1. Introduction
Uveal melanoma is a rare malignant intraocular tumor occurring mainly in adults, representing approximately 5% of the total incidence of adult melanomas. Most UM arises from melanocytes within the choroid and, in contrast to cutaneous melanomas, is characterized by a low burden of genetic alterations. Despite originating from the same embryonic origin, uveal and cutaneous melanomas differ in their molecular characteristics and how they metastasize [
1,
2,
3]. Primarily, skin-localized melanoma metastasizes in various organs, while UM is characterized by a high frequency of metastases in the liver, which are particularly resistant to treatment [
4]. Primary UM can be effectively treated with enucleation, followed by stereotactic radiotherapy. While a few patients have detectable metastases at the time of diagnosis, about half of all patients will develop liver metastases within 5 years [
5].
Up to now, no systemic therapy for patients with metastatic uveal melanoma has received regulatory approval. These patients have a dismal prognosis because of the limited efficacy of the available treatments [
6]. The current chemotherapeutic approaches for UM metastases tested in clinical trials yield very low response rates [
7,
8]. A limited therapeutic effect was reported in an immunological clinical study using the adoptive transfer of tumor-infiltrating lymphocytes [
9]. These therapeutic failures highlight the need for more targeted therapy that would specifically attack or prevent UM metastases [
10].
Tumor cells with the potential to metastasize and be released into circulation retain their original homing characteristics and possess the ability to re-home to their site of origin [
11]. Tumor-cell-derived extracellular vesicles (EVs) mimic the contents of their parent cell, suggesting that their therapeutically modified EVs will be biodistributed preferentially to tumors [
12]. Consequently, systemically injected tumor-derived EVs will home to their original tumor tissues. Tsering with colleagues revealed that the cargo of UM-EVs play a role in premetastatic niche formation and metastasis in the liver. During metastatic progression, EVs from UM tumor cells are involved in micro-metastasis formation not detectable using current techniques. Furthermore, accumulations of progression factors promote the metastatic process [
13].
In our previous work, we utilized the tumor-specific tropism of mesenchymal stem/stromal cells (MSCs) to develop a targeted prodrug gene therapy [
14]. This therapy is mediated by MSCs transduced with a retrovirus, yielding cells engineered to express the suicide gene
yCD::UPRT. Our retrovirus vector had the
yCD::UPRT gene linked by an internal ribosome entry site sequence (IRES) with the neo gene, which allowed us to produce a homogenous cell population of
yCD::UPRT-transduced cells by G418 antibiotic selection. Ultimately, the suicide
yCD::UPRT gene is integrated into the cell DNA with a strong retrovirus promoter, the DNA provirus is expressed, and yeast cytosine deaminase catalyzed by
UPRT converts the nontoxic prodrug 5-fluorocytosine to the effective chemotherapeutic drug 5-fluorouracil [
15]. Later, we found
yCD::UPRT transduced MSCs excreted EVs possessing mRNA of the suicide gene in their cargo [
16]. Furthermore, we reported that a replicative-deficient retrovirus possessing the
yCD::UPRT gene is able to infect several different human tumors, leading to the transduction of tumor cells with the retrovirus vector integrated into the cell DNA of the recipient tumor cells. Similar to the
yCD::UPRT- MSCs, the tumor cells secrete EVs possessing the
yCD::UPRT gene mRNA in their cargo [
17]. Uveal melanoma, like all tumor cells, release EVs with diverse biological functions, delivering them to recipient cells. Composition and biogenesis analyses of UM EVs indicate that they induce a prometastatic microenvironment through macrophage migration inhibitory factor [
18]. In addition, UM EVs have been shown to prime the liver for metastatic growth by activating stellate cells in the premetastatic niche, resulting in pro-fibrogenic properties [
19]. Based on these results, as well as on our previous work, we hypothesized that the
yCD::UPRT suicide gene-EVs secreted by primary human UM cells can attack micro-metastases and/or destroy the formation of a premetastatic niche.
In the present study, we report the transduction of several primary uveal melanoma cells and UM-associated stromal cells with the yCD::UPRT gene. We characterized the secreted EVs in regard to their biologic activity, tumor targeting ability, and capability to effectively kill tumor cells of different types in vitro.
3. Discussion
Extracellular vesicles, especially sEVs secreted in large numbers from cancer cells, play an important role in metastases formation. Furthermore, an increasing amount of evidence indicates that sEVs secreted from cancer cells play a key role in the establishment of the premetastatic niche [
21,
22]. Generally, sEVs secreted from tumor cells are organ-tropic [
23,
24]. Moreover, sEVs mimic the roles performed by their cells of origin and can reach the appropriate target cells at distant sites and organs. Thanks to these features, tumor-derived sEVs are attractive targets for suicide gene modification [
25]. We have previously shown in many experiments that enrichment of MSCs or tumor cells with the
yCD::UPRT suicide gene via retrovirus construct leads to its DNA integration and expression. Consequently, the
yCD::UPRT-transduced cells will transfer the mRNA into excreted sEVs [
16,
17]. Obviously, the yeast origin of the expressed gene in human cells is not compatible with cell homeostasis and is, therefore, excreted. All isolated primary uveal melanoma cells transduced with
yCD::UPRT-retrovirus behaved similarly. They secreted EVs possessing mRNA information for the
yCD::UPRT-suicide gene in their cargo. All tested primary
yCD::UPRT-gene-transduced uveal melanoma cells (
yCD::UPRT-UM) proliferated at a similar rate as their parental naïve cells. Further analysis of CM from
yCD::UPRT-UM using size-exclusion chromatography provided evidence that sEVs in CM are responsible for their tumor cell killing activity. The presence of
yCD::UPRT mRNA in these sEVs was probed with RT-PCR. The standard characterization of EVs, such as through using Western blot analysis and TEM, confirmed the EVs’ characteristic of nanoparticles, produced both in the transduced and naïve UMs. The concentration measurement with the NanoSight fluctuated around 4–10
10 per ml of CM in dependence of the time–duration of the CM harvest.
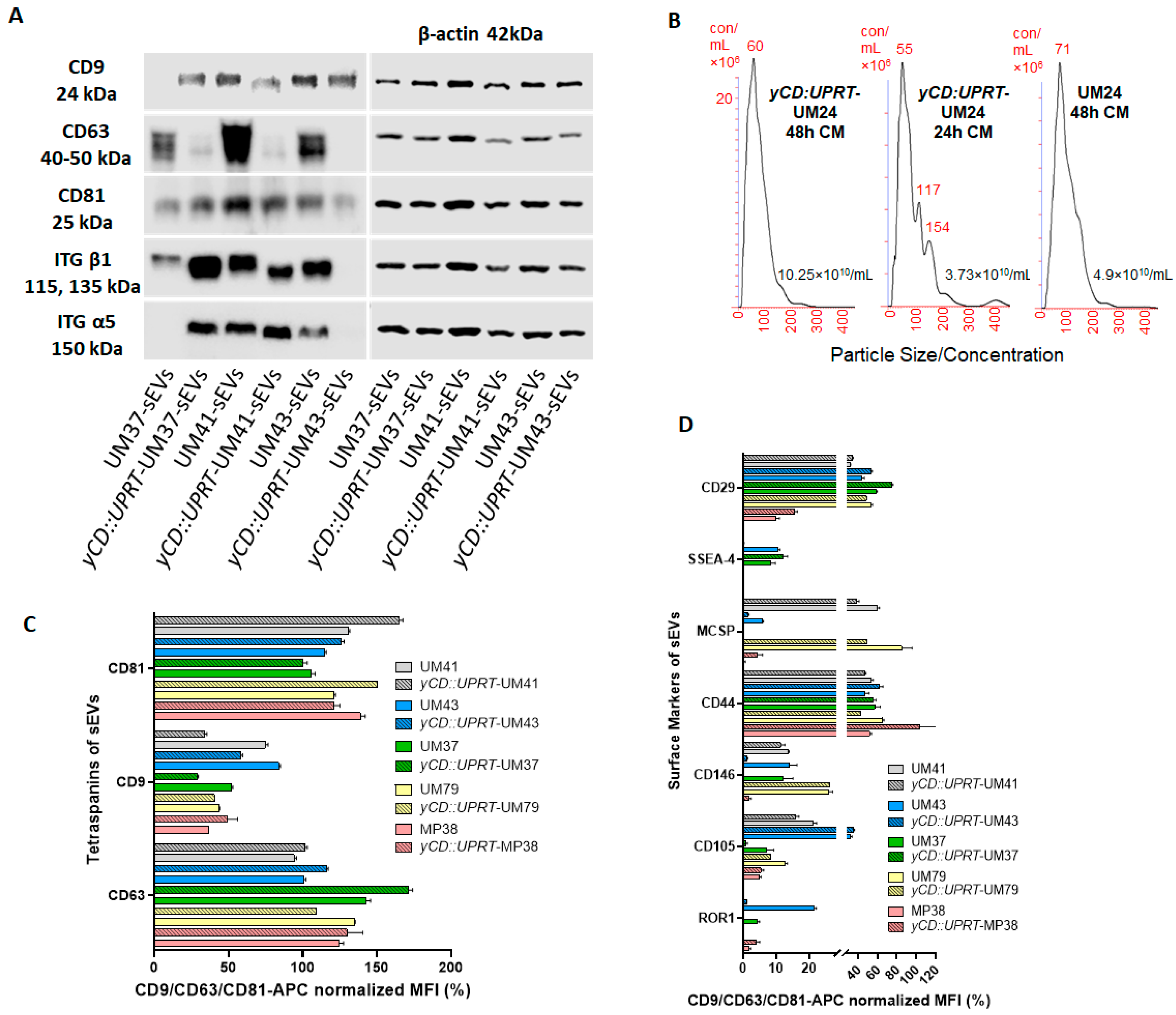
The data obtained from the bead-based multiplex flow cytometry assay confirmed the assumption that sEVs excreted from
yCD::UPRT-UMs are the sEVs from naïve UMs enriched with mRNA of the
yCD::UPRT-gene. The sEVs were both derived through the endosomal sorting complexes required for transport (ESCRT)-dependent sorting and exhibited integrins. The percentage value variations in the median fluorescence intensity normalized to the mean CD9/CD63/CD81 of the sEVs from individual primary UMs and sEVs excreted from the corresponding
yCD::UPRT-UM-gene-transduced cells may reflect their individual ability to proliferate, migrate, and communicate with the environment dissemination and metastasis [
26]. For the cell surface adhesion receptor CD44, we found it to be highly expressed at the same level in the sEVs from the naïve and
yCD::UPRT-gene-transduced UMs. The CD44 receptor is highly expressed in many cancers and regulates metastasis via recruitment of CD44 to the cell surface. The same conclusion can be drawn from the observed expression data of the integrin beta-1, known as CD29, on sEVs secreted from naïve and
yCD::UPRT-gene-transduced UMs. Integrin beta-1 activation has been shown to regulate the transition from cell dormancy to metastasis. Interestingly enough, the sEVs of MP38, both naïve and
yCD::UPRT gene transduced, differ from the other UMs through the low expression of CD29 but again at the same level. The expression of inactive tyrosine-protein kinase transmembrane receptor ROR1 on sEVs of UMs was found to be extremely low compared to other markers, with the exception of sEVs from naive UM43. Whether the observed difference is related to the immortalized character of the UM cell line, which is likely, remains to be elucidated. This may be related to the heterogeneity of the primary UMs introduced to the growth in vitro. The
yCD::UPRT-UM-43 cells, being selected for homogenous
yCD::UPRT-gene-transduced cells morphologically, resemble stromal fibroblastoid cells. Generally, all of these cytometric data show that the transduction of primary UMs with
yCD::UPRT retroviral construct is not leading to the dramatic changes in the immunologically detected sEVs markers, despite the random integration of retrovirus into the cell DNA, which could cause changes in the transcription of neighboring genes near to the integrated provirus and general heterogeneity of tumor-derived extracellular vesicles.
The host range of the yCD::UPRT-UM-sEVs in the tumor cell growth inhibition in the presence of prodrug was found not to be restricted to UMs only. Furthermore, the specificity of the sEVs from the individual primary UM and from them derived yCD::UPRT-UM-sEVs were found not to show differences between patients from whom the cells were obtained. Generally, highly proliferating cells that are metabolically more active excrete higher numbers of EVs. However, some UM cell lines, primary UM cells, and yCD::UPRT-gene-transduced cells proliferate very slowly. In addition, the proliferation of primary UMs drops with an increasing number of passages.
The proliferating activity of individual UMs is in good agreement with the histopathologic findings of cyclin D1 expression in individual uveal melanoma tumor tissue. This could be the reason why some biomarkers in UM37 and yCD::UPRT-UM43 sEVs tested negative in the WB analysis. Cyclin D1 was found to be highly overexpressed in UM49, UM55, and UM41 cells, while in UM37 and UM43 cells, the cyclin D1 expression was at the limit of detection. The heterogeneity of the sEVs populations is probably responsible for the broad host range of the yCD::UPRT-UM-sEVs tested in vitro on the human and mouse tumor cell lines of various types.
We hypothesized that sEVs excreted from UM modified with suicide gene messages may be targeted to liver metastases and/or prevent the formation of premetastatic niches. Human hepatocyte carcinoma Hep G2 was found to be sensitive to
yCD::UPRT-UM-sEVs present in CM in our in vitro experiments. We speculate that
yCD::UPRT-gene-modified sEVs secreted from primary UMs being targeted to liver may act as a preventive vaccine against liver UM metastases. The
yCD::UPRT-UM-EVs together with the prodrug systematically injected could prevent metastases formation by the inhibition of premetastatic niche creation. This assumption is supported by the evidence that
yCD::UPRT-MSC-EVs together with the prodrug was not toxic in rats, we found, when injected systematically to treat intracerebral rat glioblastoma with curative outcome [
27]. Several recent reports support this notion. For example, genetically modified tumors releasing sEVs were found to possess antitumor potential [
28]. Recently, engineered tumor exosomes were found to act as a dendritic cell (DC)-primed vaccine, which boosted antitumor immunity in breast cancer [
29]. Furthermore, MSC exosomes modified with the
yCD::UPRT gene used for the experimental treatment of glioblastoma-carrying rats prevented the recurrence of glioblastoma in cured rats [
27]. The prevention of UM metastases formation via the inhibition of premetastatic niche creation by
yCD::UPRT-UM-sEVs seems to be possible. In vivo experiments with primary UM using patient-derived xenografts in immunocompromised mice will be the next translational approach.
EVs secreted from cells, as a natural cell product, have many advantages over conventional nanocarriers [
30]. They have low immunogenicity, good biocompatibility, can penetrate the natural blood–brain barrier, and have the capacity for gene delivery. They can be loaded with anticancer drugs; consequently, EVs are absorbed in much greater proportions than other nanocarrier vesicles, such as liposomes or polystyrene NPs [
31]. Cell-secreted sEVs have a distinct cell tropism and homing selectivity that can be used to target them into certain tissues or organs [
32]. For instance, sEVs derived from dental pulp MSCs with gemcitabine as a cargo have an inhibitory effect on the growth of pancreatic carcinoma cell lines in vitro [
33].
4. Materials and Methods
4.1. Establishment of Cell Cultures from Human Uveal Melanoma
Uveal melanoma cells were obtained from eye globes with uveal melanoma (n = 59) enucleated at the Department of Ophthalmology (Medical Faculty of the Comenius University, Bratislava, Slovakia) between 2019 and 2020. All of the enucleated eye globes with uveal melanoma were histopathologically verified. Donors were informed about the nature of the study, and they provided their written informed consent. The study was approved by the Ruzinov Hospital Bratislava Ethics Committee on 12 December 2018 (number: EK/250/2018). Each UM tumor was processed within two hours after enucleation, disintegrated by mincing, and followed by enzymatic digestion of the tumor fragments with a 0.25% trypsin-EDTA solution (Sigma Aldrich, St. Louis, MO, USA) at 37 °C for 15 min. The cells were cultured as either monolayers or multicellular spheroids at 37 °C in a humidified atmosphere of 95% air and 5% CO2 with MEMα medium (Gibco, Thermofisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS) (Biosera, Nuaille, France). Serial passages were performed by trypsinization (0.25% trypsin-EDTA solution) of subconfluent monolayers. The cells were seeded in a tissue culture dish (Corning Life Sciences, Corning, NY, USA) at a density of 5 × 104 cells/cm2, and the culture medium was replaced every 2–3 days.
4.2. Isolation of Uveal Melanoma-Associated Fibroblasts (UMAFs)
In order to isolate the stromal cells, uveal melanoma tissue fragments were adhered to the plastic tissue culture dish and surrounded with a complete culture medium of low-glucose DMEM (1 g/L) supplemented with 5% human platelet lysate (HPL). The next day, the fragments were overlaid with medium and further cultivated at 37 °C in a humidified atmosphere with 5% CO2. Two–three days later, the stromal cells—UMAFs—started to migrate from the adhered UM tissue explants.
4.3. Tumor Cell Lines’ Maintenance
The lines used in this study were the human melanoma cell line A375, mouse melanoma B16-F0, and human prostate carcinoma cell line PC3. All were maintained in high-glucose DMEM with 5% FBS. The cells were authenticated using STR profiling. The uveal melanoma cell line MP38 and human hepatocyte carcinoma Hep G2 cell line were purchased from ATCC (American Type Culture Collection, Manassas, VA, USA) and maintained in high-glucose DMEM supplemented with 10% FBS. All cell lines were routinely tested for mycoplasma infection using PCR, and all experiments were performed using mycoplasma-free cells.
4.4. Preparation of Replication-Deficient yCD::UPRT Suicide Gene Retrovirus
The procedure for the preparation of cells producing
yCD::UPRT-retrovirus was performed as previously described [
15]. Briefly, we used a bicistronic retroviral vector containing the fusion gene yeast cytosine deaminase uracilphosphoribosyl transferase (
yCD::UPRT) separated by
IRES sequences from the
neo gene allowing for the selection of a pure population of transfected cells with Geneticin (G418) antibiotics. The cells that produced the virus with mixed eco- and ampho- glycoprotein virus envelopes were generated with four ping-pong rounds of infections of helper cell lines GP+E-86 and GP+envAm12.
4.5. Transduction of UM Cells with yCD::UPRT-Retrovirus
In order to obtain a transduced population of primary UM, the semi-confluent cell culture was infected with yCD::UPRT-retrovirus supplemented with protamine sulphate (5 μg/mL). A homogenous population of yCD::UPRT-gene-transduced UM cells was obtained by cultivation in selection media with G418. We used a pretested concentration of G418 for selection in every primary UM cell culture using a range over 0.4 to 1.0 mg/mL.
4.6. Cell Growth Assessment using the IncuCyte Live Cell Monitoring System
The growth of the UM cell cultures and various cancerous cell lines under the designed treatment was monitored using the IncuCyte ZOOM Kinetic Imaging system (Essen BioScience, Royston, UK). Cell viability data obtained from the IncuCyte ZOOM system were compared with data obtained with the standard MTT test—CellTiter 96 Aqueous One Solution Cell Proliferation Assay (Promega, Madison, WI, USA).
4.7. Conditioned Media and Extracellular Vesicles Isolation
Conditioned medium (CM) was used as the source of the sEVs in the experiments where the biological activity of the yCD::UPRT-UM-sEVs was evaluated and for the particle size estimation. The isolation of the sEVs was performed for the downstream analyses, such as Western blotting and PCR. The CM was harvested from the semi-confluent UM cell cultures, washed three times with phosphate buffered saline (PBS) to remove any debris, and then cultured in FBS-free MEMα for 24 or 48 h. The 24 h CM was collected every second day, and the 48 h CM was collected every third day, alternating with the growth culture fluid exchange. The usual number of cells was 2 × 106 cells in a 100 mm Petri dish. The harvested CM was centrifuged at 800× g for 5 min to remove any possible residual cells and cell debris and then filtered through a 0.22 μm syringe filter to remove microvesicles. The sEVs were isolated from the CM using size-exclusion chromatography, Amicon® Ultra-15 ultrafiltration, TFF-Easy—tangential flow filtration EV concentrator 1 (HansaBioMed Life Sciences Ltd., Tallinn, Estonia), or ExoQuick TC (System Biosciences, Mountain View, CA, USA). Isolation of RNA from the sEVs was performed with a SeraMir Exosome RNA Column Purification Kit (System Biosciences, Mountain View, CA, USA). The sEVs for the Western blot analysis were concentrated 20–40 times using an Amicon® Ultra-15 Centrifugal Filter Unit 100 kDA MWCO (Merck KGaA, Darmstadt, Germany) and precipitated with ExoQuick TC (System Biosciences, Mountain View, CA, USA).
4.8. Separation of the sEVs from Conditioned Media with Size-Exclusion Chromatography
The CM from the yCD::UPRT-transduced uveal melanoma cells was separated in a PD-10 column (GE Healthcare, 105 mm × 15 mm) filled with Sephacryl S-500 (GE Healthcare, Chicago, IL, USA), pre-equilibrated with PBS, and stored at 4 °C in 20% ethanol. Before use, the column was equilibrated to room temperature and washed with PBS (two void volumes of the column) using a peristaltic pump P-1 (Pharmacia Lkb, Uppsala, Sweden). Subsequently, 1 mL of filtered CM (0.22 µm PES filter) was applied in the column, and 22 fractions (0.5 mL each) were collected. All fractions were tested for tumor cell proliferation (in the absence of 5-FC) and growth inhibition (in the presence of 5-FC) on A375 cells monitored with an IncuCyte S3 Live-Cell Analysis System.
4.9. Evaluation of sEVs’ Concentration and Size Distribution
The assessment of the particle size, quantity, and distribution was performed using a NanoSight NS500 instrument (Malvern Instruments Ltd., Great Malvern, UK) equipped with a 405 nm laser and sCMOS trigger camera. Data were analyzed using Nanoparticle Tracking Analysis (NTA) 2.3 software.
4.10. Cell DNA Isolation and PCR Amplification
DNA extraction from cell pellets was performed using a QIAmp DNA Mini kit (Qiagen, Venlo, The Netherlands) following the manufacturer’s instructions. A NanoDrop® ND-1000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) was used to control for extracted DNA quantity and quality. Standard PCR using 50 ng of genomic DNA was then performed with the same sets of primers and chemistry as qPCR employing a Bio-Rad T100 thermal cycler (Bio-Rad, Hercules, CA, USA). The products were visualized by agarose gel electrophoresis (2%) using a 50 bp DNA ladder.
4.11. Cell RNA Isolation and RT-PCR
Total RNA was extracted from pellets of yCD::UPRT-UM cells and corresponding control cells with RNeasy Mini Kit (Qiagen, Venlo, The Netherlands). A NanoDrop® ND-1000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) was used to assess the RNA quality and quantity. Reverse transcription was performed using a RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, Loughborough, UK). Real-time PCR (RT-PCR) was performed with identical primers while following the same protocol used for exoRNA. The GAPDH gene was used as an amplification control.
4.12. Exosome RNA Isolation
The sEVs from the cultivation media were isolated using the Complete SeraMir Exosome RNA Amplification kit (System Biosciences, Mountain View, CA, USA) containing Exoquick-TC (System Biosciences, Mountain View, CA, USA), using the protocol recommended by the supplier. Briefly, CM was centrifuged at 3000× g for 15 min to remove cells and cell debris. Then, 5 mL of supernatant was mixed with 1 mL of Exoquick solution, mixed, and then incubated overnight at 4 °C. Subsequently, the tubes were centrifuged at 16,000× g for 2 min, and the sEVs pellet was reconstituted in 350 μL of lysis buffer. After vortexing, followed by incubation for 5 min at room temperature to allow for complete lysis, 200 μL of 100% ethanol was added, and 600 μL of the mixture was filtered through the spin column at 16,000× g for 1 min. Two wash steps were then performed with 400 μL of wash buffer, followed by centrifugation at 16,000× g for 1 min, which was then followed by the elution of total RNA into 30 μL of elution buffer. The quality of the eluted exoRNA was then evaluated using the Agilent RNA 6000 Nano Kit (Agilent Technologies, Santa Clara, CA, USA).
4.13. cDNA Synthesis from Total RNA
Five microliters of the eluate was then used for reverse transcription using the Complete SeraMir Exosome RNA Amplification kit (System Biosciences, Mountain View, CA, USA). For the poly A reaction, exoRNA eluted from the spin column (5 µL), 5× polyA buffer (2 µL), 25 mM MnCl2 (1 µL), 5 mM ATP (1.5 µL), and polyA polymerase (0.5 µL) were incubated for 30 min in 37 °C. Then, 0.5 µL of SeraMir Adaptor Oligo was added, and the mix was incubated for 5 min at 60 °C. After subsequent incubation at room temperature for 2 min, the RT reaction was performed in a 20 µL volume and consisted of polyA exoRNA from the previous step (10 µL), 5× RT master mix (4 µL), 5’ SeraMir Switch Oligo (1 µL), reverse transcriptase (1 µL), and RNA-se-free water (4 µL). Incubation in a BioRad T100 thermal cycler (Bio-Rad, Hercules, CA, USA) at 42 °C for 30 min was followed by 95 °C for 10 min to terminate the reaction.
4.14. Real-Time PCR Profiling Using exo-cDNA
Real-time PCR (RT-PCR) was then performed using 20 µL of the cDNA generated in the previous step, using 10 μL of 2× GoTaq
® qPCR Master Mix (Promega, Madison, WI, USA), 1 μL (0.5 μM) forward primer, 1 μL (0.5 μM) reverse primer, 7 μL ultrapure DNase/RNase-free water, and 1 μL of cDNA. Primer sequences were as published previously [
17].
yCD::UPRT forward: 5’-ATGGACATTGCCTATGAGGA-3’;
yCD::UPRT reverse: 5’-TTCTCCAGGGTGCTGATCTC-3’ (product size 167 bp);
GAPDH forward: 5’-GAAGGTGAAGGTCGGAGTC-3’;
GAPDH reverse: 5’-GAAGATGGTGATGGGATTTC-3’ (product size 226 bp).
All samples were analyzed in triplicate on a Bio-Rad CFX96 real-time PCR detection system (Bio-Rad, Hercules, CA, USA). The following cycling conditions were applied: 95 °C for 10 min, 40 cycles at 95 °C for 15 s, 60 °C for 30 s, and 72 °C for 30 s. Given the missing yCD::UPRT transcripts in negative controls, the presence of specific transcripts was visualized using agarose gel electrophoresis.
4.15. Western Blot Analysis
The sEVs were lysed in RIPA buffer with a protease inhibitor cocktail (Complete ULTRA, Roche, Switzerland), a phosphatase (PhosSTOP, Roche, Switzerland), and sonicated. The proteins (30 μg per well) were loaded onto 8% or 10% SDS polyacrylamide gels and subjected to electrophoresis. Separated proteins were transferred to a 0.45 µm Nitrocellulose Transfer Membrane (Thermo Fisher Scientific) using the Mini Trans-Blot Cell Module (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Membranes were incubated overnight at 4 °C with the appropriate primary antibodies (CD9 #10626D, CD63 #10628D, and CD81 #10630D purchased from Invitrogen, Integrin α5/CD49e #610633 from BD Biosciences, and Integrin β1 Antibody #4706S from CST). The applied dilutions are shown in
Supplementary Materials Table S1. A primary monoclonal antibody against β-actin (# A1978 Sigma-Aldrich, St-Louis, MO, USA) served as a loading control. The specific binding of the antibodies was detected with the appropriate peroxidase-conjugated Alexa Fluor 680 secondary antibody and visualized using an LI-COR
® instrument (LI-COR
®, Lincoln, NE, USA).
4.16. Bead-Based Multiplex Flow Cytometry Assay
The sEVs isolated from the primary UMs and sEVs from the corresponding yCD::UPRT-UMs were subjected to a surface-marker characterization using a flow cytometry bead-based multiplex analysis (MACSPlex Exosome Kit, human, Miltenyi Biotec. 130-108-813). The samples were processed according to the manufacturer’s protocol. Briefly, sEVs were isolated via CM concentration using a TFF-Easy EV concentrator 1 (TFF). With TFF, a filter unit separates the CM into retentate containing sEVs and flow-through permeate. One-third of the 50-fold concentrated CM sEVs were mixed with the manufacturer’s buffer to a final volume of 120 µL, followed by the addition of 15 µL of MACSPlex Exosome Capture Beads. After, the samples were incubated overnight at room temperature, protected from the light, and on rotation—450 rpm. Then, 500 µL of the MACSPlex buffer was added, and the samples were centrifuged at 3000× g for 5 min. The same volume of supernatant was discarded, and 15 µL of the detection antibody cocktail—5 µL of each MACSPlex exosome detection reagent: CD9, CD63, and CD81—was added to each sample. The samples were incubated for 1 hour at room temperature, protected from the light, and on rotation—450 rpm. Later, 500 µL of buffer was added to each sample, centrifuged at 3000× g for 5 min, followed by discarding of the same volume of supernatant, and washed again with 500 μL of buffer. The samples were incubated for 15 min at room temperature on rotation, protected from light, and then centrifuged for 5 min at 3000× g. Subsequently, 500 µL of the supernatant was removed, and approximately 150 µL of each sample was re-suspended and used for the analysis. The flow cytometry analysis was performed using the MACSQuant® Analyzer 10 (Miltenyi Biotec), and results were processed with MACSQuant Analyzer 10 software (Miltenyi Biotec). The 39 single bead populations were gated to determine the APC signal intensity on each bead population, and the median fluorescence intensity (MFI) for each capture bead was measured. For each population, the background was corrected by subtracting the respective MFI values from the non-EVs controls that were treated exactly like the EV samples and, furthermore, normalized to the mean of the APC-MFI detected for the CD9, CD63, and CD81 Exosome Capture Beads (mean CD9, CD63, and CD81 = 100%). Only positive markers are shown in the graphics.
4.17. Statistical Analysis
Unless noted otherwise, all experiments were repeated at least three times to enable statistical analysis, and all results were similar among replicates. Data from the MACSPlex analysis were statistically evaluated using the one-way ANOVA nonparametric test (Friedman test). For multiple comparisons, Dunn’s test was used. The statistical analysis was carried out using GraphPad Prism 9 software (GraphPadPrism Software, San Diego, CA, USA).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}