases? Open Biol. 2016, 6, 160057. [Google Scholar] [CrossRef]
- Austefjord, M.W.; Gerdes, H.-H.; Wang, X. Tunneling Nanotubes. Commun. Integr. Biol. 2014, 7, e27934. [Google Scholar] [CrossRef]
- Tóth, E.A.; Oszvald, Á.; Péter, M.; Balogh, G.; Osteikoetxea-Molnár, A.; Bozó, T.; Szabó-Meleg, E.; Nyitrai, M.; Derényi, I.; Kellermayer, M.; et al. Nanotubes Connecting B Lymphocytes: High Impact of Differentiation-Dependent Lipid Composition on Their Growth and Mechanics. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2017, 1862, 991–1000. [Google Scholar] [CrossRef]
- Mahapatra, A.; Uysalel, C.; Rangamani, P. The Mechanics and Thermodynamics of Tubule Formation in Biological Membranes. J. Membr. Biol. 2021, 254, 273–291. [Google Scholar] [CrossRef]
- Stachowiak, J.C.; Hayden, C.C.; Sasaki, D.Y. Steric Confinement of Proteins on Lipid Membranes Can Drive Curvature and Tubulation. Proc. Natl. Acad. Sci. USA 2010, 107, 7781–7786. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, A.; Masuda, M.; Ohki, T.; Onishi, H.; Mochizuki, N. A Novel Actin Bundling/Filopodium-Forming Domain Conserved in Insulin Receptor Tyrosine Kinase Substrate P53 and Missing in Metastasis Protein. J. Biol. Chem. 2004, 279, 14929–14936. [Google Scholar] [CrossRef] [PubMed]
- Cordero Cervantes, D.; Zurzolo, C. Peering into Tunneling Nanotubes—The Path Forward. EMBO J. 2021, 40, e105789. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hoelzle, M.; Disanza, A.; Scita, G.; Svitkina, T. Coordination of Membrane and Actin Cytoskeleton Dynamics during Filopodia Protrusion. PLoS ONE 2009, 4, e5678. [Google Scholar] [CrossRef] [PubMed]
- Teodorof, C.; Bae, J.I.L.; Kim, S.-M.; Oh, H.J.; Kang, Y.S.; Choi, J.; Chun, J.-S.; Song, W.K. SPIN90-IRSp53 Complex Participates in Rac-Induced Membrane Ruffling. Exp. Cell Res. 2009, 315, 2410–2419. [Google Scholar] [CrossRef] [PubMed]
- Expression Atlas. Available online: https://www.ebi.ac.uk/gxa/search?geneQuery=%5B%7B%22value%22%3A%22IRSp53%22%7D%5D&species=Macaca%20mulatta&conditionQuery=%5B%5D&bs=%7B%22macaca%20mulatta%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline (accessed on 27 July 2023).
- Expression Atlas. Available online: https://www.ebi.ac.uk/gxa/experiments/E-MTAB-3079/Results?geneQuery=%5B%7B%22value%22%3A%22ensmusg00000025372%22%7D%5D (accessed on 27 July 2023).
- Sudhaharan, T.; Hariharan, S.; Lim, J.S.Y.; Liu, J.Z.; Koon, Y.L.; Wright, G.D.; Chiam, K.H.; Ahmed, S. Superresolution Microscopy Reveals Distinct Localisation of Full Length IRSp53 and Its I-BAR Domain Protein within Filopodia. Sci. Rep. 2019, 9, 2524. [Google Scholar] [CrossRef]
- Mandal, K. Review of PIP2 in Cellular Signaling, Functions and Diseases. Int. J. Mol. Sci. 2020, 21, 8342. [Google Scholar] [CrossRef]
- Bompard, G.; Sharp, S.J.; Freiss, G.; Machesky, L.M. Involvement of Rac in Actin Cytoskeleton Rearrangements Induced by MIM-B. J. Cell Sci. 2005, 118, 5393–5403. [Google Scholar] [CrossRef][Green Version]
- Disanza, A.; Bisi, S.; Winterhoff, M.; Milanesi, F.; Ushakov, D.S.; Kast, D.; Marighetti, P.; Romet-Lemonne, G.; Müller, H.-M.; Nickel, W.; et al. CDC42 Switches IRSp53 from Inhibition of Actin Growth to Elongation by Clustering of VASP. EMBO J. 2013, 32, 2735–2750. [Google Scholar] [CrossRef]
- Yin, H.L.; Janmey, P.A. Phosphoinositide Regulation of the Actin Cytoskeleton. Annu. Rev. Physiol. 2003, 65, 761–789. [Google Scholar] [CrossRef]
- Defacque, H.; Egeberg, M.; Habermann, A.; Diakonova, M.; Roy, C.; Mangeat, P.; Voelter, W.; Marriott, G.; Pfannstiel, J.; Faulstich, H.; et al. Involvement of Ezrin/Moesin in de Novo Actin Assembly on Phagosomal Membranes. EMBO J. 2000, 19, 199–212. [Google Scholar] [CrossRef]
- Feng, Z.; Lee, S.; Jia, B.; Jian, T.; Kim, E.; Zhang, M. IRSp53 Promotes Postsynaptic Density Formation and Actin Filament Bundling. J. Cell Biol. 2022, 221, e202105035. [Google Scholar] [CrossRef]
- Tsai, F.-C.; Henderson, J.M.; Jarin, Z.; Kremneva, E.; Senju, Y.; Pernier, J.; Mikhajlov, O.; Manzi, J.; Kogan, K.; Le Clainche, C.; et al. Activated I-BAR IRSp53 Clustering Controls the Formation of VASP-Actin–Based Membrane Protrusions. Sci. Adv. 2022, 8, eabp8677. [Google Scholar] [CrossRef] [PubMed]
- Chou, A.M.; Sem, K.P.; Wright, G.D.; Sudhaharan, T.; Ahmed, S. Dynamin1 Is a Novel Target for IRSp53 Protein and Works with Mammalian Enabled (Mena) Protein and Eps8 to Regulate Filopodial Dynamics. J. Biol. Chem. 2014, 289, 24383–24396. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.I.; Lim, K.B.; Sudhaharan, T.; Sem, K.P.; Bu, W.; Chou, A.M.; Ahmed, S. MDia1 and WAVE2 Proteins Interact Directly with IRSp53 in Filopodia and Are Involved in Filopodium Formation. J. Biol. Chem. 2012, 287, 4702–4714. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.B.; Bu, W.; Goh, W.I.; Koh, E.; Ong, S.H.; Pawson, T.; Sudhaharan, T.; Ahmed, S. The Cdc42 Effector IRSp53 Generates Filopodia by Coupling Membrane Protrusion with Actin Dynamics. J. Biol. Chem. 2008, 283, 20454–20472. [Google Scholar] [CrossRef]
- Önfelt, B.; Nedvetzki, S.; Yanagi, K.; Davis, D.M. Cutting Edge: Membrane Nanotubes Connect Immune Cells. J. Immunol. 2004, 173, 1511–1513. [Google Scholar] [CrossRef]
- Mesarec, L.; Drab, M.; Penič, S.; Kralj-Iglič, V.; Iglič, A. On the Role of Curved Membrane Nanodomains and Passive and Active Skeleton Forces in the Determination of Cell Shape and Membrane Budding. Int. J. Mol. Sci. 2021, 22, 2348. [Google Scholar] [CrossRef]
- Vaggi, F.; Disanza, A.; Milanesi, F.; di Fiore, P.P.; Menna, E.; Matteoli, M.; Gov, N.S.; Scita, G.; Ciliberto, A. The Eps8/IRSp53/VASP Network Differentially Controls Actin Capping and Bundling in Filopodia Formation. PLoS Comput. Biol. 2011, 7, e1002088. [Google Scholar] [CrossRef]
- Kast, D.J.; Yang, C.; Disanza, A.; Boczkowska, M.; Madasu, Y.; Scita, G.; Svitkina, T.; Dominguez, R. Mechanism of IRSp53 Inhibition and Combinatorial Activation by Cdc42 and Downstream Effectors. Nat. Struct. Mol. Biol. 2014, 21, 413–422. [Google Scholar] [CrossRef]
- Millard, T.H.; Dawson, J.; Machesky, L.M. Characterisation of IRTKS, a Novel IRSp53/MIM Family Actin Regulator with Distinct Filament Bundling Properties. J. Cell Sci. 2007, 120, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Jarin, Z.; Pak, A.J.; Bassereau, P.; Voth, G.A. Lipid-Composition-Mediated Forces Can Stabilize Tubular Assemblies of I-BAR Proteins. Biophys. J. 2021, 120, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef] [PubMed]
- Spudich, J.A.; Watt, S. The Regulation of Rabbit Skeletal Muscle Contraction. I. Biochemical Studies of the Interaction of the Tropomyosin-Troponin Complex with Actin and the Proteolytic Fragments of Myosin. J. Biol. Chem. 1971, 246, 4866–4871. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Pantaloni, D.; Korn, E.D. Fluorescence Measurements of the Binding of Cations to High-Affinity and Low-Affinity Sites on ATP-G-Actin. J. Biol. Chem. 1986, 261, 10778–10784. [Google Scholar] [CrossRef] [PubMed]
- Kouyama, T.; Mihashi, K. Fluorimetry Study of N-(1-Pyrenyl)Iodoacetamide-Labelled F-Actin. Local Structural Change of Actin Protomer Both on Polymerization and on Binding of Heavy Meromyosin. Eur. J. Biochem. 1981, 114, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Waller, A.P. Investigations into Membrane Protrusions Mediated by IRSp53 IMD. Ph.D. Thesis, University of Birmingham, Birmingham, UK, 2010. [Google Scholar]
- Manders, E.M.M.; Verbeek, F.J.; Aten, J.A. Measurement of Co-Localization of Objects in Dual-Colour Confocal Images. J. Microsc. 1993, 169, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Hencz, A.J.; Somogyi, P.; Halász, H.; Szabó-Meleg, E. Visualization of the Effect of TR100 Anti-Cancer Compound on Membrane Nanotubes with SR-SIM Microscopy. Resolut. Discov. 2022, 6, 12–19. [Google Scholar] [CrossRef]
- Barkó, S.; Bugyi, B.; Carlier, M.-F.; Gombos, R.; Matusek, T.; Mihály, J.; Nyitrai, M. Characterization of the Biochemical Properties and Biological Function of the Formin Homology Domains of Drosophila DAAM. J. Biol. Chem. 2010, 285, 13154–13169. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Matkó, J.; Tóth, E.A. Membrane Nanotubes Are Ancient Machinery for Cell-to-Cell Communication and Transport. Their Interference with the Immune System. Biol. Futur. 2021, 72, 25–36. [Google Scholar] [CrossRef] [PubMed]











| Parameter | A20 Control | I-BAR Overexpression | IRSp53 Overexpression |
|---|---|---|---|
| Number of filopodia per cell | 4.33 ± 0.51 | 16.30 ± 2.12 | 13.86 ± 1.65 |
| Average length of filopodia (µm) | 1.24 ± 0.06 | 3.05 ± 0.16 | 2.18 ± 0.11 |
| Relative frequency of NTs | 0.16 ± 0.03 | 0.12 ± 0.03 | 0.19 ± 0.04 |
| Average diameter of the NTs (µm) | 1.35 ± 0.10 | 0.93 ± 0.12 | 1.24 ±0.08 |
| Length of the NTs (µm) | 26.33 ± 5.59 | 21.36 ± 2.29 | 31.75 ± 4.17 |
| Relative branching | 0.12 ± 0.80 | 0.55 ± 0.19 | 1.00 ± 0.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madarász, T.; Brunner, B.; Halász, H.; Telek, E.; Matkó, J.; Nyitrai, M.; Szabó-Meleg, E. Molecular Relay Stations in Membrane Nanotubes: IRSp53 Involved in Actin-Based Force Generation. Int. J. Mol. Sci. 2023, 24, 13112. https://doi.org/10.3390/ijms241713112
Madarász T, Brunner B, Halász H, Telek E, Matkó J, Nyitrai M, Szabó-Meleg E. Molecular Relay Stations in Membrane Nanotubes: IRSp53 Involved in Actin-Based Force Generation. International Journal of Molecular Sciences. 2023; 24(17):13112. https://doi.org/10.3390/ijms241713112
Chicago/Turabian StyleMadarász, Tamás, Brigitta Brunner, Henriett Halász, Elek Telek, János Matkó, Miklós Nyitrai, and Edina Szabó-Meleg. 2023. "Molecular Relay Stations in Membrane Nanotubes: IRSp53 Involved in Actin-Based Force Generation" International Journal of Molecular Sciences 24, no. 17: 13112. https://doi.org/10.3390/ijms241713112
APA StyleMadarász, T., Brunner, B., Halász, H., Telek, E., Matkó, J., Nyitrai, M., & Szabó-Meleg, E. (2023). Molecular Relay Stations in Membrane Nanotubes: IRSp53 Involved in Actin-Based Force Generation. International Journal of Molecular Sciences, 24(17), 13112. https://doi.org/10.3390/ijms241713112