
Isolation and Identification of a Novel Anti-Dry Eye Peptide from Tilapia Skin Peptides Based on In Silico, In Vitro, and In Vivo Approaches


Abstract
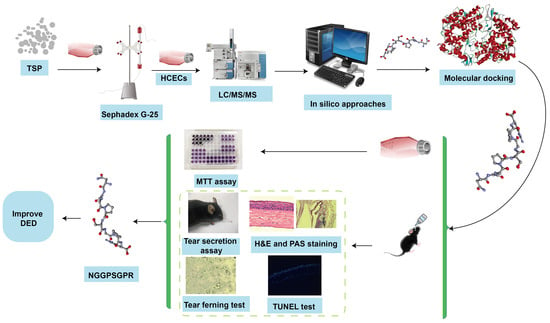
1. Introduction
2. Results and Discussion
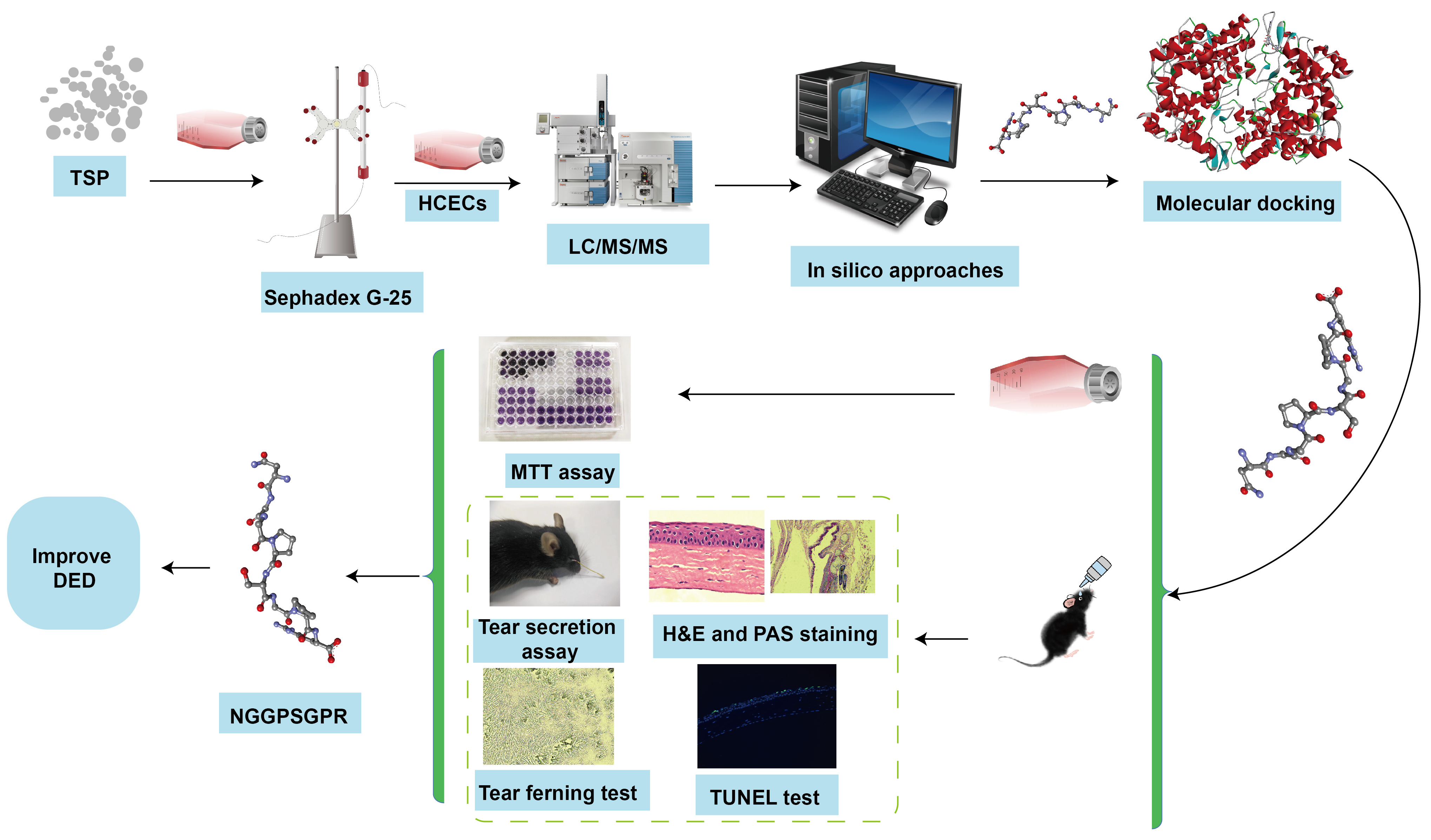
2.1. Effects of TSP on Cell Viability of HCECs
2.2. Identification of Peptides from TSP- II via LC/MS/MS
2.3. Screening of Bioactive Peptides via ToxinPred and PeptideRanker Web Servers
2.4. Screening of AIPs and Allergenicity Prediction
2.5. Screening Anti-DED Peptides by Molecule Docking with COX-2
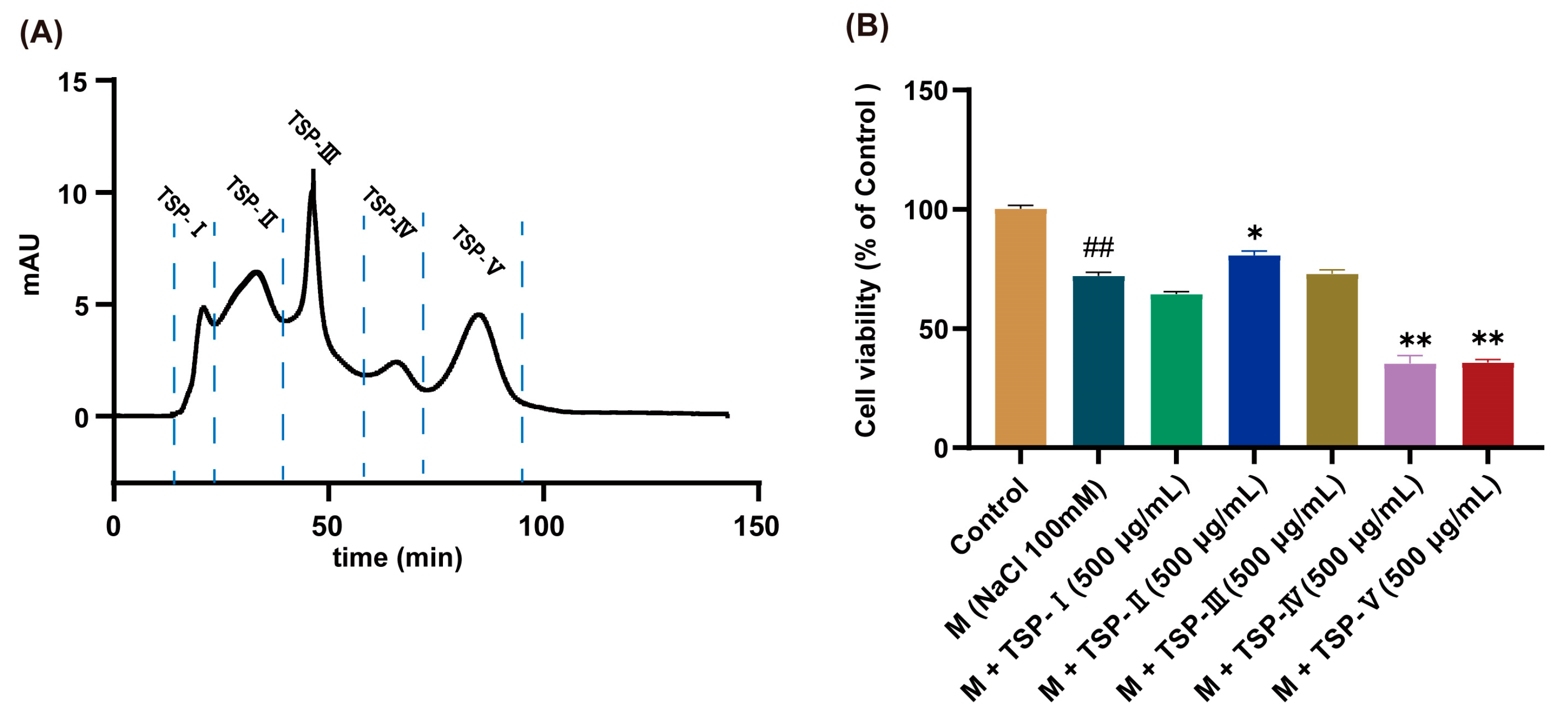
2.6. Assay of In Vitro Anti-DED Activity of NGG
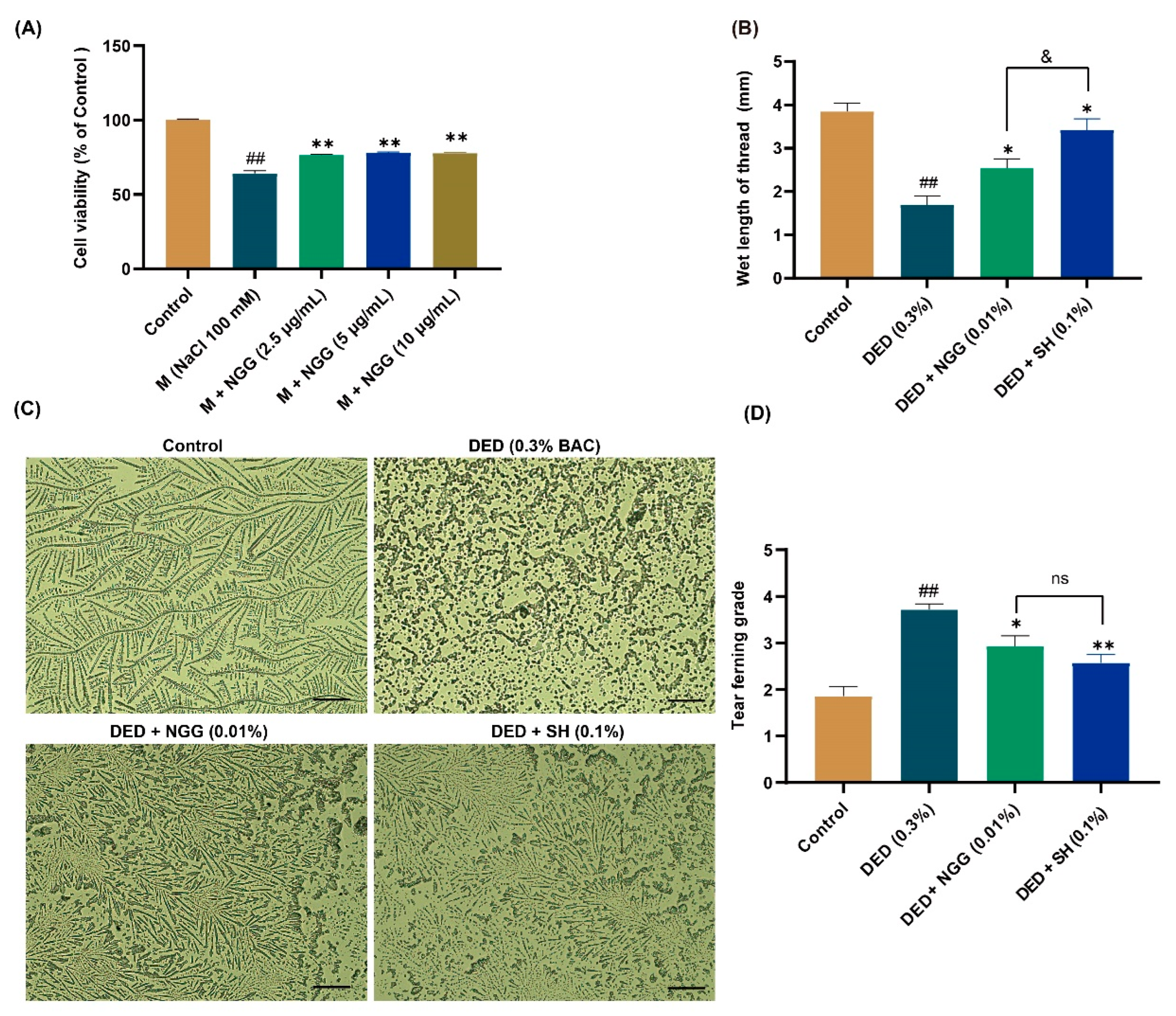
2.7. Effects of NGG on Tear Production and Tear Ferning in DED Mice
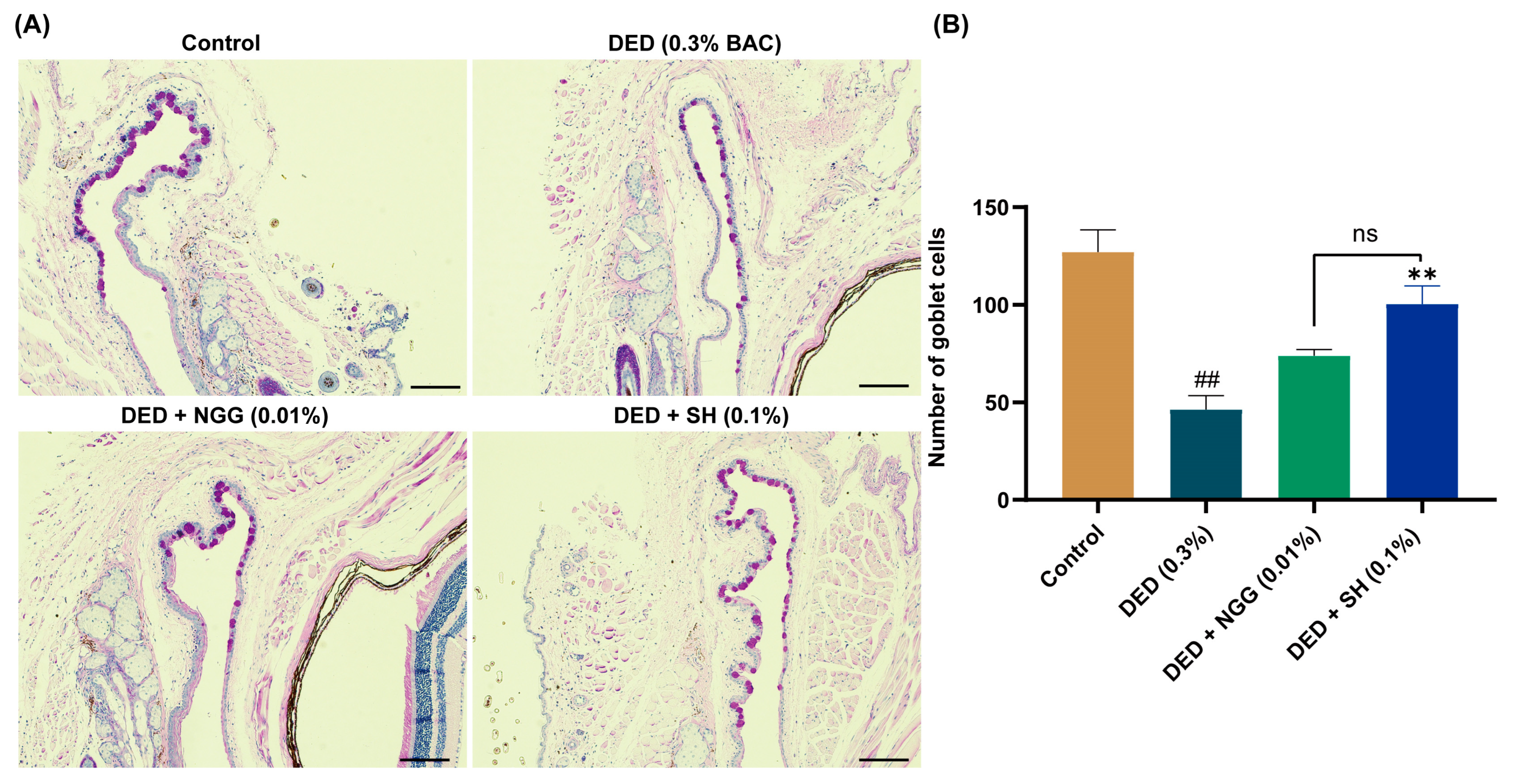
2.8. Effects of NGG on Conjunctival Goblet Cells (CGCs) in DED Mice
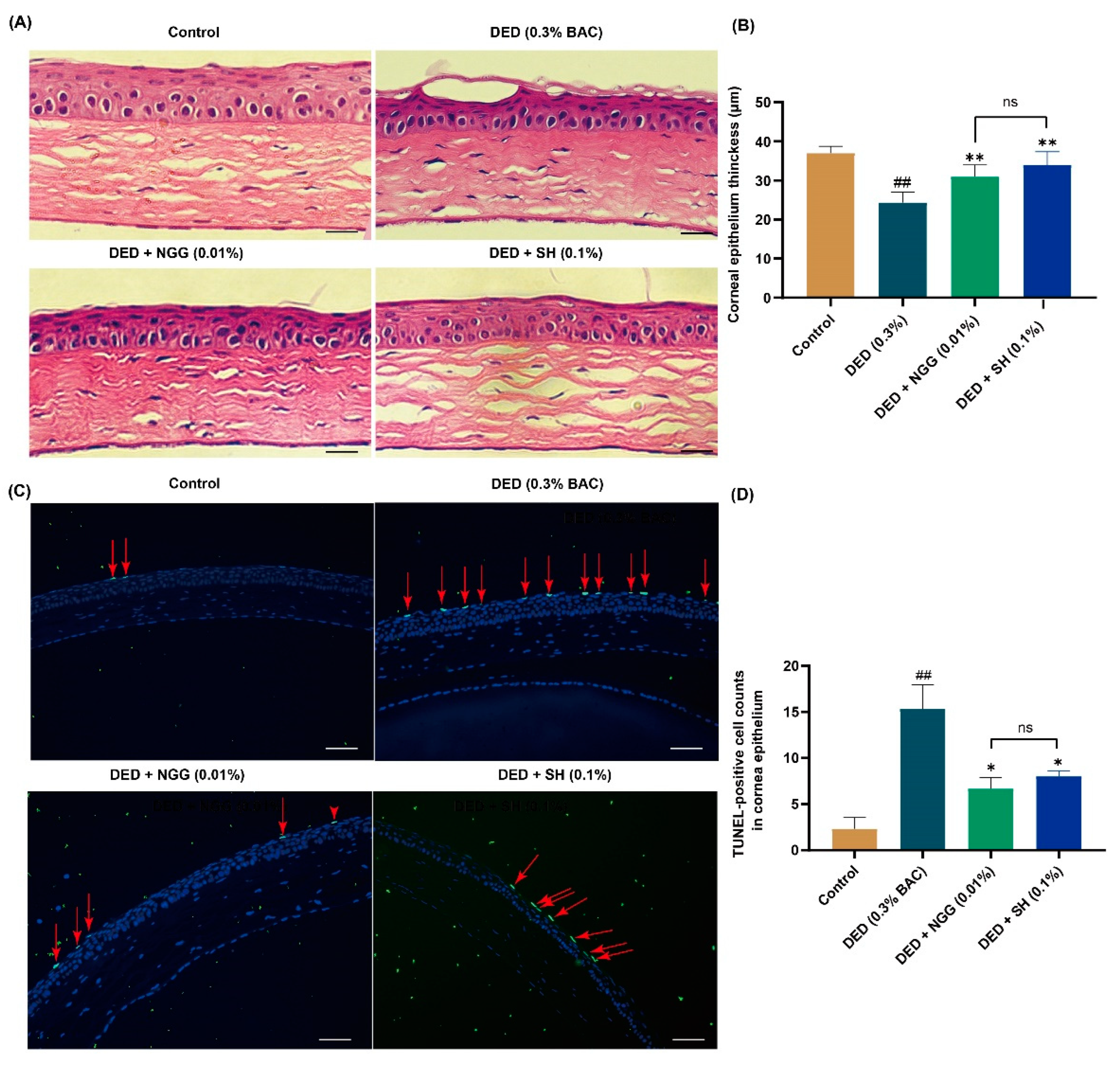
2.9. Effects of NGG on Corneal Epithelium in DED Mice
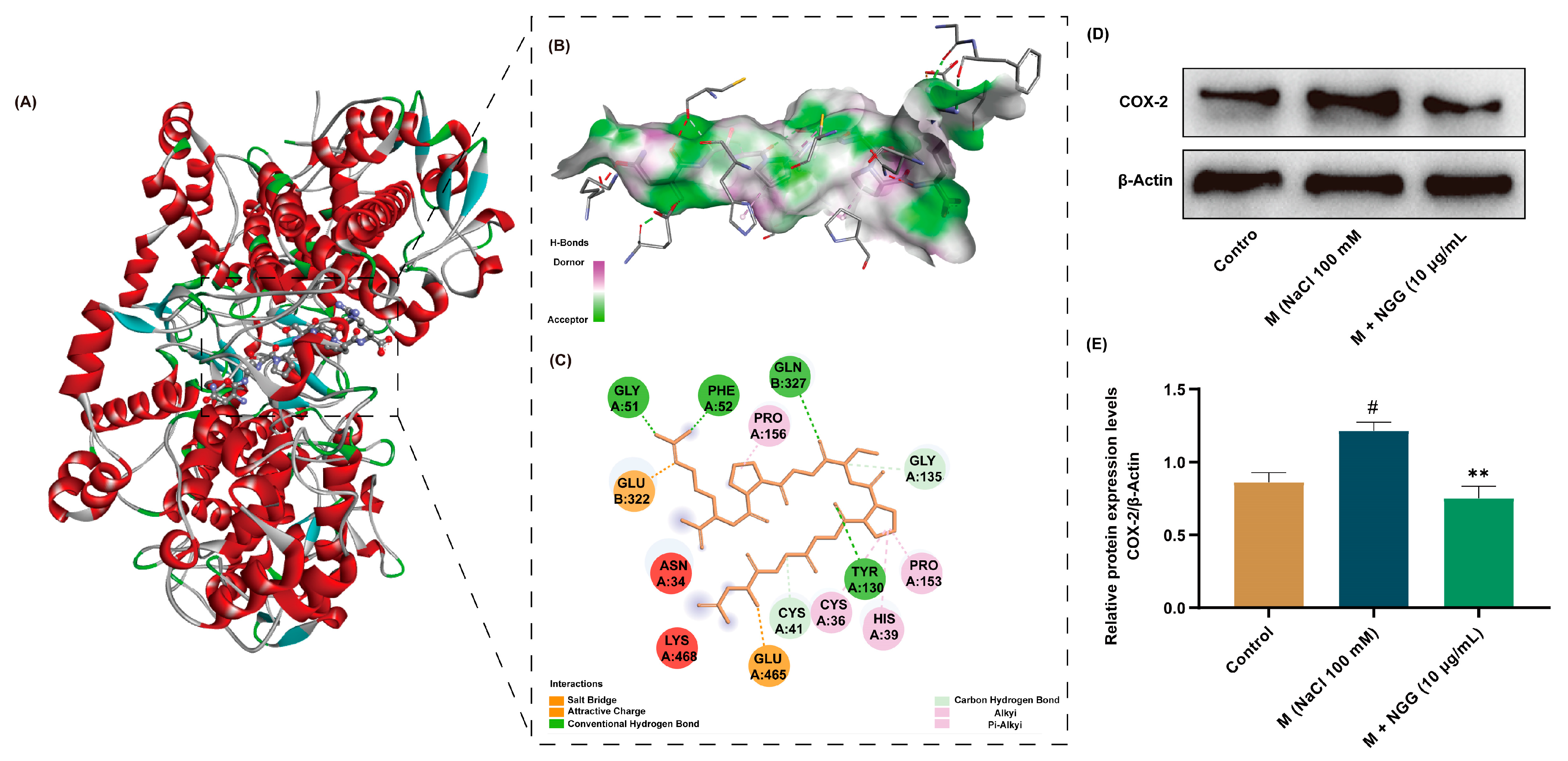
2.10. Analysis of COX-2–NGG Interactions and Effects of NGG on COX-2 Protein Expression
3. Materials and Methods
3.1. Materials
3.2. Preparation of TSP
3.3. Isolation of TSP and Screening of Its Active Fractions
3.4. Peptides Sequence Identification of TSP via LC/MS/MS
3.5. Screening the Bioactive Peptides via In Silico Methods
3.6. Molecular Docking and Peptide Chemical Synthesis
3.7. Cell Culture and Treatment
3.8. Animals and Treatment
3.9. Tear Production and Tear Ferning Test
3.10. Tissue Preparation for Histological Assessment
3.11. H&E Staining and PAS Staining Analysis
3.12. In Situ TUNEL Staining
3.13. Western Blot
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, Z.; Liu, T.; Zhou, X.; Yang, Y.; Liu, Y.; Zhou, H.; Wei, S.; Zhai, Z.; Wu, Y.; Sun, F.; et al. Rapid and quantitative detection of tear MMP-9 for dry eye patients using a novel silicon nanowire-based biosensor. Biosens. Bioelectron. 2022, 214, 114498. [Google Scholar] [CrossRef]
- Kawashima, M. Systemic Health and Dry Eye. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES138–DES142. [Google Scholar] [CrossRef]
- Garcia-Marques, J.V.; Talens-Estarelles, C.; Garcia-Lazaro, S.; Wolffsohn, J.S.; Cervino, A. Systemic, environmental and lifestyle risk factors for dry eye disease in a mediterranean caucasian population. Contact Lens Anterior Eye 2022, 45, 101539. [Google Scholar] [CrossRef]
- Tellefsen Noland, S.; Badian, R.A.; Utheim, T.P.; Utheim, O.A.; Stojanovic, A.; Tashbayev, B.; Raeder, S.; Dartt, D.A.; Chen, X. Sex and age differences in symptoms and signs of dry eye disease in a Norwegian cohort of patients. Ocul. Surf. 2021, 19, 68–73. [Google Scholar] [CrossRef]
- Garcia-Alfaro, P.; Garcia, S.; Rodriguez, I.; Verges, C. Dry eye disease symptoms and quality of life in perimenopausal and postmenopausal women. Climacteric 2021, 24, 261–266. [Google Scholar] [CrossRef]
- Muntz, A.; Turnbull, P.R.; Kim, A.D.; Gokul, A.; Wong, D.; Tsay, T.S.; Zhao, K.; Zhang, S.; Kingsnorth, A.; Wolffsohn, J.S.; et al. Extended screen time and dry eye in youth. Contact Lens Anterior Eye 2022, 45, 101541. [Google Scholar] [CrossRef]
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.J.; Gundersen, K.G.; Tong, L.; Utheim, T.P. Dry eye disease and proteomics. Ocul. Surf. 2022, 24, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Gong, Q.; Lu, B.; Huang, K.; Tong, Y.; Mutsvene, T.E.; Lin, M.; Xu, Z.; Lu, F.; Li, X.; et al. Anti-inflammatory and antioxidative effects of gallic acid on experimental dry eye: In vitro and in vivo studies. Eye Vis. 2023, 10, 17. [Google Scholar] [CrossRef]
- Li, S.; Lu, Z.Y.; Huang, Y.; Wang, Y.; Jin, Q.; Shentu, X.C.; Ye, J.; Ji, J.; Yao, K.; Han, H.J. Anti-Oxidative and Anti-Inflammatory Micelles: Break the Dry Eye Vicious Cycle. Adv. Sci. 2022, 9, e2200435. [Google Scholar] [CrossRef] [PubMed]
- Foulks, G.N.; Forstot, S.L.; Donshik, P.C.; Forstot, J.Z.; Goldstein, M.H.; Lemp, M.A.; Nelson, J.D.; Nichols, K.K.; Pflugfelder, S.C.; Tanzer, J.M.; et al. Clinical Guidelines for Management of Dry Eye Associated with Sjogren Disease. Ocul. Surf. 2015, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Kuklinski, E.; Asbell, P.A. Sjogren’s syndrome from the perspective of ophthalmology. Clin. Immunol. 2017, 182, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Wang, L.; Liu, C.; Liang, Y.; Lin, Q. Bioactive peptides from foods: Production, function, and application. Food Funct. 2021, 12, 7108–7125. [Google Scholar] [CrossRef] [PubMed]
- Zhi, T.; Hong, D.; Zhang, Z.; Li, S.; Xia, J.; Wang, C.; Wu, Y.; Jia, Y.; Ma, A. Anti-inflammatory and gut microbiota regulatory effects of walnut protein derived peptide LPF in vivo. Food Res. Int. 2022, 152, 110875. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Yuan, Q.; Xie, H.; Shi, J.; Ju, X. Separation and purification of an anti-tumor peptide from rapeseed (Brassica campestris L.) and the effect on cell apoptosis. Food Funct. 2016, 7, 2239–2248. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, S.; Wang, Q.; Yuan, Q.; Li, Y.; Reboredo-Rodriguez, P.; Varela-Lopez, A.; He, Z.; Wu, F.; Hu, H.; et al. Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake. Int. J. Mol. Sci. 2022, 23, 12086. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Fan, N.W.; Yeh, S.I.; Chen, S.L.; Tsao, Y.P. The Therapeutic Effects of a PEDF-Derived Short Peptide on Murine Experimental Dry Eye Involves Suppression of MMP-9 and Inflammation. Transl. Vis. Sci. Technol. 2022, 11, 12. [Google Scholar] [CrossRef]
- Kim, D.U.; Chung, H.C.; Choi, J.; Sakai, Y.; Lee, B.Y. Oral Intake of Low-Molecular-Weight Collagen Peptide Improves Hydration, Elasticity, and Wrinkling in Human Skin: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2018, 10, 826. [Google Scholar] [CrossRef]
- Song, W.K.; Liu, D.; Sun, L.L.; Li, B.F.; Hou, H. Physicochemical and Biocompatibility Properties of Type I Collagen from the Skin of Nile Tilapia (Oreochromis niloticus) for Biomedical Applications. Mar. Drugs 2019, 17, 137. [Google Scholar] [CrossRef]
- Li, D.D.; Li, W.J.; Kong, S.Z.; Li, S.D.; Guo, J.Q.; Guo, M.H.; Cai, T.T.; Li, N.; Chen, R.Z.; Luo, R.Q.; et al. Protective effects of collagen polypeptide from tilapia skin against injuries to the liver and kidneys of mice induced by d-galactose. Biomed. Pharmacother. 2019, 117, 109204. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Yin, H.; Hu, C.; Zeng, J.; Zhang, S.; Chen, S.; Zheng, W.; Li, M.; Jin, L.; Liu, Y.; et al. Tilapia Skin Peptides Ameliorate Cyclophosphamide-Induced Anxiety- and Depression-Like Behavior via Improving Oxidative Stress, Neuroinflammation, Neuron Apoptosis, and Neurogenesis in Mice. Front. Nutr. 2022, 9, 882175. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Yin, H.; Hu, C.; Zeng, J.; Shi, X.; Chen, S.; Zhang, K.; Zheng, W.; Wu, W.; Liu, S. Tilapia skin peptides restore cyclophosphamide-induced premature ovarian failure via inhibiting oxidative stress and apoptosis in mice. Food Funct. 2022, 13, 1668–1679. [Google Scholar] [CrossRef]
- Zeng, J.; Hu, C.; Lin, C.; Zhang, S.; Deng, K.; Du, J.; Yang, Z.; Liu, S.; Wu, W.; Zhao, Y.-T. Tilapia Skin Peptides Inhibit Apoptosis, Inflammation, and Oxidative Stress to Improve Dry Eye Disease In Vitro and In Vivo. J. Food Biochem. 2023, 2023, 6761792. [Google Scholar] [CrossRef]
- Wenhui, T.; Shumin, H.; Yongliang, Z.; Liping, S.; Hua, Y. Identification of in vitro angiotensin-converting enzyme and dipeptidyl peptidase IV inhibitory peptides from draft beer by virtual screening and molecular docking. J. Sci. Food Agric. 2022, 102, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P.; Pliszka, M.; Mogut, D.; Darewicz, M. Characteristics of Biopeptides Released In Silico from Collagens Using Quantitative Parameters. Foods 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Jiang, X.; Chou, Y.; Wei, S.; Hao, R.; Su, J.; Li, X. In vivo Confocal Microscopic Evaluation of Previously Neglected Oval Cells in Corneal Nerve Vortex: An Inflammatory Indicator of Dry Eye Disease. Front. Med. 2022, 9, 906219. [Google Scholar] [CrossRef]
- Manavalan, B.; Shin, T.H.; Kim, M.O.; Lee, G. AIPpred: Sequence-Based Prediction of Anti-inflammatory Peptides Using Random Forest. Front. Pharmacol. 2018, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Wongsrangsap, N.; Chukiatsiri, S. Purification and Identification of Novel Antioxidant Peptides from Enzymatically Hydrolysed Samia ricini Pupae. Molecules 2021, 26, 2588. [Google Scholar] [CrossRef]
- Tu, M.; Liu, H.; Cheng, S.; Mao, F.; Chen, H.; Fan, F.; Lu, W.; Du, M. Identification and characterization of a novel casein anticoagulant peptide derived from in vivo digestion. Food Funct. 2019, 10, 2552–2559. [Google Scholar] [CrossRef]
- Patel, B.Y.; Volcheck, G.W. Food Allergy: Common Causes, Diagnosis, and Treatment. Mayo Clin. Proc. 2015, 90, 1411–1419. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef]
- Fang, Y.; Pan, X.; Zhao, E.; Shi, Y.; Shen, X.; Wu, J.; Pei, F.; Hu, Q.; Qiu, W. Isolation and identification of immunomodulatory selenium-containing peptides from selenium-enriched rice protein hydrolysates. Food Chem. 2019, 275, 696–702. [Google Scholar] [CrossRef]
- Vo, T.S.; Kim, Y.S.; Ngo, D.H.; Le, P.U.; Kim, S.Y.; Kim, S.K. Spirulina maxima peptides suppress mast cell degranulation via inactivating Akt and MAPKs phosphorylation in RBL-2H3 cells. Int. J. Biol. Macromol. 2018, 118, 2224–2229. [Google Scholar] [CrossRef]
- Montoya-Rodríguez, A.; de Mejía, E.G.; Dia, V.P.; Reyes-Moreno, C.; Milán-Carrillo, J. Extrusion improved the anti-inflammatory effect of amaranth (Amaranthus hypochondriacus) hydrolysates in LPS-induced human THP-1 macrophage-like and mouse RAW 264.7 macrophages by preventing activation of NF-κB signaling. Mol. Nutr. Food Res. 2014, 58, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Su, M.; Hong, J.; La Kim, E.; Moon, H.R.; Chung, H.Y.; Kim, S.; Jung, J.H. Design of balanced COX inhibitors based on anti-inflammatory and/or COX-2 inhibitory ascidian metabolites. Eur. J. Med. Chem. 2019, 180, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-C.; Ha, J.-H.; Lee, J.K.; Jung, S.H.; Kim, J.-C. In Vivo Anti-Inflammation Potential of Aster koraiensis Extract for Dry Eye Syndrome by the Protection of Ocular Surface. Nutrients 2020, 12, 3245. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.W.; Seo, Y.; Choi, W.; Yeo, A.; Noh, H.; Kim, E.K.; Lee, H.K. Dry Eye-Induced CCR7+CD11b+ Cell Lymph Node Homing Is Induced by COX-2 Activities. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6829–6838. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, K.; Liu, Q.; Zhang, X. Discovery of monoamine oxidase A inhibitory peptides from hairtail (Trichiurus japonicus) using in vitro simulated gastrointestinal digestion and in silico studies. Bioorg. Chem. 2020, 101, 104032. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, S.; Yue, H.; Wang, M.; Zeng, J.; Wu, W.; Wang, J.; Zheng, H.; Xue, C.; Zhao, Y.-T. Isolation, identification and in silico analysis of two novel cytoprotective peptides from tilapia skin against oxidative stress-induced ovarian granulosa cell damage. J. Funct. Foods 2023, 107, 105629. [Google Scholar] [CrossRef]
- Willcox, M.D.P.; Argueso, P.; Georgiev, G.A.; Holopainen, J.M.; Laurie, G.W.; Millar, T.J.; Papas, E.B.; Rolland, J.P.; Schmidt, T.A.; Stahl, U.; et al. TFOS DEWS II Tear Film Report. Ocul. Surf. 2017, 15, 366–403. [Google Scholar] [CrossRef]
- Cher, I. A new look at lubrication of the ocular surface: Fluid mechanics behind the blinking eyelids. Ocul. Surf. 2008, 6, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jeyalatha, M.V.; Qu, Y.; He, X.; Ou, S.; Bu, J.; Jia, C.; Wang, J.; Wu, H.; Liu, Z.; et al. Dry Eye Management: Targeting the Ocular Surface Microenvironment. Int. J. Mol. Sci. 2017, 18, 1398. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Chen, Z.Y.; Tang, Y.J.; Tsai, C.H.; Chuang, Y.L.; Hsieh, E.H.; Tucker, L.; Lin, I.C.; Tseng, C.L. Development of Lutein-Containing Eye Drops for the Treatment of Dry Eye Syndrome. Pharmaceutics 2021, 13, 1801. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Liu, X.; Zhou, T.; Wang, Y.; Bai, L.; He, H.; Liu, Z. A mouse dry eye model induced by topical administration of benzalkonium chloride. Mol. Vis. 2011, 17, 257–264. [Google Scholar] [PubMed]
- Tang, Y.J.; Chang, H.H.; Tsai, C.Y.; Chen, L.Y.; Lin, D.P. Establishment of a Tear Ferning Test Protocol in the Mouse Model. Transl. Vis. Sci. Technol. 2020, 9, 1. [Google Scholar] [CrossRef]
- Kim, C.S.; Jo, K.; Lee, I.S.; Kim, J. Topical Application of Apricot Kernel Extract Improves Dry Eye Symptoms in a Unilateral Exorbital Lacrimal Gland Excision Mouse. Nutrients 2016, 8, 750. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, J.; Bu, J.; Tang, L.; Yang, Y.; Ouyang, W.; Lin, X.; Liu, Z.; Huang, C.; Quantock, A.J.; et al. High-fat diet induces dry eye-like ocular surface damages in murine. Ocul. Surf. 2020, 18, 267–276. [Google Scholar] [CrossRef]
- Baudouin, C.; Rolando, M.; Del Castillo, J.M.B.; Messmer, E.M.; Figueiredo, F.C.; Irkec, M.; Van Setten, G.; Labetoulle, M. Reconsidering the central role of mucins in dry eye and ocular surface diseases. Prog. Retin. Eye Res. 2019, 71, 68–87. [Google Scholar] [CrossRef]
- Alam, J.; de Paiva, C.S.; Pflugfelder, S.C. Immune–Goblet cell interaction in the conjunctiva. Ocul. Surf. 2020, 18, 326–334. [Google Scholar] [CrossRef]
- Barbosa, F.L.; Xiao, Y.; Bian, F.; Coursey, T.G.; Ko, B.Y.; Clevers, H.; de Paiva, C.S.; Pflugfelder, S.C. Goblet Cells Contribute to Ocular Surface Immune Tolerance-Implications for Dry Eye Disease. Int. J. Mol. Sci. 2017, 18, 978. [Google Scholar] [CrossRef]
- Li, J.; Tan, G.; Ding, X.Y.; Wang, Y.H.; Wu, A.H.; Yang, Q.C.; Ye, L.; Shao, Y. A mouse dry eye model induced by topical administration of the air pollutant particulate matter 10. Biomed. Pharmacother. 2017, 96, 524–534. [Google Scholar] [CrossRef]
- Shih, Y.H.; Chen, F.A.; Wang, L.F.; Hsu, J.L. Discovery and Study of Novel Antihypertensive Peptides Derived from Cassia obtusifolia Seeds. J. Agric. Food Chem. 2019, 67, 7810–7820. [Google Scholar] [CrossRef]
- Zhong, C.; Sun, L.C.; Yan, L.J.; Lin, Y.C.; Liu, G.M.; Cao, M.J. Production, optimisation and characterisation of angiotensin converting enzyme inhibitory peptides from sea cucumber (Stichopus japonicus) gonad. Food Funct. 2018, 9, 594–603. [Google Scholar] [CrossRef]
- Park, B.; Lee, I.S.; Hyun, S.W.; Jo, K.; Lee, T.G.; Kim, J.S.; Kim, C.S. The Protective Effect of Polygonum cuspidatum (PCE) Aqueous Extract in a Dry Eye Model. Nutrients 2018, 10, 1550. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S.; Discovery, O.S.D. In Silico Approach for Predicting Toxicity of Peptides and Proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, I.; Flower, D.R.; Doytchinova, I. AllerTOP—A server for in silico prediction of allergens. BMC Bioinform. 2013, 14 (Suppl. S6), 1–9. [Google Scholar] [CrossRef]
- Xiao, C.; Zhou, L.; Gao, J.; Jia, R.; Zheng, Y.; Zhao, S.; Zhao, M.; Toldra, F. Musculus senhousei as a promising source of bioactive peptides protecting against alcohol-induced liver injury. Food Chem. Toxicol. 2023, 174, 113652. [Google Scholar] [CrossRef]
- Ying, F.; Lin, S.Q.; Li, J.Y.; Zhang, X.W.; Chen, G. Identification of monoamine oxidases inhibitory peptides from soybean protein hydrolysate through ultrafiltration purification and in silico studies. Food Biosci. 2021, 44, 101355. [Google Scholar] [CrossRef]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef] [PubMed]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chao, H.; Chen, L.; Craig, P.A.; Crichlow, G.V.; Dalenberg, K.; Duarte, J.M.; et al. RCSB Protein Data Bank (RCSB.org): Delivery of experimentally-determined PDB structures alongside one million computed structure models of proteins from artificial intelligence/machine learning. Nucleic Acids Res. 2023, 51, D488–D508. [Google Scholar] [CrossRef]
- Qu, M.; Qi, X.; Wang, Q.; Wan, L.; Li, J.; Li, W.; Li, Y.; Zhou, Q. Therapeutic Effects of STAT3 Inhibition on Experimental Murine Dry Eye. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3776–3785. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Peptide | Length | HPEPDOCK Score | Allergenicity | Uniprot | BIOPEP-UWM |
|---|---|---|---|---|---|---|
| 1 | NGGPSGPR | 8 | −227.219 | No | No | No |
| 2 | DFCPPGFNTK | 10 | −189.871 | No | No | No |
| 3 | EAPDPLRN | 8 | −139.07 | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, J.; Lin, C.; Zhang, S.; Yin, H.; Deng, K.; Yang, Z.; Zhang, Y.; Liu, Y.; Hu, C.; Zhao, Y.-T. Isolation and Identification of a Novel Anti-Dry Eye Peptide from Tilapia Skin Peptides Based on In Silico, In Vitro, and In Vivo Approaches. Int. J. Mol. Sci. 2023, 24, 12772. https://doi.org/10.3390/ijms241612772
Zeng J, Lin C, Zhang S, Yin H, Deng K, Yang Z, Zhang Y, Liu Y, Hu C, Zhao Y-T. Isolation and Identification of a Novel Anti-Dry Eye Peptide from Tilapia Skin Peptides Based on In Silico, In Vitro, and In Vivo Approaches. International Journal of Molecular Sciences. 2023; 24(16):12772. https://doi.org/10.3390/ijms241612772
Chicago/Turabian StyleZeng, Jian, Cuixian Lin, Shilin Zhang, Haowen Yin, Kaishu Deng, Zhiyou Yang, Yongping Zhang, You Liu, Chuanyin Hu, and Yun-Tao Zhao. 2023. "Isolation and Identification of a Novel Anti-Dry Eye Peptide from Tilapia Skin Peptides Based on In Silico, In Vitro, and In Vivo Approaches" International Journal of Molecular Sciences 24, no. 16: 12772. https://doi.org/10.3390/ijms241612772
APA StyleZeng, J., Lin, C., Zhang, S., Yin, H., Deng, K., Yang, Z., Zhang, Y., Liu, Y., Hu, C., & Zhao, Y.-T. (2023). Isolation and Identification of a Novel Anti-Dry Eye Peptide from Tilapia Skin Peptides Based on In Silico, In Vitro, and In Vivo Approaches. International Journal of Molecular Sciences, 24(16), 12772. https://doi.org/10.3390/ijms241612772