
Insights into the Mechanism of Curaxin CBL0137 Epigenetic Activity: The Induction of DNA Demethylation and the Suppression of BET Family Proteins
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
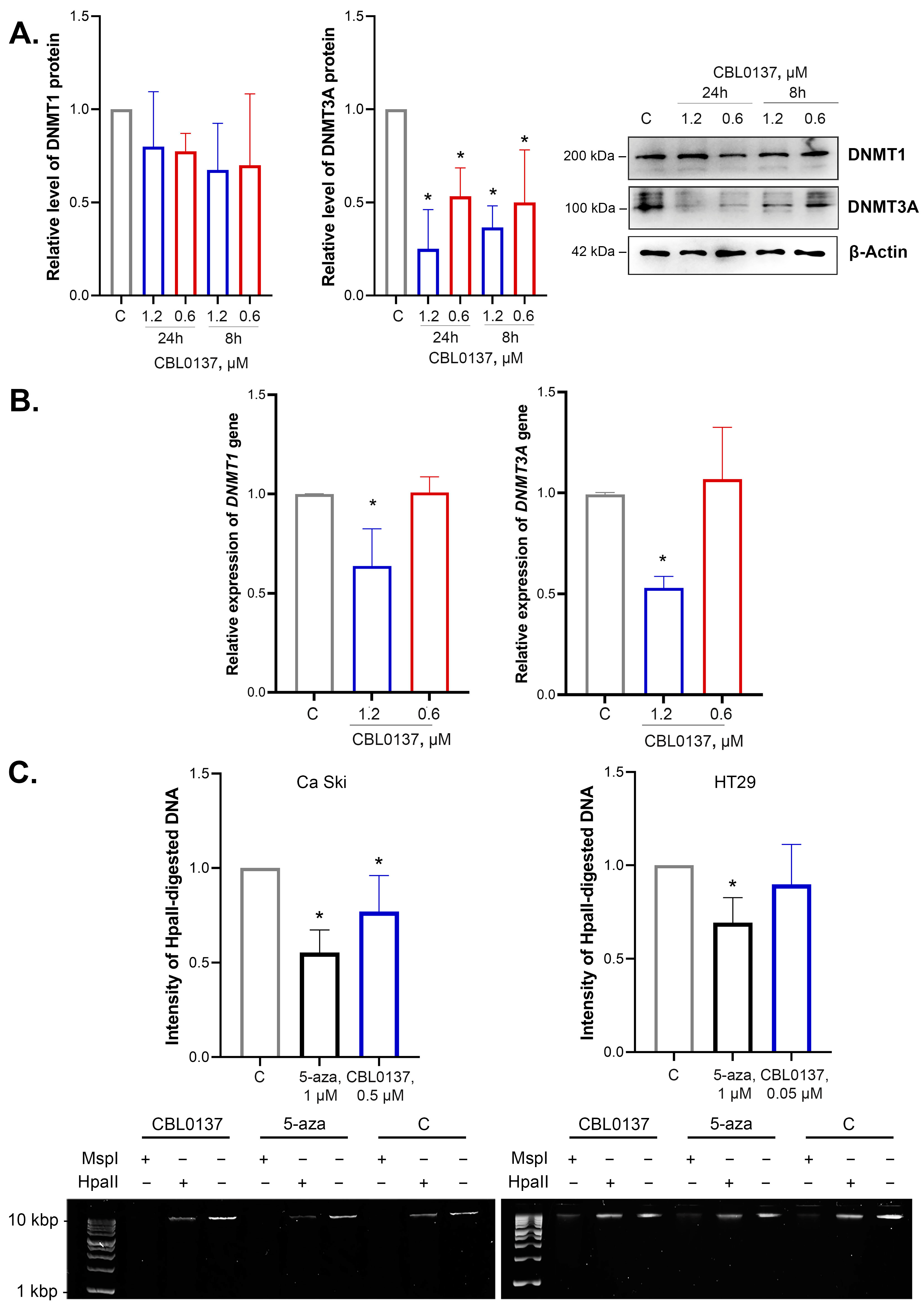
2.1. CBL0137 Inhibited DNMT1 and DNMT3A Expression
2.2. CBL0137 Decreased the Level of DNA Methylation in Ca Ski Cells but Did Not Affect DNA Methylation in HT29 Cells
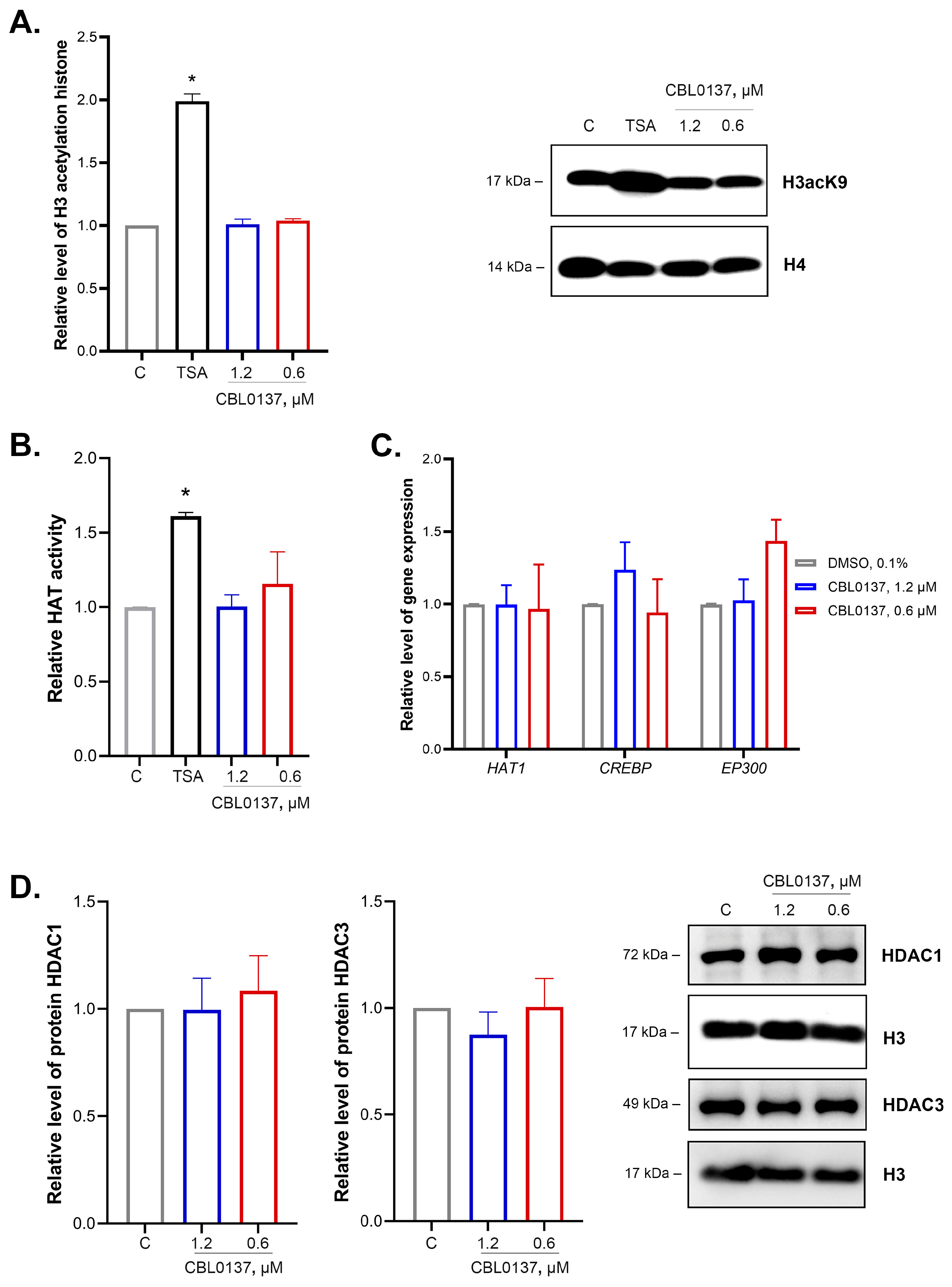
2.3. CBL0137 Did Not Affect the Level of Histone H3 Acetylation, as well as the Activity and Expression of Histone Acetyltransferases or HDAC1 and HDAC3 Expression
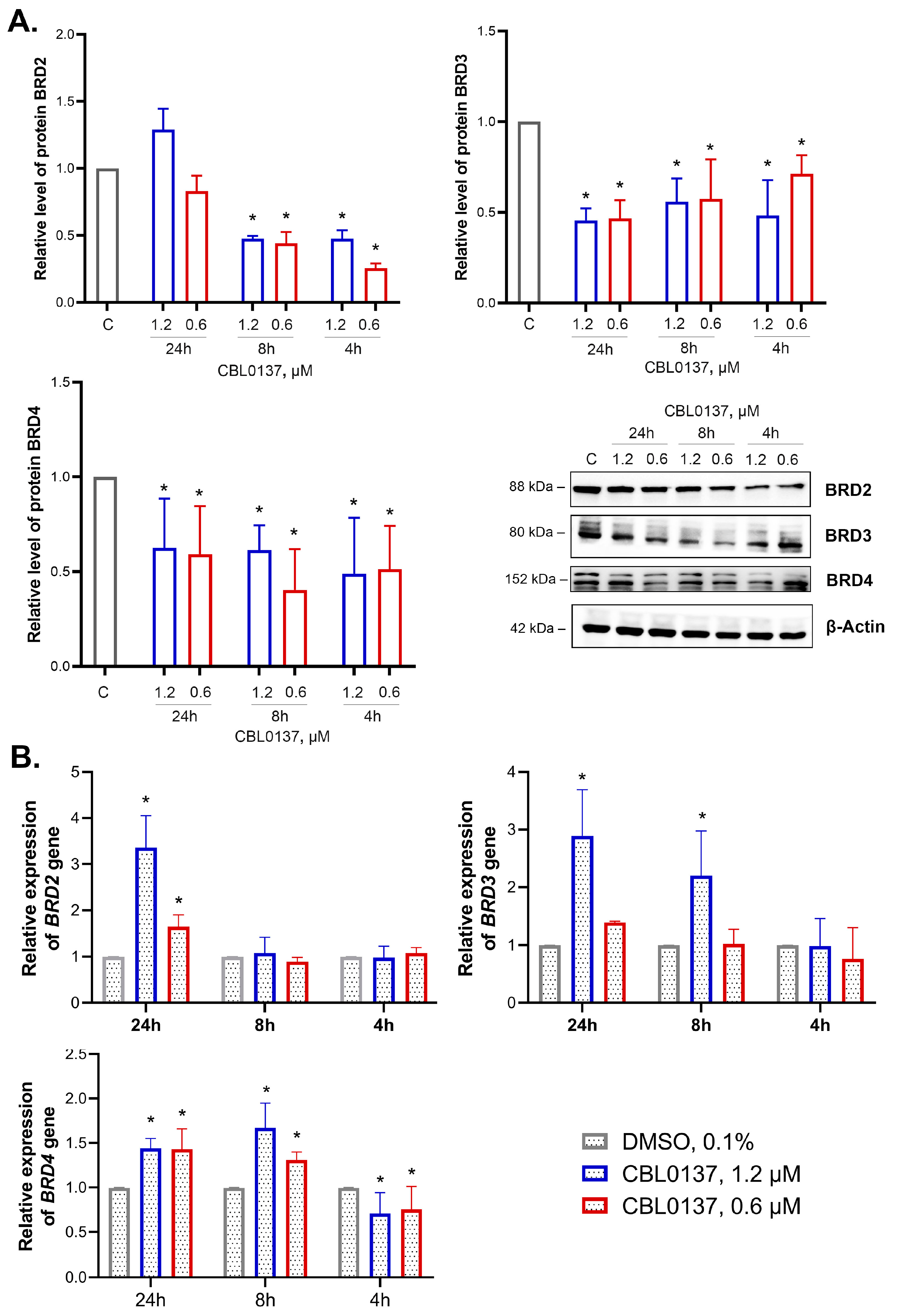
2.4. CBL0137 Inhibited the Expression of BET Family Proteins and Affected BRD2, BRD3 and BRD4 Gene Expression
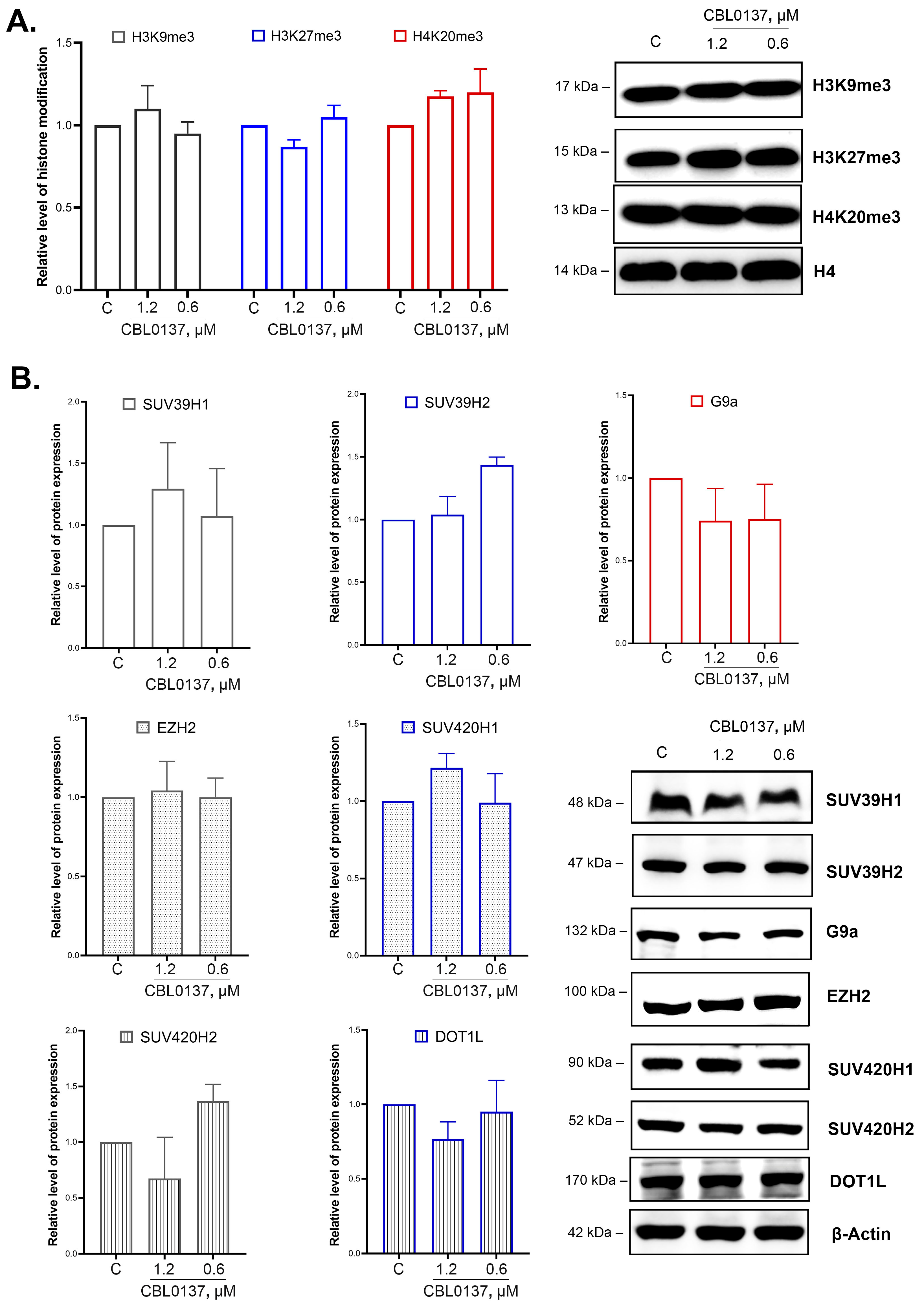
2.5. CBL0137 Did Not Affect the Integral Level of H3K9me3, H3K27me3, and H4K20me3 Histone Modifications in HeLa TI Cells
2.6. CBL0137 Did Not Affect SUV39H1/H2, SUV420H1/H2, G9a, EZH2, and DOT1L Expression Levels
3. Discussion
4. Materials and Methods
4.1. Cell Cultivating
4.2. MTT Test
4.3. DNA Extraction
4.4. MSRE Assay
4.5. Protein Extraction
4.5.1. Histones
4.5.2. Total Protein Fraction
4.5.3. Nuclear Protein Fraction
4.6. Western Blotting
4.6.1. Histone Modifications
4.6.2. DNMTs, HMTs, and BET Family Proteins
4.6.3. HDACs
4.7. HAT Activity Assay
4.8. RNA Extraction and cDNA Synthesis
4.9. Reverse-Transcription Quantitative PCR
4.10. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Chan, Y.T.; Tan, H.Y.; Li, S.; Wang, N.; Feng, Y. Epigenetic regulation in human cancer: The potential role of epi-drug in cancer therapy. Mol. Cancer 2020, 19, 79. [Google Scholar] [CrossRef] [PubMed]
- Tulsyan, S.; Aftab, M.; Sisodiya, S.; Khan, A.; Chikara, A.; Tanwar, P.; Hussain, S. Molecular basis of epigenetic regulation in cancer diagnosis and treatment. Front. Genet. 2022, 13, 885635. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, T.; Luo, H.; Liu, Y.; Liu, D. Targeting Epigenetic Regulatory Enzymes for Cancer Therapeutics: Novel Small-Molecule Epidrug Development. Front. Oncol. 2022, 12, 848221. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Cellular reprogramming to model and study epigenetic alterations in cancer. Stem Cell Res. 2020, 49, 102062. [Google Scholar] [CrossRef]
- Tao, F.; Zhang, Z. Editorial: Epigenetic drugs and therapeutic resistance for epithelial malignancies. Front. Pharmacol. 2023, 14, 1208518. [Google Scholar] [CrossRef]
- Cheng, Y.; He, C.; Wang, M.; Ma, X.; Mo, F.; Yang, S.; Han, J.; Wei, X. Targeting epigenetic regulators for cancer therapy: Mechanisms and advances in clinical trials. Signal Transduct. Target. Ther. 2019, 4, 62. [Google Scholar] [CrossRef]
- Guo, L.; Lee, Y.T.; Zhou, Y.; Huang, Y. Targeting epigenetic regulatory machinery to overcome cancer therapy resistance. Semin. Cancer Biol. 2022, 83, 487–502. [Google Scholar] [CrossRef]
- Morel, D.; Jeffery, D.; Aspeslagh, S.; Almouzni, G.; Postel-Vinay, S. Combining epigenetic drugs with other therapies for solid tumours—Past lessons and future promise. Nat. Rev. Clin. Oncol. 2020, 17, 91–107. [Google Scholar] [CrossRef]
- Ganai, S.A.; Ganai, S.A. Modulating Epigenetic Modification Enzymes Through Relevant Epidrugs as a Timely Strategy in Anticancer Therapy. In Histone Deacetylase Inhibitors in Combinatorial Anticancer Therapy; Springer: Singapore, 2020; pp. 137–157. [Google Scholar] [CrossRef]
- Maksimova, V.P.; Usalka, O.G.; Makus, Y.V.; Popova, V.G.; Trapeznikova, E.S.; Khayrieva, G.I.; Sagitova, G.R.; Zhidkova, E.M.; Prus, A.Y.; Yakubovskaya, M.G.; et al. Aberrations of DNA methylation in cancer. Adv. Mol. Oncol. 2022, 9, 24–40. [Google Scholar] [CrossRef]
- Majchrzak-Celinska, A.; Warych, A.; Szoszkiewicz, M. Novel Approaches to Epigenetic Therapies: From Drug Combinations to Epigenetic Editing. Genes 2021, 12, 208. [Google Scholar] [CrossRef]
- Gasparian, A.V.; Burkhart, C.A.; Purmal, A.A.; Brodsky, L.; Pal, M.; Saranadasa, M.; Bosykh, D.A.; Commane, M.; Guryanova, O.A.; Pal, S.; et al. Curaxins: Anticancer compounds that simultaneously suppress NF-kappaB and activate p53 by targeting FACT. Sci. Transl. Med. 2011, 3, 95ra74. [Google Scholar] [CrossRef]
- Fetisov, T.I.; Borunova, A.A.; Antipova, A.S.; Antoshina, E.E.; Trukhanova, L.S.; Gorkova, T.G.; Zuevskaya, S.N.; Maslov, A.; Gurova, K.; Gudkov, A.; et al. Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo. Biomedicines 2023, 11, 230. [Google Scholar] [CrossRef]
- Jin, M.Z.; Xia, B.R.; Xu, Y.; Jin, W.L. Curaxin CBL0137 Exerts Anticancer Activity via Diverse Mechanisms. Front. Oncol. 2018, 8, 598. [Google Scholar] [CrossRef]
- Kirsanov, K.; Fetisov, T.; Lesovaya, E.A.; Maksimova, V.; Trukhanova, L.; Antoshina, E.; Gor’kova, T.; Morozova, O.; Safina, A.; Fleyshman, D.; et al. Prevention of Colorectal Carcinogenesis by DNA-Binding Small-Molecule Curaxin CBL0137 Involves Suppression of Wnt Signaling. Cancer Prev. Res. 2020, 13, 53–64. [Google Scholar] [CrossRef]
- Song, H.; Xi, S.; Chen, Y.; Pramanik, S.; Zeng, J.; Roychoudhury, S.; Harris, H.; Akbar, A.; Elhag, S.S.; Coulter, D.W.; et al. Histone chaperone FACT complex inhibitor CBL0137 interferes with DNA damage repair and enhances sensitivity of medulloblastoma to chemotherapy and radiation. Cancer Lett. 2021, 520, 201–212. [Google Scholar] [CrossRef]
- Xiao, L.; Karsa, M.; Ronca, E.; Bongers, A.; Kosciolek, A.; El-Ayoubi, A.; Revalde, J.L.; Seneviratne, J.A.; Cheung, B.B.; Cheung, L.C.; et al. The Combination of Curaxin CBL0137 and Histone Deacetylase Inhibitor Panobinostat Delays KMT2A-Rearranged Leukemia Progression. Front. Oncol. 2022, 12, 863329. [Google Scholar] [CrossRef]
- Kantidze, O.L.; Gurova, K.V.; Studitsky, V.M.; Razin, S.V. The 3D Genome as a Target for Anticancer Therapy. Trends Mol. Med. 2020, 26, 141–149. [Google Scholar] [CrossRef]
- Leonova, K.; Safina, A.; Nesher, E.; Sandlesh, P.; Pratt, R.; Burkhart, C.; Lipchick, B.; Gitlin, I.; Frangou, C.; Koman, I.; et al. TRAIN (Transcription of Repeats Activates INterferon) in response to chromatin destabilization induced by small molecules in mammalian cells. eLife 2018, 7, e30842. [Google Scholar] [CrossRef]
- Maksimova, V.; Shalginskikh, N.; Vlasova, O.; Usalka, O.; Beizer, A.; Bugaeva, P.; Fedorov, D.; Lizogub, O.; Lesovaya, E.; Katz, R.; et al. HeLa TI cell-based assay as a new approach to screen for chemicals able to reactivate the expression of epigenetically silenced genes. PLoS ONE 2021, 16, e0252504. [Google Scholar] [CrossRef]
- Poleshko, A.; Einarson, M.B.; Shalginskikh, N.; Zhang, R.; Adams, P.D.; Skalka, A.M.; Katz, R.A. Identification of a functional network of human epigenetic silencing factors. J. Biol. Chem. 2010, 285, 422–433. [Google Scholar] [CrossRef]
- Maksimova, V.; Makus, J.; Popova, V.; Prus, A.; Usalka, O.; Trapeznikova, E.; Zhidkova, E.; Belitsky, G.; Yakubovskaya, M.; Kirsanov, K. Histone Methyltransferases as a New Target for Epigenetic Action of Vorinostat. Biochemistry 2023, 88, 968–978. [Google Scholar] [CrossRef]
- Feng, C.; Dong, J.; Chang, W.; Cui, M.; Xu, T. The Progress of Methylation Regulation in Gene Expression of Cervical Cancer. Int. J. Genom. 2018, 2018, 8260652. [Google Scholar] [CrossRef]
- Bibikova, M.; Lin, Z.; Zhou, L.; Chudin, E.; Garcia, E.W.; Wu, B.; Doucet, D.; Thomas, N.J.; Wang, Y.; Vollmer, E.; et al. High-throughput DNA methylation profiling using universal bead arrays. Genome Res. 2006, 16, 383–393. [Google Scholar] [CrossRef]
- Da Silva, F.A.; Feldberg, E.; Goll, L.G.; Carvalho-Zilse, G.A.; Gross, M.C.; Schneider, C.H.; Matoso, D.A. An HpaII/MspI-PCR assay to measure methylation of DNA in Hoplosternum littorale (Callichthyidae, Siluriformes) from a polluted environment in the central Amazon basin. Environ. Technol. Innov. 2019, 14, 100354. [Google Scholar] [CrossRef]
- Kantidze, O.L.; Luzhin, A.V.; Nizovtseva, E.V.; Safina, A.; Valieva, M.E.; Golov, A.K.; Velichko, A.K.; Lyubitelev, A.V.; Feofanov, A.V.; Gurova, K.V.; et al. The anti-cancer drugs curaxins target spatial genome organization. Nat. Commun. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Volokh, O.I.; Sivkina, A.L.; Moiseenko, A.V.; Popinako, A.V.; Karlova, M.G.; Valieva, M.E.; Kotova, E.Y.; Kirpichnikov, M.P.; Formosa, T.; Studitsky, V.M.; et al. Mechanism of curaxin-dependent nucleosome unfolding by FACT. Front. Mol. Biosci. 2022, 9, 1048117. [Google Scholar] [CrossRef]
- Lu, K.; Liu, C.; Liu, Y.; Luo, A.; Chen, J.; Lei, Z.; Kong, J.; Xiao, X.; Zhang, S.; Wang, Y.Z.; et al. Curaxin-Induced DNA Topology Alterations Trigger the Distinct Binding Response of CTCF and FACT at the Single-Molecule Level. Biochemistry 2021, 60, 494–499. [Google Scholar] [CrossRef]
- Safina, A.; Cheney, P.; Pal, M.; Brodsky, L.; Ivanov, A.; Kirsanov, K.; Lesovaya, E.; Naberezhnov, D.; Nesher, E.; Koman, I.; et al. FACT is a sensor of DNA torsional stress in eukaryotic cells. Nucleic Acids Res. 2017, 45, 1925–1945. [Google Scholar] [CrossRef]
- Sergeev, A.; Vorobyov, A.; Yakubovskaya, M.; Kirsanova, O.; Gromova, E. Novel anticancer drug curaxin CBL0137 impairs DNA methylation by eukaryotic DNA methyltransferase Dnmt3a. Bioorg. Med. Chem. Lett. 2020, 30, 127296. [Google Scholar] [CrossRef]
- Khrabrova, D.A.; Loiko, A.G.; Tolkacheva, A.A.; Cherepanova, N.A.; Zvereva, M.I.; Kirsanova, O.V.; Gromova, E.S. Functional Analysis of DNMT3A DNA Methyltransferase Mutations Reported in Patients with Acute Myeloid Leukemia. Biomolecules 2019, 10, 8. [Google Scholar] [CrossRef]
- Czibere, A.; Bruns, I.; Kroger, N.; Platzbecker, U.; Lind, J.; Zohren, F.; Fenk, R.; Germing, U.; Schroder, T.; Graf, T.; et al. 5-Azacytidine for the treatment of patients with acute myeloid leukemia or myelodysplastic syndrome who relapse after allo-SCT: A retrospective analysis. Bone Marrow Transpl. 2010, 45, 872–876. [Google Scholar] [CrossRef]
- Pradeepa, M.M. Causal role of histone acetylations in enhancer function. Transcription 2017, 8, 40–47. [Google Scholar] [CrossRef]
- Zhou, D.; Wu, Z.; Park, J.G.; Fiches, G.N.; Li, T.W.; Ma, Q.; Huang, H.; Biswas, A.; Martinez-Sobrido, L.; Santoso, N.G.; et al. FACT subunit SUPT16H associates with BRD4 and contributes to silencing of interferon signaling. Nucleic Acids Res. 2022, 50, 8700–8718. [Google Scholar] [CrossRef] [PubMed]
- Altendorfer, E.; Mochalova, Y.; Mayer, A. BRD4: A general regulator of transcription elongation. Transcription 2022, 13, 70–81. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Picaud, S.; Mangos, M.; Keates, T.; Lambert, J.P.; Barsyte-Lovejoy, D.; Felletar, I.; Volkmer, R.; Muller, S.; Pawson, T.; et al. Histone recognition and large-scale structural analysis of the human bromodomain family. Cell 2012, 149, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Tian, J.; Wu, T. BRD4 in physiology and pathology: “BET” on its partners. Bioessays 2021, 43, e2100180. [Google Scholar] [CrossRef]
- Manzotti, G.; Ciarrocchi, A.; Sancisi, V. Inhibition of BET Proteins and Histone Deacetylase (HDACs): Crossing Roads in Cancer Therapy. Cancers 2019, 11, 304. [Google Scholar] [CrossRef]
- Wang, C.; Xu, Q.; Zhang, X.; Day, D.S.; Abraham, B.J.; Lun, K.; Chen, L.; Huang, J.; Ji, X. BRD2 interconnects with BRD3 to facilitate Pol II transcription initiation and elongation to prime promoters for cell differentiation. Cell Mol. Life Sci. 2022, 79, 338. [Google Scholar] [CrossRef]
- Zanconato, F.; Battilana, G.; Forcato, M.; Filippi, L.; Azzolin, L.; Manfrin, A.; Quaranta, E.; Di Biagio, D.; Sigismondo, G.; Guzzardo, V.; et al. Transcriptional addiction in cancer cells is mediated by YAP/TAZ through BRD4. Nat. Med. 2018, 24, 1599–1610. [Google Scholar] [CrossRef]
- Devaiah, B.N.; Case-Borden, C.; Gegonne, A.; Hsu, C.H.; Chen, Q.; Meerzaman, D.; Dey, A.; Ozato, K.; Singer, D.S. BRD4 is a histone acetyltransferase that evicts nucleosomes from chromatin. Nat. Struct. Mol. Biol. 2016, 23, 540–548. [Google Scholar] [CrossRef]
- Xiao, L.; Somers, K.; Murray, J.; Pandher, R.; Karsa, M.; Ronca, E.; Bongers, A.; Terry, R.; Ehteda, A.; Gamble, L.D.; et al. Dual Targeting of Chromatin Stability by The Curaxin CBL0137 and Histone Deacetylase Inhibitor Panobinostat Shows Significant Preclinical Efficacy in Neuroblastoma. Clin. Cancer Res. 2021, 27, 4338–4352. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, J.; Liu, P.Y.; Atmadibrata, B.; Bradner, J.E.; Marshall, G.M.; Lock, R.B.; Liu, T. The Bromodomain Inhibitor JQ1 and the Histone Deacetylase Inhibitor Panobinostat Synergistically Reduce N-Myc Expression and Induce Anticancer Effects. Clin. Cancer Res. 2016, 22, 2534–2544. [Google Scholar] [CrossRef]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef]
- Link, A.J.; LaBaer, J. Trichloroacetic acid (TCA) precipitation of proteins. Cold Spring Harb. Protoc. 2011, 2011, 993–994. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford method for protein quantitation. Methods Mol. Biol. 1994, 32, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Hebert, J.D.; Lee, T.A.; Xing, H.; Boussommier-Calleja, A.; Hynes, R.O.; Lauffenburger, D.A.; Kamm, R.D. Macrophage-Secreted TNFalpha and TGFbeta1 Influence Migration Speed and Persistence of Cancer Cells in 3D Tissue Culture via Independent Pathways. Cancer Res. 2017, 77, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| DNMT1 | GAGCCACAGATGCTGACAAA | TGCCATTAACACCACCTTCA |
| DNMT3A | AGCCCAAGGTCAAGGAGATT | CAGCAGATGGTGCAGTAGGA |
| CREBBP | CTGGCAGACCTCGGAAAGAA | CTGGCGCCGCAAAAACT |
| EP300 | CGCTTTGTCTACACCTGCAA | TGCTGGTTGTTGCTCTCATC |
| HAT1 | GCGATAGAGGCACAACAGAA | TGTATTGTTCGGCATCACTCA |
| BRD2 | CGGCTTATGTTCTCCAACTGCTA | GGCAGTAGAGACTGGTAAAGGC |
| BRD3 | CCAACCATCACTGCAAACGTCAC | GGAGTGGTTGTGTCTGCTTTCC |
| BRD4 | CGCTATGTCACCTCCTGTTTGC | ACTCTGAGGACGAGAAGCCCTT |
| RPLP0 | CCTTCTCCTTTGGGCTGGTCATCCA | CAGACACTGGCAACATTGCGGACAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maksimova, V.; Popova, V.; Prus, A.; Lylova, E.; Usalka, O.; Sagitova, G.; Zhidkova, E.; Makus, J.; Trapeznikova, E.; Belitsky, G.; et al. Insights into the Mechanism of Curaxin CBL0137 Epigenetic Activity: The Induction of DNA Demethylation and the Suppression of BET Family Proteins. Int. J. Mol. Sci. 2023, 24, 12874. https://doi.org/10.3390/ijms241612874
Maksimova V, Popova V, Prus A, Lylova E, Usalka O, Sagitova G, Zhidkova E, Makus J, Trapeznikova E, Belitsky G, et al. Insights into the Mechanism of Curaxin CBL0137 Epigenetic Activity: The Induction of DNA Demethylation and the Suppression of BET Family Proteins. International Journal of Molecular Sciences. 2023; 24(16):12874. https://doi.org/10.3390/ijms241612874
Chicago/Turabian StyleMaksimova, Varvara, Valeriia Popova, Anzhelika Prus, Evgeniya Lylova, Olga Usalka, Guzel Sagitova, Ekaterina Zhidkova, Julia Makus, Ekaterina Trapeznikova, Gennady Belitsky, and et al. 2023. "Insights into the Mechanism of Curaxin CBL0137 Epigenetic Activity: The Induction of DNA Demethylation and the Suppression of BET Family Proteins" International Journal of Molecular Sciences 24, no. 16: 12874. https://doi.org/10.3390/ijms241612874
APA StyleMaksimova, V., Popova, V., Prus, A., Lylova, E., Usalka, O., Sagitova, G., Zhidkova, E., Makus, J., Trapeznikova, E., Belitsky, G., Yakubovskaya, M., & Kirsanov, K. (2023). Insights into the Mechanism of Curaxin CBL0137 Epigenetic Activity: The Induction of DNA Demethylation and the Suppression of BET Family Proteins. International Journal of Molecular Sciences, 24(16), 12874. https://doi.org/10.3390/ijms241612874