
Heterodimerization of Chemoreceptors TAS1R3 and mGlu2 in Human Blood Leukocytes

Abstract
:1. Introduction
2. Results
2.1. Gene Transcripts of All GRMs and TAS1Rs Are Expressed in Human PMNs and T Cells
2.2. Immunocytochemistry Revealed Co-Expression of mGlu2 and TAS1R3 in Isolated PMNs
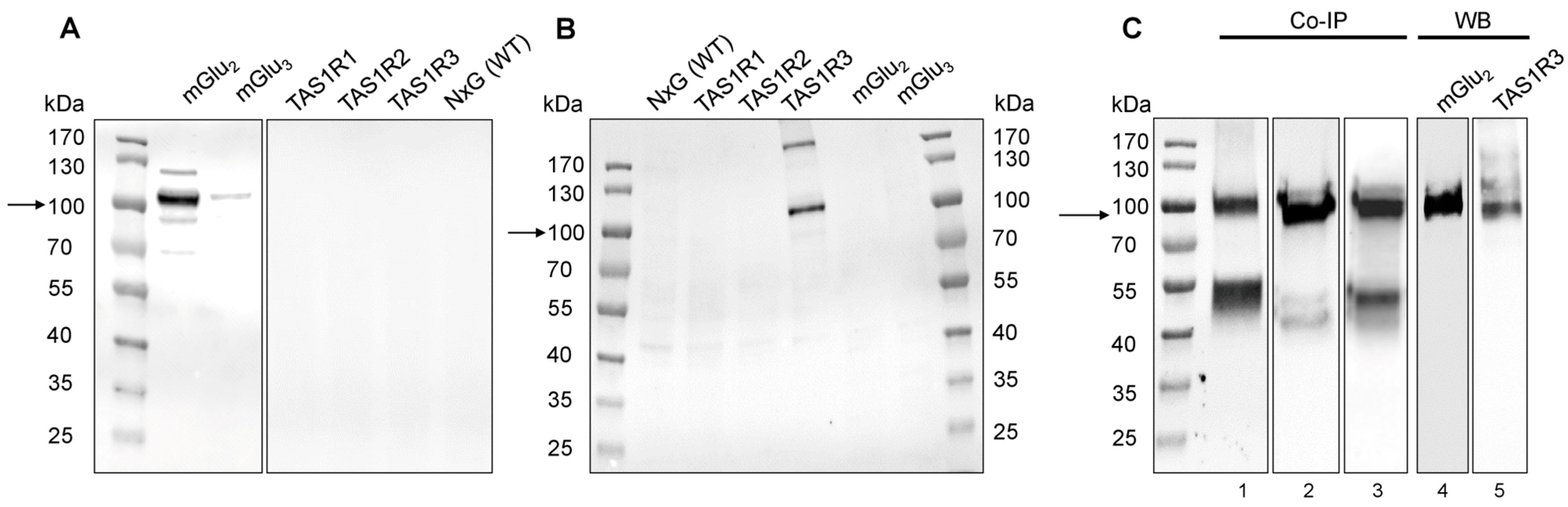
2.3. Western Blot and co-IP Revealed Heterodimerization of mGlu2 and TAS1R3 in Isolated PMNs and T Cells
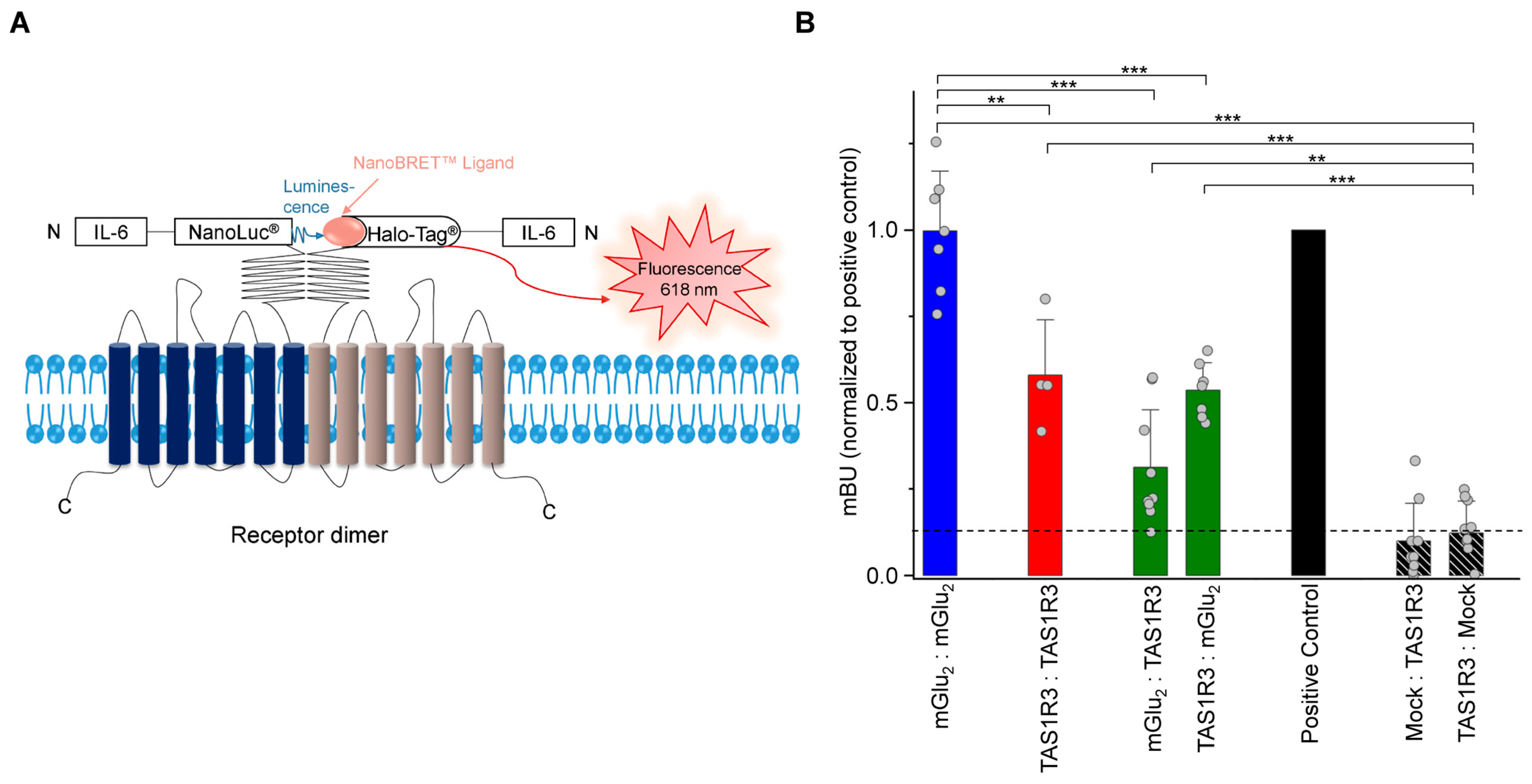
2.4. Recombinant mGlu2 and TAS1R3 Form Heteromeric Complexes in HEK-293 Cells
2.5. Functional Analysis of Recombinant mGlu2 and TAS1R3 Revealed a Heterodimer-Related Gain-of-Function in Response to MSG in HEK-293 Cells
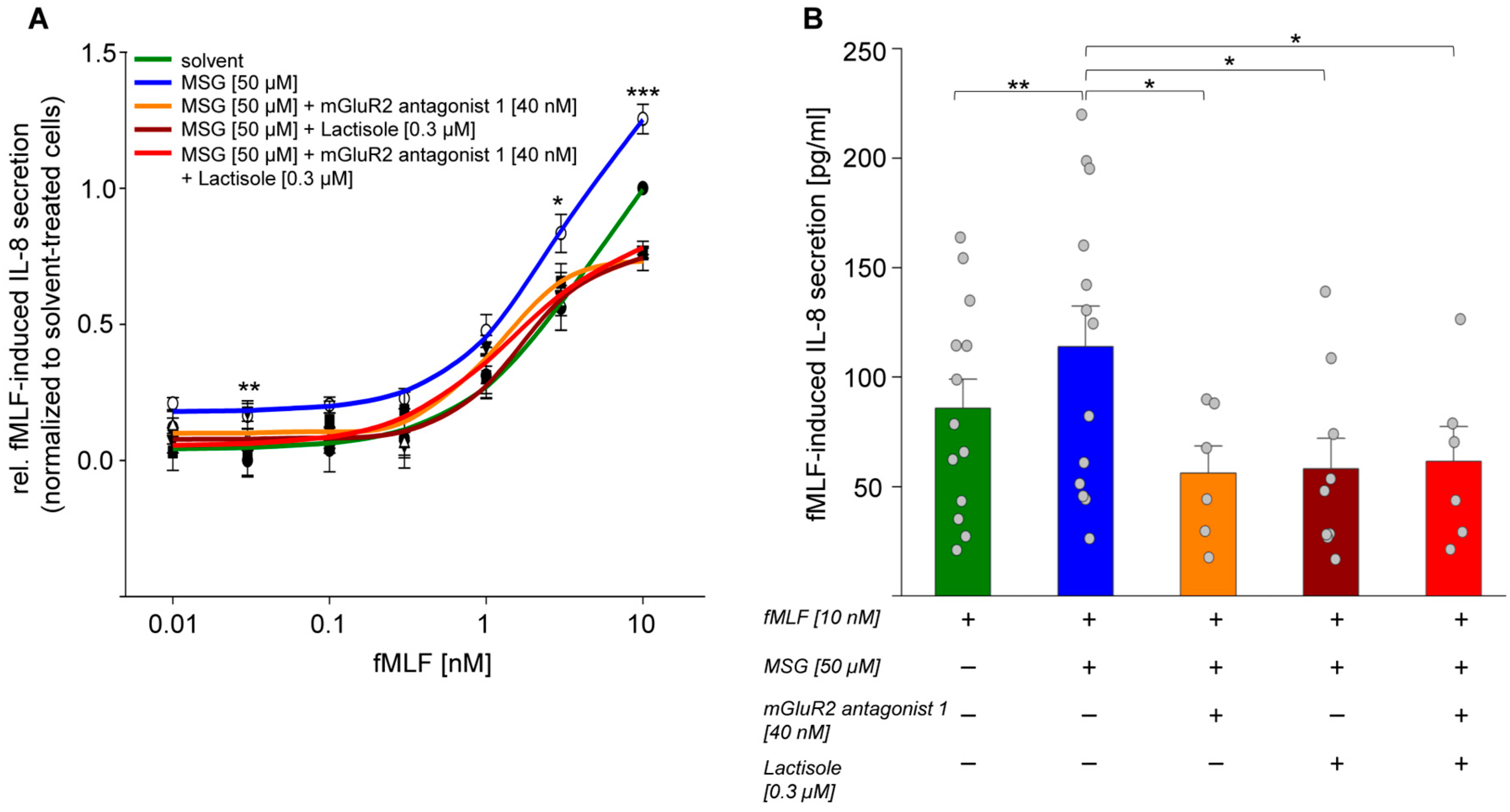
2.6. MSG via mGlu2/TAS1R3 Facilitated an fMLF-Induced IL-8 Secretion in Isolated PMNs In Vitro
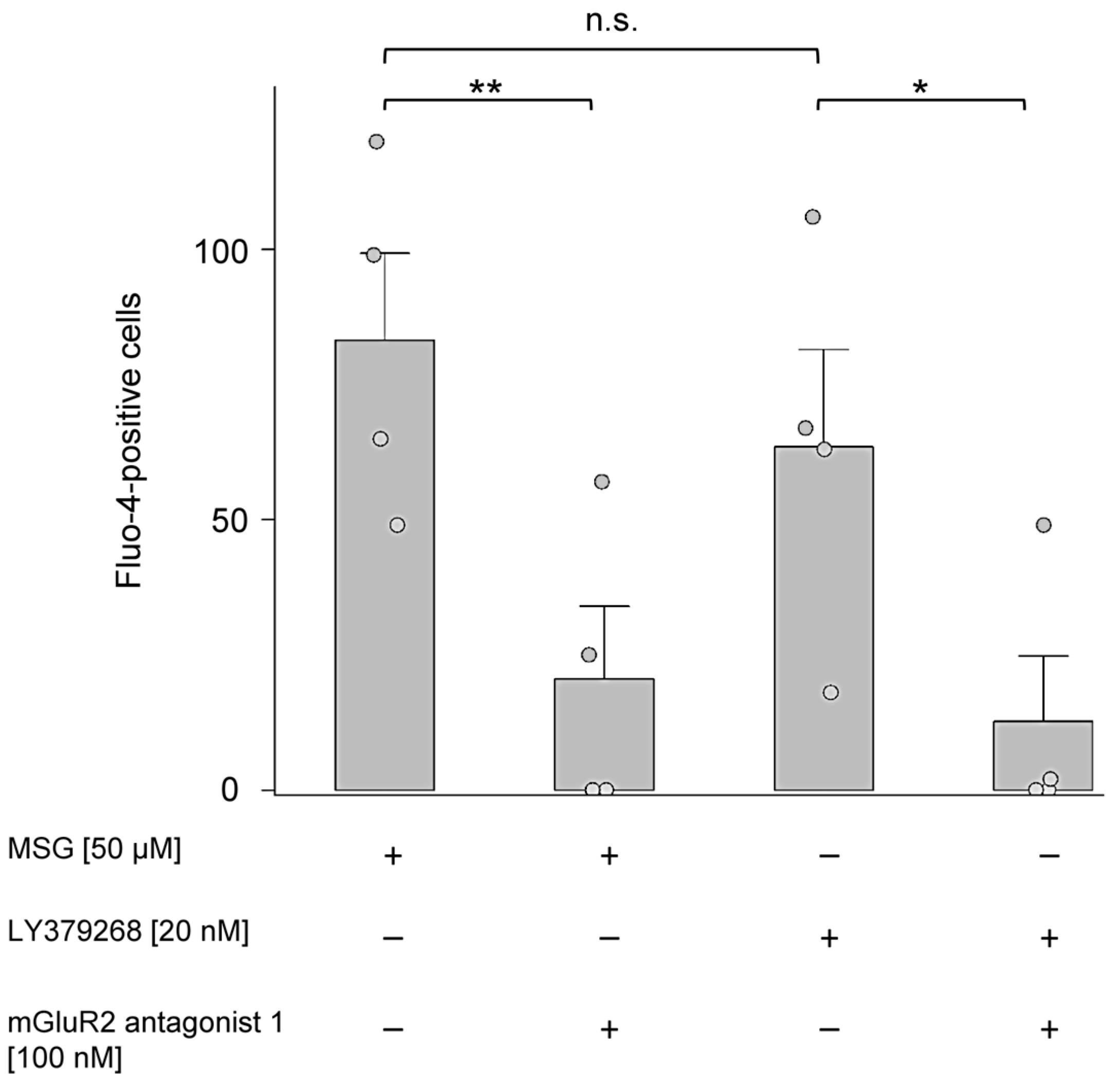
2.7. MSG and LY379268 Increased Intracellular Ca2+ in Isolated PMNs
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Human Blood Cell Purification
4.3. RNA Isolation and cDNA Synthesis
4.4. RT-PCR and qPCR
4.5. ddPCR
4.6. Immunocytochemistry
4.7. Bioluminescence Resonance Energy Transfer (BRET) Assay
4.8. Western Blot and Co-IP
4.9. Molecular Cloning
4.10. Cell Culture and Transient DNA Transfection
4.11. cAMP Luminescence Assay
4.12. Data Analysis of cAMP Luminescence Assay
4.13. ELISA
4.14. Intracellular Ca2+ Fluorescence Measured Using Laser-Guided Flow Cytometry
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milligan, G.; Ward, R.J.; Marsango, S. GPCR homo-oligomerization. Curr. Opin. Cell Biol. 2019, 57, 40–47. [Google Scholar] [CrossRef]
- Ferre, S.; Casado, V.; Devi, L.A.; Filizola, M.; Jockers, R.; Lohse, M.J.; Milligan, G.; Pin, J.P.; Guitart, X. G protein-coupled receptor oligomerization revisited: Functional and pharmacological perspectives. Pharmacol. Rev. 2014, 66, 413–434. [Google Scholar] [CrossRef]
- Lohse, M.J. Dimerization in GPCR mobility and signaling. Curr. Opin. Pharmacol. 2010, 10, 53–58. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Gurevich, E.V. GPCR monomers and oligomers: It takes all kinds. Trends Neurosci. 2008, 31, 74–81. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suarez-Boomgaard, D.; Rivera, A.; et al. The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Brito, I.; Narvaez, M.; Savelli, D.; Shumilov, K.; Di Palma, M.; Sartini, S.; Skieterska, K.; Van Craenenbroeck, K.; Valladolid-Acebes, I.; Zaldivar-Oro, R.; et al. Searching the GPCR Heterodimer Network (GPCR-hetnet) Database for Information to Deduce the Receptor–Receptor Interface and Its Role in the Integration of Receptor Heterodimer Functions. In Receptor-Receptor Interactions in the Central Nervous System; Humana Press: New York, NY, USA, 2018; pp. 283–298. [Google Scholar] [CrossRef]
- Zhao, G.Q.; Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Erlenbach, I.; Ryba, N.J.; Zuker, C.S. The receptors for mammalian sweet and umami taste. Cell 2003, 115, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.; Zuker, C.S. An amino-acid taste receptor. Nature 2002, 416, 199–202. [Google Scholar] [CrossRef]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [PubMed]
- Romano, C.; Yang, W.L.; O’Malley, K.L. Metabotropic glutamate receptor 5 is a disulfide-linked dimer. J. Biol. Chem. 1996, 271, 28612–28616. [Google Scholar] [CrossRef]
- Pandya, N.J.; Klaassen, R.V.; van der Schors, R.C.; Slotman, J.A.; Houtsmuller, A.; Smit, A.B.; Li, K.W. Group 1 metabotropic glutamate receptors 1 and 5 form a protein complex in mouse hippocampus and cortex. Proteomics 2016, 16, 2698–2705. [Google Scholar] [CrossRef]
- Doumazane, E.; Scholler, P.; Zwier, J.M.; Trinquet, E.; Rondard, P.; Pin, J.P. A new approach to analyze cell surface protein complexes reveals specific heterodimeric metabotropic glutamate receptors. FASEB J. 2011, 25, 66–77. [Google Scholar] [CrossRef]
- Levitz, J.; Habrian, C.; Bharill, S.; Fu, Z.; Vafabakhsh, R.; Isacoff, E.Y. Mechanism of Assembly and Cooperativity of Homomeric and Heteromeric Metabotropic Glutamate Receptors. Neuron 2016, 92, 143–159. [Google Scholar] [CrossRef]
- Kammermeier, P.J. Functional and pharmacological characteristics of metabotropic glutamate receptors 2/4 heterodimers. Mol. Pharmacol. 2012, 82, 438–447. [Google Scholar] [CrossRef]
- Moreno Delgado, D.; Moller, T.C.; Ster, J.; Giraldo, J.; Maurel, D.; Rovira, X.; Scholler, P.; Zwier, J.M.; Perroy, J.; Durroux, T.; et al. Pharmacological evidence for a metabotropic glutamate receptor heterodimer in neuronal cells. eLife 2017, 6, e25233. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Noetzel, M.J.; Johnson, K.A.; Zamorano, R.; Jalan-Sakrikar, N.; Gregory, K.J.; Conn, P.J.; Niswender, C.M. Selective actions of novel allosteric modulators reveal functional heteromers of metabotropic glutamate receptors in the CNS. J. Neurosci. 2014, 34, 79–94. [Google Scholar] [CrossRef]
- Lattin, J.; Zidar, D.A.; Schroder, K.; Kellie, S.; Hume, D.A.; Sweet, M.J. G-protein-coupled receptor expression, function, and signaling in macrophages. J. Leukoc. Biol. 2007, 82, 16–32. [Google Scholar] [CrossRef]
- Kehrl, J.H. G-protein-coupled receptor signaling, RGS proteins, and lymphocyte function. Crit. Rev. Immunol. 2004, 24, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, C.; Lind, S.; Mårtensson, J.; Björkman, L.; Wu, Y.; Sundqvist, M.; Forsman, H. G protein coupled pattern recognition receptors expressed in neutrophils: Recognition, activation/modulation, signaling and receptor regulated functions. Immunol. Rev. 2022, 314, 69–92. [Google Scholar] [CrossRef]
- Futosi, K.; Fodor, S.; Mocsai, A. Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int. Immunopharmacol. 2013, 17, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly, Y.M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PLoS ONE 2011, 6, e21205. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Lira, S.A.; Furtado, G.C. The biology of chemokines and their receptors. Immunol. Res. 2012, 54, 111–120. [Google Scholar] [CrossRef]
- Sharma, M. Chemokines and their receptors: Orchestrating a fine balance between health and disease. Crit. Rev. Biotechnol. 2010, 30, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.A. Role of the complement in experimental sepsis. J. Leukoc. Biol. 2008, 83, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ye, D. Molecular biology for formyl peptide receptors in human diseases. J. Mol. Med. 2013, 91, 781–789. [Google Scholar] [CrossRef]
- Migeotte, I.; Communi, D.; Parmentier, M. Formyl peptide receptors: A promiscuous subfamily of G protein-coupled receptors controlling immune responses. Cytokine Growth Factor Rev. 2006, 17, 501–519. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wang, L.; Guo, J.; Sun, D.; Wang, Y.; Liu, W.; Xu, H.E.; Zhang, C. Molecular recognition of formylpeptides and diverse agonists by the formylpeptide receptors FPR1 and FPR2. Nat. Commun. 2022, 13, 1054. [Google Scholar] [CrossRef]
- Malki, A.; Fiedler, J.; Fricke, K.; Ballweg, I.; Pfaffl, M.W.; Krautwurst, D. Class I odorant receptors, TAS1R and TAS2R taste receptors, are markers for subpopulations of circulating leukocytes. J. Leukoc. Biol. 2015, 97, 533–545. [Google Scholar] [CrossRef]
- Marcinek, P.; Geithe, C.; Krautwurst, D. Chemosensory G Protein-Coupled Receptors (GPCR) in Blood Leukocytes. In Taste and Smell; Krautwurst, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 151–173. [Google Scholar] [CrossRef]
- Skurk, T.; Kraemer, T.; Marcinek, P.; Malki, A.; Lang, R.; Krautwurst, T.; Hofmann, T.; Krautwurst, D. Sweetener system intervention shifted neutrophils from homeostasis to priming. Nutrients 2023, 15, 1260. [Google Scholar] [CrossRef]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef]
- Lee, R.J.; Cohen, N.A. Taste receptors in innate immunity. Cell. Mol. Life Sci. CMLS 2015, 72, 217–236. [Google Scholar] [CrossRef]
- Howitt, M.R.; Cao, Y.G.; Gologorsky, M.B.; Li, J.A.; Haber, A.L.; Biton, M.; Lang, J.; Michaud, M.; Regev, A.; Garrett, W.S. The Taste Receptor TAS1R3 Regulates Small Intestinal Tuft Cell Homeostasis. Immunohorizons 2020, 4, 23–32. [Google Scholar] [CrossRef]
- Qin, Y.; Palayyan, S.R.; Zheng, X.; Tian, S.; Margolskee, R.F.; Sukumaran, S.K. Type II taste cells participate in mucosal immune surveillance. PLoS Biol. 2023, 21, e3001647. [Google Scholar] [CrossRef]
- Sukumaran, S.K.; Lewandowski, B.C.; Qin, Y.; Kotha, R.; Bachmanov, A.A.; Margolskee, R.F. Whole transcriptome profiling of taste bud cells. Sci. Rep. 2017, 7, 7595. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Lima, M.M.; Procopio, J.; Pithon-Curi, T.C.; Doi, S.Q.; Bazotte, R.B.; Curi, R. Glutamine and glutamate as vital metabolites. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas E Biol. 2003, 36, 153–163. [Google Scholar] [CrossRef]
- Xue, H.; Field, C.J. New role of glutamate as an immunoregulator via glutamate receptors and transporters. Front. Biosci. 2011, 3, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Conn, P.J.; Pin, J.P. Pharmacology and functions of metabotropic glutamate receptors. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 205–237. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.J.; Goudet, C. International Union of Basic and Clinical Pharmacology. CXI. Pharmacology, Signaling, and Physiology of Metabotropic Glutamate Receptors. Pharmacol. Rev. 2021, 73, 521–569. [Google Scholar] [CrossRef]
- Chaudhari, N.; Pereira, E.; Roper, S.D. Taste receptors for umami: The case for multiple receptors. Am. J. Clin. Nutr. 2009, 90, 738s–742s. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Chattopadhyay, D. Glutamate is the chemotaxis-inducing factor in placental extracts. Amino Acids 2009, 37, 359–366. [Google Scholar] [CrossRef]
- Gupta, R.; Palchaudhuri, S.; Chattopadhyay, D. Glutamate induces neutrophil cell migration by activating class I metabotropic glutamate receptors. Amino Acids 2013, 44, 757–767. [Google Scholar] [CrossRef]
- Miglio, G.; Varsaldi, F.; Dianzani, C.; Fantozzi, R.; Lombardi, G. Stimulation of group I metabotropic glutamate receptors evokes calcium signals and c-jun and c-fos gene expression in human T cells. Biochem. Pharmacol. 2005, 70, 189–199. [Google Scholar] [CrossRef]
- Chiocchetti, A.; Miglio, G.; Mesturini, R.; Varsaldi, F.; Mocellin, M.; Orilieri, E.; Dianzani, C.; Fantozzi, R.; Dianzani, U.; Lombardi, G. Group I mGlu receptor stimulation inhibits activation-induced cell death of human T lymphocytes. Br. J. Pharmacol. 2006, 148, 760–768. [Google Scholar] [CrossRef]
- Fallarino, F.; Volpi, C.; Fazio, F.; Notartomaso, S.; Vacca, C.; Busceti, C.; Bicciato, S.; Battaglia, G.; Bruno, V.; Puccetti, P.; et al. Metabotropic glutamate receptor-4 modulates adaptive immunity and restrains neuroinflammation. Nat. Med. 2010, 16, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.; Ciruela, F.; Casado, V.; Mallol, J.; Gallart, T.; Lluis, C.; Franco, R. Group I metabotropic glutamate receptors mediate a dual role of glutamate in T cell activation. J. Biol. Chem. 2004, 279, 33352–33358. [Google Scholar] [CrossRef]
- Pacheco, R.; Gallart, T.; Lluis, C.; Franco, R. Role of glutamate on T-cell mediated immunity. J. Neuroimmunol. 2007, 185, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Poulopoulou, C.; Davaki, P.; Koliaraki, V.; Kolovou, D.; Markakis, I.; Vassilopoulos, D. Reduced expression of metabotropic glutamate receptor 2mRNA in T cells of ALS patients. Ann. Neurol. 2005, 58, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Pin, J.P.; Galvez, T.; Prezeau, L. Evolution, structure, and activation mechanism of family 3/C G-protein-coupled receptors. Pharmacol. Ther. 2003, 98, 325–354. [Google Scholar] [CrossRef]
- Zhang, F.; Klebansky, B.; Fine, R.M.; Xu, H.; Pronin, A.; Liu, H.; Tachdjian, C.; Li, X. Molecular mechanism for the umami taste synergism. Proc. Natl. Acad. Sci. USA 2008, 105, 20930–20934. [Google Scholar] [CrossRef]
- Mafi, A.; Kim, S.K.; Chou, K.C.; Guthrie, B.; Goddard, W.A., 3rd. Predicted Structure of Fully Activated Tas1R3/1R3’ Homodimer Bound to G Protein and Natural Sugars: Structural Insights into G Protein Activation by a Class C Sweet Taste Homodimer with Natural Sugars. J. Am. Chem. Soc. 2021, 143, 16824–16838. [Google Scholar] [CrossRef]
- Brauner-Osborne, H.; Wellendorph, P.; Jensen, A.A. Structure, pharmacology and therapeutic prospects of family C G-protein coupled receptors. Curr. Drug Targets 2007, 8, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Boss, V.; Nutt, K.M.; Conn, P.J. L-cysteine sulfinic acid as an endogenous agonist of a novel metabotropic receptor coupled to stimulation of phospholipase D activity. Mol. Pharmacol. 1994, 45, 1177–1182. [Google Scholar] [PubMed]
- Meldrum, B.S. Glutamate as a Neurotransmitter in the Brain: Review of Physiology and Pathology. J. Nutr. 2000, 130, 1007S–1015S. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Zhu, W.L.; Ning, Y.L.; Li, P.; Zhao, Y.; Yang, N.; Chen, X.; Jiang, Y.L.; Yang, W.Q.; Jiang, D.P.; et al. Dramatic increases in blood glutamate concentrations are closely related to traumatic brain injury-induced acute lung injury. Sci. Rep. 2017, 7, 5380. [Google Scholar] [CrossRef]
- Suzuki, S.; Koshimizu, H.; Adachi, N.; Matsuoka, H.; Fushimi, S.; Ono, J.; Ohta, K.I.; Miki, T. Functional interaction between BDNF and mGluR II in vitro: BDNF down-regulated mGluR II gene expression and an mGluR II agonist enhanced BDNF-induced BDNF gene expression in rat cerebral cortical neurons. Peptides 2017, 89, 42–49. [Google Scholar] [CrossRef]
- Jørgensen, C.V.; Klein, A.B.; El-Sayed, M.; Knudsen, G.M.; Mikkelsen, J.D. Metabotropic glutamate receptor 2 and corticotrophin-releasing factor receptor-1 gene expression is differently regulated by BDNF in rat primary cortical neurons. Synapse 2013, 67, 794–800. [Google Scholar] [CrossRef]
- Avila, J.R.; Lee, J.S.; Torii, K.U. Co-Immunoprecipitation of Membrane-Bound Receptors. Arab. Book 2015, 13, e0180. [Google Scholar] [CrossRef]
- Hall, R.A. Co-immunoprecipitation as a strategy to evaluate receptor-receptor or receptor-protein interactions. In G Protein-Coupled Receptor–Protein Interactions; Dowd, B.F., George, S.R., Eds.; Wiley: Hoboken, NJ, USA, 2005; pp. 165–178. [Google Scholar]
- Hamprecht, B. Structural, electrophysiological, biochemical, and pharmacological properties of neuroblastoma-glioma cell hybrids in cell culture. Int. Rev. Cytol. 1977, 49, 99–170. [Google Scholar] [CrossRef]
- Machleidt, T.; Woodroofe, C.C.; Schwinn, M.K.; Mendez, J.; Robers, M.B.; Zimmerman, K.; Otto, P.; Daniels, D.L.; Kirkland, T.A.; Wood, K.V. NanoBRET—A Novel BRET Platform for the Analysis of Protein-Protein Interactions. ACS Chem. Biol. 2015, 10, 1797–1804. [Google Scholar] [CrossRef]
- Binkowski, B.; Fan, F.; Wood, K. Engineered luciferases for molecular sensing in living cells. Curr. Opin. Biotechnol. 2009, 20, 14–18. [Google Scholar] [CrossRef]
- Noe, F.; Frey, T.; Fiedler, J.; Geithe, C.; Nowak, B.; Krautwurst, D. IL-6–HaloTag® enables live-cell plasma membrane staining, flow cytometry, functional expression, and de-orphaning of recombinant odorant receptors. J. Biol. Methods 2017, 4, e81. [Google Scholar] [CrossRef] [PubMed]
- Noe, F.; Geithe, C.; Fiedler, J.; Krautwurst, D. A bi-functional IL-6-HaloTag® as a tool to measure the cell-surface expression of recombinant odorant receptors and to facilitate their activity quantification. J. Biol. Methods 2017, 4, e82. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Masu, M.; Ishii, T.; Shigemoto, R.; Nakanishi, S. A family of metabotropic glutamate receptors. Neuron 1992, 8, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Insel, P.A.; Ostrom, R.S. Forskolin as a tool for examining adenylyl cyclase expression, regulation, and G protein signaling. Cell. Mol. Neurobiol. 2003, 23, 305–314. [Google Scholar] [CrossRef]
- Seamon, K.B.; Daly, J.W. Forskolin: A unique diterpene activator of cyclic AMP-generating systems. J. Cycl. Nucleotide Res. 1981, 7, 201–224. [Google Scholar]
- Cubillos, S.; Norgauer, J.; Lehmann, K. Toxins-useful biochemical tools for leukocyte research. Toxins 2010, 2, 428–452. [Google Scholar] [CrossRef]
- Katada, T. The inhibitory G protein G(i) identified as pertussis toxin-catalyzed ADP-ribosylation. Biol. Pharm. Bull. 2012, 35, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.L. Subunit structure and enzymic activity of pertussis toxin. Microbiol. Sci. 1988, 5, 285–287. [Google Scholar]
- Sundd, P.; Pospieszalska, M.K.; Ley, K. Neutrophil rolling at high shear: Flattening, catch bond behavior, tethers and slings. Mol. Immunol. 2013, 55, 59–69. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Acher, F.; Battaglia, G.; Bräuner-Osborne, H.; Conn, P.J.; Duvoisin, R.; Ferraguti, F.; Flor, P.J.; Goudet, C.; Gregory, K.J.; Hampson, D.; et al. Metabotropic Glutamate Receptors in GtoPdb v.2023.1. Available online: http://journals.ed.ac.uk/gtopdb-cite/article/view/8689 (accessed on 1 June 2023).
- Monn, J.A.; Valli, M.J.; Massey, S.M.; Hansen, M.M.; Kress, T.J.; Wepsiec, J.P.; Harkness, A.R.; Grutsch, J.L., Jr.; Wright, R.A.; Johnson, B.G.; et al. Synthesis, pharmacological characterization, and molecular modeling of heterobicyclic amino acids related to (+)-2-aminobicyclo[3.1.0] hexane-2,6-dicarboxylic acid (LY354740): Identification of two new potent, selective, and systemically active agonists for group II metabotropic glutamate receptors. J. Med. Chem. 1999, 42, 1027–1040. [Google Scholar] [CrossRef]
- Hubner, K.; Surovtsova, I.; Yserentant, K.; Hansch, M.; Kummer, U. Ca2+ dynamics correlates with phenotype and function in primary human neutrophils. Biophys. Chem. 2013, 184, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Watson, F.; Edwards, S.W. Stimulation of primed neutrophils by soluble immune complexes: Priming leads to enhanced intracellular Ca2+ elevations, activation of phospholipase D, and activation of the NADPH oxidase. Biochem. Biophys. Res. Commun. 1998, 247, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Kuhns, D.B.; Young, H.A.; Gallin, E.K.; Gallin, J.I. Ca2+-dependent production and release of IL-8 in human neutrophils. J. Immunol. 1998, 161, 4332–4339. [Google Scholar] [CrossRef]
- Vogt, K.L.; Summers, C.; Chilvers, E.R.; Condliffe, A.M. Priming and de-priming of neutrophil responses in vitro and in vivo. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12967. [Google Scholar] [CrossRef]
- Krause, K.H.; Campbell, K.P.; Welsh, M.J.; Lew, D.P. The calcium signal and neutrophil activation. Clin. Biochem. 1990, 23, 159–166. [Google Scholar] [CrossRef]
- Clemens, R.A.; Lowell, C.A. Store-operated calcium signaling in neutrophils. J. Leukoc. Biol. 2015, 98, 497–502. [Google Scholar] [CrossRef]
- Chen, L.W.; Jan, C.R. Mechanisms and modulation of formyl-methionyl-leucyl-phenylalanine (fMLP)-induced Ca2+ mobilization in human neutrophils. Int. Immunopharmacol. 2001, 1, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Pettit, E.J.; Hallett, M.B. Two distinct Ca2+ storage and release sites in human neutrophils. J. Leukoc. Biol. 1998, 63, 225–232. [Google Scholar] [CrossRef]
- Gama, L.; Wilt, S.G.; Breitwieser, G.E. Heterodimerization of calcium sensing receptors with metabotropic glutamate receptors in neurons. J. Biol. Chem. 2001, 276, 39053–39059. [Google Scholar] [CrossRef]
- Senga, T.; Iwamoto, S.; Yoshida, T.; Yokota, T.; Adachi, K.; Azuma, E.; Hamaguchi, M.; Iwamoto, T. LSSIG is a novel murine leukocyte-specific GPCR that is induced by the activation of STAT3. Blood 2003, 101, 1185–1187. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.; Krauss-Etschmann, S.; Koller, B.; Hordijk, P.L.; Kuijpers, T.W.; Hoffmann, F.; Hector, A.; Eber, E.; Marcos, V.; Bittmann, I.; et al. Infiltrated neutrophils acquire novel chemokine receptor expression and chemokine responsiveness in chronic inflammatory lung diseases. J. Immunol. 2008, 181, 8053–8067. [Google Scholar] [CrossRef] [PubMed]
- Comerford, I.; Harata-Lee, Y.; Bunting, M.D.; Gregor, C.; Kara, E.E.; McColl, S.R. A myriad of functions and complex regulation of the CCR7/CCL19/CCL21 chemokine axis in the adaptive immune system. Cytokine Growth Factor Rev. 2013, 24, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.T.; Herz, C.; Ruf, P.; Stetter, R.; Lamy, E. Human T2R38 Bitter Taste Receptor Expression in Resting and Activated Lymphocytes. Front. Immunol. 2018, 9, 2949. [Google Scholar] [CrossRef]
- Foster, S.R.; Porrello, E.R.; Purdue, B.; Chan, H.W.; Voigt, A.; Frenzel, S.; Hannan, R.D.; Moritz, K.M.; Simmons, D.G.; Molenaar, P.; et al. Expression, regulation and putative nutrient-sensing function of taste GPCRs in the heart. PLoS ONE 2013, 8, e64579. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Selvam, B.; Sanematsu, K.; Shigemura, N.; Shukla, D.; Procko, E. Structural architecture of a dimeric class C GPCR based on co-trafficking of sweet taste receptor subunits. J. Biol. Chem. 2019, 294, 4759–4774. [Google Scholar] [CrossRef]
- Gassmann, M.; Haller, C.; Stoll, Y.; Abdel Aziz, S.; Biermann, B.; Mosbacher, J.; Kaupmann, K.; Bettler, B. The RXR-type endoplasmic reticulum-retention/retrieval signal of GABAB1 requires distant spacing from the membrane to function. Mol. Pharmacol. 2005, 68, 137–144. [Google Scholar] [CrossRef]
- Margeta-Mitrovic, M.; Jan, Y.N.; Jan, L.Y. A trafficking checkpoint controls GABA(B) receptor heterodimerization. Neuron 2000, 27, 97–106. [Google Scholar] [CrossRef]
- Masubuchi, Y.; Nakagawa, Y.; Ma, J.; Sasaki, T.; Kitamura, T.; Yamamoto, Y.; Kurose, H.; Kojima, I.; Shibata, H. A novel regulatory function of sweet taste-sensing receptor in adipogenic differentiation of 3T3-L1 cells. PLoS ONE 2013, 8, e54500. [Google Scholar] [CrossRef]
- Xu, H.; Staszewski, L.; Tang, H.; Adler, E.; Zoller, M.; Li, X. Different functional roles of T1R subunits in the heteromeric taste receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 14258–14263. [Google Scholar] [CrossRef]
- Önnheim, K.; Christenson, K.; Gabl, M.; Burbiel, J.C.; Müller, C.E.; Oprea, T.I.; Bylund, J.; Dahlgren, C.; Forsman, H. A novel receptor cross-talk between the ATP receptor P2Y2 and formyl peptide receptors reactivates desensitized neutrophils to produce superoxide. Exp. Cell. Res. 2014, 323, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Andersen, G.; Kahlenberg, K.; Krautwurst, D.; Somoza, V. [6]-Gingerol facilitates CXCL8 secretion and ROS production in primary human neutrophils by targeting the TRPV1 channel. Mol. Nutr. Food Res. 2022, 67, e2200434. [Google Scholar] [CrossRef]
- McCormick, B.; Chu, J.Y.; Vermeren, S. Cross-talk between Rho GTPases and PI3K in the neutrophil. Small GTPases 2019, 10, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Hafkamp, F.M.J.; Varela, L.; Simkhada, N.; Taanman-Kueter, E.W.; Tas, S.W.; Wauben, M.H.M.; Groot Kormelink, T.; de Jong, E.C. Efficient Neutrophil Activation Requires Two Simultaneous Activating Stimuli. Int. J. Mol. Sci. 2021, 22, 10106. [Google Scholar] [CrossRef]
- Abreu, N.; Acosta-Ruiz, A.; Xiang, G.; Levitz, J. Mechanisms of differential desensitization of metabotropic glutamate receptors. Cell Rep. 2021, 35, 109050. [Google Scholar] [CrossRef] [PubMed]
- Iacovelli, L.; Molinaro, G.; Battaglia, G.; Motolese, M.; Di Menna, L.; Alfiero, M.; Blahos, J.; Matrisciano, F.; Corsi, M.; Corti, C.; et al. Regulation of group II metabotropic glutamate receptors by G protein-coupled receptor kinases: mGlu2 receptors are resistant to homologous desensitization. Mol. Pharmacol. 2009, 75, 991–1003. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Vandesompele, J.; Kubista, M.; Pfaffl, M.W. Reference gene validation software for improved normalization. In Real-Time PCR: Current Technology and Applications; Logan, J., Edwards, K., Saunders, N., Eds.; Caister Academic Press: Norfolk, UK, 2009; pp. 47–64. [Google Scholar]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Geithe, C.; Andersen, G.; Malki, A.; Krautwurst, D. A Butter Aroma Recombinate Activates Human Class-I Odorant Receptors. J. Agric. Food Chem. 2015, 63, 9410–9420. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G-Protein | mGlu2 | mGlu2/TAS1R3 | TAS1R3 |
|---|---|---|---|
| GαGustducin | 56.14 ± 10.12 a | 25.66 ± 6.26 A* | n.d. |
| Gαi1 | 18.11 ± 2.91 a | 5.88 ± 0.97 B* | n.d. |
| Gαi2 | 19.88 ± 2.46 a | 7.79 ± 0.52 A* | n.d. |
| Gαi3 | 8.38 ± 0.50 b | 4.97 ± 0.59 B* | n.d. |
| Receptors | IC50 Value |
|---|---|
| mGlu2 | 8.38 ± 0.50 a |
| mGlu2/TAS1R3 | 4.97 ± 0.59 b |
| mGlu2/TAS1R2 | 10.84 ± 2.13 a |
| mGlu2/TAS1R1 | 10.62 ± 1.26 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ball, L.; Bauer, J.; Krautwurst, D. Heterodimerization of Chemoreceptors TAS1R3 and mGlu2 in Human Blood Leukocytes. Int. J. Mol. Sci. 2023, 24, 12942. https://doi.org/10.3390/ijms241612942
Ball L, Bauer J, Krautwurst D. Heterodimerization of Chemoreceptors TAS1R3 and mGlu2 in Human Blood Leukocytes. International Journal of Molecular Sciences. 2023; 24(16):12942. https://doi.org/10.3390/ijms241612942
Chicago/Turabian StyleBall, Lena, Julia Bauer, and Dietmar Krautwurst. 2023. "Heterodimerization of Chemoreceptors TAS1R3 and mGlu2 in Human Blood Leukocytes" International Journal of Molecular Sciences 24, no. 16: 12942. https://doi.org/10.3390/ijms241612942
APA StyleBall, L., Bauer, J., & Krautwurst, D. (2023). Heterodimerization of Chemoreceptors TAS1R3 and mGlu2 in Human Blood Leukocytes. International Journal of Molecular Sciences, 24(16), 12942. https://doi.org/10.3390/ijms241612942