

Crystal Structure of the Catalytic Domain of a Botulinum Neurotoxin Homologue from Enterococcus faecium: Potential Insights into Substrate Recognition
 , , , and
, , , and
Abstract
1. Introduction
2. Results and Discussion
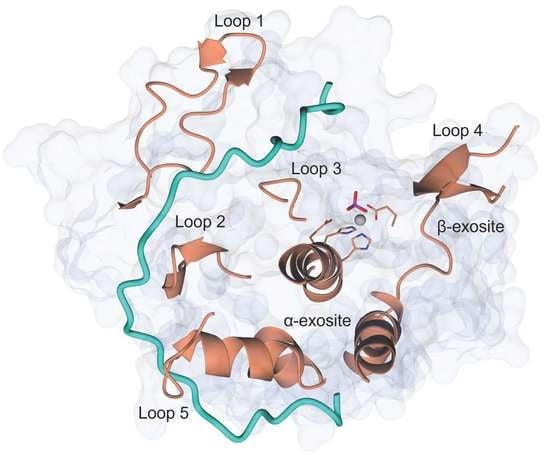
2.1. Crystal Structure of LC/En
2.2. The Active Site and Subsites
2.3. Putative Substrate Binding Exosites
2.4. The α-Exosite
2.5. LC/En Undergoes Autoproteolysis
3. Materials and Methods
3.1. Cloning, Expression, and Purification of LC/En
3.2. Crystallisation and Structure Determination
3.3. Trypsin-Digested Electrospray Ionisation Mass Spectrometry (ESI-MS) of LC/En
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, B. The Destructive Effect of Botulinum Neurotoxins on the SNARE Protein: SNAP-25 and Synaptic Membrane Fusion. PeerJ 2015, 3, e1065. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Stenmark, P. The Structure and Classification of Botulinum Toxins. Botulinum Toxin Ther. 2021, 263, 11–33. [Google Scholar] [CrossRef]
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and Tetanus Neurotoxins. Annu. Rev. Biochem. 2019, 88, 811–837. [Google Scholar] [CrossRef]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum Neurotoxins: Biology, Pharmacology, and Toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.S.; Acharya, K.R. A Comprehensive Structural Analysis of Clostridium Botulinum Neurotoxin A Cell-Binding Domain from Different Subtypes. Toxins 2023, 15, 92. [Google Scholar] [CrossRef]
- Gardner, A.P.; Barbieri, J.T. Light Chain Diversity among the Botulinum Neurotoxins. Toxins 2018, 10, 268. [Google Scholar] [CrossRef]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal Structure of Botulinum Neurotoxin Type A and Implications for Toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef]
- Kumaran, D.; Eswaramoorthy, S.; Furey, W.; Navaza, J.; Sax, M.; Swaminathan, S. Domain Organization in Clostridium Botulinum Neurotoxin Type E Is Unique: Its Implication in Faster Translocation. J. Mol. Biol. 2009, 386, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Košenina, S.; Martínez-Carranza, M.; Davies, J.R.; Masuyer, G.; Stenmark, P. Structural Analysis of Botulinum Neurotoxins Type B and E by Cryo-EM. Toxins 2021, 14, 14. [Google Scholar] [CrossRef]
- Brunger, A.T.; Breidenbach, M.A.; Jin, R.; Fischer, A.; Santos, J.S.; Montal, M. Botulinum Neurotoxin Heavy Chain Belt as an Intramolecular Chaperonefor the Light Chain. PLoS Pathog. 2006, 2, 0922–0932. [Google Scholar] [CrossRef]
- Orrell, K.E.; Melnyk, R.A. Large Clostridial Toxins: Mechanisms and Roles in Disease. Microbiol. Mol. Biol. Rev. 2021, 85, e00064-21. [Google Scholar] [CrossRef]
- Foster, K.A. The Dual-Receptor Recognition of Botulinum Neurotoxins. In Molecular Aspects of Botulinum Neurotoxin; Springer: Berlin/Heidelberg, Germany, 2014; pp. 129–149. ISBN 9781461494546. [Google Scholar]
- Karalewitz, A.P.-A.; Fu, Z.; Baldwin, M.R.; Kim, J.-J.P.; Barbieri, J.T. Botulinum Neurotoxin Serotype C Associates with Dual Ganglioside Receptors to Facilitate Cell Entry. J. Biol. Chem. 2012, 287, 40806–40816. [Google Scholar] [CrossRef]
- Dong, M.; Yeh, F.; Tep, W.H.; Chapman, P.; Dean, C.; Johnson, E.A.; Janz, R.; Chapman, E.R. SV2 Is the Protein Receptor for Botulinum Neurotoxin A. Science 2006, 312, 592–596. [Google Scholar] [CrossRef]
- Peng, L.; Tepp, W.H.; Johnson, E.A.; Dong, M. Botulinum Neurotoxin D Uses Synaptic Vesicle Protein SV2 and Gangliosides as Receptors. PLoS Pathog. 2011, 7, e1002008. [Google Scholar] [CrossRef]
- Dong, M.; Liu, H.; Tepp, W.H.; Johnson, E.A.; Janz, R.; Chapman, E.R. Glycosylated SV2A and SV2B Mediate the Entry of Botulinum Neurotoxin E into Neurons. Mol. Biol. Cell 2008, 19, 5226–5237. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Chen, C.; Barbieri, J.T.; Kim, J.J.P.; Baldwin, M.R. Glycosylated SV2 and Gangliosides as Dual Receptors for Botulinum Neurotoxin Serotype F. Biochemistry 2009, 48, 5631–5641. [Google Scholar] [CrossRef]
- Dong, M.; Richards, D.A.; Goodnough, M.C.; Tepp, W.H.; Johnson, E.A.; Chapman, E.R. Synaptotagmins I and II Mediate Entry of Botulinum Neurotoxin B into Cells. J. Cell Biol. 2003, 162, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Karnath, T.; Henke, T.; Bigalke, H.; Binz, T. Synaptotagmins I and II Act as Nerve Cell Receptors for Botulinum Neurotoxin G. J. Biol. Chem. 2004, 279, 30865–30870. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Swaminathan, S.; Eswaramoorthy, S. Structural Analysis of the Catalytic and Binding Sites of Clostridium Botulinum Neurotoxin B. Nat. Struct. Biol. 2000, 7, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.R.; Liu, S.M.; Acharya, K.R. Variations in the Botulinum Neurotoxin Binding Domain and the Potential for Novel Therapeutics. Toxins 2018, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Solabre Valois, L.; Wilkinson, K.A.; Nakamura, Y.; Henley, J.M. Endocytosis, Trafficking and Exocytosis of Intact Full-Length Botulinum Neurotoxin Type a in Cultured Rat Neurons. Neurotoxicology 2020, 78, 80–87. [Google Scholar] [CrossRef]
- Koriazova, L.K.; Montal, M. Translocation of Botulinum Neurotoxin Light Chain Protease through the Heavy Chain Channel. Nat. Struct. Biol. 2003, 10, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Montal, M. Translocation of Botulinum Neurotoxin Light Chain Protease by the Heavy Chain Protein-Conducting Channel. Toxicon 2009, 54, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.; Matsui, T.; Perry, K.; Rummel, A.; Bowen, M.E.; Guo, Z.; Weisemann, J.; Krez, N.; Jin, R. A Viral-Fusion-Peptide-like Molecular Switch Drives Membrane Insertion of Botulinum Neurotoxin A1. Nat. Commun. 2018, 9, 5367. [Google Scholar] [CrossRef]
- Vaidyanathan, V.V.; Yoshino, K.-I.; Jahnz, M.; Dörries, C.; Bade, S.; Nauenburg, S.; Niemann, H.; Binz, T. Proteolysis of SNAP-25 Isoforms by Botulinum Neurotoxin Types A, C, and E: Domains and Amino Acid Residues Controlling the Formation of Enzyme-Substrate Complexes and Cleavage. J. Neurochem. 1999, 72, 327–337. [Google Scholar] [CrossRef]
- Chen, S.; Hall, C.; Barbieri, J.T. Substrate Recognition of VAMP-2 by Botulinum Neurotoxin B and Tetanus Neurotoxin. J. Biol. Chem. 2008, 283, 21153–21159. [Google Scholar] [CrossRef]
- Masuyer, G.; Davies, J.R.; Moore, K.; Chaddock, J.A.; Acharya, K.R. Structural Analysis of Clostridium Botulinum Neurotoxin Type D as a Platform for the Development of Targeted Secretion Inhibitors. Sci. Rep. 2015, 5, 13397. [Google Scholar] [CrossRef]
- Schiavo, G.; Shone, C.C.; Rossetto, O.; Alexander, F.C.G.; Montecucco, C. Botulinum Neurotoxin Serotype F Is a Zinc Endopeptidase Specific for VAMP/Synaptobrevin. J. Biol. Chem. 1993, 268, 11516–11519. [Google Scholar] [CrossRef]
- Schiavo, G.; Malizios, C.; Trimblell, W.S.; Polverino De Lauretos, P.; Milant, G.; Sugiyamas, H.; Johnsons, E.A.; Montecuccoi, C. Botulinum G Neurotoxin Cleaves VAMP/Synaptobrevin at a Single Ala-Ala Peptide Bond. J. Biol. Chem. 1994, 269, 20213–20216. [Google Scholar] [CrossRef]
- Masuyer, G.; Zhang, S.; Barkho, S.; Shen, Y.; Henriksson, L.; Košenina, S.; Dong, M.; Stenmark, P. Structural Characterisation of the Catalytic Domain of Botulinum Neurotoxin X–High Activity and Unique Substrate Specificity. Sci. Rep. 2018, 8, 4518. [Google Scholar] [CrossRef] [PubMed]
- Foran, P.; Lawrence, G.W.; Shone, C.C.; Foster, K.A.; Dolly, J.O. Botulinum Neurotoxin C1 Cleaves Both Syntaxin and SNAP-25 in Intact and Permeabilized Chromaffin Cells: Correlation with Its Blockade of Catecholamine Release. Biochemistry 1996, 35, 2630–2636. [Google Scholar] [CrossRef] [PubMed]
- Blasi, J.; Chapman, E.R.; Yamasaki, S.; Binz, T.; Niemann, H.; Jahn, R. Botulinum Neurotoxin C1 Blocks Neurotransmitter Release by Means of Cleaving HPC-1/Syntaxin. EMBO J. 1993, 12, 4821–4828. [Google Scholar] [CrossRef]
- Fonfria, E.; Maignel, J.; Lezmi, S.; Martin, V.; Splevins, A.; Shubber, S.; Kalinichev, M.; Foster, K.; Picaut, P.; Krupp, J. The Expanding Therapeutic Utility of Botulinum Neurotoxins. Toxins 2018, 10, 208. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Fujisawa, T.; Mochizuki, T.; Kaminuma, E.; Suzuki, Y.; Nakamura, Y.; Tohno, M. Draft Genome Sequence of Weissella Oryzae SG25T, Isolated from Fermented Rice Grains. Genome Announc. 2014, 2, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, M.J.; Adams, J.B.; Doxey, A.C. Botulinum Neurotoxin Homologs in Non-Clostridium Species. FEBS Lett. 2014, 589, 342–348. [Google Scholar] [CrossRef]
- Brunt, J.; Carter, A.T.; Stringer, S.C.; Peck, M.W. Identification of a Novel Botulinum Neurotoxin Gene Cluster in Enterococcus. FEBS Lett. 2018, 592, 310–317. [Google Scholar] [CrossRef]
- Zhang, S.; Lebreton, F.; Mansfield, M.J.; Miyashita, S.I.; Zhang, J.; Schwartzman, J.A.; Tao, L.; Masuyer, G.; Martínez-Carranza, M.; Stenmark, P.; et al. Identification of a Botulinum Neurotoxin-like Toxin in a Commensal Strain of Enterococcus Faecium. Cell Host Microbe 2018, 23, 169–176. [Google Scholar] [CrossRef]
- Zhou, X.; Willems, R.J.L.; Friedrich, A.W.; Rossen, J.W.A.; Bathoorn, E. Enterococcus Faecium: From Microbiological Insights to Practical Recommendations for Infection Control and Diagnostics. Antimicrob. Resist. Infect. Control. 2020, 9, 130. [Google Scholar] [CrossRef]
- Newman, J. Novel Buffer Systems for Macromolecular Crystallization. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 610–612. [Google Scholar] [CrossRef]
- Boivin, S.; Kozak, S.; Meijers, R. Optimization of Protein Purification and Characterization Using Thermofluor Screens. Protein Expr. Purif. 2013, 91, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Košenina, S.; Masuyer, G.; Zhang, S.; Dong, M.; Stenmark, P. Crystal Structure of the Catalytic Domain of the Weissella Oryzae Botulinum-like Toxin. FEBS Lett. 2019, 593, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Voss, N.R.; Gerstein, M. 3V: Cavity, Channel and Cleft Volume Calculator and Extractor. Nucleic Acids Res. 2010, 38, W555–W562. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS Database of Proteolytic Enzymes, Their Substrates and Inhibitors in 2017 and a Comparison with Peptidases in the PANTHER Database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Swaminathan, S. Molecular Structures and Functional Relationships in Clostridial Neurotoxins. FEBS J. 2011, 278, 4467–4485. [Google Scholar] [CrossRef]
- Breidenbach, M.A.; Brunger, A.T. Substrate Recognition Strategy for Botulinum Neurotoxin Serotype A. Nature 2004, 432, 925–929. [Google Scholar] [CrossRef]
- Kumaran, D.; Rawat, R.; Ahmed, S.A.; Swaminathan, S. Substrate Binding Mode and Its Implication on Drug Design for Botulinum Neurotoxin A. PLoS Pathog. 2008, 4, e1000165. [Google Scholar] [CrossRef]
- Binz, T.; Bade, S.; Rummel, A.; Kollewe, A.; Alves, J. Arg36 and Tyr365 of the Botulinum Neurotoxin Type A Light Chain Are Involved in Transition State Stabilization. Biochemistry 2002, 41, 1717–1723. [Google Scholar] [CrossRef]
- Agarwal, R.; Schmidt, J.J.; Stafford, R.G.; Swaminathan, S. Mode of VAMP Substrate Recognition and Inhibition of Clostridium Botulinum Neurotoxin F. Nat. Struct. Mol. Biol. 2009, 16, 789–794. [Google Scholar] [CrossRef]
- Chen, S.; Kim, J.-J.P.; Barbieri, J.T. Mechanism of Substrate Recognition by Botulinum Neurotoxin Serotype A. J. Biol. Chem. 2007, 282, 9621–9627. [Google Scholar] [CrossRef]
- Lam, K.; Tremblay, J.M.; Perry, K.; Ichtchenko, K.; Shoemaker, C.B.; Jin, R. Probing the Structure and Function of the Protease Domain of Botulinum Neurotoxins Using Single-Domain Antibodies. PLoS Pathog. 2022, 18, e1010169. [Google Scholar] [CrossRef] [PubMed]
- Barthels, F.; Schirmeister, T.; Kersten, C. BANΔIT: B’-Factor Analysis for Drug Design and Structural Biology. Mol. Inf. 2021, 40, 202000144. [Google Scholar] [CrossRef] [PubMed]
- Segelke, B.; Knapp, M.; Kadkhodayan, S.; Balhorn, R.; Rupp, B. Crystal Structure of Clostridium Botulinum Neurotoxin Protease in a Product-Bound State: Evidence for Noncanonical Zinc Protease Activity. Proc. Natl. Acad. Sci. USA 2004, 101, 6888–6893. [Google Scholar] [CrossRef] [PubMed]
- DasGupta, B.R.; Antharavally, B.S.; Tepp, W.; Evenson, M.L. Botulinum Neurotoxin Types A, B, and E: Fragmentations by Autoproteolysis and Other Mechanisms Including by O-Phenanthroline–Dithiothreitol, and Association of the Dinucleotides NAD+/NADH with the Heavy Chain of the Three Neurotoxins. Protein J. 2005, 24, 337–368. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, B.R.; Foley, J.C. Botulinum Neurotoxin Types A and E: Isolated Light Chain Breaks down into Two Fragments. Comparison of Their Amino Acid Sequences with Tetanus Neurotoxin. Biochimie 1989, 71, 1193–1200. [Google Scholar] [CrossRef]
- Beecher, D.J.; DasGupta, B.R. Botulinum Neurotoxin Type A: Limited Proteolysis by Endoproteinase Glu-C and α-Chymotrypsin Enhanced Following Reduction; Identification of the Cleaved Sites and Fragments. J. Protein Chem. 1997, 16, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Mizanur, R.M.; Frasca, V.; Swaminathan, S.; Bavari, S.; Webb, R.; Smith, L.A.; Ahmed, S.A. The C Terminus of the Catalytic Domain of Type A Botulinum Neurotoxin May Facilitate Product Release from the Active Site. J. Biol. Chem. 2013, 288, 24223–24233. [Google Scholar] [CrossRef]
- Winter, G.; Waterman, D.G.; Parkhurst, J.M.; Brewster, A.S.; Gildea, R.J.; Gerstel, M.; Fuentes-Montero, L.; Vollmar, M.; Michels-Clark, T.; Young, I.D.; et al. DIALS: Implementation and Evaluation of a New Integration Package. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 85–97. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 Suite and Current Developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser Crystallographic Software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the Refinement of Macromolecular Crystal Structures. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and Better Reference Data for Improved All-Atom Structure Validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- McNicholas, S.; Potterton, E.; Wilson, K.S.; Noble, M.E.M. Presenting Your Structures: The CCP4mg Molecular-Graphics Software. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 386–394. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- University of Bath, Material and Chemical Characterisation Facility (MC2). [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beamline | I04 (Diamond Light Source) |
| Wavelength | 0.9537 Å |
| Calculated dose | 10.0 MGy |
| Crystallographic statistics | |
| Resolution (Å) | 101.75–2.0 (2.05–2.0) |
| Space group | P 42212 |
| Unit cell dimensions | |
| a, b, c (Å) | 143.894, 143.894, 51.695 |
| α, β, γ (°) | 90.00, 90.00, 90.00 |
| No. reflections | 982,082 |
| No. unique reflections | 37,319 (2683) |
| Completeness (%) | 100 (100) |
| Rmerge | 0.131 (3.877) |
| Rpim | 0.036 (0.763) |
| <I/σ(I)> | 13.2 (0.9) |
| CC1/2 | 1.00 (0.626) |
| Multiplicity | 26.3 (26.6) |
| Refinement statistics | |
| Rwork/Rfree | 0.19/0.23 |
| RMSD bond length (Å) | 0.0106 |
| RMSD bond angles (°) | 1.910 |
| Ramachandran plot statistics (%) | |
| Favoured | 97% |
| Allowed | 3.00% |
| Outliers | 0.00% |
| Average B-factors (Å2) | |
| Amino acids | 58.78 |
| Zinc | 48.17 |
| Ligands | 69.23 |
| Water | 54.73 |
| Loop 1 | 50.04 |
| Loop 2 | 90.04 |
| Loop 3 | 52.84 |
| Loop 4 | 80.84 |
| Loop 5 | 72.09 |
| Number of atoms | |
| Amino acids | 3226 |
| Zinc | 1 |
| Ligands | 24 |
| Water | 156 |
| S1 | S1′ | S2′ | S3′ | S4′ | S5′ | |
|---|---|---|---|---|---|---|
| LC/En | G169 | R364 T222 M166 V372 P217 K168 K195 | R364 V369 V372 | W258 E263 V265 Y367 V369 P370 | A167 | W258 E263 S260 L442 P370 |
| LC/A | E164 | R363 T220 I161 D370 T215 F163 F194 | R363 N368 D370 | Y251 L256 V258 Y366 N368 F369 | Q162 | Y251 L256 S254 F423 F369 |
| LC/En (Residues 1–425) | Belt (BoNT/En residues 484–514) |
|---|---|
| Q112 | L484 |
| K341 | S485 |
| D342 | N486 |
| D345 | |
| K321 | Y487 |
| Y235 | |
| R119 | D488 |
| R119 | K491 |
| E313 | T496 |
| E313 | S497 |
| M133 | P500 |
| T135 | I502 |
| M133 | |
| T135 | V504 |
| S176 | D506 |
| Y125 | |
| K175 | P507 |
| L173 | E509 |
| R177 | |
| R177 | |
| K171 | D511 |
| K171 | |
| T63 | |
| S62 | |
| K171 | D513 |
| K171 |
| Loop | LC/En | LC/A | LC/F | LC/X |
|---|---|---|---|---|
| 1 | 23 (20–42) | 13 (21–33) | 14 (21–34) | 18 (21–38) |
| 2 | 11 (123–133) | 13 (117–129) | 15 (117–131) | 11 (121–131) |
| 3 | 6 (172–177) | 10 (167–176) | 15 (169–183) | 7 (169–175) |
| 4 | 23 (244–266) | 18 (242–259) | 17 (247–263) | 18 (246–263) |
| 5 | 9 (306–314) | 15 (300–314) | 14 (303–316) | 8 (304–311) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregory, K.S.; Hall, P.-R.; Onuh, J.P.; Mojanaga, O.O.; Liu, S.M.; Acharya, K.R. Crystal Structure of the Catalytic Domain of a Botulinum Neurotoxin Homologue from Enterococcus faecium: Potential Insights into Substrate Recognition. Int. J. Mol. Sci. 2023, 24, 12721. https://doi.org/10.3390/ijms241612721
Gregory KS, Hall P-R, Onuh JP, Mojanaga OO, Liu SM, Acharya KR. Crystal Structure of the Catalytic Domain of a Botulinum Neurotoxin Homologue from Enterococcus faecium: Potential Insights into Substrate Recognition. International Journal of Molecular Sciences. 2023; 24(16):12721. https://doi.org/10.3390/ijms241612721
Chicago/Turabian StyleGregory, Kyle S., Peter-Rory Hall, Jude Prince Onuh, Otsile O. Mojanaga, Sai Man Liu, and K. Ravi Acharya. 2023. "Crystal Structure of the Catalytic Domain of a Botulinum Neurotoxin Homologue from Enterococcus faecium: Potential Insights into Substrate Recognition" International Journal of Molecular Sciences 24, no. 16: 12721. https://doi.org/10.3390/ijms241612721
APA StyleGregory, K. S., Hall, P.-R., Onuh, J. P., Mojanaga, O. O., Liu, S. M., & Acharya, K. R. (2023). Crystal Structure of the Catalytic Domain of a Botulinum Neurotoxin Homologue from Enterococcus faecium: Potential Insights into Substrate Recognition. International Journal of Molecular Sciences, 24(16), 12721. https://doi.org/10.3390/ijms241612721