
CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Introduction of Mutations in the HY5 Gene Using CRISPR/Cas9 Technology for HY5-Deficient Chinese Cabbage Mutants
2.2. CRISPR/Cas9-Mediated Gene-Editing Results in Efficient Loss-of-Function HY5 Mutations in Transgenic Chinese Cabbage
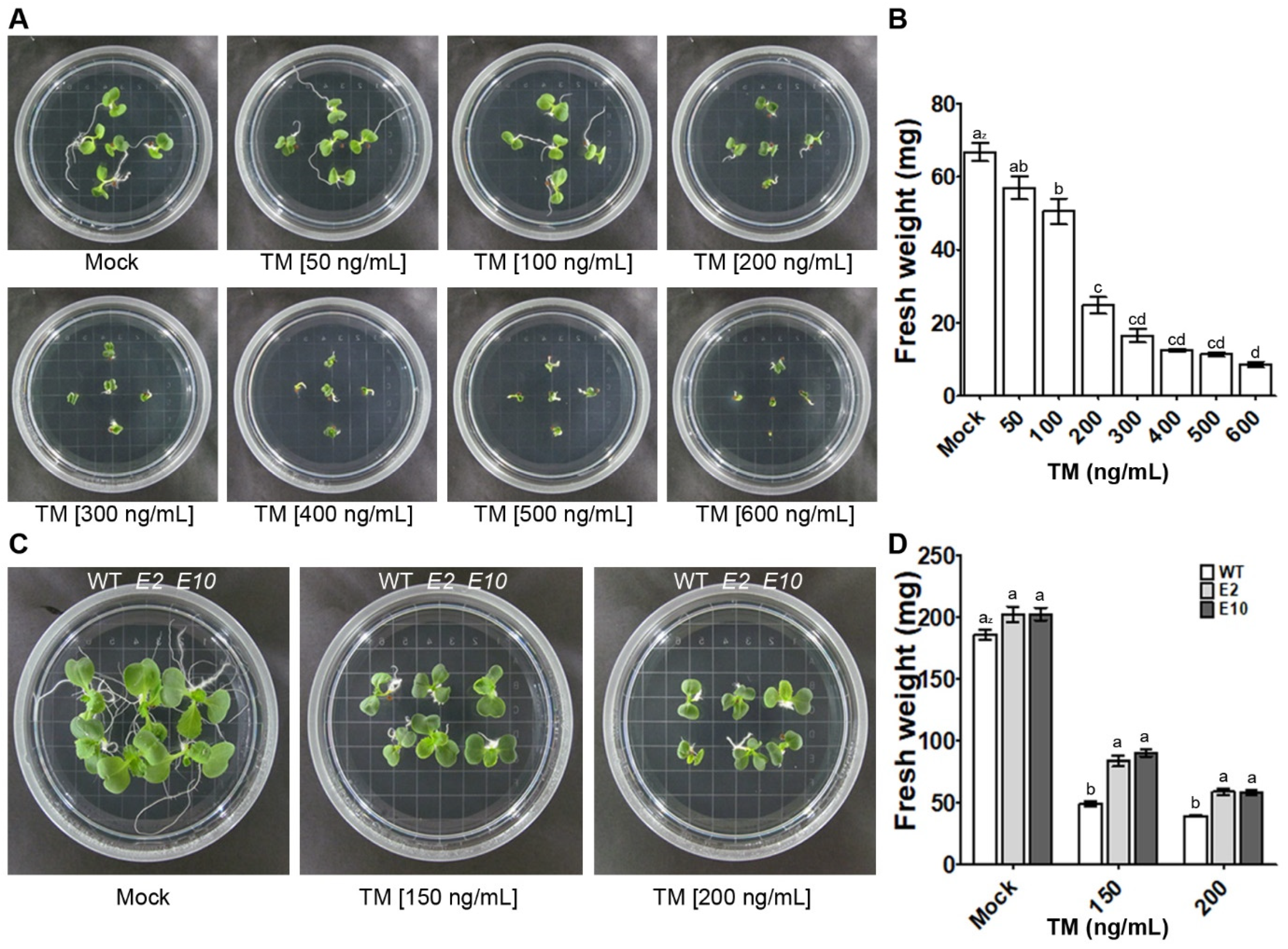
2.3. Mutations in the HY5 Gene Confer Resistance to Tunicamycin-Induced ER Stress in Chinese Cabbage
2.4. Mutations in the HY5 Gene Reduce ROS Generation in Chinese Cabbage
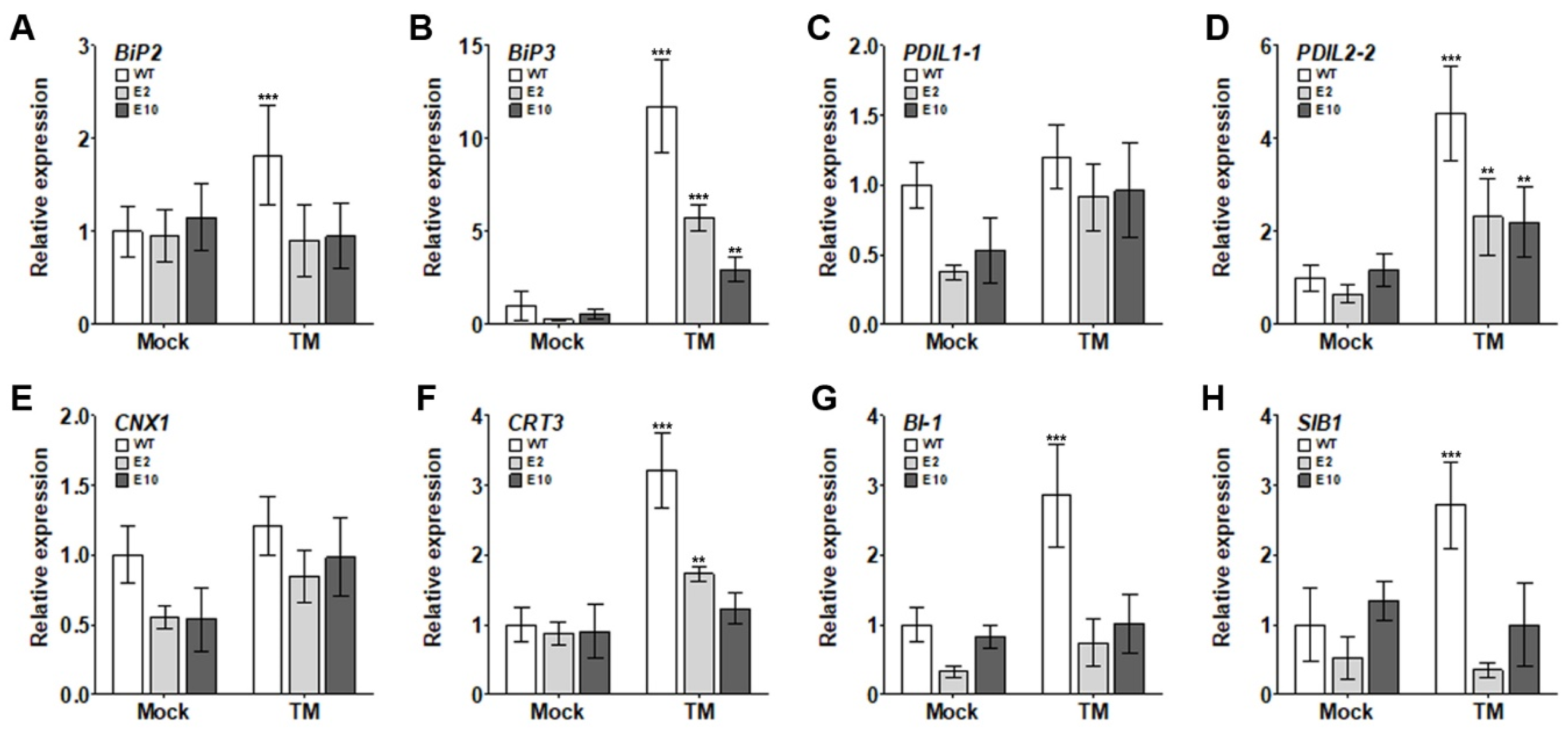
2.5. Alleviation of Growth Inhibition in hy5 Mutants under ER Stress Induced by TM Is Associated with Decreased Expression of UPR- and Cell-Death-Related Genes
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Sequence Information and Analysis
3.3. Selection of Target Sequences and Vector Construction
3.4. Agrobacterium-Mediated Transformation and Selection Procedures
3.5. Mutation Detection and Deep Sequence Analysis
3.6. Artificial Fertilization of Edited Plants
3.7. Tunicamycin (TM) Treatment
3.8. Nitroblue Tetrazolium (NBT) and 3,3′-Diaminobenzidine (DAB) Staining
3.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Xie, F.; Zha, J.; Tang, H.; Xu, Y.; Liu, X.; Wan, Z. Combining ability and heterosis analysis for mineral elements by using cytoplasmic male-sterile systems in non-heading Chinese cabbage (Brassica rapa). Crop Pasture Sci. 2018, 69, 296–302. [Google Scholar] [CrossRef]
- Dai, Y.; Sun, X.; Wang, C.; Li, F.; Zhang, S.; Zhang, H.; Li, G.; Yuan, L.; Chen, G.; Sun, R. Gene co-expression network analysis reveals key pathways and hub genes in Chinese cabbage (Brassica rapa L.) during vernalization. BMC Genom. 2021, 22, 236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dai, Y.; Yue, L.; Chen, G.; Yuan, L.; Zhang, S.; Li, F.; Zhang, H.; Li, G.; Zhu, S.; et al. Heat stress response in Chinese cabbage (Brassica rapa L.) revealed by transcriptome and physiological analysis. PeerJ 2022, 10, e13427. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Li, G.; Dai, Y.; Sun, X.; Li, F.; Zhang, S.; Zhang, H.; Sun, R. Gene co-expression network analysis of the heat-responsive core transcriptome identifies hub genes in Brassica rapa. Planta 2021, 253, 111. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Gao, S.; Zhang, B.; Ge, W.; Zhang, J.; Ji, R. Chinese Cabbage BrCAP Has Potential Resistance against Plasmodiophora brassicae. Horticulturae 2023, 9, 517. [Google Scholar] [CrossRef]
- Wei, X.; Rahim, M.A.; Zhao, Y.; Yang, S.; Wang, Z.; Su, H.; Li, L.; Niu, L.; Harun-Ur-Rashid, M.; Yuan, Y.; et al. Comparative Transcriptome Analysis of Early- and Late-Bolting Traits in Chinese Cabbage (Brassica rapa). Front. Genet. 2021, 12, 590830. [Google Scholar] [CrossRef]
- Wang, Y.; Zafar, N.; Ali, Q.; Manghwar, H.; Wang, G.; Yu, L.; Ding, X.; Ding, F.; Hong, N.; Jin, S. CRISPR/Cas Genome Editing Technologies for Plant Improvement against Biotic and Abiotic Stresses: Advances, Limitations, and Future Perspectives. Cells 2022, 11, 3928. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, S.K.; Lee, H.J.; Choi, C.S.; Park, S.T. Impacts of climate change on the growth, morphological and physiological responses, and yield of Kimchi cabbage leaves. Hortic. Environ. Biotechnol. 2016, 57, 470–477. [Google Scholar] [CrossRef]
- Qiu, N.; Liu, Q.; Li, J.; Zhang, Y.; Wang, F.; Gao, J. Physiological and Transcriptomic Responses of Chinese Cabbage (Brassica rapa L. ssp. Pekinensis) to Salt Stress. Int. J. Mol. Sci. 2017, 18, 1953. [Google Scholar] [CrossRef]
- Liu, J.X.; Howell, S.H. Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell 2010, 22, 2930–2942. [Google Scholar] [CrossRef]
- Duwi Fanata, W.I.; Lee, S.Y.; Lee, K.O. The unfolded protein response in plants: A fundamental adaptive cellular response to internal and external stresses. J. Proteom. 2013, 93, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Angelos, E.; Ruberti, C.; Kim, S.J.; Brandizzi, F. Maintaining the factory: The roles of the unfolded protein response in cellular homeostasis in plants. Plant J. 2017, 90, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, J.; Zhang, Z.; Quan, R.; Zhang, H.; Deng, X.W.; Ma, L.; Huang, R. Ethylene promotes hypocotyl growth and HY5 degradation by enhancing the movement of COP1 to the nucleus in the light. PLoS Genet. 2013, 9, e1004025. [Google Scholar] [CrossRef]
- Vitale, A.; Boston, R.S. Endoplasmic reticulum quality control and the unfolded protein response: Insights from plants. Traffic 2008, 9, 1581–1588. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, N.; Ujino, T.; Sano, H.; Chrispeels, M.J. Overexpression of a gene that encodes the first enzyme in the biosynthesis of asparagine-linked glycans makes plants resistant to tunicamycin and obviates the tunicamycin-induced unfolded protein response. Plant Physiol. 1999, 121, 353–361. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Ko, K.S.; Vu, B.N.; Lee, Y.E.; Yoon, S.H.; Pham, T.T.; Kim, J.-Y.; Lim, J.-M.; Kang, Y.J.; Hong, J.C.; et al. N-acetylglucosaminyltransferase II Is Involved in Plant Growth and Development Under Stress Conditions. Front. Plant Sci. 2021, 12, 761064. [Google Scholar] [CrossRef]
- Ko, K.S.; Yoo, J.Y.; Kim, K.H.; Hwang, B.Y.; Vu, B.N.; Lee, Y.E.; Choi, H.N.; Lee, Y.N.; Yun, J.; Park, J.Y.; et al. P58IPK facilitates plant recovery from ER stress by enhancing protein synthesis. Plant Biotechnol. Rep. 2022, 16, 665–681. [Google Scholar] [CrossRef]
- Ko, K.S.; Yoo, J.Y.; Vu, B.N.; Lee, Y.E.; Choi, H.N.; Lee, Y.N.; Fanata, W.I.D.; Harmoko, R.; Chung, W.S.; Hong, J.C.; et al. The role of protein phosphatase 2A (PP2A) in the unfolded protein response (UPR) of plants. Biochem. Biophys. Res. Commun. 2023, 670, 94–101. [Google Scholar] [CrossRef]
- Iwata, Y.; Koizumi, N. An Arabidopsis transcription factor, AtbZIP60, regulates the endoplasmic reticulum stress response in a manner unique to plants. Proc. Natl. Acad. Sci. USA 2005, 102, 5280–5285. [Google Scholar] [CrossRef]
- Liu, J.X.; Srivastava, R.; Che, P.; Howell, S.H. An endoplasmic reticulum stress response in Arabidopsis is mediated by proteolytic processing and nuclear relocation of a membrane-associated transcription factor, bZIP28. Plant Cell 2007, 19, 4111–4119. [Google Scholar] [CrossRef]
- Oyama, T.; Shimura, Y.; Okada, K. The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl. Genes. Dev. 1997, 11, 2983–2995. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.; Neff, M.M.; Hong, S.W.; Zhang, H.; Deng, X.W.; Xiong, L. Integration of light and abscisic acid signaling during seed germination and early seedling development. Proc. Natl. Acad. Sci. USA 2008, 105, 4495–4500. [Google Scholar] [CrossRef]
- De Lucas, M.; Daviere, J.M.; Rodriguez-Falcon, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blazquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef]
- Feng, S.; Martinez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Iglesias-Pedraz, J.M.; Kircher, S.; et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature 2008, 451, 475–479. [Google Scholar] [CrossRef]
- Li, J.; Li, G.; Gao, S.; Martinez, C.; He, G.; Zhou, Z.; Huang, X.; Lee, J.H.; Zhang, H.; Shen, Y.; et al. Arabidopsis transcription factor ELONGATED HYPOCOTYL5 plays a role in the feedback regulation of phytochrome A signaling. Plant Cell 2010, 22, 3634–3649. [Google Scholar] [CrossRef]
- Shi, Q.M.; Yang, X.; Song, L.; Xue, H.W. Arabidopsis MSBP1 is activated by HY5 and HYH and is involved in photomorphogenesis and brassinosteroid sensitivity regulation. Mol. Plant 2011, 4, 1092–1104. [Google Scholar] [CrossRef]
- Shin, D.H.; Choi, M.; Kim, K.; Bang, G.; Cho, M.; Choi, S.B.; Choi, G.; Park, Y.I. HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis. FEBS Lett. 2013, 587, 1543–1547. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-G.; Lee, G.-H.; Park, Y.-D. Characterization of a Drought-Tolerance Gene, BrDSR, in Chinese Cabbage. Hortic. Sci. Technol. 2016, 34, 102–111. [Google Scholar]
- Gangappa, S.N.; Botto, J.F. The Multifaceted Roles of HY5 in Plant Growth and Development. Mol. Plant 2016, 9, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, M.T.; Hardtke, C.S.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; He, H.; Wang, X.; Yang, X.; Li, L.; Deng, X.W. Genome-wide mapping of the HY5-mediated gene networks in Arabidopsis that involve both transcriptional and post-transcriptional regulation. Plant J. 2011, 65, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lory, N.; Stauber, J.; Hoecker, U. Photoreceptor Specificity in the Light-Induced and COP1-Mediated Rapid Degradation of the Repressor of Photomorphogenesis SPA2 in Arabidopsis. PLoS Genet. 2015, 11, e1005516. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Gohda, K.; Osterlund, M.T.; Oyama, T.; Okada, K.; Deng, X.W. HY5 stability and activity in arabidopsis is regulated by phosphorylation in its COP1 binding domain. EMBO J. 2000, 19, 4997–5006. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, Y.; Yang, W.; Chang, G.; Li, P.; Wei, J.; Yuan, X.; Huang, J.; Hu, X. The hydrogen sulfide signal enhances seed germination tolerance to high temperatures by retaining nuclear COP1 for HY5 degradation. Plant Sci. 2019, 285, 34–43. [Google Scholar] [CrossRef]
- Xu, D.; Li, J.; Gangappa, S.N.; Hettiarachchi, C.; Lin, F.; Andersson, M.X.; Jiang, Y.; Deng, X.W.; Holm, M. Convergence of Light and ABA signaling on the ABI5 promoter. PLoS Genet. 2014, 10, e1004197. [Google Scholar] [CrossRef]
- Lu, D.P.; Christopher, D.A. Light enhances the unfolded protein response as measured by BiP2 gene expression and the secretory GFP-2SC marker in Arabidopsis. Physiol. Plant 2008, 134, 360–368. [Google Scholar] [CrossRef]
- Nawkar, G.M.; Kang, C.H.; Maibam, P.; Park, J.H.; Jung, Y.J.; Chae, H.B.; Chi, Y.H.; Jung, I.J.; Kim, W.Y.; Yun, D.J.; et al. HY5, a positive regulator of light signaling, negatively controls the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 2084–2089. [Google Scholar] [CrossRef]
- Alquezar, B.; Bennici, S.; Carmona, L.; Gentile, A.; Pena, L. Generation of Transfer-DNA-Free Base-Edited Citrus Plants. Front. Plant Sci. 2022, 13, 835282. [Google Scholar] [CrossRef]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef]
- Li, Q.; Sapkota, M.; Van Der Knaap, E. Perspectives of CRISPR/Cas-mediated cis-engineering in horticulture: Unlocking the neglected potential for crop improvement. Hortic. Res. 2020, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Habben, J.E.; Archibald, R.L.; Drummond, B.J.; Chamberlin, M.A.; Williams, R.W.; Lafitte, H.R.; Weers, B.P. Overexpression of ARGOS Genes Modifies Plant Sensitivity to Ethylene, Leading to Improved Drought Tolerance in Both Arabidopsis and Maize. Plant Physiol. 2015, 169, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2017, 15, 207–216. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, X.; Su, W.; Hao, Y.; Liu, H.; Chen, R.; Song, S. BcSOC1 Promotes Bolting and Stem Elongation in Flowering Chinese Cabbage. Int. J. Mol. Sci. 2022, 23, 3459. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, W.; Zhou, Z.; Chen, H.; Xie, C.; Lin, Y. A new rice breeding method: CRISPR/Cas9 system editing of the Xa13 promoter to cultivate transgene-free bacterial blight-resistant rice. Plant Biotechnol. J. 2020, 18, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef]
- Catala, R.; Medina, J.; Salinas, J. Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 16475–16480. [Google Scholar] [CrossRef]
- Yang, B.; Song, Z.; Li, C.; Jiang, J.; Zhou, Y.; Wang, R.; Wang, Q.; Ni, C.; Liang, Q.; Chen, H.; et al. RSM1, an Arabidopsis MYB protein, interacts with HY5/HYH to modulate seed germination and seedling development in response to abscisic acid and salinity. PLoS Genet. 2018, 14, e1007839. [Google Scholar] [CrossRef]
- Yang, J.; Qu, X.; Li, T.; Gao, Y.; Du, H.; Zheng, L.; Ji, M.; Zhang, P.; Zhang, Y.; Hu, J.; et al. HY5-HDA9 orchestrates the transcription of HsfA2 to modulate salt stress response in Arabidopsis. J. Integr. Plant Biol. 2023, 65, 45–63. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Ko, K.S.; Yoo, J.Y.; Ramasamy, N.K.; Harmoko, R.; Vũ, B.N.T.; Park, J.Y.; Lee, K.O. Cell cycle arrest mediated by WEE1 is involved in the unfolded protein response in plants. Plant Biotechnol. Rep. 2018, 12, 315–328. [Google Scholar] [CrossRef]
- Ozgur, R.; Turkan, I.; Uzilday, B.; Sekmen, A.H. Endoplasmic reticulum stress triggers ROS signalling, changes the redox state, and regulates the antioxidant defence of Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Ruberti, C.; Lai, Y.; Brandizzi, F. Recovery from temporary endoplasmic reticulum stress in plants relies on the tissue-specific and largely independent roles of bZIP28 and bZIP60, as well as an antagonizing function of BAX-Inhibitor 1 upon the pro-adaptive signaling mediated by bZIP28. Plant J. 2018, 93, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Li, Z.; Li, M.; Dogra, V.; Lv, S.; Liu, R.; Lee, K.P.; Kim, C. Uncoupled Expression of Nuclear and Plastid Photosynthesis-Associated Genes Contributes to Cell Death in a Lesion Mimic Mutant. Plant Cell 2019, 31, 210–230. [Google Scholar] [CrossRef]
- Engler, C.; Youles, M.; Gruetzner, R.; Ehnert, T.M.; Werner, S.; Jones, J.D.; Patron, N.J.; Marillonnet, S. A golden gate modular cloning toolbox for plants. ACS Synth. Biol. 2014, 3, 839–843. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar]
- Shokralla, S.; Porter, T.M.; Gibson, J.F.; Dobosz, R.; Janzen, D.H.; Hallwachs, W.; Golding, G.B.; Hajibabaei, M. Massively parallel multiplex DNA sequencing for specimen identification using an Illumina MiSeq platform. Sci. Rep. 2015, 5, 9687. [Google Scholar] [CrossRef]
- Ramel, F.; Sulmon, C.; Bogard, M.; Couee, I.; Gouesbet, G. Differential patterns of reactive oxygen species and antioxidative mechanisms during atrazine injury and sucrose-induced tolerance in Arabidopsis thaliana plantlets. BMC Plant Biol. 2009, 9, 28. [Google Scholar] [CrossRef]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B. Histochemical Detection of Superoxide and H2O2 Accumulation in Brassica juncea Seedlings. Bio-Protoc. 2014, 4, e1108. [Google Scholar] [CrossRef]
- Radke, S.E.; Turner, J.C.; Facciotti, D. Transformation and regeneration of Brassica rapa using Agrobacterium tumefaciens. Plant Cell Rep. 1992, 11, 499–505. [Google Scholar] [CrossRef]
- Lee, M.-K.; Kim, H.-S.; Kim, J.-S.; Kim, S.-H.; Park, Y.-D. Agrobacterium-mediated transformation system for large-scale producion of transgenic chinese cabbage (Brassica rapa L. ssp. pekinensis) plants for insertional mutagenesis. J. Plant Biol. 2004, 47, 300–306. [Google Scholar] [CrossRef]
- Khan, U.M.; Shaheen, N.; Farooq, A.; Maqbool, R.; Khan, S.H.; Azhar, M.T.; Rana, I.A.; Seo, H. Optimization of Regeneration and Agrobacterium-Mediated Transformation Protocols for Bi and Multilocular Varieties of Brassica rapa. Plants 2022, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.S.; Cao, J.; Ren, J.P.; Earle, E.D. Control of Lepidopteran insect pests in transgenic Chinese cabbage (Brassica rapa ssp. pekinensis) transformed with a synthetic Bacillus thuringiensis cry1C gene. Plant Cell Rep. 2001, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Siddique, M.I.; Kim, D.S.; Lee, E.S.; Han, K.; Kim, S.G.; Lee, H.E. CRISPR/Cas9-mediated gene editing to confer turnip mosaic virus (TuMV) resistance in Chinese cabbage (Brassica rapa). Hortic. Res. 2023, 10, uhad078. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleotide Sequence (5′->3′) | Type | Mutant Frequency (%) | Reads Number | Total Reads Number | ||
|---|---|---|---|---|---|---|
| E2 | gRNA1 | GTGTAAATAAAAAGGTTGTTGAGG | WT | 4.38 | 1337 | 30,547 |
| GTGTAAATAAAAAG– –GTTGAGG | −3 | 62.3 | 19,039 | |||
| GTGTAAATAAAAAGGTTGTTTGAGG | +1 | 33.3 | 10,171 | |||
| gRNA2 | GGAAAATAGAGTGAAAGACTTGG | WT | 7.91 | 3775 | 47,718 | |
| GGAAAATAGAGTGAA– –ACTTGG | −2 | 57.5 | 27,429 | |||
| GGAAAATAGAGTGA– –ACTTGG | −3 | 34.6 | 16,514 | |||
| E10 | gRNA1 | GTGTAAATAAAAAGGTTGTTGAGG | WT | 2.7 | 946 | 34,503 |
| GTGTAAATAAAAAG– –GTTGAGG | −3 | 62.2 | 21,454 | |||
| GTGTAAATAAAAAGGTTGTTTGAGG | +1 | 35.1 | 12,103 | |||
| gRNA2 | GGAAAATAGAGTGAAAGACTTGG | WT | 2.9 | 1320 | 46,112 | |
| GGAAAATAGAGTGAA– –ACTTGG | −2 | 56.3 | 25,961 | |||
| GGAAAATAGAGTGA– –ACTTGG | −3 | 40.8 | 18,831 |
| Gene | Arabidopsis | Gene | Brassica rapa | Gene Annotation |
|---|---|---|---|---|
| CNX1 | AT5G61790 | BrCNX1 | Bra035913 | Calnexin 1, unfolded protein binding |
| CRT3 | AT1G08450 | BrCRT3 | Bra031627 | Calreticulin 3, unfolded protein binding |
| BiP2 | AT5G42020 | BrBiP2 | Bra015047 | Heat shock protein 70 (Hsp 70) family protein |
| BiP3 | AT1G09080 | BrBiP3 | Bra031657 | Heat shock protein 70 (Hsp 70) family protein |
| PDIL1-1 | AT1G21750 | BrPDI1-1 | Bra016405 | PDI-like 1-1, protein disulfide isomerase activity |
| PDIL2-2 | AT1G04980 | BrPDI2-2 | Bra015375 | PDI-like 2-2, protein disulfide isomerase activity |
| BI-1 | AT5G47120 | BrBI-1 | Bra022106 | BAX inhibitor 1, Functions as an attenuator of biotic and abiotic types of cell death |
| SIB1 | AT3G56710 | BrSIB1 | Bra007265 | sigma factor binding protein 1, defense responses |
| TUB | AT5G12250 | BrTUB | Bra008903 | beta-6 tubulin, internal control |
| Primer Name | Sequence (5′ to 3′) | Gene |
|---|---|---|
| CNX1 F | ATCCCTGACAAGACCATCC | BrCNX1 |
| CNX1 R | CCTCCCACATACCATCTTCC | BrCNX1 |
| CRT3 F | TCTTCTCTCTTCTCACTCTCAC | BrCRT3 |
| CRT3 R | CTTTATTGTCAGGATCGCCG | BrCRT3 |
| BiP2 F | GGGAAGCCGTACATTCAAG | BrBiP2 |
| BiP2 R | GAGACCAGCAATAACACCAG | BrBiP2 |
| BiP3 F | CTGACTTCTCTGAGCCTTTAAC | BrBiP3 |
| BiP3 R | TCTTCACCACCTTCACCAC | BrBiP3 |
| PDIL1-1 F | AAAATCTCAACCCATCCCAACC | BrPDIL1-1 |
| PDIL1-1 R | ATGACACAGCGACTTCGTCC | BrPDIL1-1 |
| PDIL2-2 F | CTTGAAGCCAATGCTGGAC | BrPDIL2-2 |
| PDIL2-2 R | ACCATATCCTCCAACTCCAAC | BrPDIL2-2 |
| BI-1 F | CATCCTCATCACTGCGTTTG | BrBI-1 |
| BI-1 R | AGTGTCCACCACCATGTATC | BrBI-1 |
| SIB1 F | TTCAGCCAACAAAGCCATC | BrSIB1 |
| SIB1 R | AACATCTCTTCACCATCCAAC | BrSIB1 |
| TUB F | TGACTGTCTTCAGGGTTTCCA | TUBULIN |
| TUB R | CACCGTGTCCGAGACCTTAG | TUBULIN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Ko, K.S.; Lee, H.E.; Lee, E.S.; Han, K.; Yoo, J.Y.; Vu, B.N.; Choi, H.N.; Lee, Y.N.; Hong, J.C.; et al. CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress. Int. J. Mol. Sci. 2023, 24, 13105. https://doi.org/10.3390/ijms241713105
Lee YR, Ko KS, Lee HE, Lee ES, Han K, Yoo JY, Vu BN, Choi HN, Lee YN, Hong JC, et al. CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress. International Journal of Molecular Sciences. 2023; 24(17):13105. https://doi.org/10.3390/ijms241713105
Chicago/Turabian StyleLee, Ye Rin, Ki Seong Ko, Hye Eun Lee, Eun Su Lee, Koeun Han, Jae Yong Yoo, Bich Ngoc Vu, Ha Na Choi, Yoo Na Lee, Jong Chan Hong, and et al. 2023. "CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress" International Journal of Molecular Sciences 24, no. 17: 13105. https://doi.org/10.3390/ijms241713105
APA StyleLee, Y. R., Ko, K. S., Lee, H. E., Lee, E. S., Han, K., Yoo, J. Y., Vu, B. N., Choi, H. N., Lee, Y. N., Hong, J. C., Lee, K. O., & Kim, D. S. (2023). CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress. International Journal of Molecular Sciences, 24(17), 13105. https://doi.org/10.3390/ijms241713105