

Porphyromonas gingivalis Peptidyl Arginine Deiminase (PPAD) in the Context of the Feed-Forward Loop of Inflammation in Periodontitis
 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
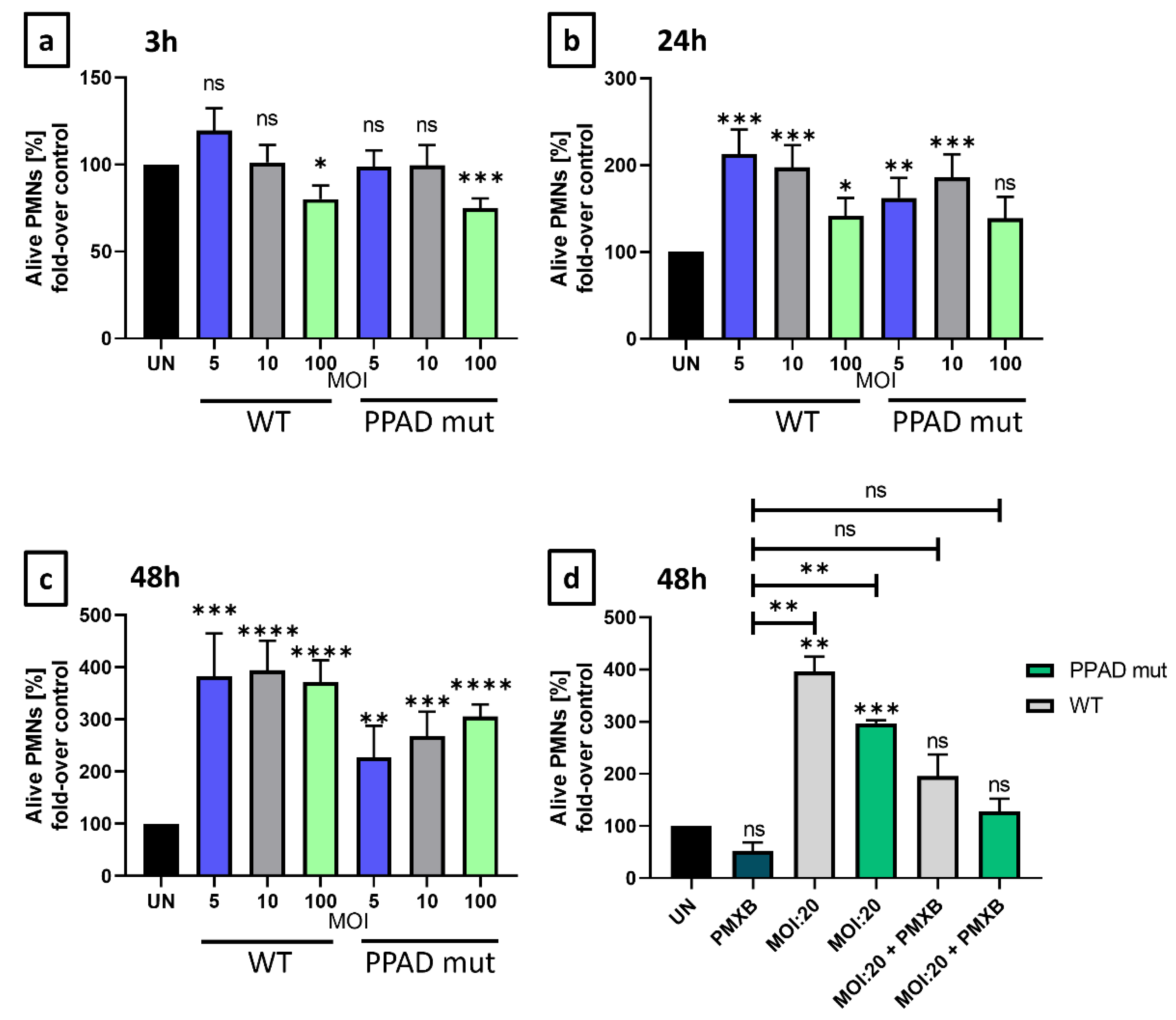
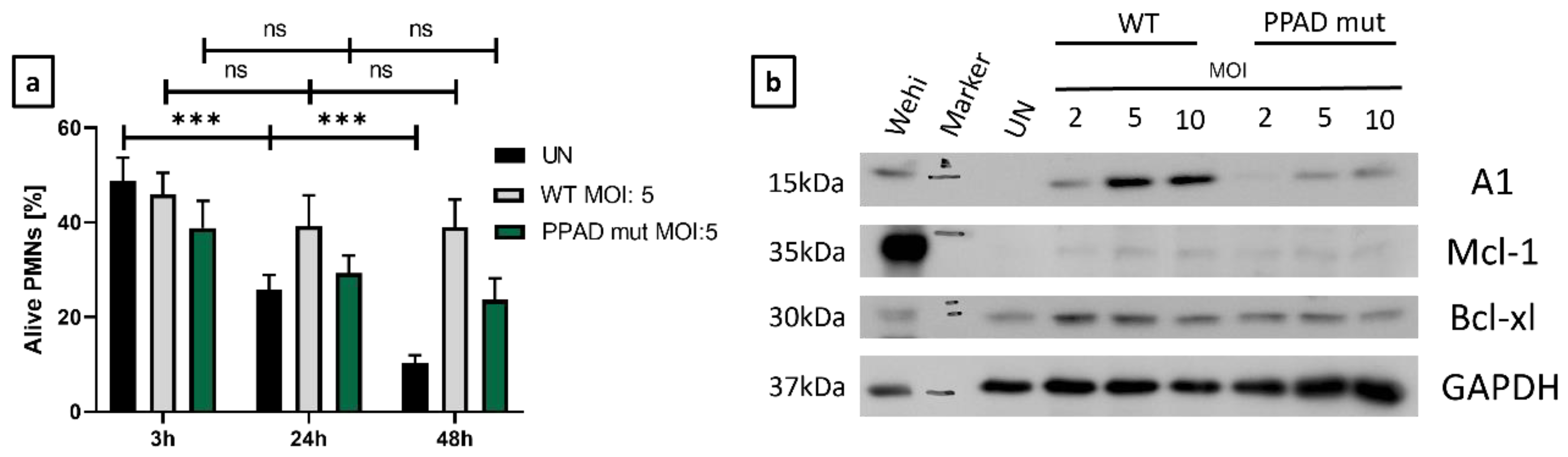
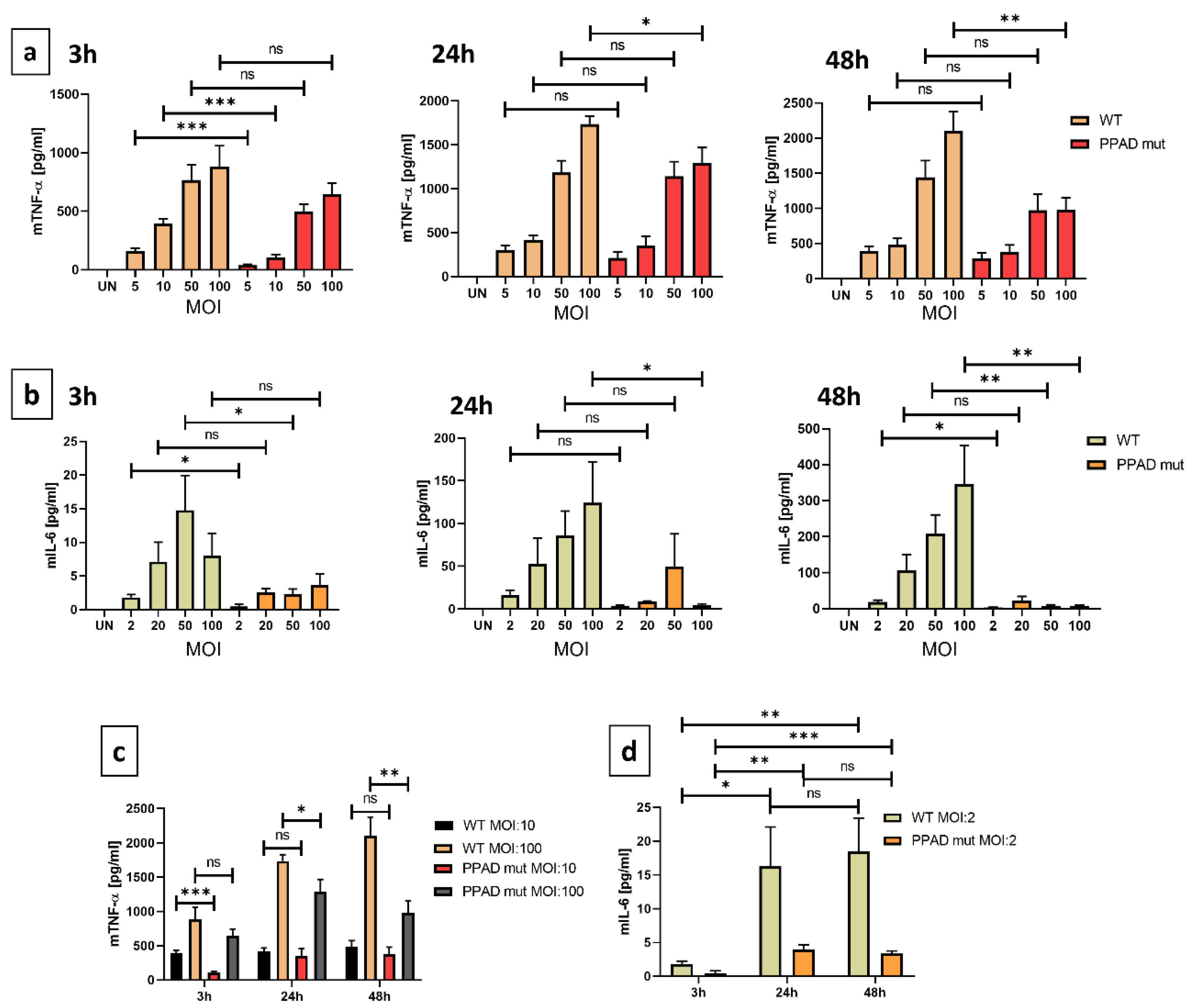
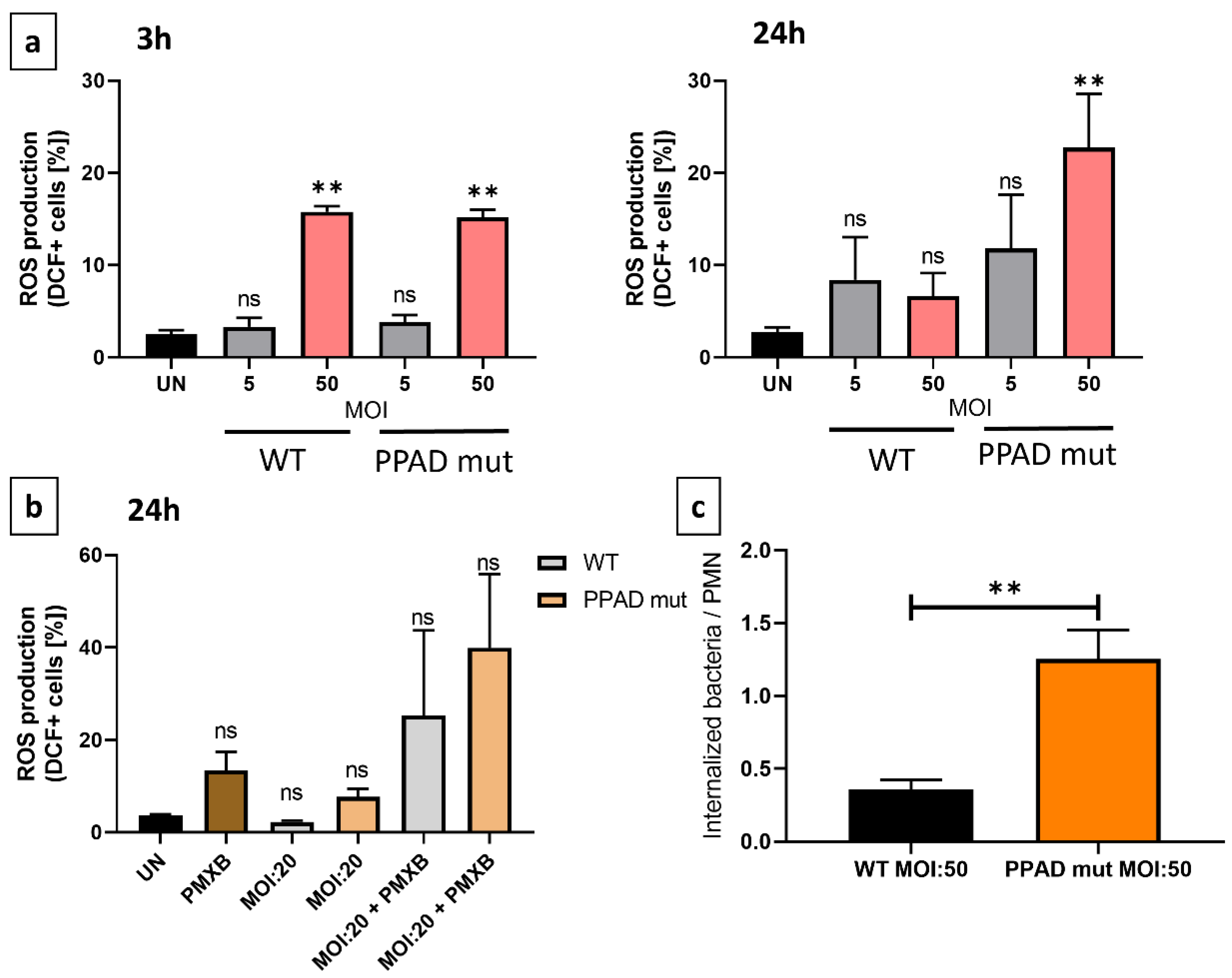
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Bacteria Strains
4.2. Flow Cytometry
4.3. Immunoblotting
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Invasion Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oral Health. Available online: https://www.who.int/news-room/fact-sheets/detail/oral-health (accessed on 23 March 2021).
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Van Dyke, T.E.; Sheilesh, D. Risk factors for periodontitis. J. Int. Acad. Periodontol. 2005, 7, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef] [PubMed]
- De Pablo, P.; Chapple, I.L.C.; Buckley, C.D.; Dietrich, T. Periodontitis in systemic rheumatic diseases. Nat. Rev. Rheumatol. 2009, 5, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Potempa, J.; Mydel, P.; Koziel, J. The case for periodontitis in the pathogenesis of rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 606–620. [Google Scholar] [CrossRef]
- McGraw, W.T.; Potempa, J.; Farley, D.; Travis, J. Purification, characterization, and sequence analysis of a potential virulence factor from Porphyromonas gingivalis, peptidylarginine deiminase. Infect. Immun. 1999, 67, 3248–3256. [Google Scholar] [CrossRef]
- Gabarrini, G.; de Smit, M.; Westra, J.; Brouwer, E.; Vissink, A.; Zhou, K.; Rossen, J.W.A.; Stobernack, T.; van Dijl, J.M.; van Winkelhoff, A.J. The peptidylarginine deiminase gene is a conserved feature of Porphyromonas gingivalis. Sci. Rep. 2015, 5, 13936. [Google Scholar] [CrossRef]
- Abdullah, S.N.; Farmer, E.A.; Spargo, L.; Logan, R.; Gully, N. Porphyromonas gingivalis peptidylarginine deiminase substrate specificity. Anaerobe 2013, 23, 102–108. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K.; Potempa, J. Citrullination as a plausible link to periodontitis, rheumatoid arthritis, atherosclerosis and Alzheimer’s disease. J. Oral Microbiol. 2018, 10, 1487742. [Google Scholar] [CrossRef]
- Vermilyea, D.M.; Ottenberg, G.K.; Davey, M.E. Citrullination mediated by PPAD constrains biofilm formation in P. gingivalis strain 381. NPJ Biofilms Microbiomes 2019, 5, 7. [Google Scholar] [CrossRef]
- Karkowska-Kuleta, J.; Bartnicka, D.; Zawrotniak, M.; Zielinska, G.; Kierońska, A.; Bochenska, O.; Ciaston, I.; Koziel, J.; Potempa, J.; Baster, Z.; et al. The activity of bacterial peptidylarginine deiminase is important during formation of dual-species biofilm by periodontal pathogen Porphyromonas gingivalis and opportunistic fungus Candida albicans. Pathog. Dis. 2018, 76, fty033. [Google Scholar] [CrossRef] [PubMed]
- Gully, N.; Bright, R.; Marino, V.; Marchant, C.; Cantley, M.; Haynes, D.; Butler, C.; Dashper, S.; Reynolds, E.; Bartold, M. Porphyromonas gingivalis peptidylarginine deiminase, a key contributor in the pathogenesis of experimental periodontal disease and experimental arthritis. PLoS ONE 2014, 9, e100838. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from Marrow to Microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Z.; Li, X.; Dong, G.; Zhang, M.; Xu, Z.; Yang, J. Neutrophil Extracellular Traps: Signaling Properties and Disease Relevance. Mediators Inflamm. 2020, 2020, 9254087. [Google Scholar] [CrossRef]
- Scott, D.A.; Krauss, J. Neutrophils in periodontal inflammation. Front. Oral Biol. 2012, 15, 56–83. [Google Scholar] [CrossRef] [PubMed]
- Vladimer, G.I.; Marty-Roix, R.; Ghosh, S.; Weng, D.; Lien, E. Inflammasomes and host defenses against bacterial infections. Curr. Opin. Microbiol. 2013, 16, 23–31. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Haslett, C.; Lee, A.; Savill, J.S.; Meagher, L.; Whyte, M.K. Apoptosis (programmed cell death) and functional changes in aging neutrophils: Modulation by inflammatory mediators. In Chest; Elsevier: Amsterdam, The Netherlands, 1991; p. 6S. [Google Scholar] [CrossRef]
- Savill, J.S.; Henson, P.M.; Haslett, C. Phagocytosis of aged human neutrophils by macrophages is mediated by a novel ’charge-sensitive’recognition mechanism. J. Clin. Investig. 1989, 84, 1518–1527. [Google Scholar] [CrossRef]
- Preshaw, P.M.; Schifferle, R.E.; Walters, J.D. Porphyromonas gingivalis lipopolysaccharide delays human polymorphonuclear leukocyte apoptosis in vitro. J. Periodontal Res. 1999, 34, 197–202. [Google Scholar] [CrossRef]
- Leira, Y.; Iglesias-Rey, R.; Gómez-Lado, N.; Aguiar, P.; Sobrino, T.; D’Aiuto, F.; Castillo, J.; Blanco, J.; Campos, F. Periodontitis and vascular inflammatory biomarkers: An experimental in vivo study in rats. Odontology 2020, 108, 202–212. [Google Scholar] [CrossRef]
- Zaric, S.; Shelburne, C.; Darveau, R.; Quinn, D.J.; Weldon, S.; Taggart, C.C.; Coulter, W.A. Impaired immune tolerance to Porphyromonas gingivalis lipopolysaccharide promotes neutrophil migration and decreased apoptosis. Infect Immun. 2010, 78, 4151–4156. [Google Scholar] [CrossRef] [PubMed]
- Sochalska, M.; Stańczyk, M.B.; Użarowska, M.; Zubrzycka, N.; Kirschnek, S.; Grabiec, A.M.; Kantyka, T.; Potempa, J. Application of the in vitro HOXB8 model system to characterize the contributions of neutrophil–lps interaction to periodontal disease. Pathogens 2020, 9, 530. [Google Scholar] [CrossRef] [PubMed]
- Kriebel, K.; Hieke, C.; Engelmann, R.; Potempa, J.; Müller-Hilke, B.; Lang, H.; Kreikemeyer, B. Porphyromonas gingivalis Peptidyl Arginine Deiminase Can Modulate Neutrophil Activity via Infection of Human Dental Stem Cells. J. Innate Immun. 2018, 10, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Aliko, A.; Kamińska, M.; Bergum, B.; Gawron, K.; Benedyk, M.; Lamont, R.J.; Malicki, S.; Delaleu, N.; Potempa, J.; Mydel, P. Impact of Porphyromonas gingivalis Peptidylarginine Deiminase on Bacterial Biofilm Formation, Epithelial Cell Invasion, and Epithelial Cell Transcriptional Landscape. Sci. Rep. 2018, 8, 14144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, J.; Xia, J.; Chen, Z.; Chen, X. Delayed apoptosis by neutrophils from COPD patients is associated with altered bak, bcl-xl, and mcl-1 mRNA expression. Diagn. Pathol. 2012, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Vier, J.; Groth, M.; Sochalska, M.; Kirschnek, S. The anti-apoptotic Bcl-2 family protein A1/Bfl-1 regulates neutrophil survival and homeostasis and is controlled via PI3K and JAK/STAT signaling. Cell Death Dis. 2016, 7, e1336. [Google Scholar] [CrossRef]
- Caton, J.G.; Armitage, G.; Berglundh, T.; Chapple, I.L.C.; Jepsen, S.; Kornman, K.S.; Mealey, B.L.; Papapanou, P.N.; Sanz, M.; Tonetti, M.S. A new classification scheme for periodontal and peri-implant diseases and conditions—Introduction and key changes from the 1999 classification. J. Clin. Periodontol. 2018, 45, S1–S8. [Google Scholar] [CrossRef]
- Prucsi, Z.; Płonczyńska, A.; Potempa, J.; Sochalska, M. Uncovering the Oral Dysbiotic Microbiota as Masters of Neutrophil Responses in the Pathobiology of Periodontitis. Front. Microbiol. 2021, 12, 2980. [Google Scholar] [CrossRef]
- Domingues, M.M.; Inácio, R.G.; Raimundo, J.M.; Martins, M.; Castanho, M.A.R.B.; Santos, N.C. Biophysical characterization of polymyxin B interaction with LPS aggregates and membrane model systems. Biopolymers 2012, 98, 338–344. [Google Scholar] [CrossRef]
- Moulding, D.A.; Akgul, C.; Derouet, M.; White, M.R.; Edwards, S.W. BCL-2 family expression in human neutrophils during delayed and accelerated apoptosis. J. Leukoc. Biol. 2001, 70, 783–792. [Google Scholar] [CrossRef]
- Kelly, G.L.; Strasser, A. Toward Targeting Antiapoptotic MCL-1 for Cancer Therapy. Annu. Rev. Cancer Biol. 2020, 4, 299–313. [Google Scholar] [CrossRef]
- Cardoso, E.M.; Reis, C.; Manzanares-Céspedes, M.C. Chronic periodontitis, inflammatory cytokines, and interrelationship with other chronic diseases. In Postgraduate Medicine; Taylor and Francis Inc.: Oxfordshire, UK, 2018; Volume 130, pp. 98–104. [Google Scholar] [CrossRef]
- Wielento, A.; Bereta, G.P.; Szczęśniak, K.; Jacuła, A.; Terekhova, M.; Artyomov, M.N.; Hasegawa, Y.; Grabiec, A.M.; Potempa, J. Accessory fimbrial subunits and PPAD are necessary for TLR2 activation by Porphyromonas gingivalis. Mol. Oral Microbiol. 2023, 38, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.K.; Jung, M.; Kim, S.H.; Lee, S.R.; Park, K.H.; Kim, D.H.; Kim, H.H.; Park, Y.G. Assessment of IL-6, IL-8 and TNF-α levels in the gingival tissue of patients with periodontitis. Exp. Ther. Med. 2013, 6, 847. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Hyun, J.; Park, E.; Lee, B.L.; Kim, K.K.; Choi, Y. Susceptibility of various oral bacteria to antimicrobial peptides and to phagocytosis by neutrophils. J. Periodontal Res. 2007, 42, 410–419. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.-C.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Fathalla, A.M.; Chow, S.H.; Naderer, T.; Zhou, Q.T.; Velkov, T.; Azad, M.A.K.; Li, J. Polymyxin-induced cell death of human macrophage-like THP-1 and neutrophil-like HL-60 cells associated with the activation of apoptotic pathways. Antimicrob. Agents Chemother. 2020, 64, e00013-20. [Google Scholar] [CrossRef]
- Dahlen, G.; Basic, A.; Bylund, J. Importance of Virulence Factors for the Persistence of Oral Bacteria in the Inflamed Gingival Crevice and in the Pathogenesis of Periodontal Disease. J. Clin. Med. 2019, 8, 1339. [Google Scholar] [CrossRef]
- Stobernack, T.; Glasner, C.; Junker, S.; Gabarrini, G.; de Smit, M.; de Jong, A.; Otto, A.; Becher, D.; Jan van Winkelhoff, A.; van Dijl, J.M. Extracellular Proteome and Citrullinome of the Oral Pathogen Porphyromonas gingivalis. J. Proteome Res. 2016, 15, 4532–4543. [Google Scholar] [CrossRef]
- Wielento, A.; Bereta, G.P.; Łagosz-Ćwik, K.B.; Eick, S.; Lamont, R.J.; Grabiec, A.M.; Potempa, J. TLR2 Activation by Porphyromonas gingivalis Requires Both PPAD Activity and Fimbriae. Front. Immunol. 2022, 13, 823685. [Google Scholar] [CrossRef]
- Akgul, C.; Moulding, D.A.; Edwards, S.W. Molecular control of neutrophil apoptosis. FEBS Lett. 2001, 487, 318–322. [Google Scholar] [CrossRef]
- Sochalska, M.; Tuzlak, S.; Egle, A.; Villunger, A. Lessons from gain- and loss-of-function models of pro-survival Bcl2 family proteins: Implications for targeted therapy. FEBS J. 2015, 282, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. In Cell Death and Differentiation; Nature Publishing Group: New York, NY, USA, 2018; Volume 25, pp. 65–80. [Google Scholar] [CrossRef]
- Dai, H.; Meng, X.W.; Kaufmann, S.H. Selective Inhibition of BFL1: It’s All about Finding the Right Partner. Cell Chem. Biol. 2020, 27, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dou, J.; You, Q.; Jiang, Z. Inhibitors of BCL2A1/Bfl-1 protein: Potential stock in cancer therapy. Eur. J. Med. Chem. 2021, 220, 113539. [Google Scholar] [CrossRef] [PubMed]
- Salamone, G.; Giordano, M.; Trevani, S.; Gamberale, R.; Vermeulen, M.; Schettinni, J.; Geffner, J.R. Promotion of Neutrophil Apoptosis by TNF-α. J. Immunol. 2001, 166, 3476–3483. [Google Scholar] [CrossRef]
- Akgul, C.; Edwards, S.W. Regulation of neutrophil apoptosis via death receptors. Cell. Mol. Life Sci. 2003, 60, 2402–2408. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Dadgostar, H.; Cheng, Q.; Shu, J.; Cheng, G. NF-κB-mediated up-regulation of Bcl-x and Bfl-1/A1 is required for CD40 survival signaling in B lymphocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 9136–9141. [Google Scholar] [CrossRef]
- Cherla, R.; Zhang, Y.; Ledbetter, L.; Zhang, G. Coxiella burnetii inhibits neutrophil apoptosis by exploiting survival pathways and antiapoptotic protein Mcl-1. Infect. Immun. 2018, 86, e00504-17. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef]
- Rathinasamy, K.; Ulaganathan, A.; Ramamurthy, S.; Ganesan, R.; Saket, P.; Alamelu, S. Estimation of TNF-α levels in saliva and serum of patients with periodontal health and chronic periodontitis: A case-control study. J. Contemp. Dent. Pract. 2020, 21, 148–151. [Google Scholar] [CrossRef]
- Batool, H.; Nadeem, A.; Kashif, M.; Shahzad, F.; Tahir, R.; Afzal, N. Salivary Levels of IL-6 and IL-17 Could Be an Indicator of Disease Severity in Patients with Calculus Associated Chronic Periodontitis. Biomed. Res. Int. 2018, 2018, 8531961. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.H.; Hardy, D.C.; Schuyler, C.A.; Slate, E.H.; Mize, T.W.; Huang, Y. Expression of periodontal interleukin-6 protein is increased across patients with neither periodontal disease nor diabetes, patients with periodontal disease alone and patients with both diseases. J. Periodontal Res. 2010, 45, 688–694. [Google Scholar] [CrossRef]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [PubMed]
- Biffl, W.L.; Moore, E.E.; Moore, F.A.; Barnett, C.C.; Carl, V.S.; Peterson, V.M. Interleukin-6 delays neutrophil apoptosis. Arch. Surg. 1996, 131, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Biffl, W.L.; Moore, E.E.; Moore, F.A.; Barnett, C.C. Interleukin-6 suppression of neutrophil apoptosis is neutrophil concentration dependent. J. Leukoc. Biol. 1995, 58, 582–584. [Google Scholar] [CrossRef]
- Pan, W.; Wang, Q.; Chen, Q. The cytokine network involved in the host immune response to periodontitis. Int. J. Oral Sci. 2019, 11, 30. [Google Scholar] [CrossRef]
- Stobernack, T.; Espina, M.D.T.; Mulder, L.M.; Medina, L.M.P.; Piebenga, D.R.; Gabarrini, G.; Zhao, X.; Janssen, K.M.J.; Hulzebos, J.; Brouwer, E.; et al. A secreted bacterial peptidylarginine deiminase can neutralize human innate immune defenses. mBio 2018, 9, e01704-18. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, E.; Scavenius, C.; Kantyka, T.; Jusko, M.; Mizgalska, D.; Szmigielski, B.; Potempa, B.; Enghild, J.J.; Prossnitz, E.R.; Blom, A.M.; et al. Peptidyl arginine deiminase from porphyromonas gingivalis abolishes anaphylatoxin C5a activity. J. Biol. Chem. 2014, 289, 32481–32487. [Google Scholar] [CrossRef]
- Maekawa, T.; Krauss, J.L.; Abe, T.; Jotwani, R.; Triantafilou, M.; Triantafilou, K.; Hashim, A.; Hoch, S.; Curtis, M.A.; Nussbaum, G.; et al. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell. Host Microbe 2014, 15, 768–778. [Google Scholar] [CrossRef]
- Goodson, J.M. Diagnosis of Periodontitis by Physical Measurement: Interpretation From Episodic Disease Hypothesis. J. Periodontol. 1992, 63, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Elashiry, M.; Meghil, M.M.; Arce, R.M.; Cutler, C.W. From manual periodontal probing to digital 3-D imaging to endoscopic capillaroscopy: Recent advances in periodontal disease diagnosis. J. Periodontal Res. 2019, 54, 1–9. [Google Scholar] [CrossRef]
- Graziani, F.; Karapetsa, D.; Alonso, B.; Herrera, D. Nonsurgical and surgical treatment of periodontitis: How many options for one disease? Periodontology 2000 2017, 75, 152–188. [Google Scholar] [CrossRef] [PubMed]
- Hancock, E.B.; Newell, D.H. Preventive strategies and supportive treatment. Periodontology 2000 2001, 25, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Waerhaug, J. Healing of the Dento-Epithelial Junction Following Subgingival Plaque Control: II: As Observed on Extracted Teeth. J. Periodontol. 1978, 49, 119–134. [Google Scholar] [CrossRef]
- National Institute of Dental and Craniofacial Research. Periodontal (Gum) Disease: Causes, Symptoms, and Treatments; National Institutes of Health (NIH): Bethesda, MD, USA, 2008.
- Wang, G.G.; Calvo, K.R.; Pasillas, M.P.; Sykes, D.B.; Häcker, H.; Kamps, M.P. Quantitative production of macrophages or neutrophils ex vivo using conditional Hoxb8. Nat. Methods 2006, 3, 287–293. [Google Scholar] [CrossRef]
- Gawron, K.; Bereta, G.; Nowakowska, Z.; Łazarz, K.; Łazarz, M.; Szmigielski, B.; Mizgalska, D.; Buda, A.; Koziel, J.; Oruba, Z.; et al. Peptidylarginine deiminase from Porphyromonas gingivalis contributes to infection of gingival fibroblasts and induction of prostaglandin E2-signaling pathway. Mol. Oral Microbiol. 2014, 29, 321–332. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prucsi, Z.; Zimny, A.; Płonczyńska, A.; Zubrzycka, N.; Potempa, J.; Sochalska, M. Porphyromonas gingivalis Peptidyl Arginine Deiminase (PPAD) in the Context of the Feed-Forward Loop of Inflammation in Periodontitis. Int. J. Mol. Sci. 2023, 24, 12922. https://doi.org/10.3390/ijms241612922
Prucsi Z, Zimny A, Płonczyńska A, Zubrzycka N, Potempa J, Sochalska M. Porphyromonas gingivalis Peptidyl Arginine Deiminase (PPAD) in the Context of the Feed-Forward Loop of Inflammation in Periodontitis. International Journal of Molecular Sciences. 2023; 24(16):12922. https://doi.org/10.3390/ijms241612922
Chicago/Turabian StylePrucsi, Zsombor, Agnieszka Zimny, Alicja Płonczyńska, Natalia Zubrzycka, Jan Potempa, and Maja Sochalska. 2023. "Porphyromonas gingivalis Peptidyl Arginine Deiminase (PPAD) in the Context of the Feed-Forward Loop of Inflammation in Periodontitis" International Journal of Molecular Sciences 24, no. 16: 12922. https://doi.org/10.3390/ijms241612922
APA StylePrucsi, Z., Zimny, A., Płonczyńska, A., Zubrzycka, N., Potempa, J., & Sochalska, M. (2023). Porphyromonas gingivalis Peptidyl Arginine Deiminase (PPAD) in the Context of the Feed-Forward Loop of Inflammation in Periodontitis. International Journal of Molecular Sciences, 24(16), 12922. https://doi.org/10.3390/ijms241612922