


Mechanistic Protective Effect of Cilostazol in Cisplatin-Induced Testicular Damage via Regulation of Oxidative Stress and TNF-α/NF-κB/Caspase-3 Pathways


Abstract
1. Introduction
2. Results
2.1. Effect on Relative Testicular Weight, Testosterone Concentration, and Epididymal Sperm Parameters
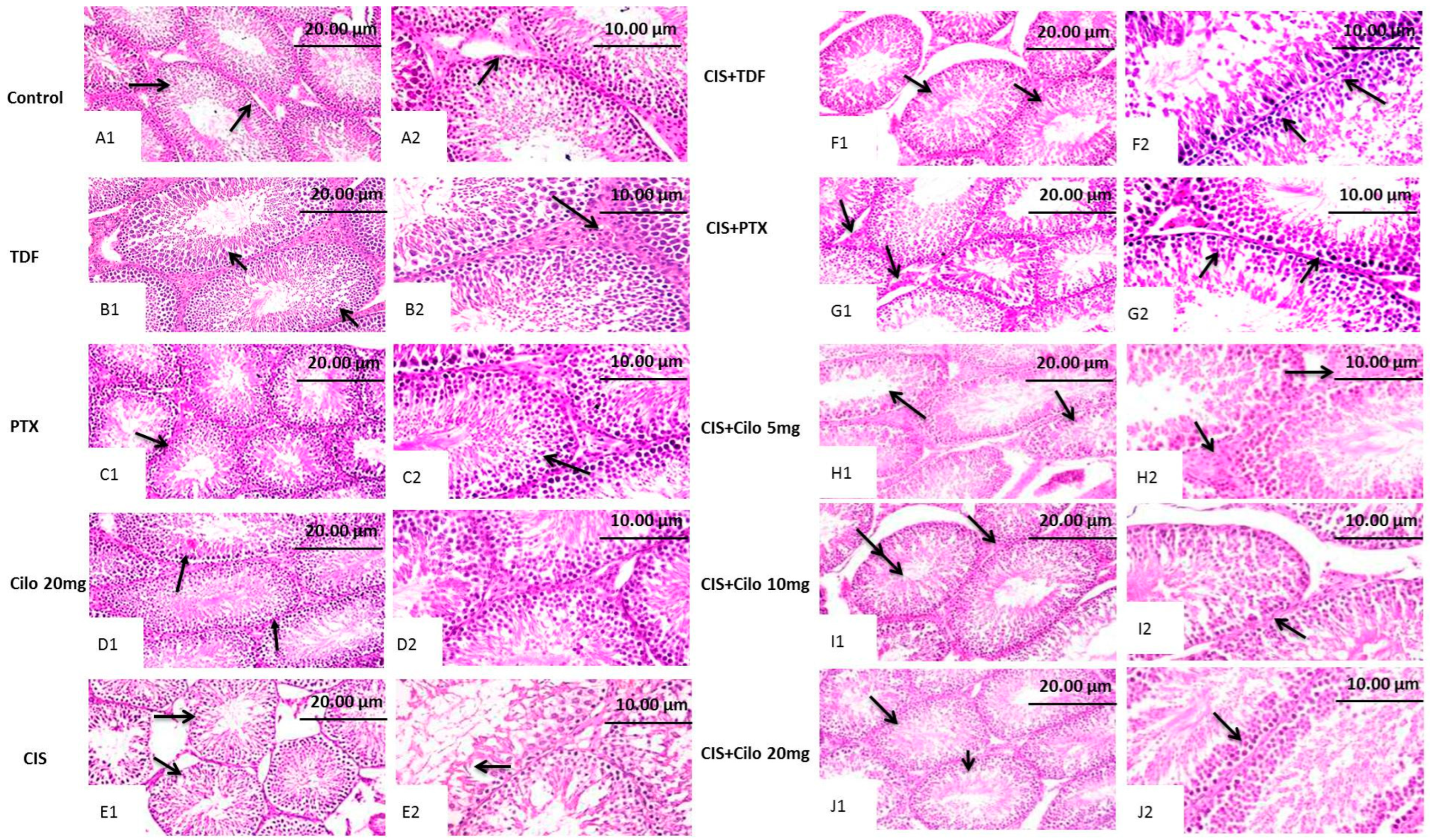
2.2. Histopathological Findings of Testicular Tissues
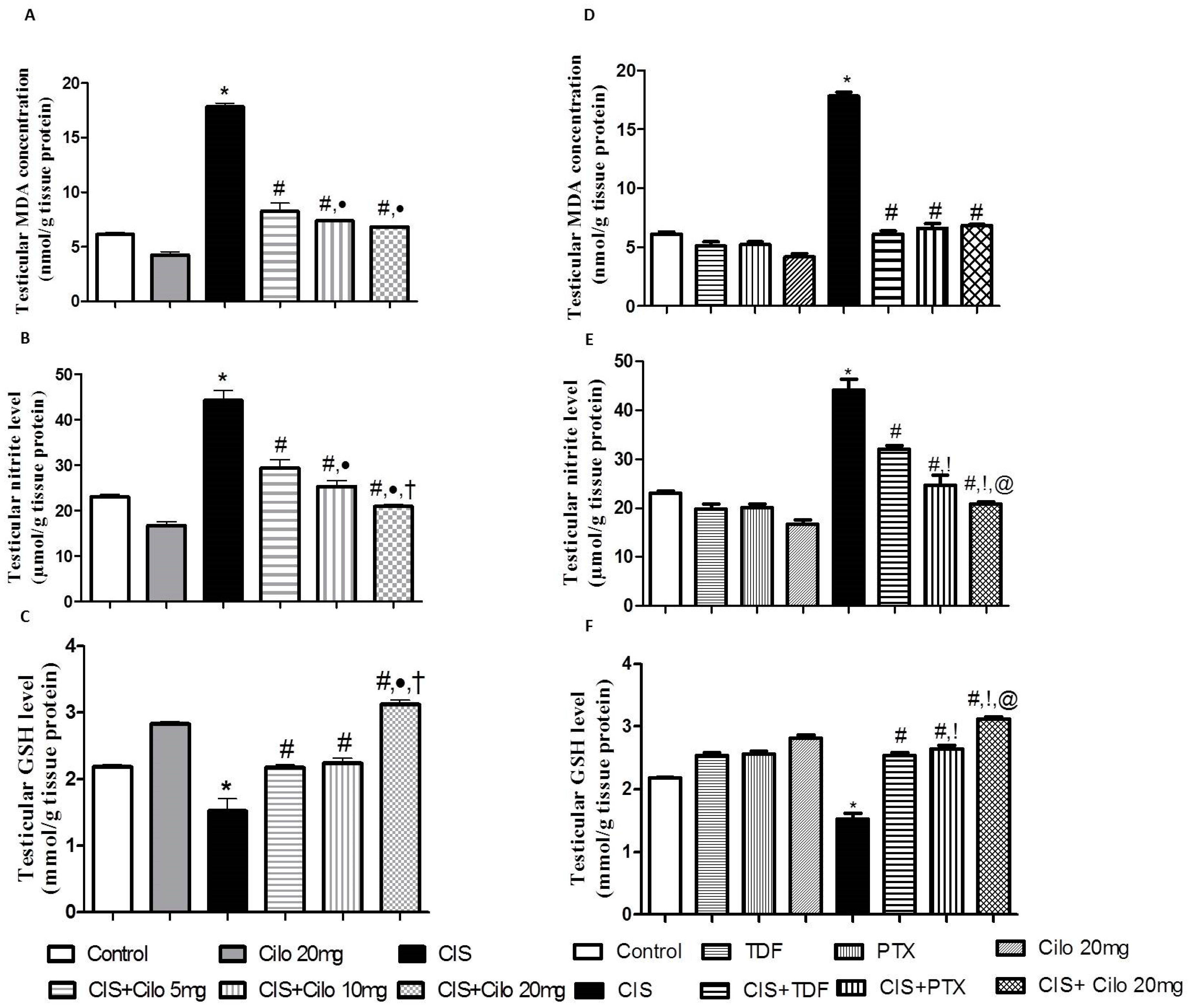
2.3. Effect on Testicular Malondialdehyde, Total Nitrite Content, and Reduced Glutathione
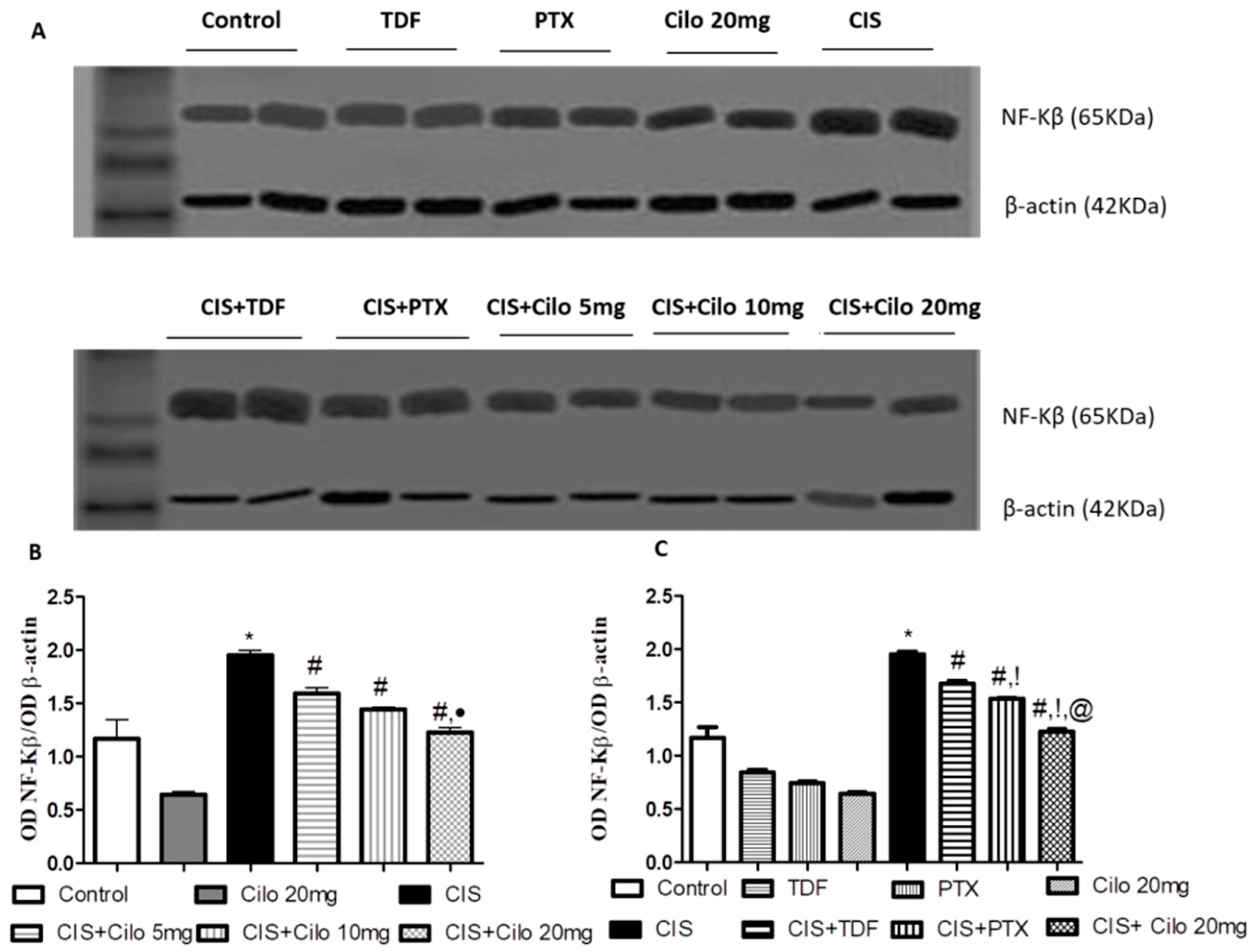
2.4. Effects on Testicular Inflammatory Mediators (TNF-α and NF-κB) Protein Expressions
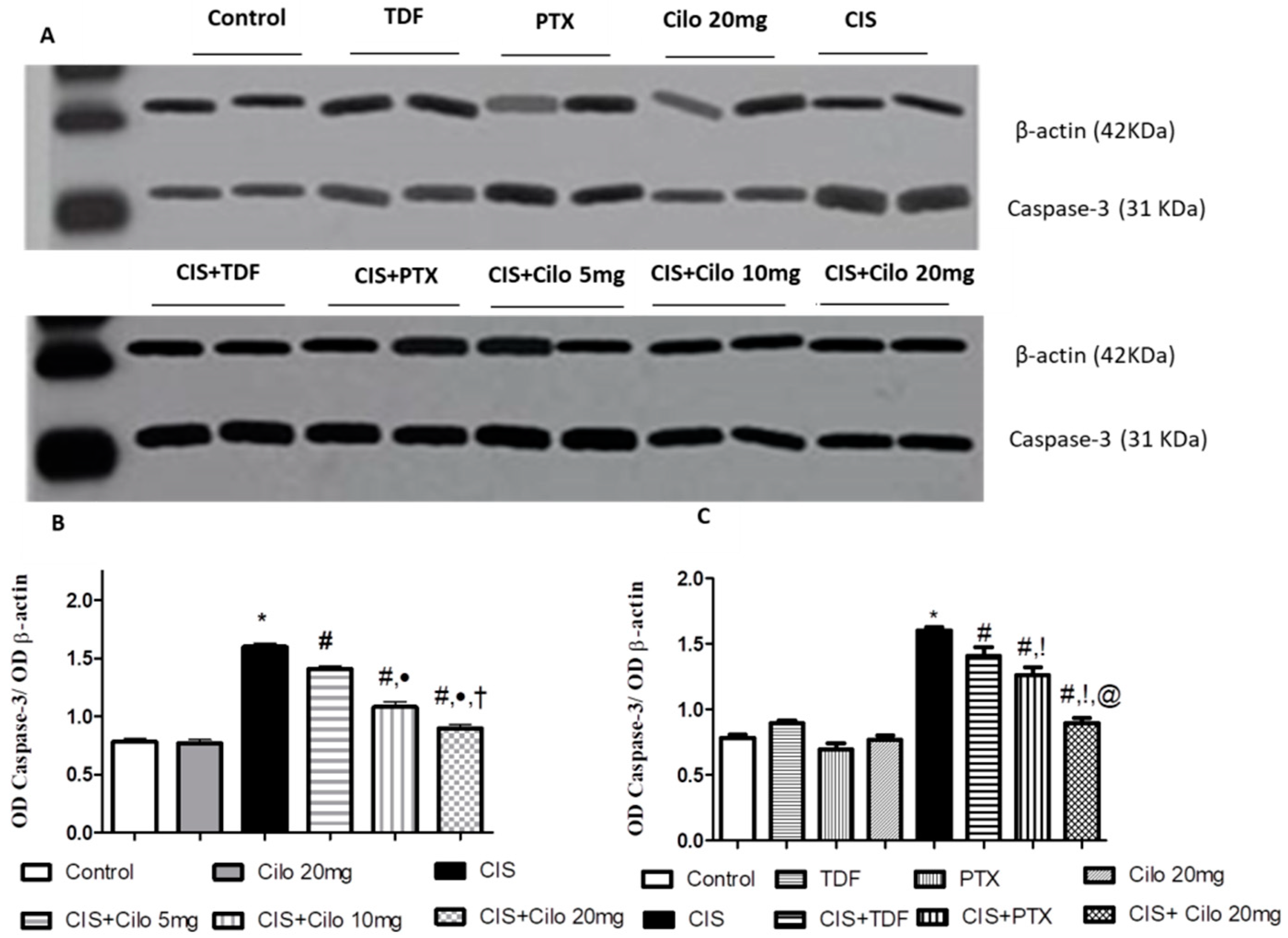
2.5. Effects on Testicular Caspase-3 Protein Expression
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Experimental Design
4.3. Collection of Samples
4.4. Sperm Motility and Count
4.5. Determination of the Serum Testosterone Level
4.6. Determination of Testicular Oxidative Stress Markers
4.7. Western Blot Analysis
4.8. Histopathological Examination
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heeba, G.H.; Hamza, A.A.; Hassanin, S.O. Induction of heme oxygenase-1 with hemin alleviates cisplatin-induced reproductive toxicity in male rats and enhances its cytotoxicity in prostate cancer cell line. Toxicol. Lett. 2016, 264, 38–50. [Google Scholar]
- Volarevic, V.; Djokovic, B.; Jankovic, M.G.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N. Molecular mechanisms of cisplatin-induced nephrotoxicity: A balance on the knife edge between renoprotection and tumor toxicity. J. Biomed. Sci. 2019, 26, 25. [Google Scholar]
- Abdel-Latif, R.; Fathy, M.; Anwar, H.A.; Naseem, M.; Dandekar, T.; Othman, E.M. Cisplatin-Induced Reproductive Toxicity and Oxidative Stress: Ameliorative Effect of Kinetin. Antioxidants 2022, 11, 863. [Google Scholar]
- Kohsaka, T.; Minagawa, I.; Morimoto, M.; Yoshida, T.; Sasanami, T.; Yoneda, Y.; Ikegaya, N.; Sasada, H. Efficacy of relaxin for cisplatin-induced testicular dysfunction and epididymal spermatotoxicity. Basic Clin. Androl. 2020, 30, 3. [Google Scholar]
- Mesbahzadeh, B.; Hassanzadeh-Taheri, M.; Aliparast, M.-s.; Baniasadi, P.; Hosseini, M. The protective effect of crocin on cisplatin-induced testicular impairment in rats. BMC Urol. 2021, 21, 117. [Google Scholar]
- Hansson, V.; Skålhegg, B.S.; Taskén, K. Cyclic-AMP-dependent protein kinase (PKA) in testicular cells. Cell specific expression, differential regulation and targeting of subunits of PKA. J. Steroid Biochem. Mol. Biol. 2000, 73, 81–92. [Google Scholar]
- Golkowski, M.; Shimizu-Albergine, M.; Suh, H.W.; Beavo, J.A.; Ong, S.E. Studying mechanisms of cAMP and cyclic nucleotide phosphodiesterase signaling in Leydig cell function with phosphoproteomics. Cell. Signal. 2016, 28, 764–778. [Google Scholar]
- Calamera, G.; Moltzau, L.R.; Levy, F.O.; Andressen, K.W. Phosphodiesterases and Compartmentation of cAMP and cGMP Signaling in Regulation of Cardiac Contractility in Normal and Failing Hearts. Int. J. Mol. Sci. 2022, 23, 2145. [Google Scholar]
- Özgür, B.C.; Telli, O.; Yuceturk, C.N.; Sarici, H.; Ozer, E.; Surer, H.; Kılınc, A.S.; Hucumenoglu, S.; Eroglu, M. The effect of sildenafil and udenafil on testicular damage following ischemia-reperfusion injury in rats. J. Urol. 2014, 192, 1272–1277. [Google Scholar]
- Yao, C.; Li, G.; Qian, Y.; Cai, M.; Yin, H.; Xiao, L.; Tang, W.; Guo, F.; Shi, B. Protection of Pentoxifylline against Testis Injury Induced by Intermittent Hypobaric Hypoxia. Oxidative Med. Cell. Longev. 2016, 2016, 3406802. [Google Scholar]
- Mohamed, M.; Hafez, H.; Zenhom, N.; Mohammed, H. Cilostazol alleviates streptozotocin-induced testicular injury in rats via PI3K/Akt pathway. Life Sci. 2018, 198, 136–142. [Google Scholar] [CrossRef]
- Kölükçü, E.; Atılgan, D.; Uluocak, N.; Deresoy, F.A.; Katar, M.; Unsal, V. Milrinone ameliorates ischaemia-reperfusion injury in experimental testicular torsion/detorsion rat model. Andrologia 2021, 53, e14128. [Google Scholar]
- Abdel-Wahab, B.A.; Alkahtani, S.A.; Elagab, E.A.M. Tadalafil alleviates cisplatin-induced reproductive toxicity through the activation of the Nrf2/HO-1 pathway and the inhibition of oxidative stress and apoptosis in male rats. Reprod. Toxicol. 2020, 96, 165–174. [Google Scholar]
- Kaku, Y.; Chiba, K.; Sato, K.; Onishi, A.; Ishida, T.; Okada, K.; Fujisawa, M. Protective effects of tadalafil against cisplatin-induced spermatogenic dysfunction. Biochem. Biophys. Res. Commun. 2022, 603, 123–129. [Google Scholar]
- Fallahzadeh, A.R.; Rezaei, Z.; Rahimi, H.R.; Barmak, M.J.; Sadeghi, H.; Mehrabi, S.; Rabani, S.M.; Kashani, I.R.; Barati, V.; Mahmoudi, R. Evaluation of the Effect of Pentoxifylline on Cisplatin-Induced Testicular Toxicity in Rats. Toxicol. Res. 2017, 33, 255–263. [Google Scholar] [CrossRef]
- Hoshino, H.; Toyoda, K.; Omae, K.; Ishida, N.; Uchiyama, S.; Kimura, K.; Sakai, N.; Okada, Y.; Tanaka, K.; Origasa, H.; et al. Dual Antiplatelet Therapy Using Cilostazol With Aspirin or Clopidogrel: Subanalysis of the CSPS.com Trial. Stroke 2021, 52, 3430–3439. [Google Scholar]
- Real, J.; Serna, M.C.; Giner-Soriano, M.; Forés, R.; Pera, G.; Ribes, E.; Alzamora, M.; Marsal, J.R.; Heras, A.; Morros, R. Safety of cilostazol in peripheral artery disease: A cohort from a primary healthcare electronic database. BMC Cardiovasc. Disord. 2018, 18, 85. [Google Scholar]
- Asal, N.J.; Wojciak, K.A. Effect of cilostazol in treating diabetes-associated microvascular complications. Endocrine 2017, 56, 240–244. [Google Scholar] [CrossRef]
- He, P.; Kawamura, H.; Takemoto, M.; Maezawa, Y.; Ishikawa, T.; Ishibashi, R.; Sakamoto, K.; Shoji, M.; Hattori, A.; Yamaga, M.; et al. Combination of cilostazol and probucol protected podocytes from lipopolysaccharide-induced injury by both anti-inflammatory and anti-oxidative mechanisms. J. Nephrol. 2017, 30, 531–541. [Google Scholar]
- Hafez, H.M.; Ibrahim, M.A.; Zedan, M.Z.; Hassan, M.; Hassanein, H. Nephroprotective effect of cilostazol and verapamil against thioacetamide-induced toxicity in rats may involve Nrf2/HO-1/NQO-1 signaling pathway. Toxicol. Mech. Methods 2019, 29, 146–152. [Google Scholar]
- Refaie, M.M.M.; Ahmed Ibrahim, R.; Shehata, S. Dose dependent effect of cilostazol in induced testicular ischemia reperfusion via modulation of HIF/VEGF and cAMP/SIRT1 pathways. Int. Immunopharmacol. 2021, 101 Pt A, 108197. [Google Scholar]
- Adelakun, S.A.; Ogunlade, B.; Fidelis, O.P.; Omotoso, O.D. Protective effect of nutritional supplementation of zinc-sulfate against cisplatin-induced spermatogonial and testicular dysfunctions in adult male Sprague-Dawley rats. Endocr. Metab. Sci. 2022, 6, 100116. [Google Scholar]
- Soni, K.K.; Kim, H.K.; Choi, B.R.; Karna, K.K.; You, J.H.; Cha, J.S.; Shin, Y.S.; Lee, S.W.; Kim, C.Y.; Park, J.K. Dose-dependent effects of cisplatin on the severity of testicular injury in Sprague Dawley rats: Reactive oxygen species and endoplasmic reticulum stress. Drug Des. Dev. Ther. 2016, 10, 3959–3968. [Google Scholar]
- Heeba, G.H.; Hamza, A.A. Rosuvastatin ameliorates diabetes-induced reproductive damage via suppression of oxidative stress, inflammatory and apoptotic pathways in male rats. Life Sci. 2015, 141, 13–19. [Google Scholar]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar]
- García, M.M.; Acquier, A.; Suarez, G.; Gomez, N.V.; Gorostizaga, A.; Mendez, C.F.; Paz, C. Cisplatin inhibits testosterone synthesis by a mechanism that includes the action of reactive oxygen species (ROS) at the level of P450scc. Chem.-Biol. Interact. 2012, 199, 185–191. [Google Scholar]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar]
- Saad, S.Y.; Najjar, T.A.; Alashari, M. Role of non-selective adenosine receptor blockade and phosphodiesterase inhibition in cisplatin-induced nephrogonadal toxicity in rats. Clin. Exp. Pharmacol. Physiol. 2004, 31, 862–867. [Google Scholar] [CrossRef]
- Dhulqarnain, A.O.; Takzaree, N.; Hassanzadeh, G.; Tooli, H.; Malekzadeh, M.; Khanmohammadi, N.; Yaghobinejad, M.; Solhjoo, S.; Rastegar, T. Pentoxifylline improves the survival of spermatogenic cells via oxidative stress suppression and upregulation of PI3K/AKT pathway in mouse model of testicular torsion-detorsion. Heliyon 2021, 7, e06868. [Google Scholar] [PubMed]
- Garg, H.; Kumar, R. An update on the role of medical treatment including antioxidant therapy in varicocele. Asian J. Androl. 2016, 18, 222–228. [Google Scholar]
- Hamza, A.A.; Elwy, H.M.; Badawi, A.M. Fenugreek seed extract attenuates cisplatin-induced testicular damage in Wistar rats. Andrologia 2016, 48, 211–221. [Google Scholar]
- Afsar, T.; Razak, S.; Khan, M.R.; Almajwal, A. Acacia hydaspica ethyl acetate extract protects against cisplatin-induced DNA damage, oxidative stress and testicular injuries in adult male rats. BMC Cancer 2017, 17, 883. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.A.A.; Eid, B.G. Cisplatin induced testicular damage through mitochondria mediated apoptosis, inflammation and oxidative stress in rats: Impact of resveratrol. Endocr. J. 2020, 67, 969–980. [Google Scholar]
- Moradi, M.; Goodarzi, N.; Faramarzi, A.; Cheraghi, H.; Hashemian, A.H.; Jalili, C. Melatonin protects rats testes against bleomycin, etoposide, and cisplatin-induced toxicity via mitigating nitro-oxidative stress and apoptosis. Biomed. Pharmacother. 2021, 138, 111481. [Google Scholar]
- Sherif, I.O.; Abdel-Aziz, A.; Sarhan, O.M. Cisplatin-induced testicular toxicity in rats: The protective effect of arjunolic acid. J. Biochem. Mol. Toxicol. 2014, 28, 515–521. [Google Scholar]
- O’Bryan, M.K.; Schlatt, S.; Gerdprasert, O.; Phillips, D.J.; de Kretser, D.M.; Hedger, M.P. Inducible nitric oxide synthase in the rat testis: Evidence for potential roles in both normal function and inflammation-mediated infertility. Biol. Reprod. 2000, 63, 1285–1293. [Google Scholar]
- Wadie, W.; Abdel-Razek, N.S.; Salem, H.A. Phosphodiesterase (1, 3 & 5) inhibitors attenuate diclofenac-induced acute kidney toxicity in rats. Life Sci. 2021, 277, 119506. [Google Scholar]
- El-Deen, R.M.; Heeba, G.H.; Abdel-Latif, R.G.; Khalifa, M.M.A. Comparative effectiveness of phosphodiesterase 3, 4, and 5 inhibitors in amelioration of high-fat diet-induced nonalcoholic fatty liver in rats. Fundam. Clin. Pharmacol. 2020, 34, 353–364. [Google Scholar]
- Elrashidy, R.A.; Hasan, R.A. Cilostazol preconditioning alleviates cyclophosphamide-induced cardiotoxicity in male rats: Mechanistic insights into SIRT1 signaling pathway. Life Sci. 2021, 266, 118822. [Google Scholar]
- El-Abhar, H.; Abd El Fattah, M.A.; Wadie, W.; El-Tanbouly, D.M. Cilostazol disrupts TLR-4, Akt/GSK-3β/CREB, and IL-6/JAK-2/STAT-3/SOCS-3 crosstalk in a rat model of Huntington’s disease. PLoS ONE 2018, 13, e0203837. [Google Scholar]
- Sakamoto, T.; Ohashi, W.; Tomita, K.; Hattori, K.; Matsuda, N.; Hattori, Y. Anti-inflammatory properties of cilostazol: Its interruption of DNA binding activity of NF-κB from the Toll-like receptor signaling pathways. Int. Immunopharmacol. 2018, 62, 120–131. [Google Scholar]
- Zidek, Z. Adenosine-cyclic AMP pathways and cytokine expression. Eur. Cytokine Netw. 1999, 10, 319–328. [Google Scholar]
- Mansour, H.M.; Salama, A.A.A.; Abdel-Salam, R.M.; Ahmed, N.A.; Yassen, N.N.; Zaki, H.F. The anti-inflammatory and anti-fibrotic effects of tadalafil in thioacetamide-induced liver fibrosis in rats. Can. J. Physiol. Pharmacol. 2018, 96, 1308–1317. [Google Scholar]
- Mostafa-Hedeab, G.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Jeandet, P.; Saad, H.M.; Batiha, G.E. A raising dawn of pentoxifylline in management of inflammatory disorders in Covid-19. Inflammopharmacology 2022, 30, 799–809. [Google Scholar] [CrossRef]
- Dong, J.; Yuan, X.; Xie, W. Pentoxifylline exerts anti-inflammatory effects on cerebral ischemia reperfusion-induced injury in a rat model via the p38 mitogen-activated protein kinase signaling pathway. Mol. Med. Rep. 2018, 17, 1141–1147. [Google Scholar]
- Amin, A.; Hamza, A.A. Effects of Roselle and Ginger on cisplatin-induced reproductive toxicity in rats. Asian J. Androl. 2006, 8, 607–612. [Google Scholar] [CrossRef]
- Wang, F.Y.; Tang, X.M.; Wang, X.; Huang, K.B.; Feng, H.W.; Chen, Z.F.; Liu, Y.N.; Liang, H. Mitochondria-targeted platinum(II) complexes induce apoptosis-dependent autophagic cell death mediated by ER-stress in A549 cancer cells. Eur. J. Med. Chem. 2018, 155, 639–650. [Google Scholar]
- Liu, Z.; Li, C.; Kang, N.; Malhi, H.; Shah, V.H.; Maiers, J.L. Transforming growth factor β (TGFβ) cross-talk with the unfolded protein response is critical for hepatic stellate cell activation. J. Biol. Chem. 2019, 294, 3137–3151. [Google Scholar]
- Casares, C.; Ramírez-Camacho, R.; Trinidad, A.; Roldán, A.; Jorge, E.; García-Berrocal, J.R. Reactive oxygen species in apoptosis induced by cisplatin: Review of physiopathological mechanisms in animal models. Eur. Arch. Oto-Rhino-Laryngol. 2012, 269, 2455–2459. [Google Scholar]
- Matsumoto, M.; Nakajima, W.; Seike, M.; Gemma, A.; Tanaka, N. Cisplatin-induced apoptosis in non-small-cell lung cancer cells is dependent on Bax- and Bak-induction pathway and synergistically activated by BH3-mimetic ABT-263 in p53 wild-type and mutant cells. Biochem. Biophys. Res. Commun. 2016, 473, 490–496. [Google Scholar] [CrossRef]
- Abdel-Aziz, A.M.; Mohamed, A.S.M.; Abdelazem, O.; Okasha, A.M.M.; Kamel, M.Y. Cilostazol protects against cyclophosphamide-induced ovarian toxicity in female rats: Role of cAMP and HO-1. Toxicol. Mech. Methods 2020, 30, 526–535. [Google Scholar]
- Tian, C.J.; Kim, Y.J.; Kim, S.W.; Lim, H.J.; Kim, Y.S.; Choung, Y.H. A combination of cilostazol and Ginkgo biloba extract protects against cisplatin-induced Cochleo-vestibular dysfunction by inhibiting the mitochondrial apoptotic and ERK pathways. Cell Death Dis. 2013, 4, e509. [Google Scholar]
- Koka, S.; Kukreja, R.C. Attenuation of Doxorubicin-induced Cardiotoxicity by Tadalafil: A Long Acting Phosphodiesterase-5 Inhibitor. Mol. Cell. Pharmacol. 2010, 2, 173–178. [Google Scholar]
- Akbari, Z.; Reisi, P.; Torkaman-Boutorabi, A.; Farahmandfar, M. Effect of Pentoxifylline on Apoptotic-Related Gene Expression Profile, Learning and Memory Impairment Induced by Systemic Lipopolysaccharide Administration in the Rat Hippocampus. Int. J. Prev. Med. 2020, 11, 151. [Google Scholar]
- Rezvanfar, M.A.; Rezvanfar, M.A.; Shahverdi, A.R.; Ahmadi, A.; Baeeri, M.; Mohammadirad, A.; Abdollahi, M. Protection of cisplatin-induced spermatotoxicity, DNA damage and chromatin abnormality by selenium nano-particles. Toxicol. Appl. Pharmacol. 2013, 266, 356–365. [Google Scholar]
- Kenjale, R.; Shah, R.; Sathaye, S. Effects of Chlorophytum borivilianum on sexual behaviour and sperm count in male rats. Phytother. Res. 2008, 22, 796–801. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. [30] Microsomal lipid peroxidation. In Methods in Enzymology; Fleischer, S., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 1978; Volume 52, pp. 302–310. [Google Scholar]
- Sastry, K.; Moudgal, R.; Mohan, J.; Tyagi, J.S.; Rao, G. Spectrophotometric Determination of Serum Nitrite and Nitrate by Copper-Cadmium Alloy. Anal. Biochem. 2002, 306, 79–82. [Google Scholar] [CrossRef]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques; Churchill Livingstone Elsevier: Oxford, UK, 2013. [Google Scholar]
- Tuglu, D.; Yuvanc, E.; Yilmaz, E.; Gencay, I.Y.; Atasoy, P.; Kisa, U.; Batislam, E. The antioxidant effect of dexmedetomidine on testicular ischemia-reperfusion injury. Acta Cir. Bras. 2015, 30, 414–421. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups (dose: mg/kg) | Relative Testicular Weight | Serum Testosterone Level (ng/mL) | Sperm Count (×106/mm3) | Sperm Motility (%) | Abnormal Sperm Morphology (%) |
|---|---|---|---|---|---|
| Control | 0.69 ± 0.044 | 2.49 ± 0.059 | 38.50 ± 0.71 | 72.67 ± 1.53 | 12.67 ± 1.52 |
| TDF 5 | 0.63 ± 0.027 | 2.58 ± 0.031 | 38.00 ± 1.00 | 68.00 ± 1.41 | 14.00 ± 1.00 |
| PTX 75 | 0.65 ± 0.033 | 2.63 ± 0.042 | 35.00 ± 2.65 | 68.50 ± 0.71 | 15.00 ± 2.00 |
| Cilo 20 | 0.65 ± 0.069 | 2.74 ± 0.042 | 35.33 ± 1.15 | 68.00 ± 1.40 | 15.50 ± 0.71 |
| CIS | 0.49 ± 0.028 * | 0.93 ± 0.051 * | 13.66 ± 1.53 * | 32.67 ± 1.52 * | 35.00 ± 1.41 * |
| CIS + TDF | 0.64 ± 0.027 # | 1.15 ± 0.059 # | 20.66 ± 0.58 # | 38.00 ± 1.00 # | 27.67 ± 0.57 # |
| CIS + PTX | 0.73 ± 0.064 # | 1.46 ± 0.044 #,! | 23.00 ± 1.00 # | 41.00 ± 1.00 # | 24.67 ± 1.53 # |
| CIS + Cilo 5 | 0.73 ± 0.027 # | 1.71 ± 0.011 # | 25.50 ± 0.71 # | 42.67 ± 1.51 # | 23.50 ± 0.71 # |
| CIS + Cilo 10 | 0.75 ± 0.061 # | 1.84 ± 0.040 #,● | 26.66 ± 1.52 # | 49.00 ± 2.00 #,● | 19.00 ± 1.00 #,● |
| CIS + Cilo 20 | 0.70 ± 0.098 # | 1.97 ± 0.020 #,●,†,!,@ | 31.50 ± 0.70 #,●,†,!,@ | 59.67 ± 1.52 #,●,†,!,@ | 19.00 ± 0.99 #,●,!,@ |
| Lesion Groups (dose: mg/kg) | Desquamation in Germinal Cells | Disorganization in Germinal Cells | Interstitial Edema | Degeneration in Germinal Cells | Reduction in Germinal Cell Counts |
|---|---|---|---|---|---|
| Control | - | - | - | - | - |
| TDF 5 | - | - | - | - | - |
| PTX 75 | - | - | - | - | - |
| Cilo 20 | - | - | - | - | - |
| CIS 7 | ++ | ++ | + | ++ | ++ |
| CIS + TDF | ++ | + | + | ++ | + |
| CIS + PTX | + | + | ++ | + | + |
| CIS + Cilo 5 | ++ | ++ | + | + | + |
| CIS + Cilo 10 | ++ | ++ | + | + | + |
| CIS + Cilo 20 | - | - | + | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othman, E.M.; Habib, H.A.; Zahran, M.E.; Amin, A.; Heeba, G.H. Mechanistic Protective Effect of Cilostazol in Cisplatin-Induced Testicular Damage via Regulation of Oxidative Stress and TNF-α/NF-κB/Caspase-3 Pathways. Int. J. Mol. Sci. 2023, 24, 12651. https://doi.org/10.3390/ijms241612651
Othman EM, Habib HA, Zahran ME, Amin A, Heeba GH. Mechanistic Protective Effect of Cilostazol in Cisplatin-Induced Testicular Damage via Regulation of Oxidative Stress and TNF-α/NF-κB/Caspase-3 Pathways. International Journal of Molecular Sciences. 2023; 24(16):12651. https://doi.org/10.3390/ijms241612651
Chicago/Turabian StyleOthman, Eman M., Heba A. Habib, Mahmoud E. Zahran, Amr Amin, and Gehan H. Heeba. 2023. "Mechanistic Protective Effect of Cilostazol in Cisplatin-Induced Testicular Damage via Regulation of Oxidative Stress and TNF-α/NF-κB/Caspase-3 Pathways" International Journal of Molecular Sciences 24, no. 16: 12651. https://doi.org/10.3390/ijms241612651
APA StyleOthman, E. M., Habib, H. A., Zahran, M. E., Amin, A., & Heeba, G. H. (2023). Mechanistic Protective Effect of Cilostazol in Cisplatin-Induced Testicular Damage via Regulation of Oxidative Stress and TNF-α/NF-κB/Caspase-3 Pathways. International Journal of Molecular Sciences, 24(16), 12651. https://doi.org/10.3390/ijms241612651