
Molecular Design of FRET Probes Based on Domain Rearrangement of Protein Disulfide Isomerase for Monitoring Intracellular Redox Status
 , , ,
, , ,  ,
,  , and
, and 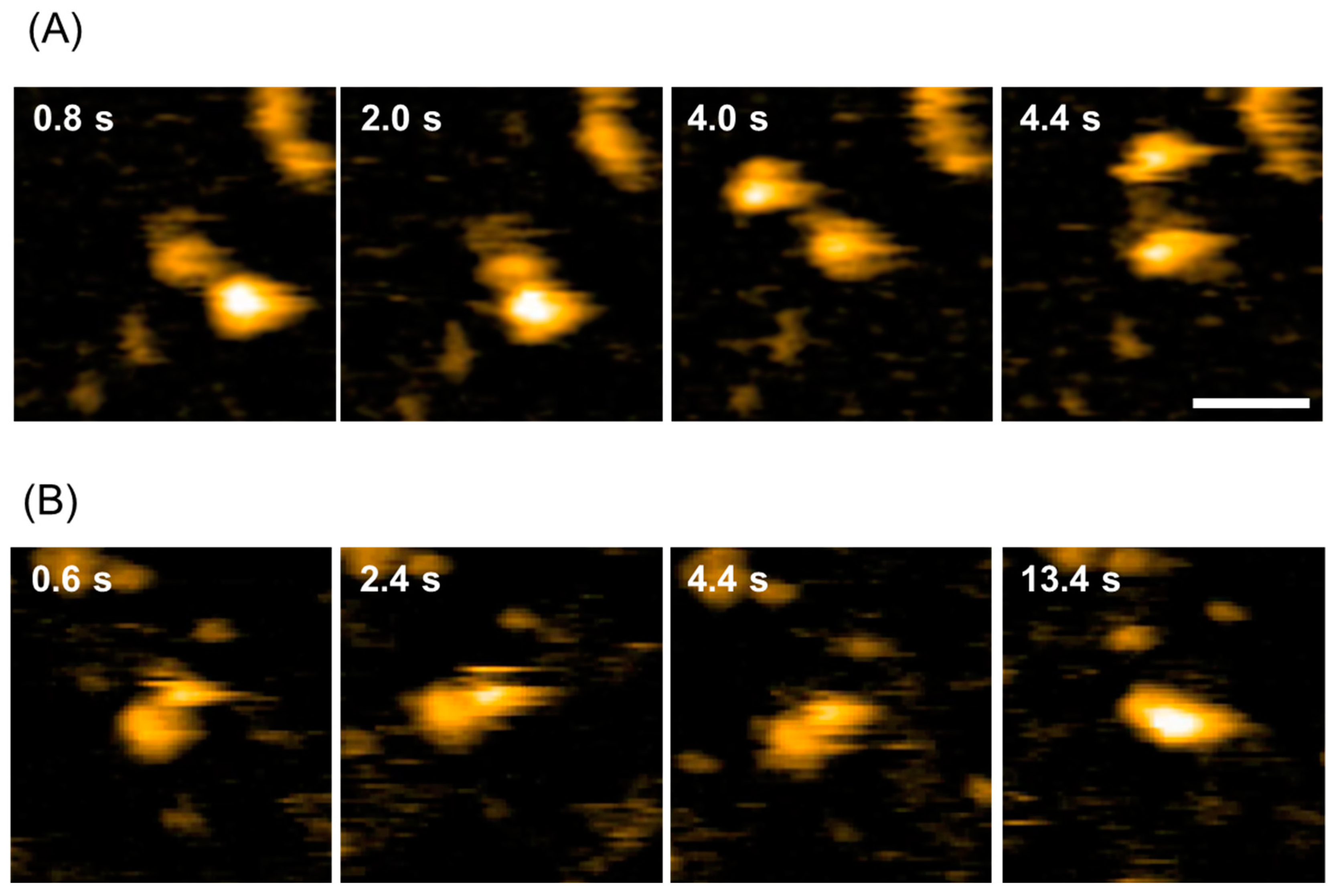{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Dynamic Domain Rearrangement of PDI-b’a’
2.2. In Vitro Characterization of PDI-b’a’-Based FRET Probes
2.3. In-Cell Characterization of PDI-b’a’-Based FRET Probes
2.4. Concluding Remarks
3. Materials and Methods
3.1. Plasmids
3.2. Bacterial Expression and Purification of Recombinant Proteins
3.3. HS-AFM
3.4. Fluorescence Measurement In Vitro
3.5. Cell Culture
3.6. FRET Imaging
3.7. CG Simulations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vogel, C.; Bashton, M.; Kerrison, N.D.; Chothia, C.; Teichmann, S.A. Structure, function and evolution of multidomain proteins. Curr. Opin. Struct. Biol. 2004, 14, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Apic, G.; Huber, W.; Teichmann, S.A. Multi-domain protein families and domain pairs: Comparison with known structures and a random model of domain recombination. J. Struct. Funct. Genom. 2003, 4, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Griesbeck, O.; Heim, R.; Tsien, R.Y. Dynamic and quantitative Ca2+ measurements using improved cameleons. Proc. Natl. Acad. Sci. USA 1999, 96, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Galligan, J.J.; Petersen, D.R. The human protein disulfide isomerase gene family. Hum. Genom. 2012, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, B.; Gilbert, H.F. Protein disulfide isomerase. Biochim. Biophys. Acta 2004, 1699, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.B.; Klappa, P.; Ruddock, L.W. Protein disulfide isomerases exploit synergy between catalytic and specific binding domains. EMBO Rep. 2002, 3, 136–140. [Google Scholar] [CrossRef]
- Serve, O.; Kamiya, Y.; Kato, K. Redox-dependent chaperoning, following PDI footsteps. In Protein Folding; Walters, E.C., Ed.; NOVA Science Publishers: New York, NY, USA, 2011; pp. 489–500. [Google Scholar]
- Inagaki, K.; Satoh, T.; Itoh, S.G.; Okumura, H.; Kato, K. Redox-dependent conformational transition of catalytic domain of protein disulfide isomerase indicated by crystal structure-based molecular dynamics simulation. Chem. Phys. Lett. 2015, 618, 203–207. [Google Scholar] [CrossRef]
- Nakasako, M.; Maeno, A.; Kurimoto, E.; Harada, T.; Yamaguchi, Y.; Oka, T.; Takayama, Y.; Iwata, A.; Kato, K. Redox-dependent domain rearrangement of protein disulfide isomerase from a thermophilic fungus. Biochemistry 2010, 49, 6953–6962. [Google Scholar] [CrossRef]
- Nguyen, V.D.; Wallis, K.; Howard, M.J.; Haapalainen, A.M.; Salo, K.E.; Saaranen, M.J.; Sidhu, A.; Wierenga, R.K.; Freedman, R.B.; Ruddock, L.W.; et al. Alternative conformations of the x region of human protein disulphide-isomerase modulate exposure of the substrate binding b’ domain. J. Mol. Biol. 2008, 383, 1144–1155. [Google Scholar] [CrossRef]
- Serve, O.; Kamiya, Y.; Maeno, A.; Nakano, M.; Murakami, C.; Sasakawa, H.; Yamaguchi, Y.; Harada, T.; Kurimoto, E.; Yagi-Utsumi, M.; et al. Redox-dependent domain rearrangement of protein disulfide isomerase coupled with exposure of its substrate-binding hydrophobic surface. J. Mol. Biol. 2010, 396, 361–374. [Google Scholar] [CrossRef]
- Tian, G.; Xiang, S.; Noiva, R.; Lennarz, W.J.; Schindelin, H. The crystal structure of yeast protein disulfide isomerase suggests cooperativity between its active sites. Cell 2006, 124, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Ren, J.; Fang, J.; Ke, H.; Gong, W.; Feng, W.; Wang, C.C. Structural insights into the redox-regulated dynamic conformations of human protein disulfide isomerase. Antioxid. Redox Signal. 2013, 19, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yu, J.; Huo, L.; Wang, L.; Feng, W.; Wang, C.C. Human protein-disulfide isomerase is a redox-regulated chaperone activated by oxidation of domain a’. J. Biol. Chem. 2012, 287, 1139–1149. [Google Scholar] [CrossRef]
- Yagi-Utsumi, M.; Satoh, T.; Kato, K. Structural basis of redox-dependent substrate binding of protein disulfide isomerase. Sci. Rep. 2015, 5, 13909. [Google Scholar] [CrossRef]
- Yang, S.; Wang, X.; Cui, L.; Ding, X.; Niu, L.; Yang, F.; Wang, C.; Wang, C.C.; Lou, J. Compact conformations of human protein disulfide isomerase. PLoS ONE 2014, 9, e103472. [Google Scholar] [CrossRef] [PubMed]
- Karamzadeh, R.; Karimi-Jafari, M.H.; Sharifi-Zarchi, A.; Chitsaz, H.; Salekdeh, G.H.; Moosavi-Movahedi, A.A. Machine Learning and Network Analysis of Molecular Dynamics Trajectories Reveal Two Chains of Red/Ox-specific Residue Interactions in Human Protein Disulfide Isomerase. Sci. Rep. 2017, 7, 3666. [Google Scholar] [CrossRef]
- Okumura, M.; Noi, K.; Kanemura, S.; Kinoshita, M.; Saio, T.; Inoue, Y.; Hikima, T.; Akiyama, S.; Ogura, T.; Inaba, K. Dynamic assembly of protein disulfide isomerase in catalysis of oxidative folding. Nat. Chem. Biol. 2019, 15, 499–509. [Google Scholar] [CrossRef]
- Inagaki, K.; Satoh, T.; Yagi-Utsumi, M.; Le Gulluche, A.C.; Anzai, T.; Uekusa, Y.; Kamiya, Y.; Kato, K. Redox-coupled structural changes of the catalytic a’ domain of protein disulfide isomerase. FEBS Lett. 2015, 589 Pt B, 2690–2694. [Google Scholar] [CrossRef]
- Chinnaraj, M.; Barrios, D.A.; Frieden, C.; Heyduk, T.; Flaumenhaft, R.; Pozzi, N. Bioorthogonal Chemistry Enables Single-Molecule FRET Measurements of Catalytically Active Protein Disulfide Isomerase. ChemBioChem 2021, 22, 134–138. [Google Scholar] [CrossRef]
- Chinnaraj, M.; Flaumenhaft, R.; Pozzi, N. Reduction of protein disulfide isomerase results in open conformations and stimulates dynamic exchange between structural ensembles. J. Biol. Chem. 2022, 298, 102217. [Google Scholar] [CrossRef]
- Oktyabrsky, O.N.; Smirnova, G.V. Redox regulation of cellular functions. Biochemistry 2007, 72, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Lennicke, C.; Cocheme, H.M. Redox metabolism: ROS as specific molecular regulators of cell signaling and function. Mol. Cell 2021, 81, 3691–3707. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sanchez-Perez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Abraham, B.G.; Santala, V.; Tkachenko, N.V.; Karp, M. Fluorescent protein-based FRET sensor for intracellular monitoring of redox status in bacteria at single cell level. Anal. Bioanal. Chem. 2014, 406, 7195–7204. [Google Scholar] [CrossRef] [PubMed]
- Hanson, G.T.; Aggeler, R.; Oglesbee, D.; Cannon, M.; Capaldi, R.A.; Tsien, R.Y.; Remington, S.J. Investigating mitochondrial redox potential with redox-sensitive green fluorescent protein indicators. J. Biol. Chem. 2004, 279, 13044–13053. [Google Scholar] [CrossRef]
- Østergaard, H.; Tachibana, C.; Winther, J.R. Monitoring disulfide bond formation in the eukaryotic cytosol. J. Cell Biol. 2004, 166, 337–345. [Google Scholar] [CrossRef]
- Sugiura, K.; Nagai, T.; Nakano, M.; Ichinose, H.; Nakabayashi, T.; Ohta, N.; Hisabori, T. Redox sensor proteins for highly sensitive direct imaging of intracellular redox state. Biochem. Biophys. Res. Commun. 2015, 457, 242–248. [Google Scholar] [CrossRef]
- Sugiura, K.; Yokochi, Y.; Fu, N.; Fukaya, Y.; Yoshida, K.; Mihara, S.; Hisabori, T. The thioredoxin (Trx) redox state sensor protein can visualize Trx activities in the light/dark response in chloroplasts. J. Biol. Chem. 2019, 294, 12091–12098. [Google Scholar] [CrossRef]
- Yano, T.; Oku, M.; Akeyama, N.; Itoyama, A.; Yurimoto, H.; Kuge, S.; Fujiki, Y.; Sakai, Y. A novel fluorescent sensor protein for visualization of redox states in the cytoplasm and in peroxisomes. Mol. Cell Biol. 2010, 30, 3758–3766. [Google Scholar] [CrossRef]
- Hiranyakorn, M.; Yagi-Utsumi, M.; Yanaka, S.; Ohtsuka, N.; Momiyama, N.; Satoh, T.; Kato, K. Mutational and Environmental Effects on the Dynamic Conformational Distributions of Lys48-Linked Ubiquitin Chains. Int. J. Mol. Sci. 2023, 24, 6075. [Google Scholar] [CrossRef]
- Seetaha, S.; Yagi-Utsumi, M.; Yamaguchi, T.; Ishii, K.; Hannongbua, S.; Choowongkomon, K.; Kato, K. Application of Site-Specific Spin Labeling for NMR Detecting Inhibitor-Induced Conformational Change of HIV-1 Reverse Transcriptase. ChemMedChem 2016, 11, 363–366. [Google Scholar] [CrossRef]
- Gobl, C.; Madl, T.; Simon, B.; Sattler, M. NMR approaches for structural analysis of multidomain proteins and complexes in solution. Prog. Nucl. Magn. Reson. Spectrosc. 2014, 80, 26–63. [Google Scholar] [CrossRef]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar]
- Nguyen, A.W.; Daugherty, P.S. Evolutionary optimization of fluorescent proteins for intracellular FRET. Nat. Biotechnol. 2005, 23, 355–360. [Google Scholar] [CrossRef]
- Nagai, T.; Yamada, S.; Tominaga, T.; Ichikawa, M.; Miyawaki, A. Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 10554–10559. [Google Scholar] [CrossRef]
- Goedhart, J.; van Weeren, L.; Hink, M.A.; Vischer, N.O.; Jalink, K.; Gadella, T.W., Jr. Bright cyan fluorescent protein variants identified by fluorescence lifetime screening. Nat. Methods 2010, 7, 137–139. [Google Scholar] [CrossRef]
- Ando, T.; Uchihashi, T.; Kodera, N.; Yamamoto, D.; Miyagi, A.; Taniguchi, M.; Yamashita, H. High-speed AFM and nano-visualization of biomolecular processes. Pflügers Arch.-Eur. J. Physiol. 2008, 456, 211–225. [Google Scholar] [CrossRef]
- Rekas, A.; Alattia, J.R.; Nagai, T.; Miyawaki, A.; Ikura, M. Crystal structure of venus, a yellow fluorescent protein with improved maturation and reduced environmental sensitivity. J. Biol. Chem. 2002, 277, 50573–50578. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef]
- Kenzaki, H.; Koga, N.; Hori, N.; Kanada, R.; Li, W.; Okazaki, K.; Yao, X.Q.; Takada, S. CafeMol: A Coarse-Grained Biomolecular Simulator for Simulating Proteins at Work. J. Chem. Theory Comput. 2011, 7, 1979–1989. [Google Scholar] [CrossRef]
- Li, W.; Terakawa, T.; Wang, W.; Takada, S. Energy landscape and multiroute folding of topologically complex proteins adenylate kinase and 2ouf-knot. Proc. Natl. Acad. Sci. USA 2012, 109, 17789–17794. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagi-Utsumi, M.; Miura, H.; Ganser, C.; Watanabe, H.; Hiranyakorn, M.; Satoh, T.; Uchihashi, T.; Kato, K.; Okazaki, K.-i.; Aoki, K. Molecular Design of FRET Probes Based on Domain Rearrangement of Protein Disulfide Isomerase for Monitoring Intracellular Redox Status. Int. J. Mol. Sci. 2023, 24, 12865. https://doi.org/10.3390/ijms241612865
Yagi-Utsumi M, Miura H, Ganser C, Watanabe H, Hiranyakorn M, Satoh T, Uchihashi T, Kato K, Okazaki K-i, Aoki K. Molecular Design of FRET Probes Based on Domain Rearrangement of Protein Disulfide Isomerase for Monitoring Intracellular Redox Status. International Journal of Molecular Sciences. 2023; 24(16):12865. https://doi.org/10.3390/ijms241612865
Chicago/Turabian StyleYagi-Utsumi, Maho, Haruko Miura, Christian Ganser, Hiroki Watanabe, Methanee Hiranyakorn, Tadashi Satoh, Takayuki Uchihashi, Koichi Kato, Kei-ichi Okazaki, and Kazuhiro Aoki. 2023. "Molecular Design of FRET Probes Based on Domain Rearrangement of Protein Disulfide Isomerase for Monitoring Intracellular Redox Status" International Journal of Molecular Sciences 24, no. 16: 12865. https://doi.org/10.3390/ijms241612865
APA StyleYagi-Utsumi, M., Miura, H., Ganser, C., Watanabe, H., Hiranyakorn, M., Satoh, T., Uchihashi, T., Kato, K., Okazaki, K.-i., & Aoki, K. (2023). Molecular Design of FRET Probes Based on Domain Rearrangement of Protein Disulfide Isomerase for Monitoring Intracellular Redox Status. International Journal of Molecular Sciences, 24(16), 12865. https://doi.org/10.3390/ijms241612865