
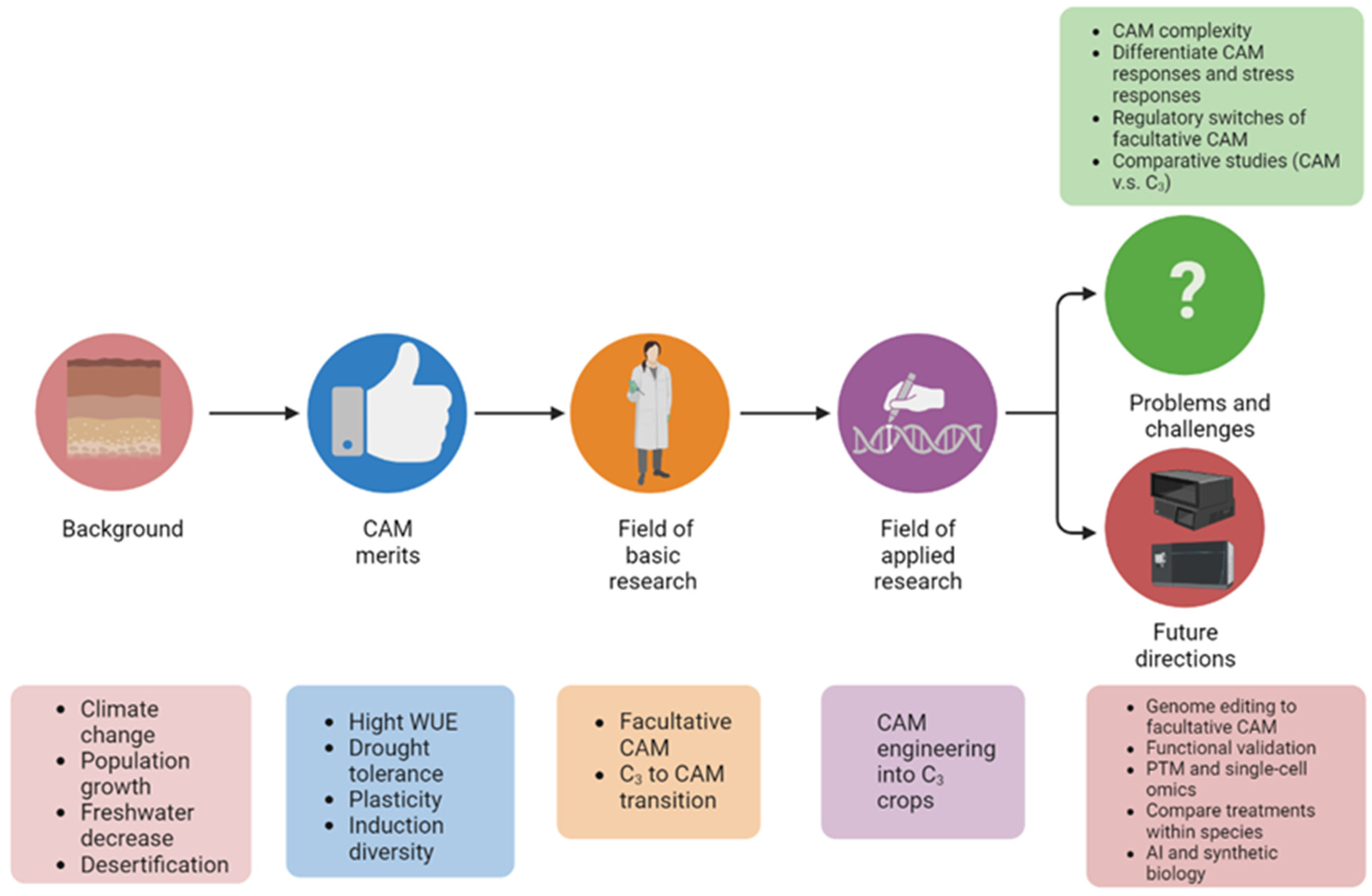
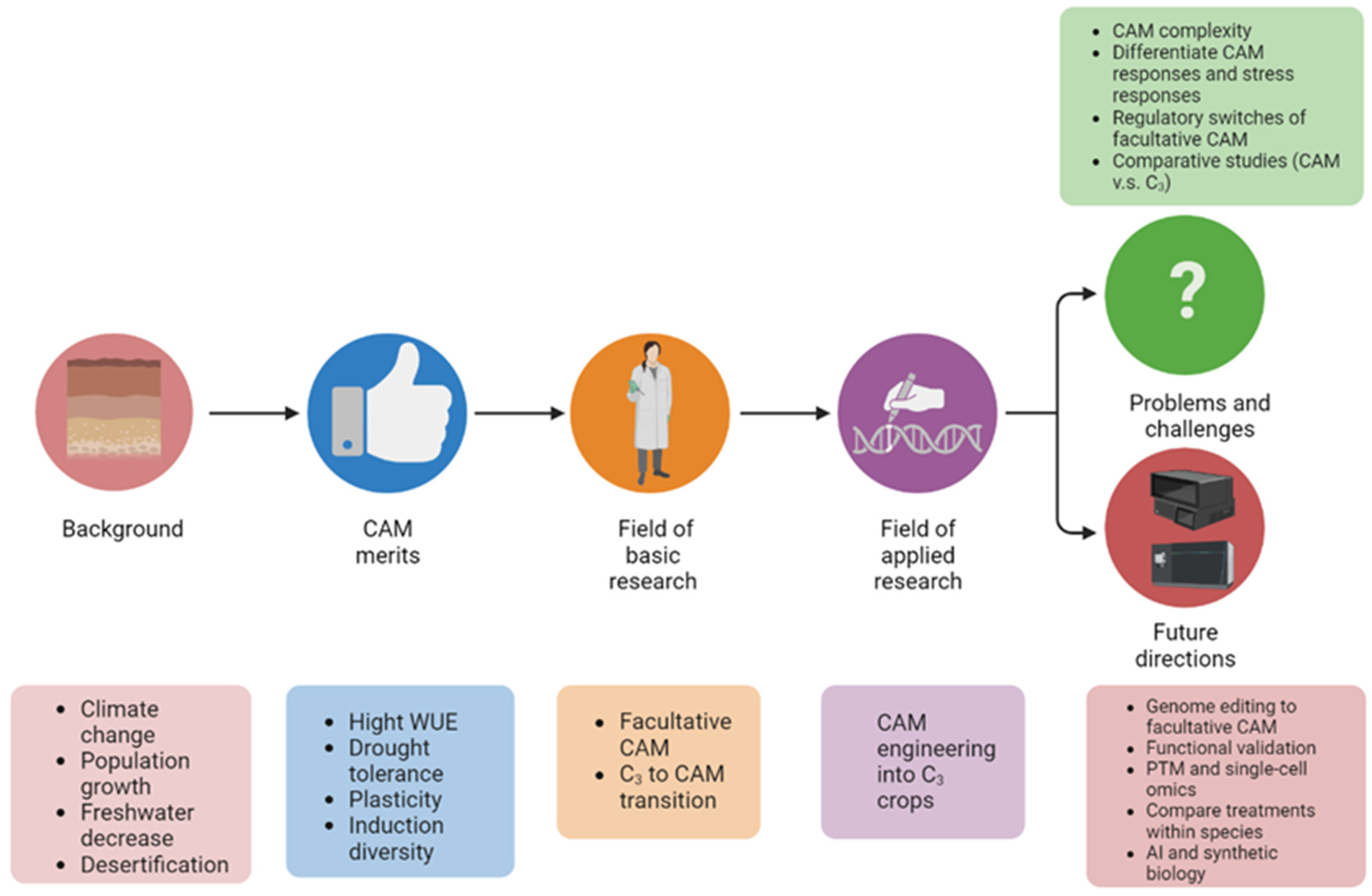
Defining Mechanisms of C3 to CAM Photosynthesis Transition toward Enhancing Crop Stress Resilience



Abstract
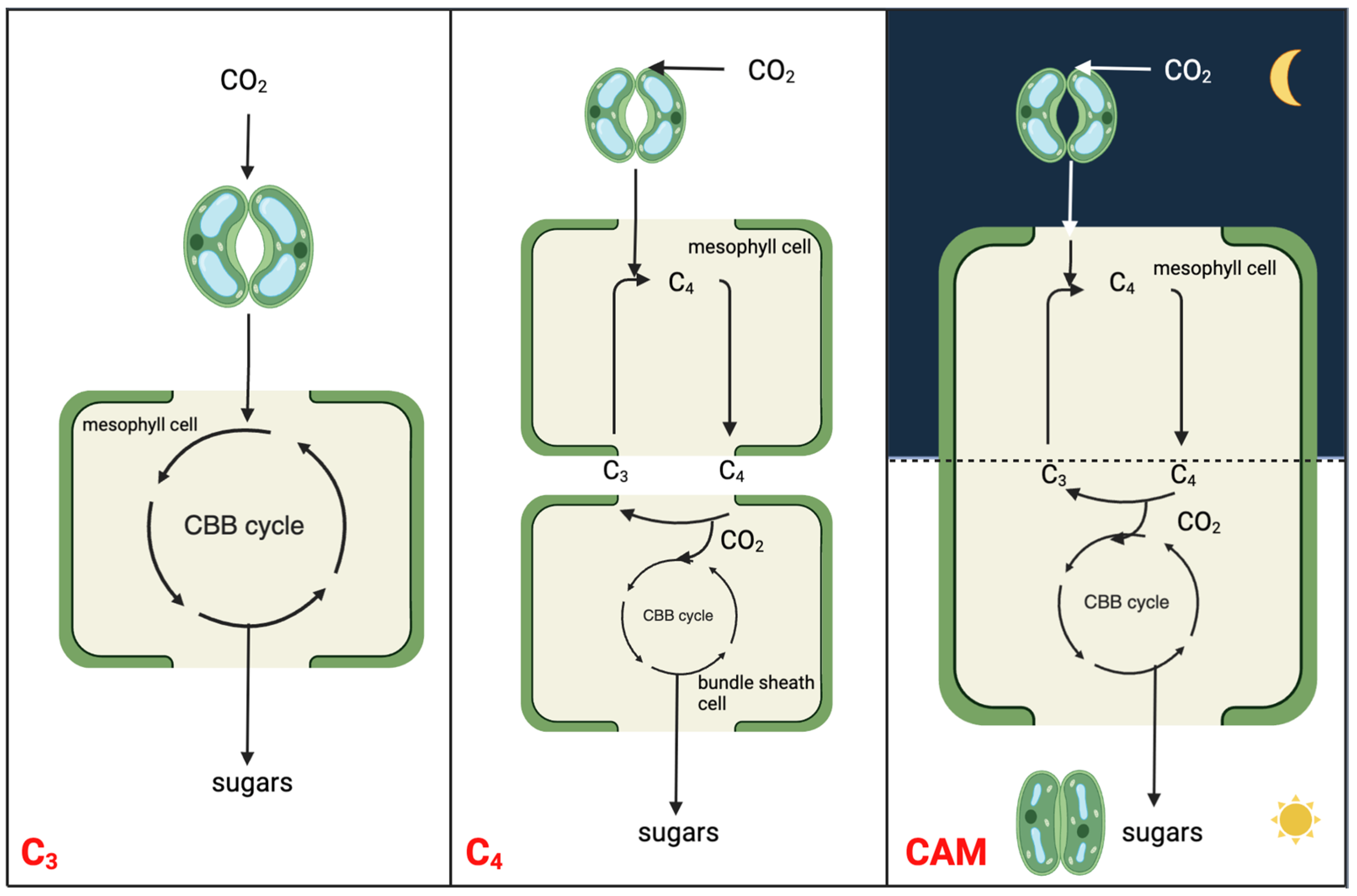
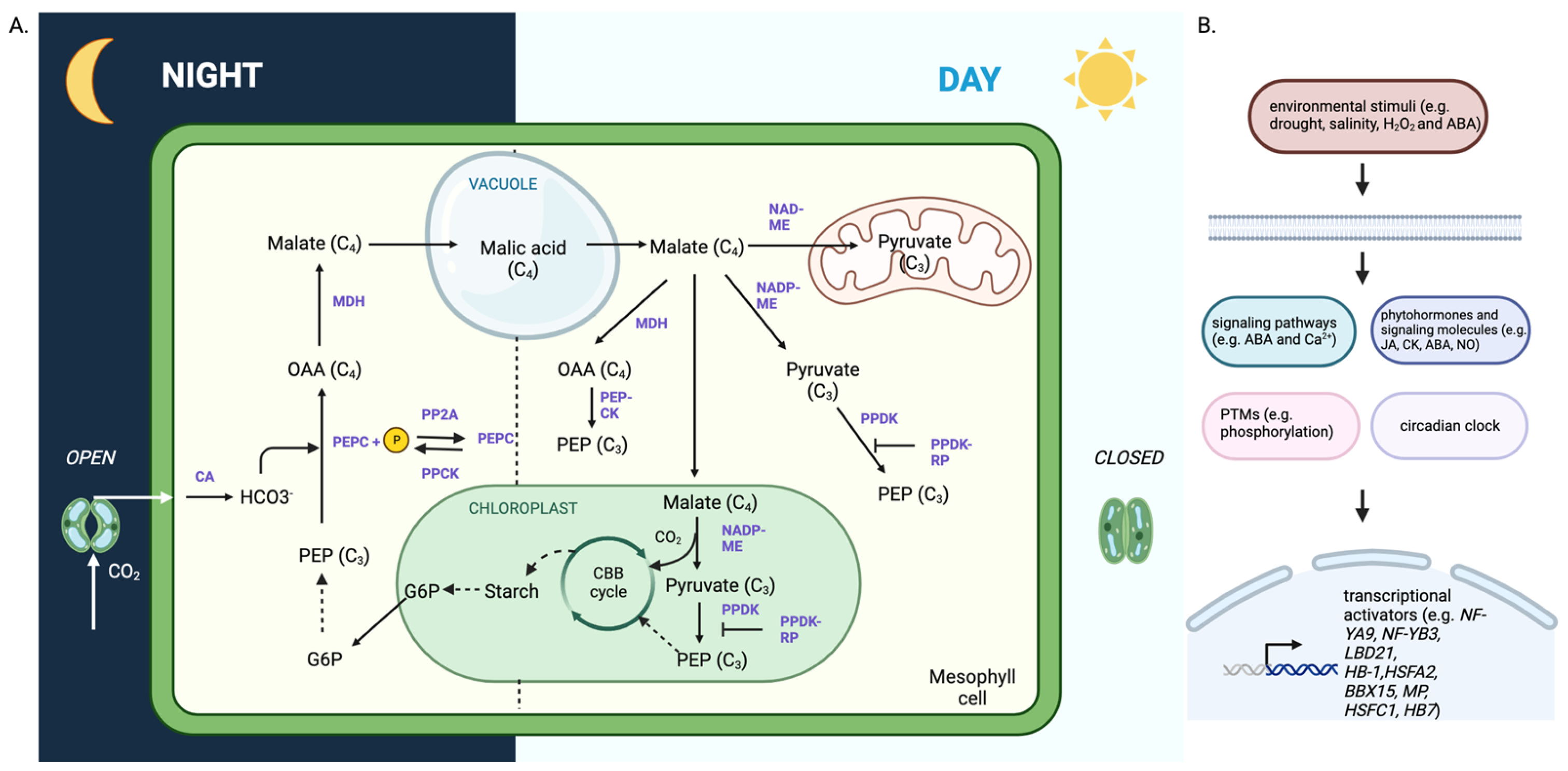
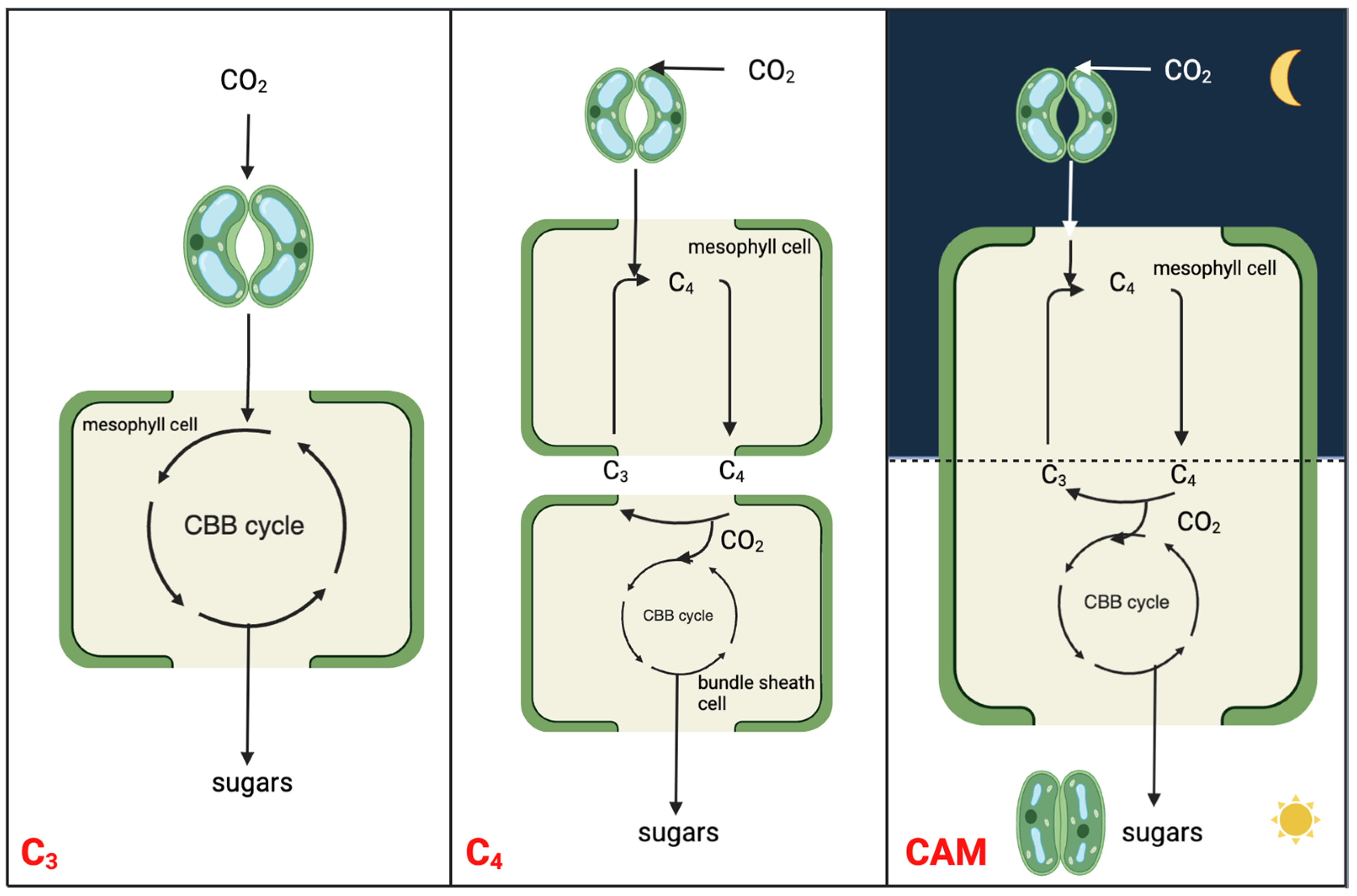
:1. Introduction
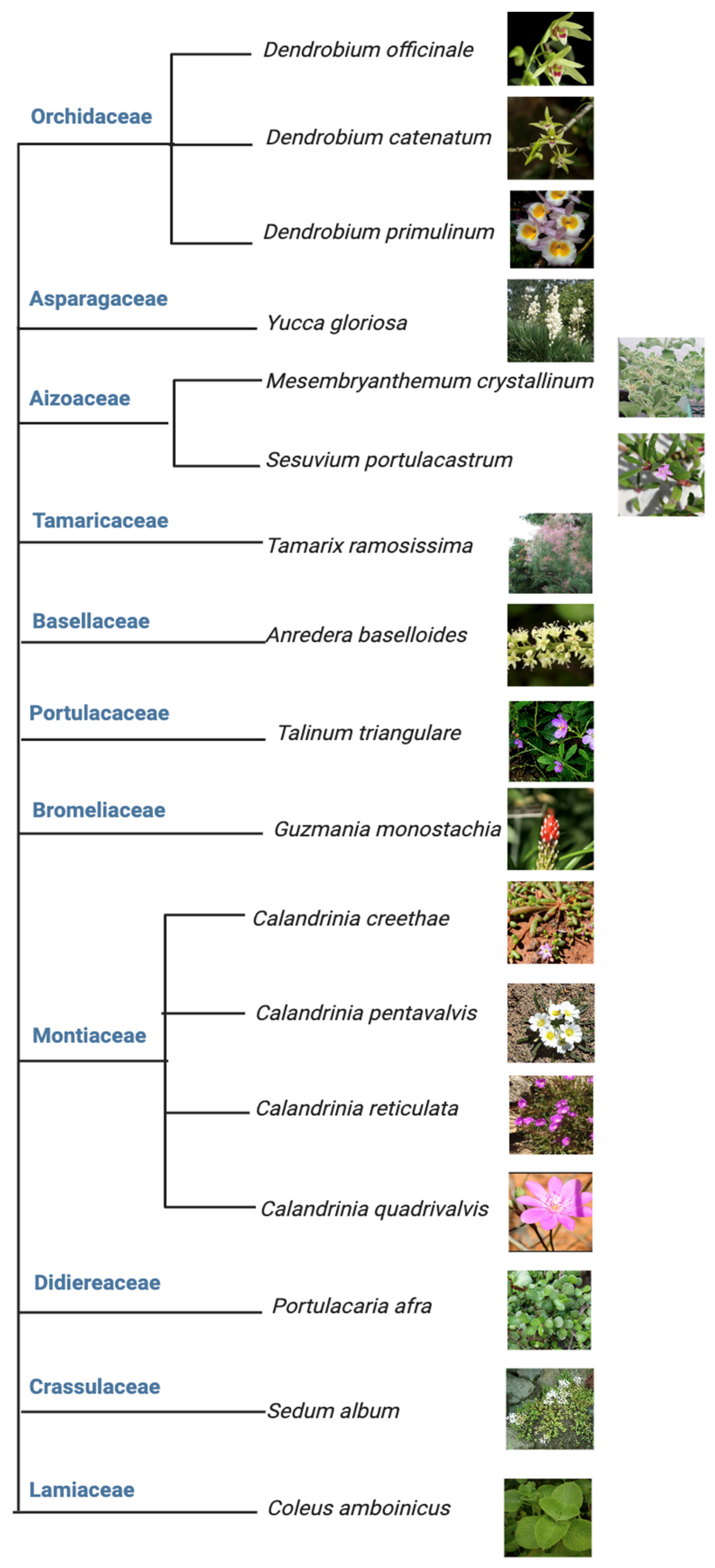
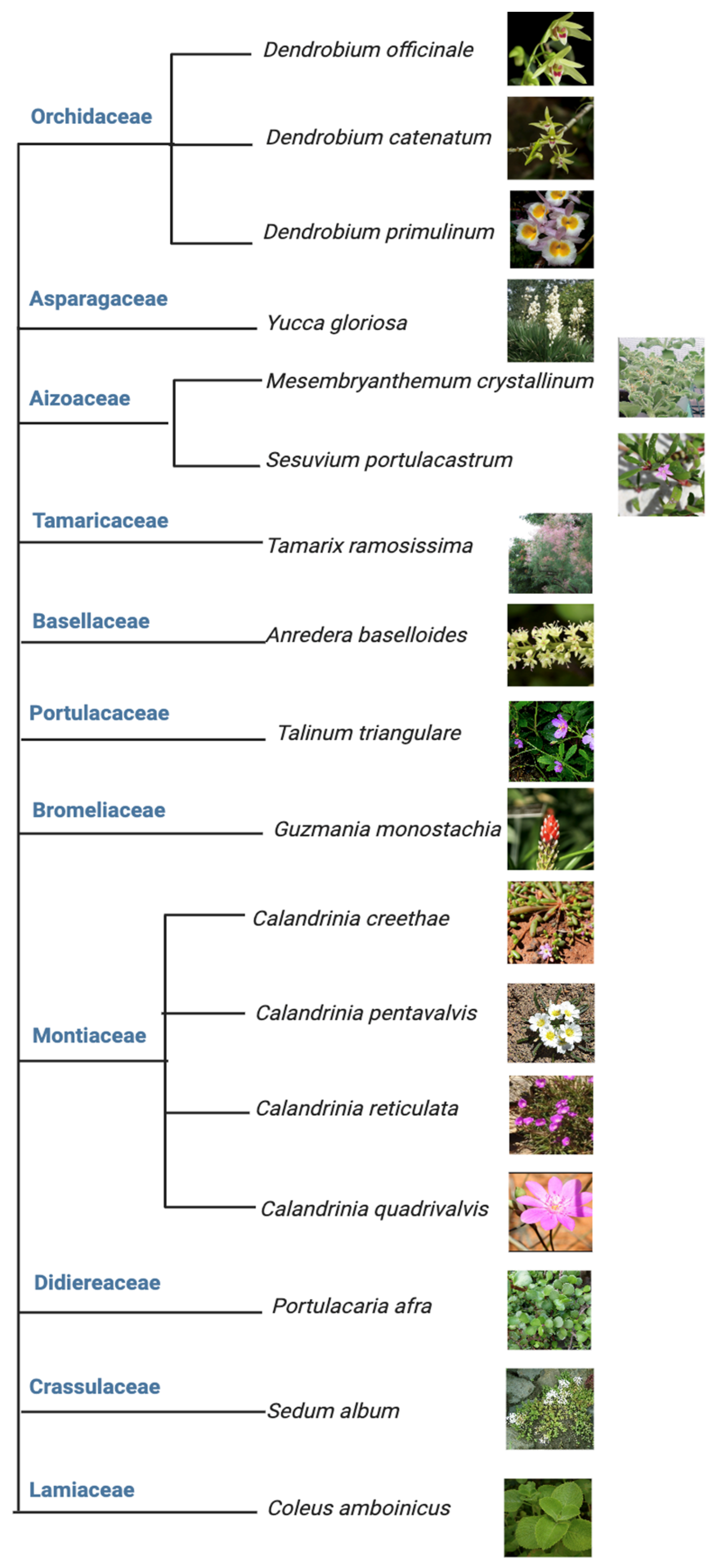
2. The Plasticity of CAM Is Best Represented in Facultative CAM
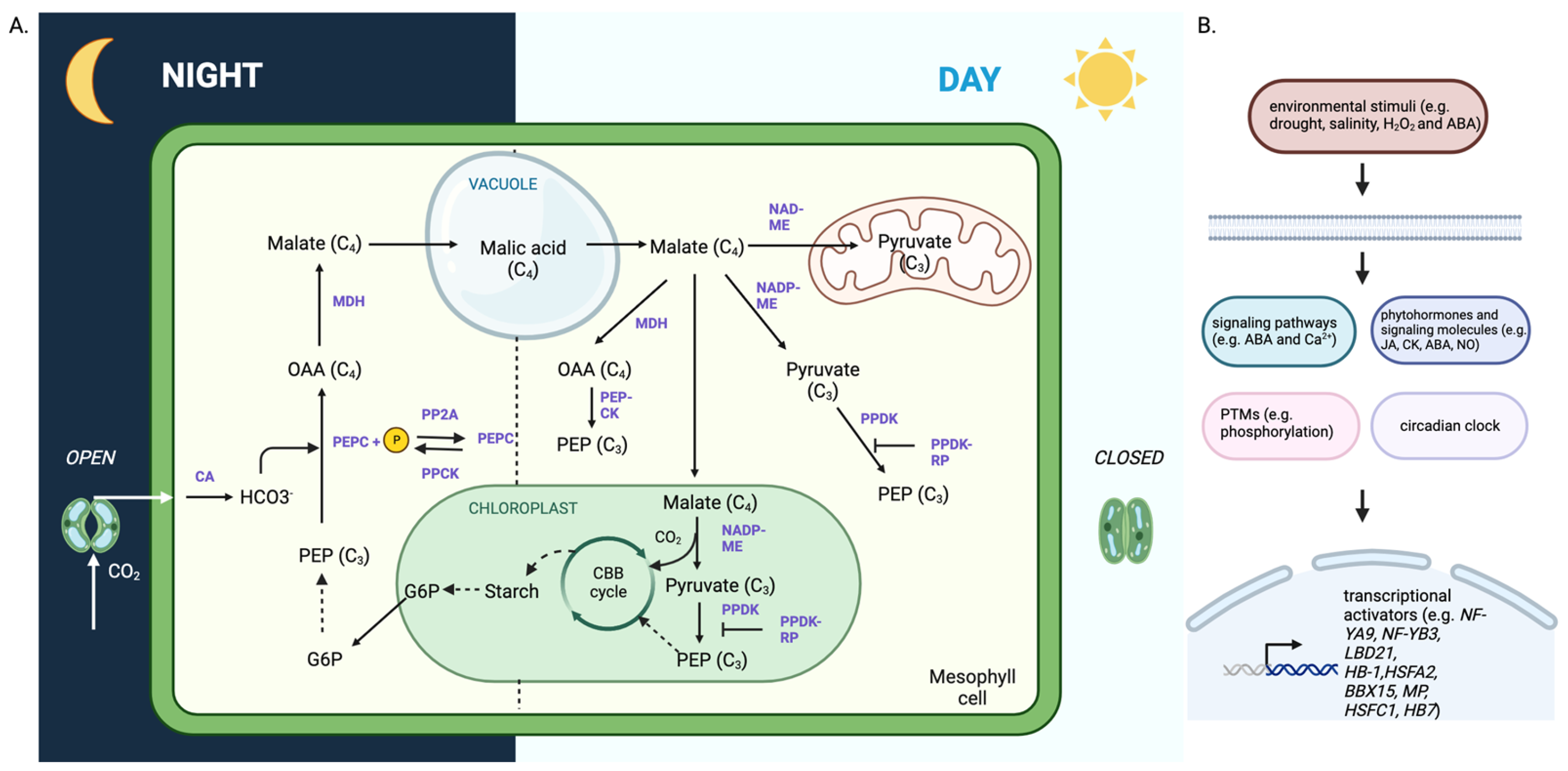
3. Studies on C3 to CAM Transition Revealed Important Molecular Players
4. CAM Engineering toward Solving the Global Climate Challenges
5. Pressing Problems and Challenges in CAM Research
6. Perspectives and Future Directions
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leisner, C.P. Review: Climate change impacts on food security- focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef] [PubMed]
- Reidmiller, D.R.; Avery, C.W.; Easterling, D.R.; Kunkel, K.E.; Lewis, K.L.M.; Maycock, T.K.; Stewart, B.C. (Eds.) USGCRP Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment; U.S. Global Change Research Program: Washington, DC, USA, 2018; Volume II, p. 1515. [Google Scholar] [CrossRef]
- Yao, J.; Liu, H.; Huang, J.; Gao, Z.; Wang, G.; Li, D.; Yu, H.; Chen, X. Accelerated dryland expansion regulates future variability in dryland gross primary production. Nat. Commun. 2020, 11, 1665. [Google Scholar] [CrossRef] [PubMed]
- Eshel, G.; Araus, V.; Undurraga, S.; Soto, D.C.; Moraga, C.; Montecinos, A.; Moyano, T.; Maldonado, J.; Díaz, F.P.; Varala, K.; et al. Plant ecological genomics at the limits of life in the Atacama Desert. Proc. Natl. Acad. Sci. USA 2021, 118, e2101177118. [Google Scholar] [CrossRef]
- Lim, S.D.; Mayer, J.A.; Yim, W.C.; Cushman, J.C. Plant tissue succulence engineering improves water-use efficiency, water-deficit stress attenuation and salinity tolerance in Arabidopsis. Plant J. 2020, 103, 1049–1072. [Google Scholar] [CrossRef]
- Hu, R.; Zhang, J.; Jawdy, S.; Sreedasyam, A.; Lipzen, A.; Wang, M.; Ng, V.; Daum, C.; Keymanesh, K.; Liu, D.; et al. Comparative genomics analysis of drought response between obligate CAM and C3 photosynthesis plants. J. Plant Physiol. 2022, 277, 153791. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 1–10. [Google Scholar]
- Crayn, D.M.; Winter, K.; Smith, J.A.C. Multiple origins of crassulacean acid metabolism and the epiphytic habit in the Neotropical family Bromeliaceae. Proc. Natl. Acad. Sci. USA 2004, 101, 3703–3708. [Google Scholar] [CrossRef]
- Ceusters, J.; Van De Poel, B. Ethylene Exerts Species-Specific and Age-Dependent Control of Photosynthesis. Plant Physiol. 2018, 176, 2601–2612. [Google Scholar] [CrossRef]
- Heyduk, K. Evolution of Crassulacean acid metabolism in response to the environment: Past, present, and future. Plant Physiol. 2022, 190, 19–30. [Google Scholar] [CrossRef]
- Amin, A.B.; Rathnayake, K.N.; Yim, W.C.; Garcia, T.M.; Wone, B.; Cushman, J.C.; Wone, B.W.M. Crassulacean Acid Metabolism Abiotic Stress-Responsive Transcription Factors: A Potential Genetic Engineering Approach for Improving Crop Tolerance to Abiotic Stress. Front. Plant Sci. 2019, 10, 129. [Google Scholar] [CrossRef]
- Wickell, D.; Kuo, L.-Y.; Yang, H.-P.; Ashok, A.D.; Irisarri, I.; Dadras, A.; de Vries, S.; de Vries, J.; Huang, Y.-M.; Li, Z.; et al. Underwater CAM photosynthesis elucidated by Isoetes genome. Nat. Commun. 2021, 12, 6348. [Google Scholar] [CrossRef]
- Osmond, C.B. Crassulacean Acid Metabolism: A Curiosity in Context. Annu. Rev. Plant Physiol. 1978, 29, 379–414. [Google Scholar] [CrossRef]
- Borland, A.M.; Guo, H.-B.; Yang, X.; Cushman, J.C. Orchestration of carbohydrate processing for crassulacean acid metabolism. Curr. Opin. Plant Biol. 2016, 31, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, J.; Leegood, R. Photosynthesis. In Plant Biochemistry; Dey, P.M., Harborne, J.B., Eds.; Academic Press: London, UK, 1997; pp. 1–4, 49–110. [Google Scholar] [CrossRef]
- Drennan, P.M.; Nobel, P.S. Responses of CAM species to increasing atmospheric CO2 concentrations. Plant Cell Environ. 2000, 23, 767–781. [Google Scholar] [CrossRef]
- Schiller, K.; Bräutigam, A. Engineering of Crassulacean Acid Metabolism. Annu. Rev. Plant Biol. 2021, 72, 77–103. [Google Scholar] [CrossRef] [PubMed]
- Kluge, M.; Ting, I.P. Crassulacean Acid Metabolism; Springer: Berlin/Heidelberg, Germany, 1978; pp. 108–152. [Google Scholar] [CrossRef]
- Kong, W.; Yoo, M.J.; Zhu, D.; Noble, J.D.; Kelley, T.M.; Li, J.; Kirst, M.; Assmann, S.M.; Chen, S. Molecular changes in Mesembryanthemum crystallinum guard cells underlying the C3 to CAM transition. Plant Mol. Biol. 2020, 103, 653–667. [Google Scholar] [CrossRef]
- Guan, Q.; Tan, B.; Kelley, T.M.; Tian, J.; Chen, S. Physiological changes in Mesembryanthemum crystallinum during the C3 to CAM transition induced by salt stress. Front. Plant Sci. 2020, 11, 283. [Google Scholar] [CrossRef]
- Ferrari, R.C.; Kawabata, A.B.; Ferreira, S.S.; Hartwell, J.; Freschi, L. A matter of time: Regulatory events behind the synchronization of C4 and crassulacean acid metabolism in Portulaca oleracea. J. Exp. Bot. 2022, 73, 4867–4885. [Google Scholar] [CrossRef]
- Gilman, I.S.; Moreno-Villena, J.J.; Lewis, Z.R.; Goolsby, E.W.; Edwards, E.J. Gene co-expression reveals the modularity and integration of C4 and CAM in Portulaca. Plant Physiol. 2022, 189, 735–753. [Google Scholar] [CrossRef]
- Ferrari, R.C.; Bittencourt, P.P.; Rodrigues, M.A.; Moreno-Villena, J.J.; Alves, F.R.R.; Gastaldi, V.D.; Boxall, S.F.; Dever, L.V.; Demarco, D.; Andrade, S.C.S.; et al. C4 and crassulacean acid metabolism within a single leaf: Deciphering key components behind a rare photosynthetic adaptation. New Phytol. 2020, 225, 1699–1714. [Google Scholar] [CrossRef]
- Gilman, I.S.; Edwards, E.J. Crassulacean acid metabolism. Curr. Biol. 2020, 30, R57–R62. [Google Scholar] [CrossRef]
- Cushman, J.C. Crassulacean acid metabolism. A plastic photosynthetic adaptation to arid environments. Plant Physiol. 2001, 127, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.; Martin, C.E.; Tezara, W.; Ballestrini, C.; Medina, E. Induction by drought of Crassulacean acid metabolism in the terrestrial bromeliad, Puya floccose. Photosynthetica 2010, 48, 383–388. [Google Scholar] [CrossRef]
- Winter, K. Ecophysiology of constitutive and facultative CAM photosynthesis. J. Exp. Bot. 2019, 70, 6495–6508. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; Holtum, J.A.M. Facultative Crassulacean acid metabolism (CAM) plants: Powerful tools for unravelling the functional elements of CAM photosynthesis. J. Exp. Bot. 2014, 65, 3425–3441. [Google Scholar] [CrossRef] [PubMed]
- Borland, A.M.; Griffiths, H. The regulation of CAM and respiratory recycling by water supply and light regime in the C3 -CAM intermediate Sedum telephium. Funct. Ecol. 1990, 4, 33. [Google Scholar] [CrossRef]
- Olivares, E.; Urich, R.; Montes, G.; Coronel, I.; Herrera, A. Occurrence of Crassulacean acid metabolism in Cissus trifoliata L. (Vitaceae). Oecologia 1984, 61, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; von Willert, D.J. NaCl-induzierter crassulaceensäurestoffwechsel bei Mesembryanthemum crystallinum. Z. Pflanzenphysiol. 1972, 67, 166–170. [Google Scholar] [CrossRef]
- Maxwell, K. Resistance is useful: Diurnal patterns of photosynthesis in C3 and Crassulacean acid metabolism epiphytic bromeliads. Funct. Plant Biol. 2002, 29, 679–687. [Google Scholar] [CrossRef]
- Taybi, T.; Cushman, J.C. Abscisic acid signaling and protein synthesis requirements for phosphoenolpyruvate carboxylase transcript induction in the common ice plant. J. Plant Physiol. 2002, 159, 1235–1243. [Google Scholar] [CrossRef]
- Brulfert, J.; Kluge, M.; Güçlü, S.; Queiroz, O. Interaction of photoperiod and drought as CAM inducing factors in Kalanchoë blossfeldiana Poelln., cv. Tom Thumb. J. Plant Physiol. 1988, 133, 222–227. [Google Scholar] [CrossRef]
- Surówka, E.; Dziurka, M.; Kocurek, M.; Goraj, S.; Rapacz, M.; Miszalski, Z. Effects of exogenously applied hydrogen peroxide on antioxidant and osmoprotectant profiles and the C3-CAM shift in the halophyte Mesembryanthemum crystallinum L. J. Plant Physiol. 2016, 200, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; Ziegler, H. Induction of Crassulacean acid metabolism in Mesembryanthemum crystallinum increases reproductive success under conditions of drought and salinity stress. Oecologia 1992, 92, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Sujkowska-rybkowska, M.; Miszalski, Z.; Kornaś, A. At the edges of photosynthetic metabolic plasticity—On the rapidity and extent of changes accompanying salinity stress-induced cam photosynthesis withdrawal. Int. J. Mol. Sci. 2021, 22, 8426. [Google Scholar] [CrossRef] [PubMed]
- Snotch from Sapporo, Hokkaido, Japan. Available online: https://commons.wikimedia.org/wiki/File:Dendrobium_officinale_Kimura_%26_Migo_J._Shanghai_Sci._Inst._3-_122_(1936)_(33547665894)_-_cropped.jpg (accessed on 10 August 2023). This Image Is Licensed under the Creative Commons Attribution 2.0 Generic License.
- Maria at Blogger. Available online: http://mariasorchids.blogspot.com/2018/02/a-display-of-dendrobiums.html (accessed on 10 August 2023). This Work Is Licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
- Ricardo. Available online: http://ricardogupi.blogspot.com/2010/10/dendrobium-primulinum-also-known-now-as.html (accessed on 10 August 2023). No License Found.
- NC State Extension Gardener. Available online: https://plants.ces.ncsu.edu/plants/yucca-gloriosa/ (accessed on 10 August 2023). No License Found.
- Dezmond Wells. Available online: https://www.inaturalist.org/taxa/130743-Sesuvium-portulacastrum (accessed on 10 August 2023). This Image Is under the Creative Commons Attribution-Share-Alike License 3.0.
- Jerzy Opioła. Available online: https://commons.wikimedia.org/wiki/File:Tamarix_ramosissima_a2.jpg (accessed on 10 August 2023). This Work Is Licensed under the Creative Commons Attribution-Share Alike 3.0 Unported License.
- Rojas-Sandoval, J.; Acevedo-Rodríguez, P. Anredera baselloides (Madeira vine). CABI Compend. 2014. [Google Scholar] [CrossRef]
- Alex Popovkin. Available online: https://ayurwiki.org/Ayurwiki/Talinum_triangulare (accessed on 10 August 2023). This Image Is Licensed under the Creative Commons Attribution 2.0 Generic License.
- Cliff at Wikipedia. Available online: https://en.wikipedia.org/wiki/Guzmania_monostachia (accessed on 10 August 2023). This Work Is under the Creative Commons Attribution-ShareAlike License 4.0.
- Roger Fryer; Jill Newland. Available online: http://www.northqueenslandplants.com/Australian%20Plant%20Families%20N-S/Portulacaceae/Calandrinia/Calandrinia%20creethae.html (accessed on 10 August 2023). This Work Is Licensed under a Creative Commons Attribution-Noncommercial 2.5 Australia License.
- Dick Culbert. Available online: https://en.wikipedia.org/wiki/Calandrinia (accessed on 10 August 2023). This Image Is Licensed under the Creative Commons Attribution 2.0 Generic License.
- Brian Freeman. Available online: https://davesgarden.com/guides/pf/go/248887/#b (accessed on 10 August 2023). No License Found.
- Photography by C. Hortin & J. Hooper. Image Used with the Permission of the Western Australian Herbarium, Department of Biodiversity, Conservation and Attractions. Available online: https://florabase.dbca.wa.gov.au/help/copyright. (accessed on 10 August 2023).
- Dinkum at Wikimedia Commons. Available online: https://commons.wikimedia.org/wiki/File:Portulacaria_afra_02.JPG (accessed on 10 August 2023). This Image Is Licensed under the Creative Commons CC0 License.
- Frank Vincentz. Available online: https://commons.wikimedia.org/wiki/File:Sedum_album_03_ies.jpg (accessed on 10 August 2023).
- Sajetpa at Malayalam Wikipedia. Available online: https://en.wikipedia.org/wiki/Coleus_amboinicus#/media/File:Leaf_-pani_koorkka.JPG (accessed on 10 August 2023). No License Found.
- Winter, K.; Garcia, M.; Virgo, A.; Smith, J.A.C. Low-level CAM photosynthesis in a succulent-leaved member of the Urticaceae, Pilea peperomioides. Funct. Plant Biol. 2021, 48, 683–690. [Google Scholar] [CrossRef]
- Bräutigam, A.; Schlüter, U.; Eisenhut, M.; Gowik, U. On the evolutionary origin of CAM photosynthesis. Plant Physiol. 2017, 174, 473–477. [Google Scholar] [CrossRef]
- Yang, X.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.C.; De Paoli, H.C.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef]
- Boxall, S.F.; Kadu, N.; Dever, L.V.; Knerová, J.; Waller, J.L.; Gould, P.J.D.; Hartwell, J. Kalanchoë PPC1 is essential for Crassulacean acid metabolism and the regulation of core circadian clock and guard cell signaling genes. Plant Cell 2020, 32, 1136–1160. [Google Scholar] [CrossRef]
- Yang, X.; Hu, R.; Yin, H.; Jenkins, J.; Shu, S.; Tang, H.; Liu, D.; Weighill, D.A.; Yim, W.C.; Ha, J.; et al. The Kalanchoë genome provides insights into convergent evolution and building blocks of crassulacean acid metabolism. Nat. Commun. 2017, 8, 1899. [Google Scholar] [CrossRef]
- Shameer, S.; Baghalian, K.; Cheung, C.Y.M.; Ratcliffe, R.G.; Sweetlove, L.J. Computational analysis of the productivity potential of CAM. Nat. Plants 2018, 4, 165–171. [Google Scholar] [CrossRef]
- Tay, I.Y.Y.; Odang, K.B.; Cheung, C.Y.M. Metabolic modeling of the C3-CAM continuum revealed the establishment of a starch/sugar-malate cycle in CAM evolution. Front. Plant Sci. 2021, 11, 573197. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.D.; Lee, S.; Choi, W.G.; Yim, W.C.; Cushman, J.C. Laying the foundation for Crassulacean acid metabolism (CAM) biodesign: Expression of the C4 metabolism cycle genes of CAM in Arabidopsis. Front. Plant Sci. 2019, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, N.; Wang, Y.; Zhou, R.; Sun, P.; Lin, H.; Chen, W.; Yu, T.; Liu, Z.; Wang, Z.; et al. High-quality ice plant reference genome analysis provides insights into genome evolution and allows exploration of genes involved in the transition from C3 to CAM pathways. Plant Biotechnol. J. 2022, 20, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; VanBuren, R.; Wai, C.M.; Tang, H.; Schatz, M.C.; Bowers, J.E.; Lyons, E.; Wang, M.-L.; Chen, J.; Biggers, E.; et al. The pineapple genome and the evolution of CAM photosynthesis. Nat. Genet. 2015, 47, 1435–1442. [Google Scholar] [CrossRef]
- Taybi, T.; Cushman, J.C.; Borland, A.M. Leaf carbohydrates influence transcriptional and post-transcriptional regulation of nocturnal carboxylation and starch degradation in the facultative CAM plant, Mesembryanthemum crystallinum. J. Plant Physiol. 2017, 218, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Szechyńska-Hebda, M.; Kornaś, A.; Miszalski, Z. Withdrawal from functional Crassulacean acid metabolism (CAM) is accompanied by changes in both gene expression and activity of antioxidative enzymes. J. Plant Physiol. 2018, 229, 151–157. [Google Scholar] [CrossRef]
- Guan, Q.; Kong, W.; Zhu, D.; Zhu, W.; Dufresne, C.; Tian, J.; Chen, S. Comparative proteomics of Mesembryanthemum crystallinum guard cells and mesophyll cells in transition from C3 to CAM. J. Proteom. 2021, 231, 104019. [Google Scholar] [CrossRef]
- Wakamatsu, A.; Mori, I.C.; Matsuura, T.; Taniwaki, Y.; Ishii, R.; Yoshida, R. Possible roles for phytohormones in controlling the stomatal behavior of Mesembryanthemum crystallinum during the salt-induced transition from C3 to Crassulacean acid metabolism. J. Plant Physiol. 2021, 262, 153448. [Google Scholar] [CrossRef]
- Winter, K. Properties of phosphoenolpyruvate carboxylase in rapidly prepared, desalted leaf extracts of the Crassulacean acid metabolism plant Mesembryanthemum crystallinum L. Planta 1982, 154, 298–308. [Google Scholar] [CrossRef]
- Häusler, R.E.; Baur, B.; Scharte, J.; Teichmann, T.; Eicks, M.; Fischer, K.L.; Flügge, U.I.; Schubert, S.; Weber, A.; Fischer, K. Plastidic metabolite transporters and their physiological functions in the inducible crassulacean acid metabolism plant Mesembryanthemum crystallinum. Plant J. 2000, 24, 285–296. [Google Scholar] [CrossRef]
- Neuhaus, H.E.; Holtum, J.A.; Latzko, E. Transport of phosphoenolpyruvate by chloroplasts from Mesembryanthemum crystallinum L. exhibiting Crassulacean acid metabolism. Plant Physiol. 1988, 87, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Cushman, J.C.; Meyer, G.; Michalowski, C.B.; Schmitt, J.M.; Bohnert, H.J. Salt stress leads to differential expression of two isogenes of phosphoenolpyruvate carboxylase during Crassulacean acid metabolism induction in the common ice plant. Plant Cell 1989, 1, 715–725. [Google Scholar] [CrossRef]
- Vaasen, A.; Begerow, D.; Hampp, R. Phosphoenolpyruvate carboxylase genes in C3, Crassulacean acid metabolism (CAM) and C3/CAM intermediate species of the genus Clusia: Rapid reversible C3/CAM switches are based on the C3 housekeeping gene. Plant Cell Environ. 2006, 29, 2113–2123. [Google Scholar] [CrossRef] [PubMed]
- Cushman, J.C.; Tillett, R.L.; Wood, J.A.; Branco, J.M.; Schlauch, K.A. Large-scale mRNA expression profiling in the common ice plant, Mesembryanthemum crystallinum, performing C3 photosynthesis and Crassulacean acid metabolism (CAM). JXB 2008, 59, 1875–1894. [Google Scholar] [CrossRef] [PubMed]
- Kore-Eda, S.; Noake, C.; Ohishi, M.; Ohnishi, J.I.; Cushman, J.C. Transcriptional profiles of organellar metabolite transporters during induction of Crassulacean acid metabolism in Mesembryanthemum crystallinum. Funct. Plant Biol. 2005, 32, 451–466. [Google Scholar] [CrossRef]
- Abraham, P.E.; Yin, H.; Borland, A.M.; Weighill, D.; Lim, S.D.; De Paoli, H.C.; Engle, N.; Jones, P.C.; Agh, R.; Weston, D.J.; et al. Transcript, protein and metabolite temporal dynamics in the CAM plant Agave. Nat. Plants 2016, 2, 16178. [Google Scholar] [CrossRef] [PubMed]
- Brilhaus, D.; Bräutigam, A.; Mettler-Altmann, T.; Winter, K.; Weber, A.P. Reversible burst of transcriptional changes during induction of Crassulacean acid metabolism in Talinum triangulare. Plant Physiol. 2016, 170, 102–122. [Google Scholar] [CrossRef]
- Maleckova, E.; Brilhaus, D.; Wrobel, T.J.; Weber, A.P.M. Transcript and metabolite changes during the early phase of abscisic acid-mediated induction of Crassulacean acid metabolism in Talinum triangulare. JXB 2019, 70, 6581–6596. [Google Scholar] [CrossRef]
- Gonçalves, A.Z.; Mercier, H. Transcriptomic and biochemical analysis reveal integrative pathways between carbon and nitrogen metabolism in Guzmania monostachia (Bromeliaceae) under drought. Front. Plant Sci. 2021, 12, 715289. [Google Scholar] [CrossRef]
- Habibi, G. Comparison of CAM expression, photochemistry and antioxidant responses in Sedum album and Portulaca oleracea under combined stress. Physiol. Plant 2020, 170, 550–568. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, B.; Zhu, D.; Dufresne, D.; Jiang, T.; Chen, S. Proteomics of homeobox7 enhanced salt tolerance in Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 6390. [Google Scholar] [CrossRef] [PubMed]
- Na, J.K.; Metzger, J.D. Chimeric promoter mediates guard cell-specific gene expression in tobacco under water deficit. Biotechnol. Lett. 2014, 36, 1893–1899. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, F.; Simeoni, F.; Francia, P.; Cominelli, E.; Conti, L.; Riboni, M.; Simoni, L.; Martin, C.R.; Tonelli, C.; Galbiati, M. The Arabidopsis thaliana MYB60 promoter provides a tool for the spatio-temporal control of gene expression in stomatal guard cells. JXB 2013, 64, 3361–3371. [Google Scholar] [CrossRef]
- Winter, K.; Smith, J.A.C. CAM photosynthesis: The acid test. New Phytol 2022, 233, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.D.; Yim, W.C.; Liu, D.; Hu, R.; Yang, X.; Cushman, J.C. A Vitis vinifera basic helix–loop–helix transcription factor enhances plant cell size, vegetative biomass and reproductive yield. Plant Biotechnol. J. 2018, 16, 1595–1615. [Google Scholar] [CrossRef]
- Kebeish, R.; Niessen, M.; Oksaksin, M.; Blume, C.; Peterhaensel, C. Constitutive and dark-induced expression of Solanum tuberosum phosphoenolpyruvate carboxylase enhances stomatal opening and photosynthetic performance of Arabidopsis thaliana. Biotechnol. Bioeng. 2012, 109, 536–544. [Google Scholar] [CrossRef]
- Liu, D.; Hu, R.; Zhang, J.; Guo, H.B.; Cheng, H.; Li, L.; Borland, A.M.; Qin, H.; Chen, J.G.; Muchero, W.; et al. Overexpression of an Agave phosphoenolpyruvate carboxylase improves plant growth and stress tolerance. Cells 2021, 10, 582. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, B.; Cheng, Z.; Zhu, D.; Jiang, T.; Chen, S. Overexpression of McHB7 transcription factor from Mesembryanthemum crystallinum improves plant salt tolerance. Int. J. Mol. Sci. 2022, 23, 7879. [Google Scholar] [CrossRef]
- Chiang, C.-P.; Yim, W.C.; Sun, Y.-H.; Ohnishi, M.; Mimura, T.; Cushman, J.C.; Yen, H.E. Identification of Ice Plant (Mesembryanthemum crystallinum L.) MicroRNAs Using RNA-Seq and Their Putative Roles in High Salinity Responses in Seedlings. Front. Plant Sci. 2016, 7, 1143. [Google Scholar] [CrossRef]
- Chu, C.; Dai, Z.; Ku, M.S.; Edwards, G.E. Induction of Crassulacean acid metabolism in the facultative halophyte Mesembryanthemum crystallinum by abscisic acid. Plant Physiol. 1990, 93, 1253–1260. [Google Scholar] [CrossRef]
- Cushman, J.C.; Borland, A.M. Induction of Crassulacean acid metabolism by water limitation. Plant Cell Environ. 2002, 25, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Heyduk, K.; Hwang, M.; Albert, V.; Silvera, K.; Lan, T.; Farr, K.; Chang, T.H.; Chan, M.T.; Winter, K.; Leebens-Mack, J. Altered gene regulatory networks are associated with the transition from C3 to Crassulacean acid metabolism in Erycina (oncidiinae: Orchidaceae). Front. Plant Sci. 2019, 9, 2000. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.G.; Davey, P.A.; Hofmann, T.A.; Borland, A.; Hartwell, J.; Lawson, T. Stomatal responses to light, CO2, and mesophyll tissue in Vicia faba and Kalanchoë fedtschenkoi. Front. Plant Sci. 2021, 12, 740534. [Google Scholar] [CrossRef]
- Cai, J.; Liu, X.; Vanneste, K.; Proost, S.; Tsai, W.-C.; Liu, K.-W.; Chen, L.-J.; He, Y.; Xu, Q.; Bian, C.; et al. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2014, 47, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Copetti, D.; Búrquez, A.; Bustamante, E.; Charboneau, J.L.M.; Childs, K.L.; Eguiarte, L.E.; Lee, S.; Liu, T.L.; McMahon, M.M.; Whiteman, N.K.; et al. Extensive gene tree discordance and hemiplasy shaped the genomes of North American columnar cacti. Proc. Natl. Acad. Sci. USA 2017, 114, 12003–12008. [Google Scholar] [CrossRef]
- Xin, H.; Wang, Y.; Li, Q.; Wan, T.; Hou, Y.; Liu, Y.; Gichuki, D.K.; Zhou, H.; Zhu, Z.; Xu, C.; et al. A genome for Cissus illustrates features underlying the evolutionary success in dry savannas. Hortic. Res. 2022, 9, uhac208. [Google Scholar] [CrossRef] [PubMed]
- Sato, R.; Kondo, Y.; Agarie, S. The first released available genome of the common ice plant (Mesembryanthemum crystallinum L.) extended the research region on salt tolerance, C3-CAM photosynthetic conversion, and halophism [version 1; peer review: 1 approved with reservations]. F1000Research 2023, 12, 448. [Google Scholar] [CrossRef]
- Liu, D.; Chen, M.; Mendoza, B.; Cheng, H.; Hu, R.; Li, L.; Trinh, C.T.; Tuskan, G.A.; Yang, X. CRISPR/Cas9-mediated targeted mutagenesis for functional genomics research of Crassulacean acid metabolism plants. JXB 2019, 70, 6621–6629. [Google Scholar] [CrossRef]
- Walker, R.P.; Chen, Z.H.; Acheson, R.M.; Leegood, R.C. Effects of phosphorylation on phosphoenolpyruvate carboxykinase from the C4 plant Guinea grass. Plant Physiol. 2002, 128, 165–172. [Google Scholar] [CrossRef]
- Walker, R.P.; Leegood, R.C. Purification, and phosphorylation in vivo and in vitro, of phosphoenolpyruvate carboxykinase from cucumber cotyledons. FEBS Lett. 1995, 362, 70–74. [Google Scholar] [CrossRef]
- Clark, N.M.; Elmore, J.M.; Walley, J.W. To the proteome and beyond: Advances in single-cell omics profiling for plant systems. Plant Physiol. 2022, 188, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Katam, R.; Lin, C.; Grant, K.; Katam, C.S.; Chen, S. Advances in plant metabolomics and its applications in stress and single-cell biology. Int. J. Mol. Sci. 2022, 23, 6985. [Google Scholar] [CrossRef] [PubMed]
- Specht, H.; Emmott, E.; Petelski, A.A.; Huffman, R.G.; Perlman, D.H.; Serra, M.; Kharchenko, P.; Koller, A.; Slavov, N. Single-cell proteomic and transcriptomic analysis of macrophage heterogeneity using SCoPE2. Genome Biol. 2021, 22, 50. [Google Scholar] [CrossRef] [PubMed]
- Nungesser, D.; Kluge, M.; Tolle, H.; Oppelt, W. A dynamic computer model of the metabolic and regulatory processes in Crassulacean acid metabolism. Planta 1984, 162, 204–214. [Google Scholar] [CrossRef]
- Burgos, A.; Miranda, E.; Vilaprinyo, E.; Meza-Canales, I.D.; Alves, R. CAM Models: Lessons and Implications for CAM Evolution. Front. Plant Sci. 2022, 13, 893095. [Google Scholar] [CrossRef]
- Töpfer, N.; Braam, T.; Shameer, S.; Ratcliffe, R.G.; Sweetlove, L.J. Alternative Crassulacean acid metabolism modes provide environment-specific water-saving benefits in a leaf metabolic model. Plant Cell 2020, 32, 3689–3705. [Google Scholar] [CrossRef]
- Libault, M.; Chen, S. Editorial: Plant single cell type systems biology. Front. Plant Sci. 2016, 7, 35. [Google Scholar] [CrossRef]
- Misra, B.B.; Assmann, S.M.; Chen, S. Plant single-cell and single-cell-type metabolomics. Trends Plant Sci. 2014, 19, 637–646. [Google Scholar] [CrossRef]
- Su, L.; Xu, C.; Zeng, S.; Su, L.; Joshi, T.; Stacey, G.; Xu, D. Large-scale integrative analysis of soybean transcriptome using an unsupervised autoencoder model. Front. Plant Sci. 2022, 13, 831204. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Focus | Key Findings | Reference |
|---|---|---|
| Identified C3-CAM transition period and temporal physiological changes |
| [20] |
| Transcriptomics of guard cells during the C3-CAM transition |
| [19] |
| ||
| Nocturnal carboxylation is coordinated with starch degradation by the products of these pathways, such as carbohydrates |
| [64] |
| Functional CAM withdrawal in the de-salted plants |
| [65] |
| Comparative proteomic changes in guard cells and mesophyll cells during the C3-CAM transition |
| [66] |
| Phytohormones in the stomatal behavior during the C3-CAM transition |
| [67] |
| Genome sequencing, transcriptomics, and comparative genomics of leaves |
| [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, B.; Chen, S. Defining Mechanisms of C3 to CAM Photosynthesis Transition toward Enhancing Crop Stress Resilience. Int. J. Mol. Sci. 2023, 24, 13072. https://doi.org/10.3390/ijms241713072
Tan B, Chen S. Defining Mechanisms of C3 to CAM Photosynthesis Transition toward Enhancing Crop Stress Resilience. International Journal of Molecular Sciences. 2023; 24(17):13072. https://doi.org/10.3390/ijms241713072
Chicago/Turabian StyleTan, Bowen, and Sixue Chen. 2023. "Defining Mechanisms of C3 to CAM Photosynthesis Transition toward Enhancing Crop Stress Resilience" International Journal of Molecular Sciences 24, no. 17: 13072. https://doi.org/10.3390/ijms241713072
APA StyleTan, B., & Chen, S. (2023). Defining Mechanisms of C3 to CAM Photosynthesis Transition toward Enhancing Crop Stress Resilience. International Journal of Molecular Sciences, 24(17), 13072. https://doi.org/10.3390/ijms241713072