
Cytokine Kinetics during Progression of COVID-19 in Rwanda Patients: Could IL-9/IFNγ Ratio Predict Disease Severity?

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Patients Characteristics
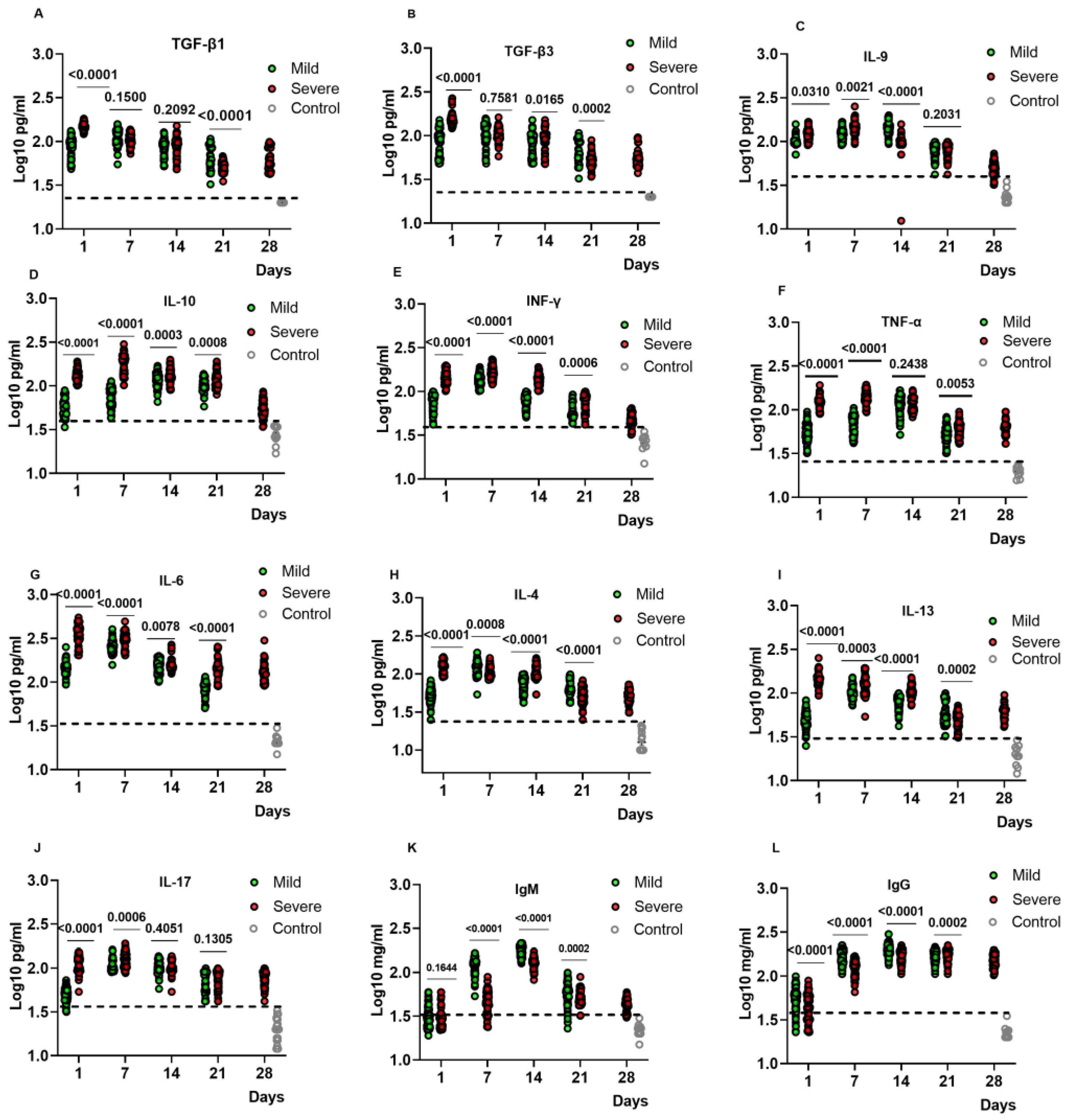
2.2. Kinetic Analysis of Cytokine and Antibody Levels in the Plasma of COVID-19 Patients
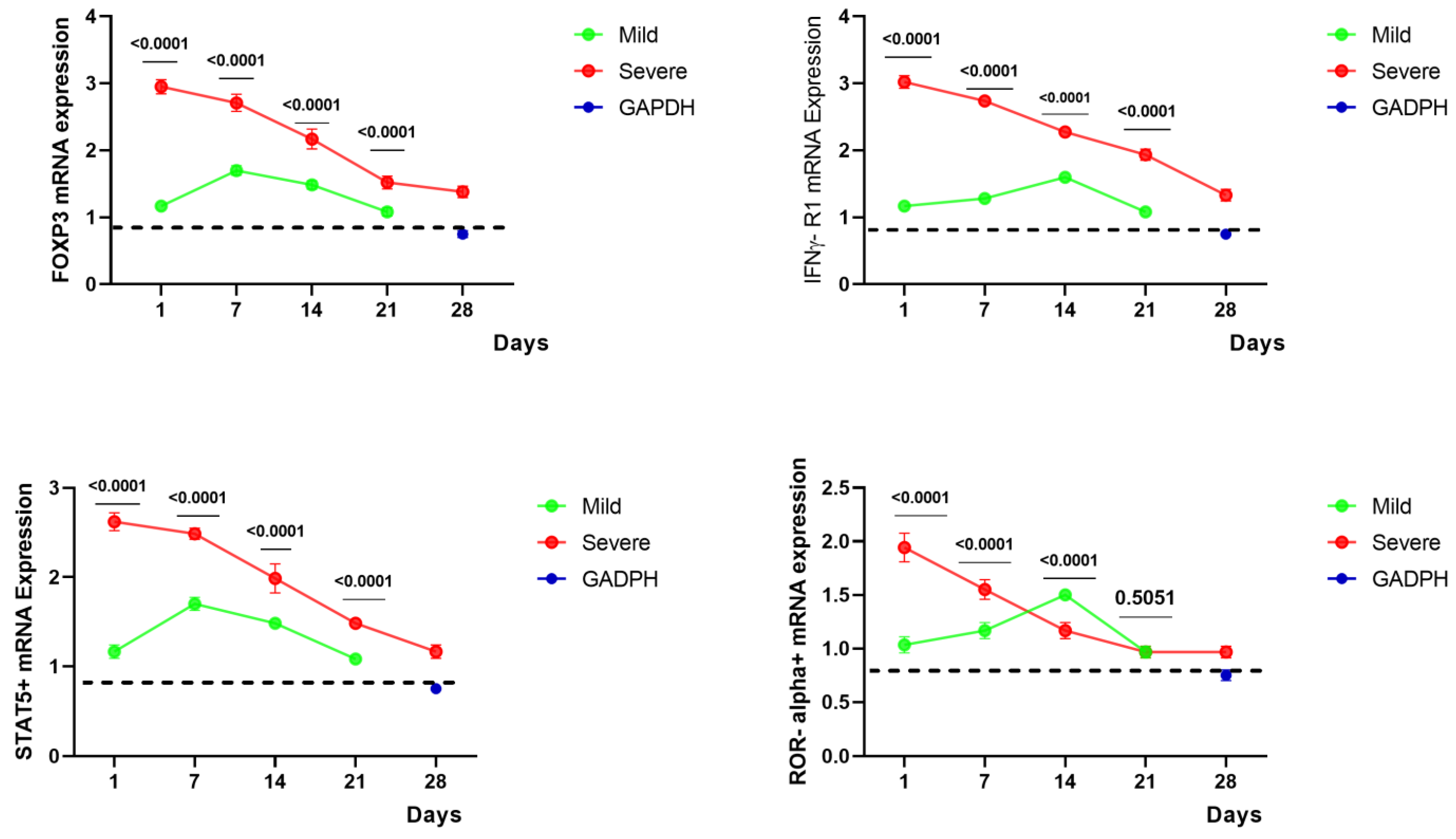
2.3. Gene Expressions of FoxP3, STAT5+, IFNγ-R1, and ROR Alpha+ of COVID-19 Patients
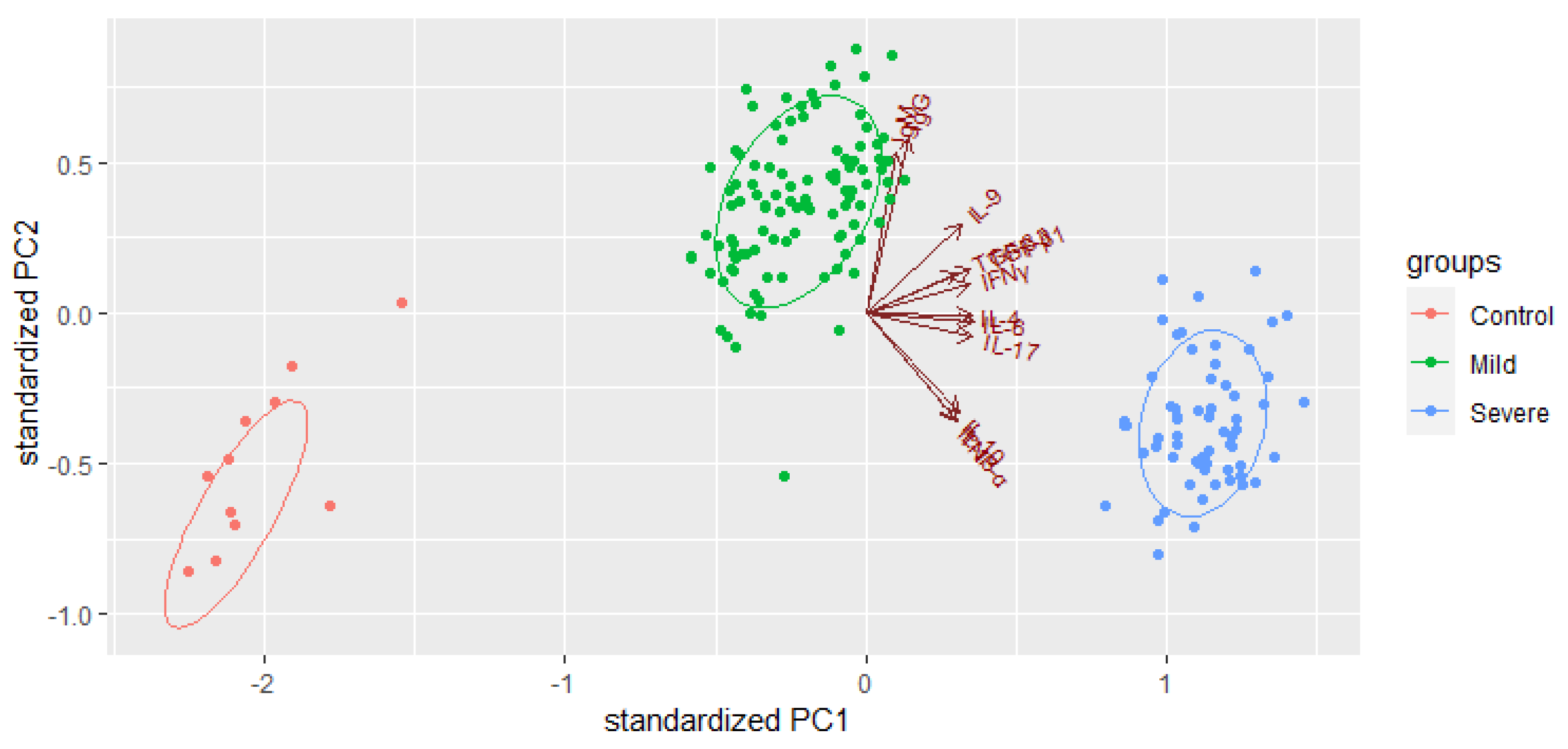
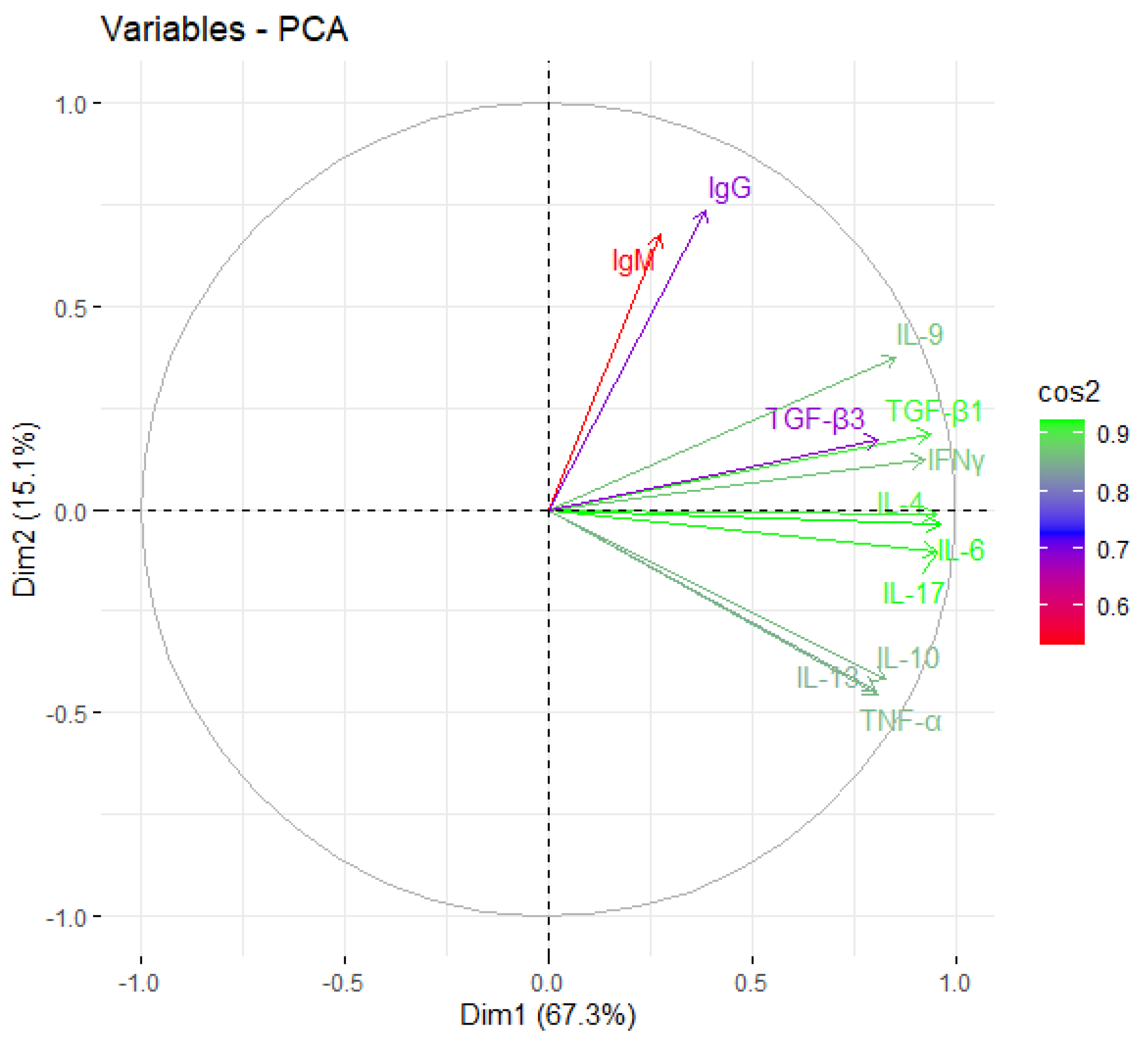
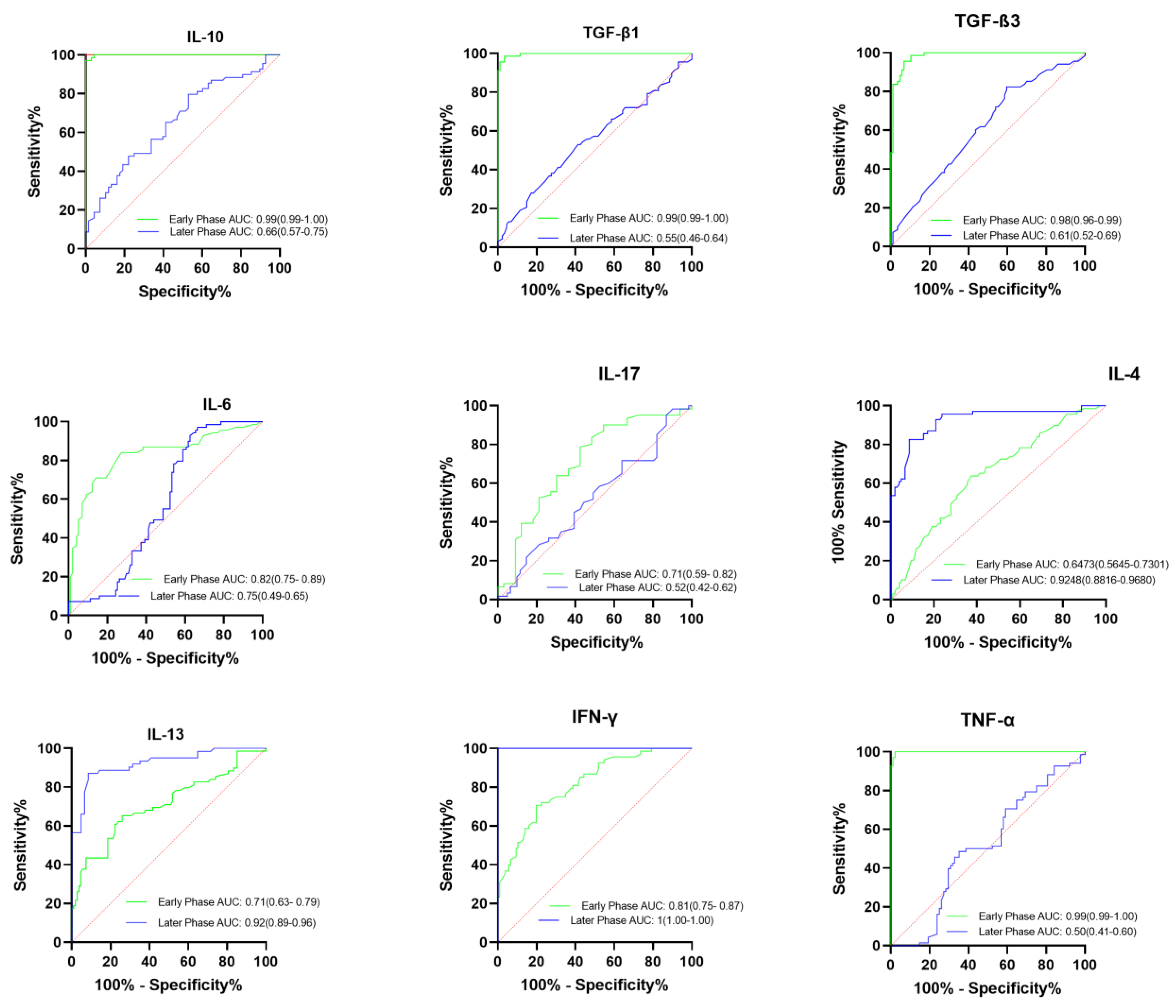
2.4. Prognostic Factors for the Identification of Severe COVID-19 Cases
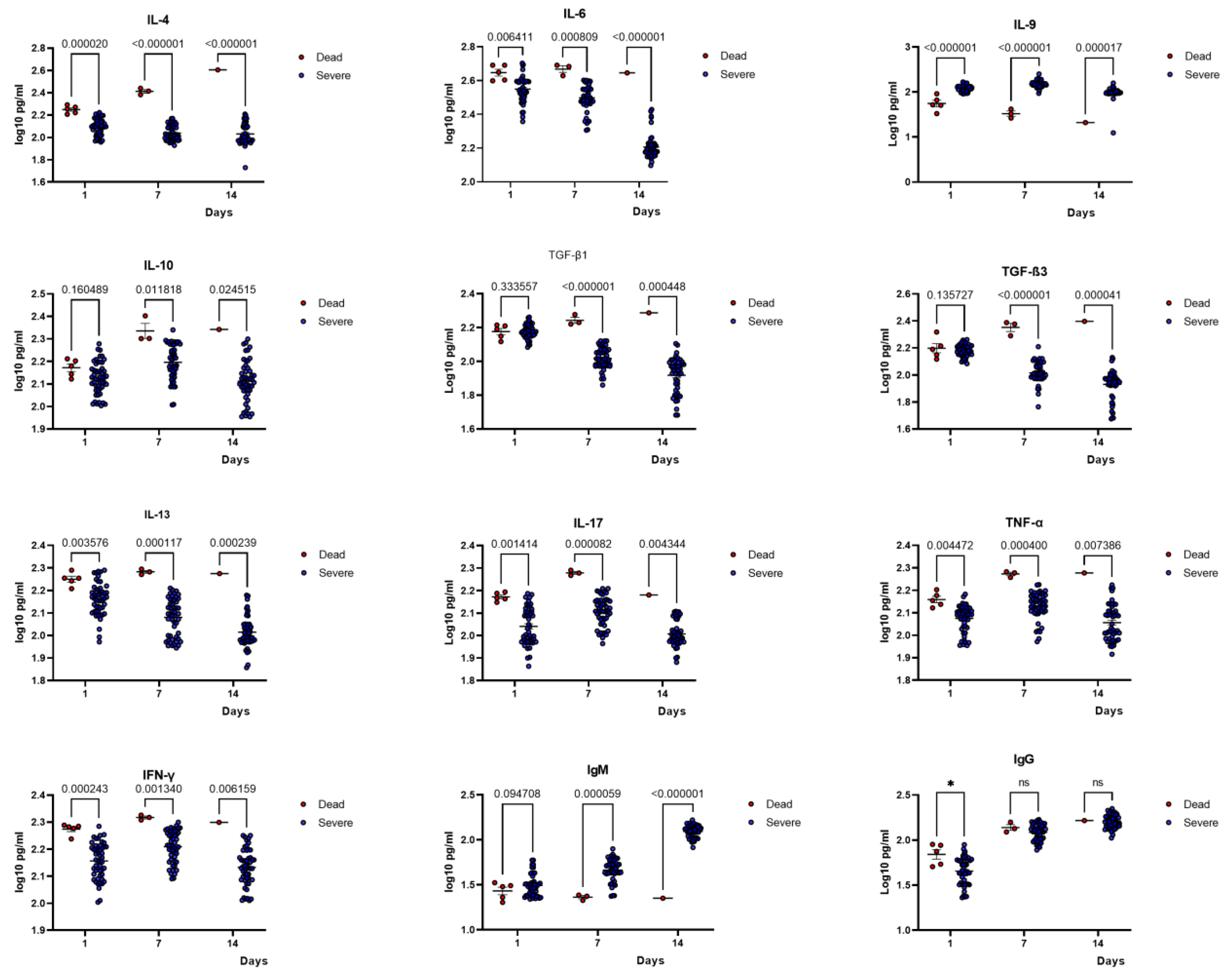
2.5. Comparison of Cytokine Profiles of Patients with Severe COVID-19 and the Deceased
2.6. The Ratio IFNγ/IL-9 and Disease Severity/Death
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Severity of COVID-19
4.3. Testing SARS-CoV-2
4.4. Cytokine Profile Using ELISA
4.5. Gene Expression Evaluation
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adimulam, T.; Arumugam, T.; Gokul, A.; Ramsuran, V. Genetic Variants within SARS-CoV-2 Human Receptor Genes May Contribute to Variable Disease Outcomes in Different Ethnicities. Int. J. Mol. Sci. 2023, 24, 8711. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, J.; Mathie, D.; Scott, F.; Mahendran, Y.; Woolhouse, M.; Stoevesandt, O.; Mduluza, T.; Mutapi, F. Peptide microarray IgM and IgG screening of pre-SARS-CoV-2 human serum samples from Zimbabwe for reactivity with peptides from all seven human coronaviruses: A cross-sectional study. Lancet 2023, 4, E215–E227. [Google Scholar] [CrossRef]
- Aissatou, A.; Fokam, J.; Semengue, E.N.J.; Takou, D.; Ka’e, A.C.; Ambe, C.C.; Nka, A.D.; Djupsa, S.C.; Beloumou, G.; Ciaffi, L.; et al. Pre-existing immunity to SARS-CoV-2 before the COVID-19 pandemic era in Cameroon: A comparative analysis according to HIV-status. Front. Immunol. 2023, 14, 1162. [Google Scholar] [CrossRef] [PubMed]
- Wachira, L.-J.; Arena, R.; Sallis, J.F.; Lambert, E.V.; Ong’Wen, O.M.; Laddu, D.R.; Onywera, V.; Oyeyemi, A.L. Why are COVID-19 effects less severe in Sub-Saharan Africa? Moving more and sitting less may be a primary reason. Prog. Cardiovasc. Dis. 2022, 71, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Wamai, R.G.; Hirsch, J.L.; Van Damme, W.; Alnwick, D.; Bailey, R.C.; Hodgins, S.; Alam, U.; Anyona, M. What Could Explain the Lower COVID-19 Burden in Africa despite Considerable Circulation of the SARS-CoV-2 Virus? Int. J. Environ. Res. Public Health 2021, 18, 8638. [Google Scholar] [CrossRef]
- Guimarães, P.O.; Quirk, D.; Furtado, R.H.; Maia, L.N.; Saraiva, J.F.; Antunes, M.O.; Filho, R.K.; Junior, V.M.; Soeiro, A.M.; Tognon, A.P.; et al. Tofacitinib in Patients Hospitalized with Covid-19 Pneumonia. New Engl. J. Med. 2021, 385, 406–415. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EbioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Wu, J.; Liu, J.; Zhao, X.; Liu, C.; Wang, W.; Wang, D.; Xu, W.; Zhang, C.; Yu, J.; Jiang, B.; et al. Clinical Characteristics of Imported Cases of Coronavirus Disease 2019 (COVID-19) in Jiangsu Province: A Multicenter Descriptive Study. Clin. Infect. Dis. 2020, 71, 706–712. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef]
- Cabaro, S.; D’esposito, V.; Di Matola, T.; Sale, S.; Cennamo, M.; Terracciano, D.; Parisi, V.; Oriente, F.; Portella, G.; Beguinot, F.; et al. Cytokine signature and COVID-19 prediction models in the two waves of pandemics. Sci. Rep. 2021, 11, 20793. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.; Prezzemolo, T.; Vanderbeke, L.; Roca, C.P.; Gerbaux, M.; Janssens, S.; Willemsen, M.; Burton, O.; Van Mol, P.; Van Herck, Y.; et al. Increased IL-10-producing regulatory T cells are characteristic of severe cases of COVID-19. Clin. Transl. Immunol. 2020, 9, e1204. [Google Scholar] [CrossRef]
- Donlan, A.N.; Sutherland, T.E.; Marie, C.; Preissner, S.; Bradley, B.T.; Carpenter, R.M.; Sturek, J.M.; Ma, J.Z.; Moreau, G.B.; Donowitz, J.R.; et al. Title: IL-13 is a driver of COVID-19 severity. JCI Insight. 2021, 6, e150107. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Gomes, M.; Kruglov, A.; Durek, P.; Heinrich, F.; Tizian, C.; Heinz, G.A.; Pascual-Reguant, A.; Du, W.; Mothes, R.; Fan, C.; et al. SARS-CoV-2 in severe COVID-19 induces a TGF-β-dominated chronic immune response that does not target itself. Nat. Commun. 2021, 12, 1961. [Google Scholar] [CrossRef]
- Sun, X.; Wang, T.; Cai, D.; Hu, Z.; Chen, J.; Liao, H.; Zhi, L.; Wei, H.; Zhang, Z.; Qiu, Y.; et al. Cytokine storm intervention in the early stages of COVID-19 pneumonia. Cytokine Growth Factor. Rev. 2020, 53, 38–42. [Google Scholar] [CrossRef]
- Bertoletti, A.; Le Bert, N.; Tan, A.T. Act Early and at the Right Location: SARS-CoV-2 T Cell Kinetics and Tissue Localization. Int. J. Mol. Sci. 2022, 23, 679. [Google Scholar] [CrossRef]
- Carrascosa, L.C.; Klein, M.; Kitagawa, Y.; Lückel, C.; Marini, F.; König, A.; Guralnik, A.; Raifer, H.; Hagner-Benes, S.; Rädler, D.; et al. Reciprocal regulation of the Il9 locus by counteracting activities of transcription factors IRF1 and IRF4. Nat. Commun. 2017, 8, 15366. [Google Scholar] [CrossRef]
- Kim, M.-H.; Salloum, S.; Wang, J.Y.; Ping, W.L.; Regan, J.; Lefteri, K.; Manickas-Hill, Z.; Gao, C.; Li, J.Z.; I Sadreyev, R.; et al. Type I, II, and III Interferon Signatures Correspond to Coronavirus Disease 2019 Severity. J. Infect. Dis. 2021, 224, 777–782. [Google Scholar] [CrossRef]
- Murugaiyan, G.; Beynon, V.; Da Cunha, A.P.; Joller, N.; Weiner, H.L. IFN-γ Limits Th9-Mediated Autoimmune Inflammation through Dendritic Cell Modulation of IL-27. J. Immunol. 2012, 189, 5277–5283. [Google Scholar] [CrossRef]
- Silva, N.S.D.L.; Orikaza, C.M.; Santana, F.R.D.; Dos Santos, L.A.; Salu, B.R.; Oliva, M.L.V.; Sinigaglia, R.d.C.; Mortara, R.A. Interleukin-9 in Immunopathology of Trypanosoma cruzi Experimental Infection. Front. Cell. Infect. Microbiol. 2021, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Ndoricyimpaye, E.L.; Van Snick, J.; Niyoyita, J.d.D.; Kanimba, P.; Mbonimpa, J.B.; Rutayisire, R.; Rutayisire, R.; Ndahindwa, V.; Cheou, P.; Coutelier, J.P.; et al. Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation. Int. J. Mol. Sci. 2022, 23, 12665. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.J.; Ohtsuki, T.; Kohno, K.; Tanabe, F.; Ushio, S.; Namba, M.; Tanimoto, T.; Torigoe, K.; Fujii, M.; Ikeda, M.; et al. Interferon-gamma-inducing factor enhances T helper 1 cytokine production by stimulated human T cells: Synergism with interleukin-12 for interferon-gamma production. Eur. J. Immunol. 1996, 26, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Mulchandani, R.; Lyngdoh, T.; Kakkar, A.K. Deciphering the COVID-19 cytokine storm: Systematic review and meta-analysis. Eur. J. Clin. Investig. 2021, 51, e13429. [Google Scholar] [CrossRef] [PubMed]
- Noël, W.; Raes, G.; Ghassabeh, G.H.; De Baetselier, P.; Beschin, A. Alternatively activated macrophages during parasite infections. Trends Parasitol. 2004, 20, 126–133. [Google Scholar] [CrossRef]
- Wollenberg, A.; Flohr, C.; Simon, D.; Cork, M.J.; Thyssen, J.P.; Bieber, T.; Bruin-Weller, M.; Weidinger, S.; Deleuran, M.; Taieb, A.; et al. European Task Force on Atopic Dermatitis statement on severe acute respiratory syndrome coronavirus 2 (SARS-Cov-2) infection and atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2020, 34, e241–e242. [Google Scholar] [CrossRef]
- Ji, J.-D.; Tassiulas, I.; Park-Min, K.-H.; Aydin, A.; Mecklenbräuker, I.; Tarakhovsky, A.; Pricop, L.; Salmon, J.E.; Ivashkiv, L.B. Inhibition of Interleukin 10 Signaling after Fc Receptor Ligation and during Rheumatoid Arthritis. J. Exp. Med. 2003, 197, 1573–1583. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, H.; Dauphars, D.J.; He, Y.-W. A Potential Role of Interleukin 10 in COVID-19 Pathogenesis. Trends Immunol. 2020, 42, 3–5. [Google Scholar] [CrossRef]
- Cecere, T.E.; Todd, S.M.; LeRoith, T. Regulatory T cells in arterivirus and coronavirus infections: Do they protect against disease or enhance it? Viruses 2012, 4, 833–846. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Jiang, L.; Dua, K.; Hansbro, P.M.; Liu, G. SARS-CoV-2 induces transcriptional signatures in human lung epithelial cells that promote lung fibrosis. Respir. Res. 2020, 21, 182. [Google Scholar] [CrossRef]
- El-Din, A.N.; Ata, K.A.E.S.; Abdel-Gawad, A.R.; Fahmy, N.F. Impact of high serum levels of mmp-7, mmp-9, tgf-β and pdgf macrophage activation markers on severity of covid-19 in obese-diabetic patients. Infect. Drug. Resist. 2021, 14, 4015–4025. [Google Scholar] [CrossRef] [PubMed]
- Basheer, M.; Saad, E.; Kananeh, M.; Asad, L.; Khayat, O.; Badarne, A.; Abdo, Z.; Arraf, N.; Milhem, F.; Bassal, T.; et al. Cytokine Patterns in COVID-19 Patients: Which Cytokines Predict Mortality and Which Protect Against? Curr. Issues Mol. Biol. 2022, 44, 4735–4747. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Hoehn, P.; Germann, T.; Rüde, E. Differential effects of interleukin-12 on the development of I mouse CD4+ T cells. Eur. J. Immunol. 1994, 24, 343–347. [Google Scholar] [CrossRef]
- Li u, Y.; Yan g, Y.; Zhan g, C.; Huan g, F.; Wan g, F.; Yua n, J.; Wan g, Z.; L i, J.; L i, J.; Fen g, C.; et al. Clinical and biochemical indexes from 2019-nCoV infected patients linked to viral loads and lung injury. Sci. China Life Sci. 2020, 63, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFβ in the Context of an Inflammatory Cytokine Milieu Supports De Novo Differentiation of IL-17-Producing T Cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Jin, Y.-H.; Kang, H.S.; Kim, B.S. Interleukin-6 (IL-6) and IL-17 Synergistically Promote Viral Persistence by Inhibiting Cellular Apoptosis and Cytotoxic T Cell Function. J. Virol. 2014, 88, 8479. [Google Scholar] [CrossRef]
- Todorov, I.; Gospodinova, M.; Bocheva, Y.; Popcheva, G. Serum amyloid A protein in the course of infectious mononucleosis. Adv. Infect. Dis. 2019, 6, 204993611881120. [Google Scholar] [CrossRef]
- Xie, N.; Li, Z.; Zuo, R.; Qi, S.; Zhu, T.; Liu, L.; Wan, L.; Yuan, J. Serum SAA1 and APOE are novel indicators for human cytomegalovirus infection. Sci. Rep. 2017, 7, 13407. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Zhang, X.X.; Wu, Y.J.; Zeng, Z.P.; She, W.M.; Chen, S.Y.; Zhang, Y.; Guo, J. Serum amyloid A levels in patients with liver diseases. World J. Gastroenterol. 2019, 25, 6440–6450. [Google Scholar] [CrossRef]
- Ather, J.L.; Dienz, O.; Boyson, J.E.; Anathy, V.; Amiel, E.; Poynter, M.E. Serum Amyloid A3 is required for normal lung development and survival following influenza infection. Sci. Rep. 2018, 8, 16571. [Google Scholar] [CrossRef]
- Laloglu, E.; Alay, H. Role of transforming growth factor-beta 1 and connective tissue growth factor levels in coronavirus disease-2019-related lung Injury: A prospective, observational, cohort study. Rev. Soc. Bras. Med. Trop. 2022, 55, e06152021. [Google Scholar] [CrossRef] [PubMed]
- Elshazli, R.M.; Toraih, E.A.; Elgaml, A.; El-Mowafy, M.; El-Mesery, M.; Amin, M.N.; Hussein, M.H.; Killackey, M.T.; Fawzy, M.S.; Kandil, E. Diagnostic and prognostic value of hematological and immunological markers in COVID-19 infection: A meta-analysis of 6320 patients. PLoS One 2020, 15, e0238160. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.A.; Li, A.P.Y.; Hachim, A.; Hui, D.S.C.; Kwan, M.Y.W.; Tsang, O.T.Y.; Chiu, S.S.; Chan, W.H.; Yau, Y.S.; Kavian, N.; et al. SARS-CoV-2 specific T cell responses are lower in children and increase with age and time after infection. Nat. Commun. 2021, 12, 4678. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, Y.; Chen, H.; Meng, J.; Li, J.; Chu, J.; Zheng, R.; Wang, H.; Pan, P.; Su, J.; et al. Weighted gene co-expression network analysis revealed T cell differentiation associated with the age-related phenotypes in COVID-19 patients. BMC Med. Genom. 2023, 16, 59. [Google Scholar] [CrossRef]
- Abdelhafiz, A.S.; Fouad, M.A.; Sayed-Ahmed, M.M.; Kamel, M.M.; Ali, A.; Fouda, M.; Khalil, M.A.; Abdel-Moneim, A.S.; Kamal, L.M. Upregulation of FOXP3 is associated with severity of hypoxia and poor outcomes in COVID-19 patients. Virology 2021, 563, 74–81. [Google Scholar] [CrossRef]
- Stephen-Victor, E.; Das, M.; Karnam, A.; Pitard, B.; Gautier, J.F.; Bayry, J. Potential of regulatory T-cell-based therapies in the management of severe COVID-19. Eur. Respir J. 2020, 56, 2002182. [Google Scholar] [CrossRef]
- Zhao, Y.; Kilian, C.; Turner, J.E.; Bosurgi, L.; Roedl, K.; Bartsch, P.; Gnirck, A.-C.; Cortesi, F.; Schultheiß, C.; Hellmig, M.; et al. Clonal expansion and activation of tissue-resident memory-like TH17 cells expressing GM-CSF in the lungs of patients with severe COVID-19. Sci. Immunol. 2021, 6, 6692. [Google Scholar] [CrossRef]
- Hassaniazad, M.; Eftekhar, E.; Inchehsablagh, B.R.; Kamali, H.; Tousi, A.; Jaafari, M.R.; Rafat, M.; Fathalipour, M.; Nikoofal-Sahlabadi, S.; Gouklani, H.; et al. A triple-blind, placebo-controlled, randomized clinical trial to evaluate the effect of curcumin-containing nanomicelles on cellular immune responses subtypes and clinical outcome in COVID-19 patients. Phytother. Res. 2021, 35, 6417. [Google Scholar] [CrossRef]
- Elyaman, W.; Bradshaw, E.M.; Uyttenhove, C.; Dardalhon, V.; Awasthi, A.; Imitola, J.; Bettelli, E.; Oukka, M.; van Snick, J.; Renauld, J.-C.; et al. IL-9 induces differentiation of TH17 cells and enhances function of FoxP3+ natural regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 12885–12890. [Google Scholar] [CrossRef]
- Lui, G.; Ling, L.; Lai, C.K.; Tso, E.Y.; Fung, K.S.; Chan, V.; Ho, T.H.Y.; Luk, F.; Chen, Z.; Ng, J.K.C.; et al. Viral dynamics of SARS-CoV-2 across a spectrum of disease severity in COVID-19. J. Infect. 2020, 81, 318. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antigen-Detection in the Diagnosis of SARS-CoV-2 Infection Using Rapid Immunoassays Interim Guidance. 2020. Available online: https://apps.who.int/iris/handle/10665/334253 (accessed on 20 November 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 20) | Mild (n = 129) | Severe (n = 68) | * p-Value | |
|---|---|---|---|---|
| Age (years-median (IQR)) | 34 (23–44) | 34 (26–44) | 42 (31–70) | <0.0001 |
| Gender (F/M) | 14/6 | 47/82 | 42/26 | - |
| Presence of comorbidities | 0 (0) | 21 (16.28) | 9 (13.43) | 0.5996 |
| Oxygen Therapy-no. (%) | 0 (0) | 0 | 68 (100) | <0.0001 |
| Mechanical Ventilation-no. (%) | 0 (0) | 0 (0) | 53 (78) | <0.0001 |
| Analgesic and antipyretic-no. (%) | 0 (0) | 97 (75) | 10 (15) | <0.0001 |
| Steroids-no. (%) | 0 (0) | 30 (23) | 59 (87) | 0.065 |
| Remdesivir-no. (%) | 0 (0) | 25 (19) | 60 (88) | 0.0636 |
| Antibiotics-no. (%) | 0 (0) | 80 (62) | 40 (59) | 0.050 |
| Hospital Mortality-no. (%) | 0 (0) | 0 (0) | 5 (7.46) | 0.004 |
| Variables | Mean | Std. Deviation | Coefficient of Dispersion | Coefficient of Variation | T-Test Ratio |
|---|---|---|---|---|---|
| Mean Centered | |||||
| Control | 0.842 | 0.064 | 0.028 | 6.8% | |
| Mild | 0.832 | 0.053 | 0.045 | 5.7% | |
| Severe | 1.002 | 0.045 | 0.033 | 4.5% | T = 8.595, p = 0.003 |
| Overall | 0.957 | 0.061 | 0.050 | 6.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndoricyimpaye, E.L.; Van Snick, J.; Robert, R.; Bikorimana, E.; Majyambere, O.; Mukantwari, E.; Nshimiyimana, T.; Mbonigaba, V.; Coutelier, J.P.; Rujeni, N. Cytokine Kinetics during Progression of COVID-19 in Rwanda Patients: Could IL-9/IFNγ Ratio Predict Disease Severity? Int. J. Mol. Sci. 2023, 24, 12272. https://doi.org/10.3390/ijms241512272
Ndoricyimpaye EL, Van Snick J, Robert R, Bikorimana E, Majyambere O, Mukantwari E, Nshimiyimana T, Mbonigaba V, Coutelier JP, Rujeni N. Cytokine Kinetics during Progression of COVID-19 in Rwanda Patients: Could IL-9/IFNγ Ratio Predict Disease Severity? International Journal of Molecular Sciences. 2023; 24(15):12272. https://doi.org/10.3390/ijms241512272
Chicago/Turabian StyleNdoricyimpaye, Ella Larissa, Jacques Van Snick, Rutayisire Robert, Emmanuel Bikorimana, Onesphore Majyambere, Enatha Mukantwari, Thaddée Nshimiyimana, Valens Mbonigaba, Jean Paul Coutelier, and Nadine Rujeni. 2023. "Cytokine Kinetics during Progression of COVID-19 in Rwanda Patients: Could IL-9/IFNγ Ratio Predict Disease Severity?" International Journal of Molecular Sciences 24, no. 15: 12272. https://doi.org/10.3390/ijms241512272
APA StyleNdoricyimpaye, E. L., Van Snick, J., Robert, R., Bikorimana, E., Majyambere, O., Mukantwari, E., Nshimiyimana, T., Mbonigaba, V., Coutelier, J. P., & Rujeni, N. (2023). Cytokine Kinetics during Progression of COVID-19 in Rwanda Patients: Could IL-9/IFNγ Ratio Predict Disease Severity? International Journal of Molecular Sciences, 24(15), 12272. https://doi.org/10.3390/ijms241512272