
Identification of a Putative CFSH Receptor Inhibiting IAG Expression in Crabs



Abstract
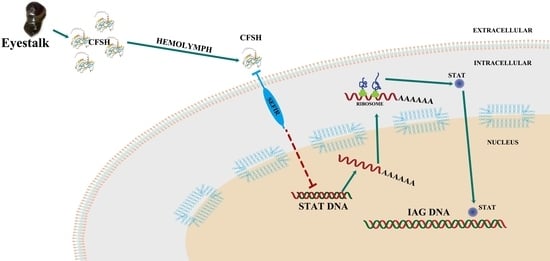
1. Introduction
2. Results
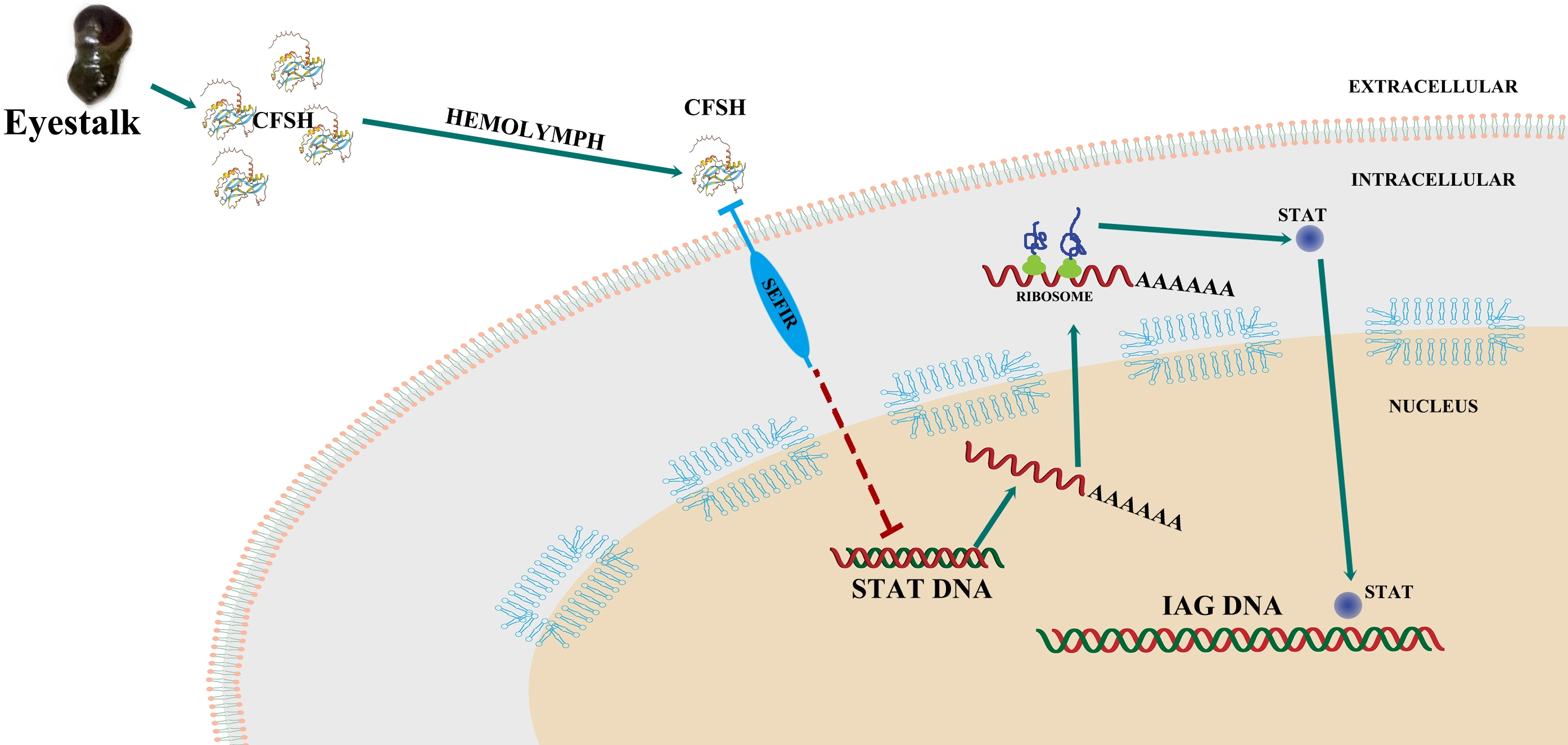
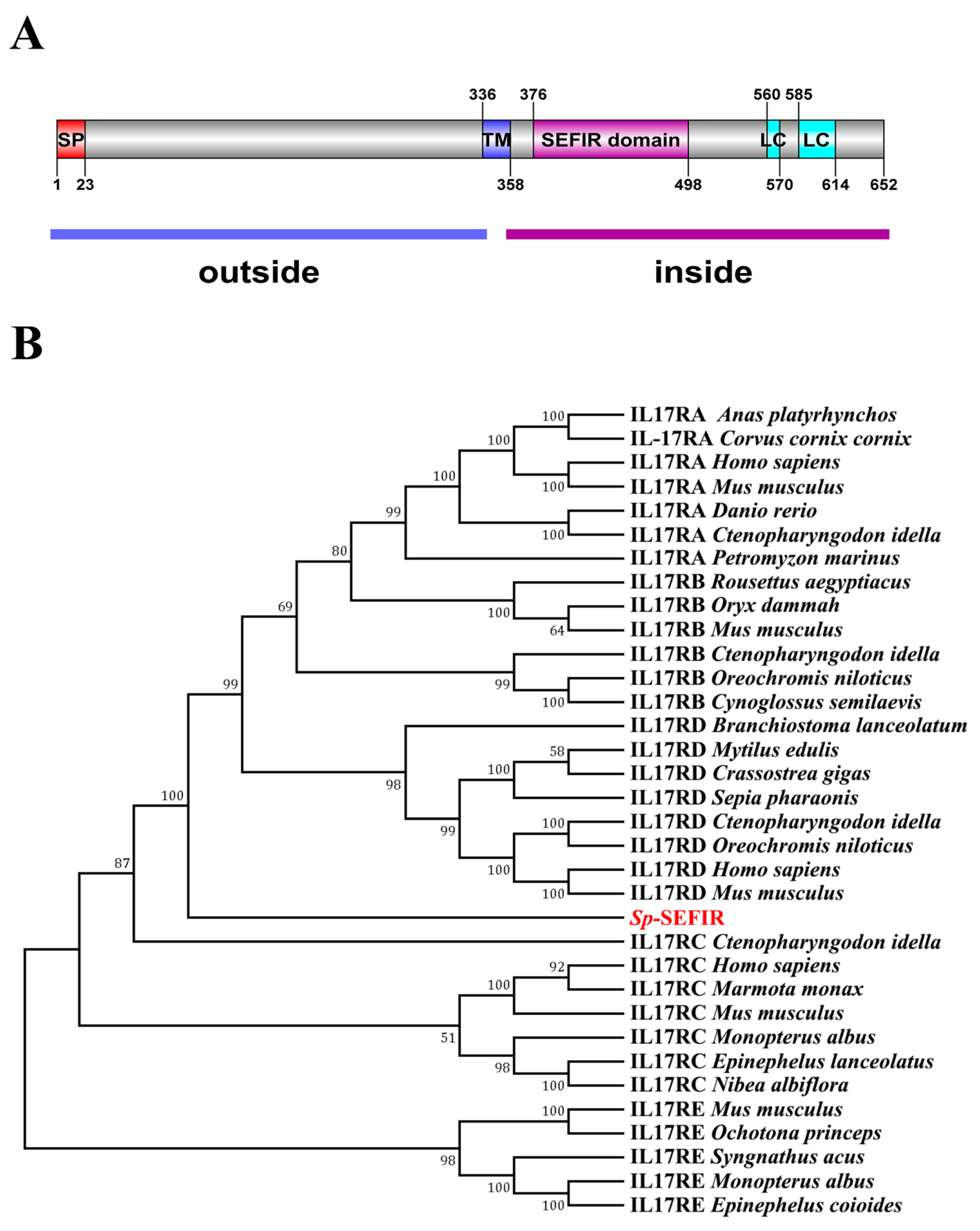
2.1. Molecular Cloning and Phylogenetics of Sp-SEFIR
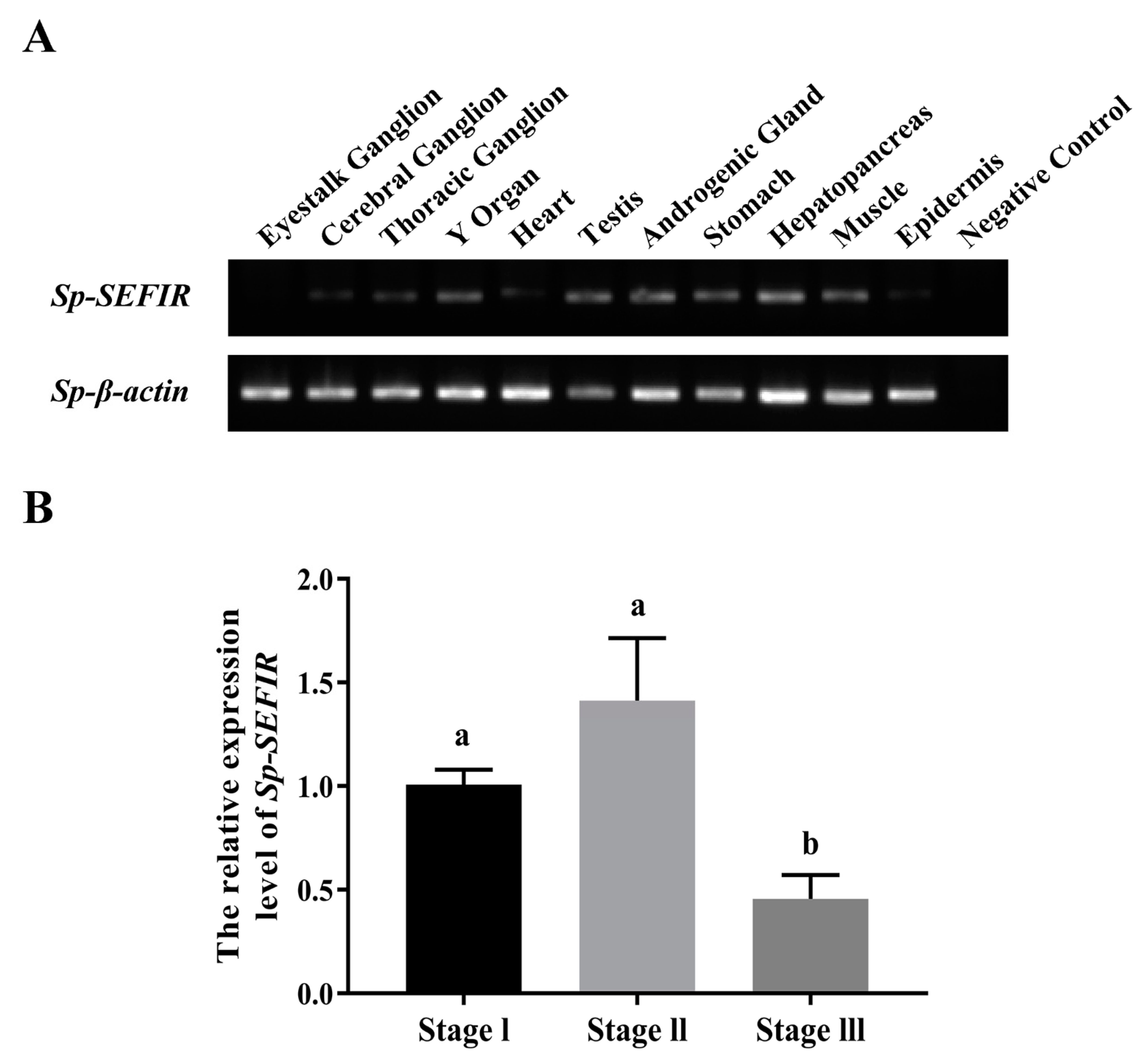
2.2. Tissue Distribution of Sp-SEFIR
2.3. Expression Profile of Sp-SEFIR during AG Development
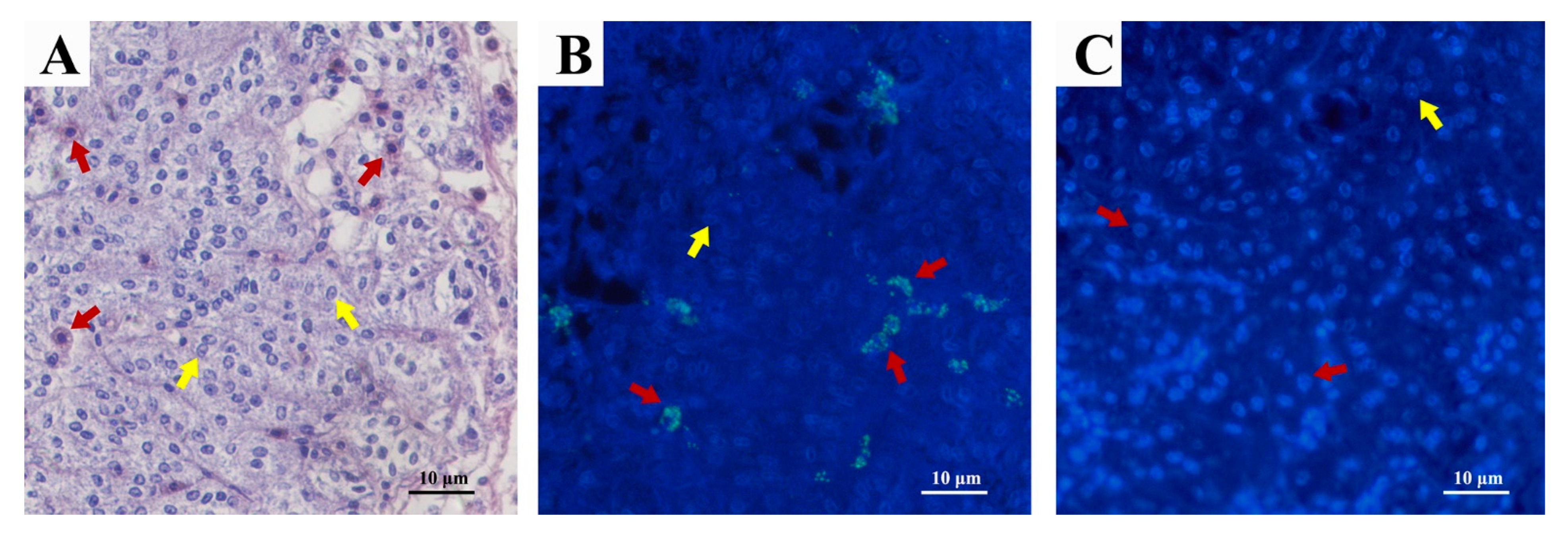
2.4. Immunofluorescence Localization of Sp-SEFIR in AG
2.5. Ligand–Receptor Interaction Analysis
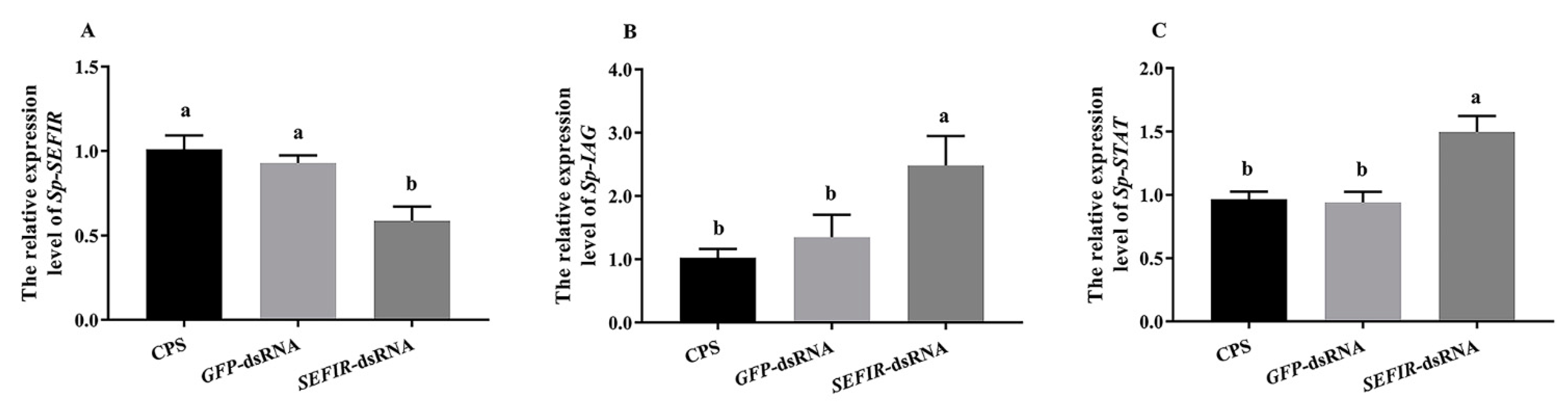
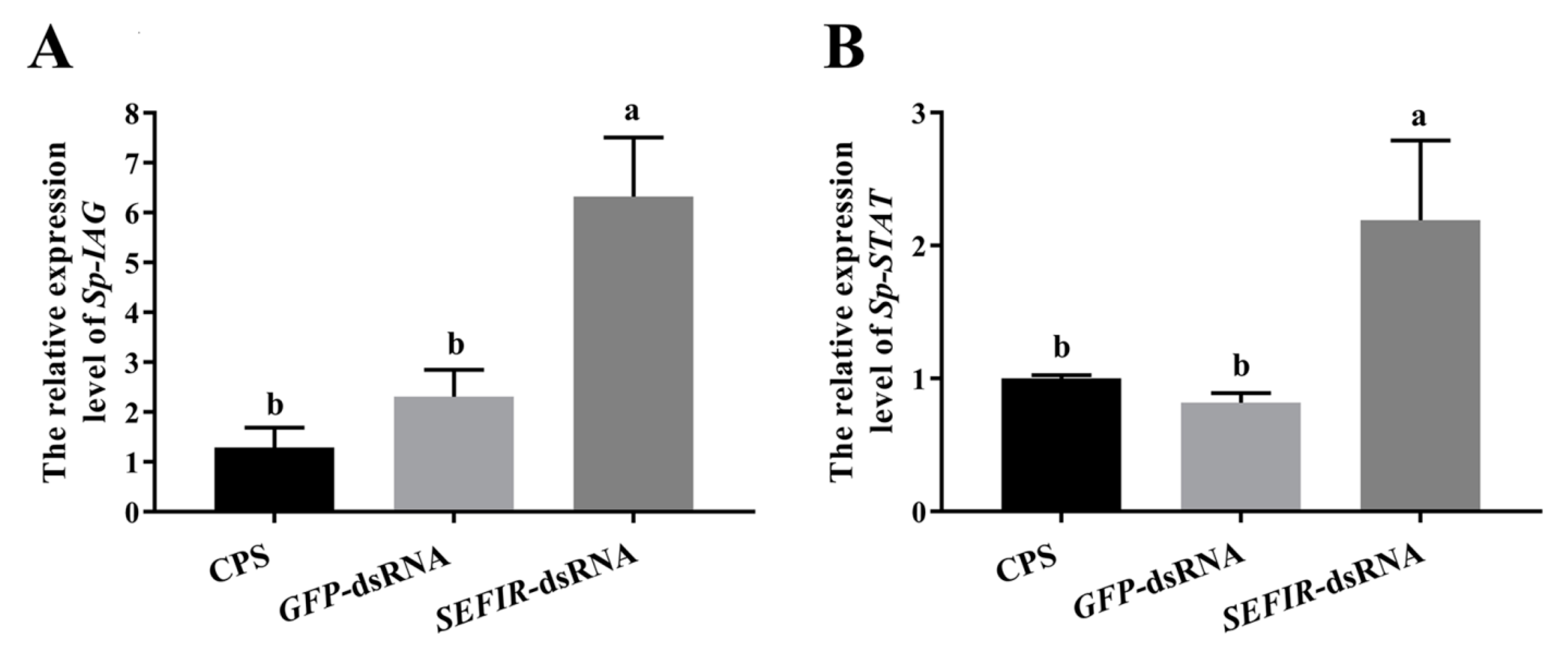
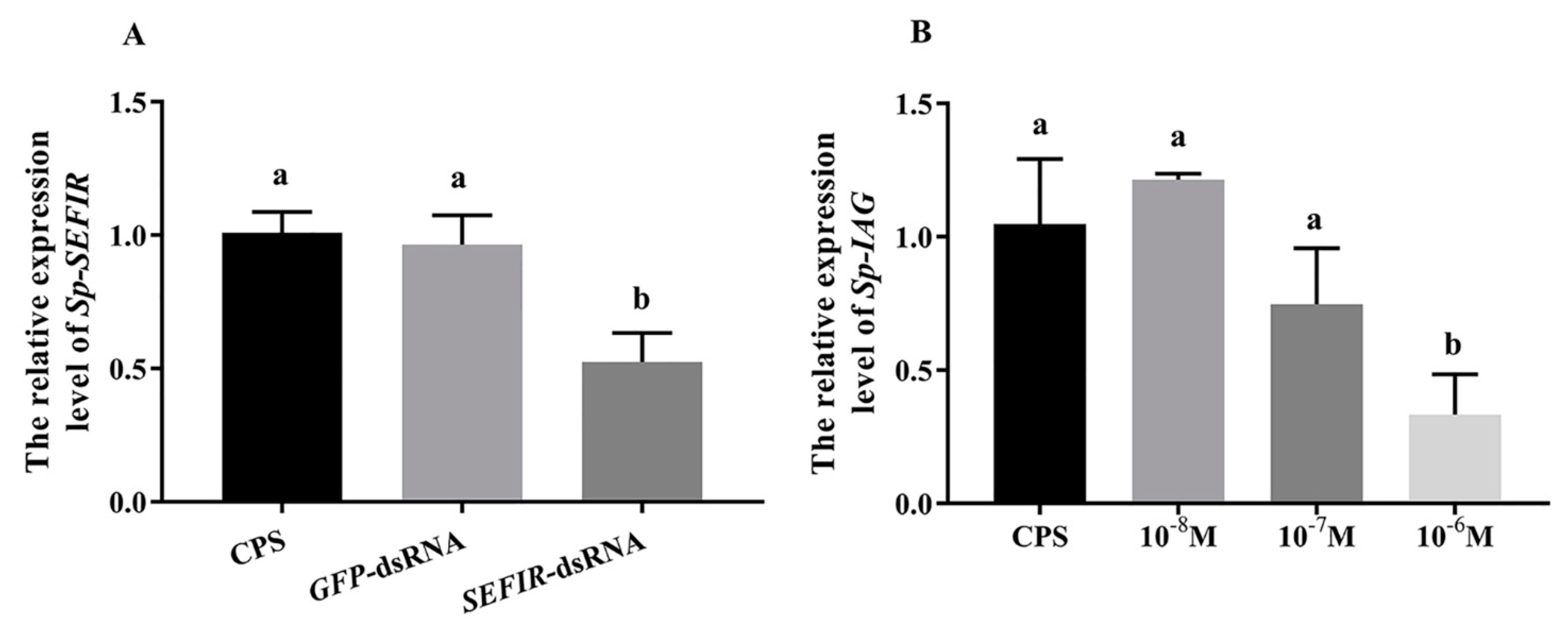
2.6. Analysis of Gene Expression in AG following Sp-SEFIR Silencing In Vivo
2.7. Analysis of Gene Expression in AG after Medication with rCFSH following Sp-SEFIR Silencing In Vitro
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. cDNA Cloning of Sp-SEFIR
4.3. Quantitative Real-Time PCR (qRT-PCR) Assays
4.4. Tissue Distribution of Sp-SEFIR
4.5. Sp-SEFIR Expression Profile throughout AG Development
4.6. Immunofluorescence Assays
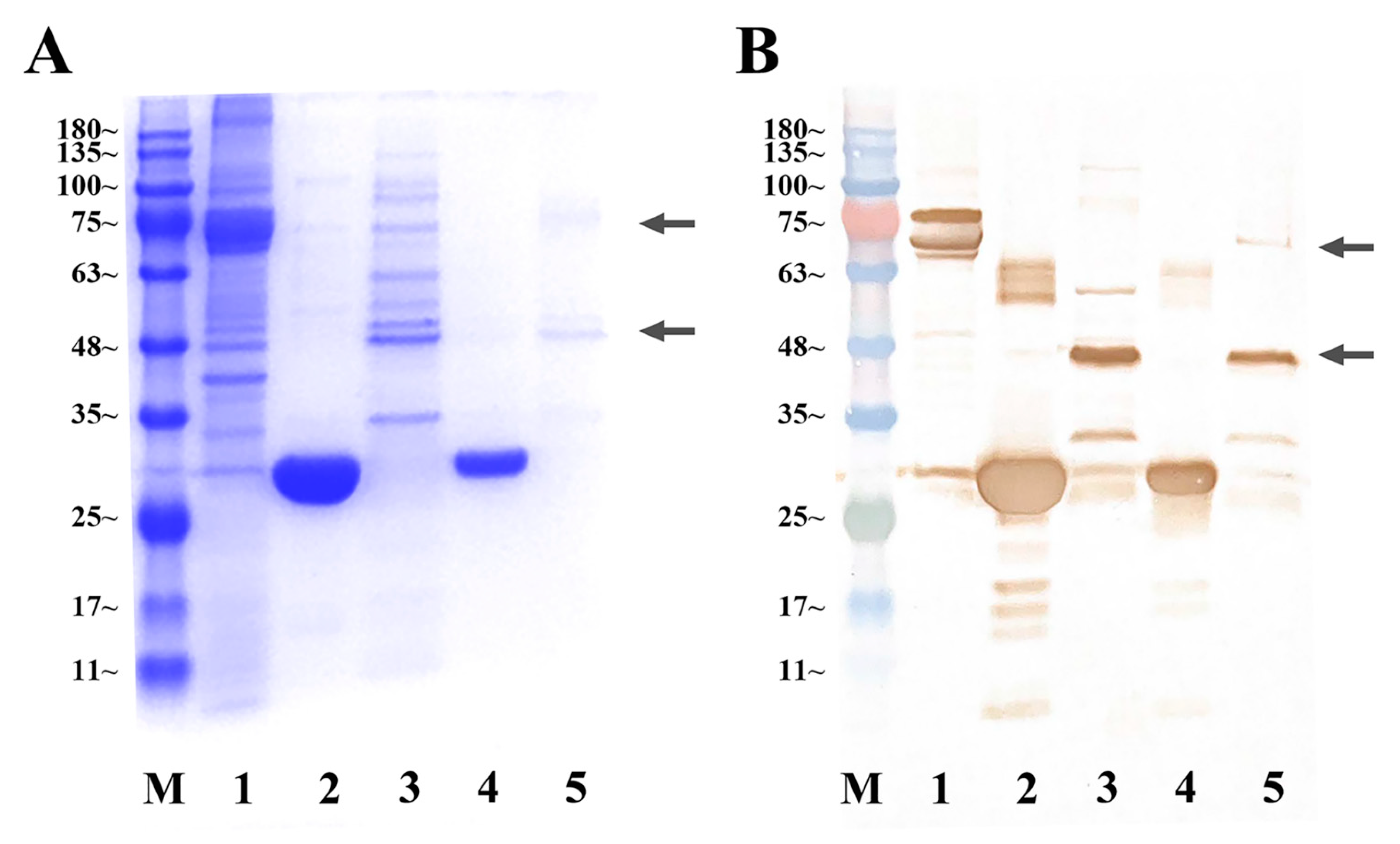
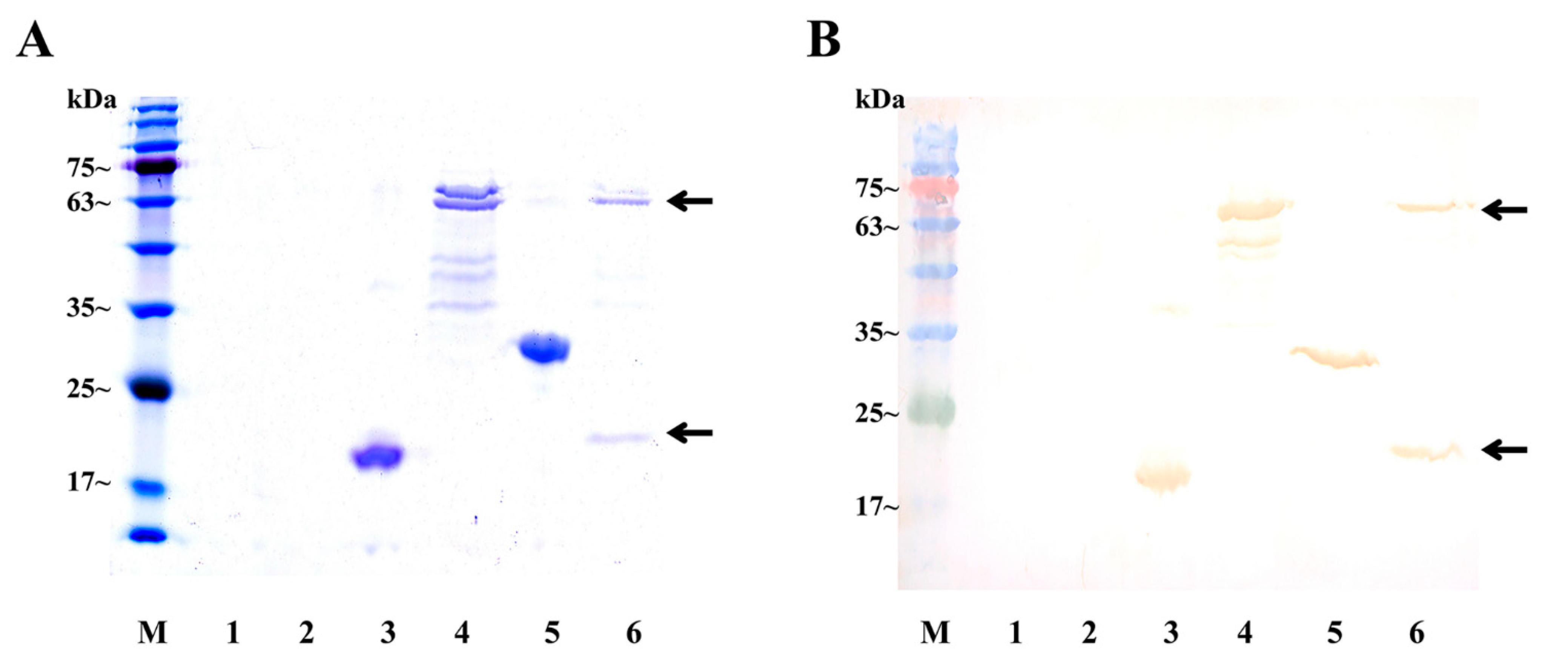
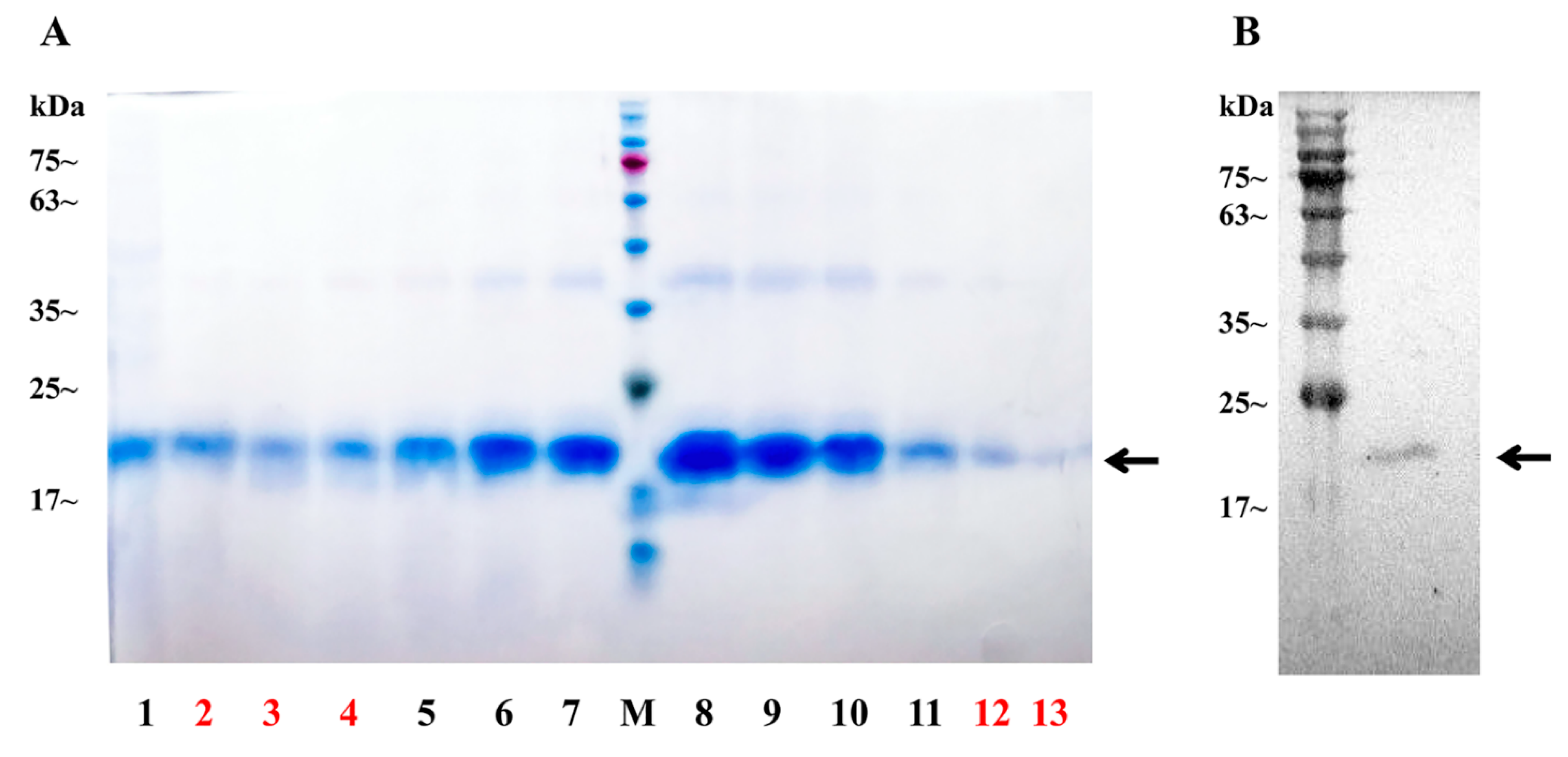
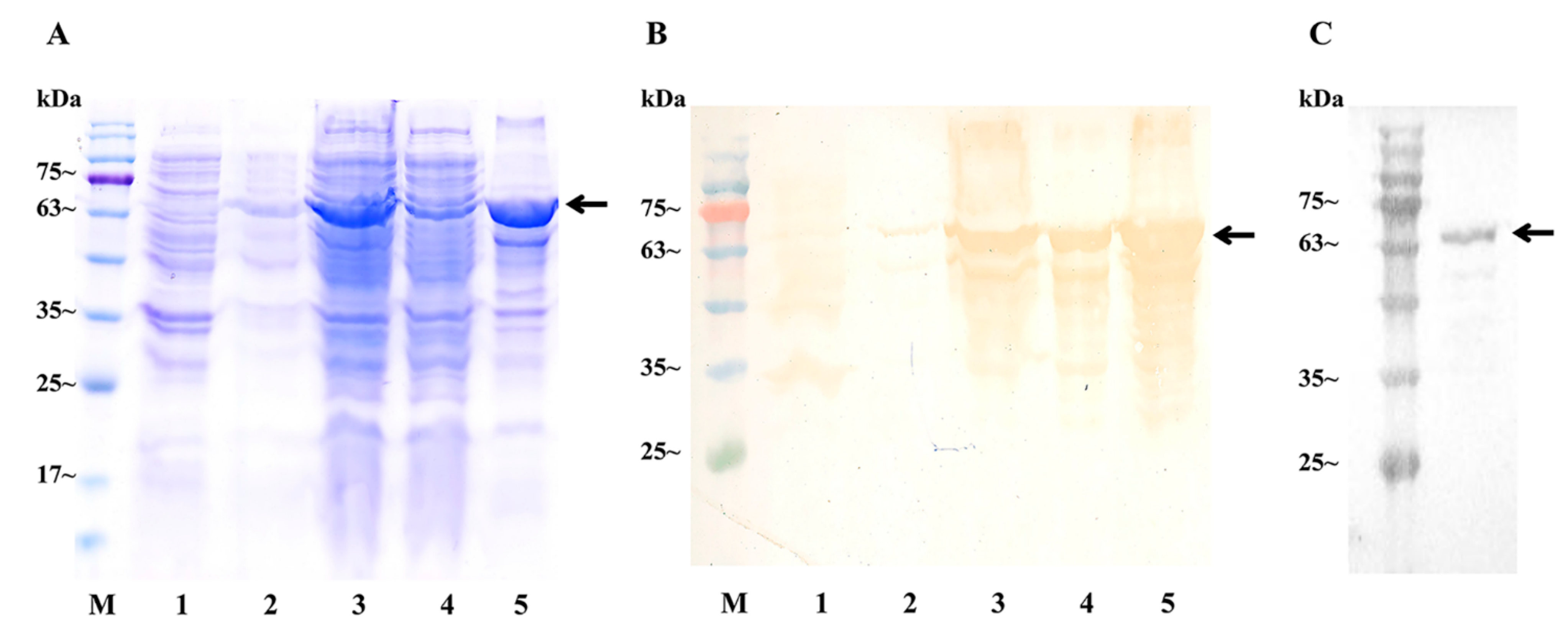
4.7. Expression of Sp-CFSH Recombinant Proteins and Sp-SEFIR Extracellular Domain Recombinant Protein
4.8. GST Pull-Down Assays
4.9. Silencing Experiment In Vivo
4.10. In Vitro Experiment: Sp-SEFIR Interference
4.11. Bioinformatics Analyses
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence (5′-3′) | Application |
|---|---|---|
| 5′R1 | GTCAAACCGCTGAATGCTGCTGATC | 5′RACE |
| 5′R2 | TCTTCATCCATCTGGTTGGGTTCTCC | |
| 3′F1 | GACCACCACCCTCCTGGTGTACCAC | 3′RACE |
| 3′F2 | TGAGGTGGTCTTGGAGAACCCAACC | |
| Sp-SEFIR-OF | GGTTGTAAACTGCAATAATACACCCTTAA | ORF validation |
| Sp-SEFIR-OR | CACATACTTTTAAATAAATGTTGGCGGA | |
| Sp-SEFIR-F | TGTGGAGAGGAAGGAGGTA | RT-RCR |
| Sp-SEFIR-R | CTGTATGCTACGAGTGGAACTA | |
| Sp-SEFIR-dsF | AAGTGAAAAATGTGCCCTGG | dsRNA synthesis |
| Sp-SEFIR-dsR | TGTCAGTCTGAACGGTGAGC | |
| GFP-F | TGGGCGTGGATAGCGGTTTG | |
| GFP-R | GGTCGGGGTAGCGGCTGAAG | |
| Sp-SEFIR-qF | CCCTGTCCCAGAGGATGAGA | qRT-PCR |
| Sp-SEFIR-qR | TATAGCAACCCTTGGTGCCG | |
| Sp-STAT-qF | CACCAGATCAAGGAGTGTGAGCGACA | |
| Sp-STAT-qR | GGTGACAAGTGAGGACAGCAAGCGA | |
| Sp-IAG-qF | ATCCTTTTCCTCCGTTTGCC | |
| Sp-IAG-qR | TCGGGTCTTCGTCTTGTTCC | |
| Sp-actinF | GAGCGAGAAATCGTTCGTGAC | Internal control |
| Sp-actinR | GGAAGGAAGGCTGGAAGAGAG | |
| Sp-SEFIR-GF | CCGGAATTCTCAGAGACAGAGGAAAGTGAAAAATGT | rSEFIR expression |
| Sp-SEFIR-GR | CTAGCTAGCCATCAGACTGGTCGGAATGTAGAAT | |
| Sp-CFSH-HF | CGCGGATCCTCCTCCATCATAGGACACATGAATTC | rHisCFSH expression |
| Sp-CFSH-HR | GGGCTAGCTTTATTCTCGCTTAAGTCGATGTAG | |
| Sp-CFSH-GF | CGCGGATCCTCCTCCATCATAGGACA | rGSTCFSH expression |
| Sp-CFSH-GR | CTAGCTAGCTTTATTCTCGCTTAAGTCGATGTAG |
| Sequence | Species | GenBank Accession Number |
|---|---|---|
| IL-17RA | Homo sapiens | AAH11624.1 |
| IL-17RA | Mus musculus | NP_032385.1 |
| IL-17RA | Danio rerio | NP_001093473.1 |
| IL-17RA | Petromyzon marinus | XP_032825579.1 |
| IL-17RA | Anas platyrhynchos | XP_027319183.2 |
| IL-17RA | Corvus cornix cornix | XP_039426789.1 |
| IL-17RA | Ctenopharyngodon idella | AXF94778.1 |
| IL-17RB | Oreochromis niloticus | XP_005478394.1 |
| IL-17RB | Ctenopharyngodon idella | AXF94779.1 |
| IL-17RB | Rousettus aegyptiacus | XP_036074512.1 |
| IL-17RB | Mus musculus | NP_062529.2 |
| IL-17RB | Cynoglossus semilaevis | XP_016891716.1 |
| IL-17RB | Oryx dammah | XP_040103149.1 |
| IL-17RC | Homo sapiens | AAM77569.1 |
| IL-17RC | Mus musculus | AAM77570.1 |
| IL-17RC | Ctenopharyngodon idella | AXF94780.1 |
| IL-17RC | Epinephelus lanceolatus | XP_033483132.1 |
| IL-17RC | Nibea albiflora | KAG8007775.1 |
| IL-17RC | Monopterus albus | XP_020466423.1 |
| IL-17RC | Marmota monax | KAH8211556.1 |
| IL-17RD | Mytilus edulis | CAG2215332.1 |
| IL-17RD | Sepia pharaonis | CAE1176245.1 |
| IL-17RD | Ctenopharyngodon idella | AXF94781.1 |
| IL-17RD | Oreochromis niloticus | XP_005453648.3 |
| IL-17RD | Crassostrea gigas | XP_034308350.1 |
| IL-17RD | Branchiostoma lanceolatum | CAH1264928.1 |
| IL-17RD | Homo sapiens | AAI11703.2 |
| IL-17RD | Mus musculus | AAI38630.1 |
| IL-17RE | Monopterus albus | XP_020466421.1 |
| IL-17RE | Mus musculus | NP_665825.2 |
| IL-17RE | Ochotona princeps | XP_004581715.1 |
| IL-17RE | Epinephelus coioides | ASU91966.1 |
| IL-17RE | Syngnathus acus | XP_037134538.1 |
| Sp-SEFIR | Scylla paramamosain | ON787957 |
Appendix B




References
- Toyota, K.; Miyakawa, H.; Hiruta, C.; Sato, T.; Katayama, H.; Ohira, T.; Iguchi, T. Sex determination and differentiation in decapod and cladoceran crustaceans: An overview of endocrine regulation. Genes 2021, 12, 305. [Google Scholar] [CrossRef]
- Lee, T.H.; Yamauchi, M.; Yamazaki, F. Sex-differentiation in the crab Eriocheir japonicus (Decapoda, Grapsidae). Invertebr. Reprod. Dev. 1994, 25, 123–137. [Google Scholar] [CrossRef]
- Li, S.H.; Li, F.H.; Yu, K.J.; Xiang, J.H. Identification and characterization of a doublesex gene which regulates the expression of insulin-like androgenic gland hormone in Fenneropenaeus chinensis. Gene 2018, 649, 1–7. [Google Scholar] [CrossRef]
- Yu, Y.Q.; Ma, W.M.; Zeng, Q.G.; Qian, Y.Q.; Yang, J.S.; Yang, W.J. Molecular cloning and sexually dimorphic expression of two Dmrt genes in the giant freshwater prawn, Macrobrachium rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Zheng, J.B.; Cheng, S.; Jia, Y.Y.; Gu, Z.M.; Li, F.; Chi, M.L.; Liu, S.L.; Jiang, W.P. Molecular identification and expression profiles of four splice variants of Sex-lethal gene in Cherax quadricarinatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 234, 26–33. [Google Scholar] [CrossRef]
- Manor, R.; Weil, S.; Oren, S.; Glazer, L.; Aflalo, E.D.; Ventura, T.; Chalifa-Caspi, V.; Lapidot, M.; Sagi, A. Insulin and gender: An insulin-like gene expressed exclusively in the androgenic gland of the male crayfish. Gen. Comp. Endocrinol. 2007, 150, 326–336. [Google Scholar] [CrossRef]
- Zmora, N.; Chung, J.S. A novel hormone is required for the development of reproductive phenotypes in adult female crabs. Endocrinology 2014, 155, 230–239. [Google Scholar] [CrossRef]
- Rosen, O.; Weil, S.; Manor, R.; Roth, Z.; Khalaila, I.; Sagi, A. A crayfish insulin-like-binding protein: Another piece in the androgenic gland insulin-like hormone puzzle is revealed. J. Biol. Chem. 2013, 288, 22289–22298. [Google Scholar] [CrossRef]
- Aizen, J.; Chandler, J.C.; Fitzgibbon, Q.P.; Sagi, A.; Battaglene, S.C.; Elizur, A.; Ventura, T. Production of recombinant insulin-like androgenic gland hormones from three decapod species: In vitro testicular phosphorylation and activation of a newly identified tyrosine kinase receptor from the Eastern spiny lobster, Sagmariasus verreauxi. Gen. Comp. Endocrinol. 2016, 229, 8–18. [Google Scholar] [CrossRef]
- Sharabi, O.; Manor, R.; Weil, S.; Aflalo, E.D.; Lezer, Y.; Levy, T.; Aizen, J.; Ventura, T.; Mather, P.B.; Khalaila, I.; et al. Identification and characterization of an insulin-like receptor involved in crustacean reproduction. Endocrinology 2016, 157, 928–941. [Google Scholar] [CrossRef]
- Guo, Q.; Li, S.H.; Lv, X.J.; Xiang, J.H.; Sagi, A.; Manor, R.; Li, F.H. A putative insulin-like androgenic gland hormone receptor gene specifically expressed in male Chinese shrimp. Endocrinology 2018, 159, 2173–2185. [Google Scholar] [CrossRef]
- Tan, K.; Li, Y.; Zhou, M.; Wang, W. siRNA knockdown of MrIR induces sex reversal in Macrobrachium rosenbergii. Aquaculture 2020, 523, 735172. [Google Scholar] [CrossRef]
- Yang, G.; Lu, Z.; Qin, Z.; Zhao, L.; Pan, G.; Shen, H.; Zhang, M.; Liang, R.; Lin, L.; Zhang, K. Insight into the regulatory relationships between the insulin-like androgenic gland hormone gene and the insulin-like androgenic gland hormone-binding protein gene in giant freshwater prawns (Macrobrachium rosenbergii). Int. J. Mol. Sci. 2020, 21, 4207. [Google Scholar] [CrossRef]
- Jiang, Q.L.; Lu, B.; Lin, D.D.; Huang, H.Y.; Chen, X.L.; Ye, H.H. Role of crustacean female sex hormone (CFSH) in sex differentiation in early juvenile mud crabs, Scylla paramamosain. Gen. Comp. Endocrinol. 2020, 289, 113383. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.Y.; Huang, L.; Wang, G.Z.; Zhu, Z.H.; Ye, H.H. Roles of crustacean female sex hormone 1a in a protandric simultaneous hermaphrodite shrimp. Front. Mar. Sci. 2021, 8, 791965. [Google Scholar] [CrossRef]
- Zhu, D.; Feng, T.; Mo, N.; Han, R.; Lu, W.; Shao, S.; Cui, Z. New insights for the regulatory feedback loop between type 1 crustacean female sex hormone (CFSH-1) and insulin-like androgenic gland hormone (IAG) in the Chinese mitten crab (Eriocheir sinensis). Front. Physiol. 2022, 13, 1054773. [Google Scholar] [CrossRef]
- Ventura, T.; Cummins, S.F.; Fitzgibbon, Q.; Battaglene, S.; Elizur, A. Analysis of the central nervous system transcriptome of the eastern rock lobster Sagmariasus verreauxi reveals its putative neuropeptidome. PLoS ONE 2014, 9, e97323. [Google Scholar] [CrossRef]
- Veenstra, J.A. The power of next-generation sequencing as illustrated by the neuropeptidome of the crayfish Procambarus clarkii. Gen. Comp. Endocrinol. 2015, 224, 84–95. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Cummins, S.F.; Elizur, A.; Ventura, T. Transcriptomic characterization and curation of candidate neuropeptides regulating reproduction in the eyestalk ganglia of the Australian crayfish, Cherax quadricarinatus. Sci. Rep. 2016, 6, 38658. [Google Scholar] [CrossRef]
- Toullec, J.Y.; Corre, E.; Mandon, P.; Gonzalez-Aravena, M.; Ollivaux, C.; Lee, C.Y. Characterization of the neuropeptidome of a Southern Ocean decapod, the Antarctic shrimp Chorismus antarcticus: Focusing on a new decapod ITP-like peptide belonging to the CHH peptide family. Gen. Comp. Endocrinol. 2017, 252, 60–78. [Google Scholar] [CrossRef]
- Kotaka, S.; Ohira, T. cDNA cloning and in situ localization of a crustacean female sex hormone-like molecule in the kuruma prawn Marsupenaeus japonicus. Fish. Sci. 2018, 84, 53–60. [Google Scholar] [CrossRef]
- Tsutsui, N.; Kotaka, S.; Ohira, T.; Sakamoto, T. Characterization of distinct ovarian isoform of crustacean female sex hormone in the kuruma prawn Marsupenaeus japonicus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 217, 7–16. [Google Scholar] [CrossRef]
- Oliphant, A.; Alexander, J.L.; Swain, M.T.; Webster, S.G.; Wilcockson, D.C. Transcriptomic analysis of crustacean neuropeptide signaling during the moult cycle in the green shore crab, Carcinus maenas. BMC Genomics 2018, 19, 711–726. [Google Scholar] [CrossRef]
- Thongbuakaew, T.; Suwansa-ard, S.; Sretarugsa, P.; Sobhon, P.; Cummins, S.F. Identification and characterization of a crustacean female sex hormone in the giant freshwater prawn, Macrobrachium rosenbergii. Aquaculture 2019, 507, 56–68. [Google Scholar] [CrossRef]
- Liu, X.; Ma, K.Y.; Liu, Z.Q.; Feng, J.B.; Ye, B.Q.; Qiu, G.F. Transcriptome analysis of the brain of the Chinese mitten crab, Eriocheir sinensis, for neuropeptide abundance profiles during ovarian development. Anim. Reprod. Sci. 2019, 201, 63–70. [Google Scholar] [CrossRef]
- Wang, Z.K.; Luan, S.; Meng, X.H.; Cao, B.X.; Luo, K.; Kong, J. Comparative transcriptomic characterization of the eyestalk in Pacific white shrimp (Litopenaeus vannamei) during ovarian maturation. Gen. Comp. Endocrinol. 2019, 274, 60–72. [Google Scholar] [CrossRef]
- Liu, A.; Liu, J.; Liu, F.; Huang, Y.Y.; Wang, G.Z.; Ye, H.H. Crustacean female sex hormone from the mud crab Scylla paramamosain is highly expressed in prepubertal males and inhibits the development of androgenic gland. Front. Physiol. 2018, 9, 924. [Google Scholar] [CrossRef]
- Jiang, Q.L.; Lu, B.; Wang, G.Z.; Ye, H.H. Transcriptional inhibition of Sp-IAG by crustacean female sex hormone in the mud crab, Scylla paramamosain. Int. J. Mol. Sci. 2020, 21, 5300. [Google Scholar] [CrossRef]
- Iyer, S.; Acharya, K.R. Tying the knot: The cystine signature and molecular-recognition processes of the vascular endothelial growth factor family of angiogenic cytokines. FEBS J. 2011, 278, 4304–4322. [Google Scholar] [CrossRef]
- Jiang, X.L.; Dias, J.A.; He, X.L. Structural biology of glycoprotein hormones and their receptors: Insights to signaling. Mol. Cell Endocrinol. 2014, 382, 424–451. [Google Scholar] [CrossRef]
- Rast, J.P.; Smith, L.C.; Loza-Coll, M.; Hibino, T.; Litman, G.W. Genomic insights into the immune system of the sea urchin. Science 2006, 314, 952–956. [Google Scholar] [CrossRef]
- Hibino, T.; Loza-Coll, M.; Messier, C.; Majeske, A.J.; Cohen, A.H.; Terwilliger, D.P.; Buckley, K.M.; Brockton, V.; Nair, S.V.; Berney, K.; et al. The immune gene repertoire encoded in the purple sea urchin genome. Dev. Biol. 2006, 300, 349–365. [Google Scholar] [CrossRef]
- Roberts, S.; Gueguen, Y.; de Lorgeril, J.; Goetz, F. Rapid accumulation of an interleukin 17 homolog transcript in Crassostrea gigas hemocytes following bacterial exposure. Dev. Comp. Immunol. 2008, 32, 1099–1104. [Google Scholar] [CrossRef]
- Wu, S.Z.; Huang, X.D.; Li, Q.; He, M.X. Interleukin-17 in pearl oyster (Pinctada fucata): Molecular cloning and functional characterization. Fish. Shellfish. Immunol. 2013, 34, 1050–1056. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zhang, Y.H.; Xiang, Z.M.; Tong, Y.; Qu, F.F.; Yu, Z.N. Genomic characterization and expression analysis of five novel IL-17 genes in the Pacific oyster, Crassostrea gigas. Fish Shellfish Immunol. 2014, 40, 455–465. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, M.; Xia, N.; Yu, S.; Chen, Y.; Wang, N. Cloning and analysis of gene expression of interleukin-17 homolog in triangle-shell pearl mussel, Hyriopsis cumingii, during pearl sac formation. Fish. Shellfish Immunol. 2016, 52, 151–156. [Google Scholar] [CrossRef]
- Vizzini, A.; Di Falco, F.; Parrinello, D.; Sanfratello, M.A.; Mazzarella, C.; Parrinello, N.; Cammarata, M. Ciona intestinalis interleukin 17-like genes expression is upregulated by LPS challenge. Dev. Comp. Immunol. 2015, 48, 129–137. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Gallardo-Escárate, C. Molecular cloning and expression of IRAK-4, IL-17 and I-κB genes in Haliotis rufescens challenged with Vibrio anguillarum. Fish. Shellfish Immunol. 2014, 36, 503–509. [Google Scholar] [CrossRef]
- Rosani, U.; Varotto, L.; Gerdol, M.; Pallavicini, A.; Venier, P. IL-17 signaling components in bivalves: Comparative sequence analysis and involvement in the immune responses. Dev. Comp. Immunol. 2015, 52, 255–268. [Google Scholar] [CrossRef]
- Subramaniam, S.V.; Cooper, R.S.; Adunyah, S.E. Evidence for the involvement of JAK/STAT pathway in the signaling mechanism of interleukin-17. Biochem. Biophys. Res. Commun. 1999, 262, 14–19. [Google Scholar] [CrossRef]
- Ye, H.H.; Li, S.J.; Huang, H.Y.; Wang, G.Z.; Lin, Q.W. Histological study on development of androgenic gland in mud crab Scylla serrata. J. Fish. Sci. China 2003, 10, 376–380. [Google Scholar]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, M.; Liu, X. Phase-specific expression of an insulin-like androgenic gland factor in a marine shrimp Lysmata wurdemanni: Implication for maintaining protandric simultaneous hermaphroditism. PLoS ONE 2017, 12, e0172782. [Google Scholar] [CrossRef]
- Levy, T.; Sagi, A. The “IAG-Switch”-a key controlling element in decapod crustacean sex differentiation. Front. Endocrinol. 2020, 11, 651. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.Y.; Ye, H.H.; Zeng, C.S.; Zhu, Z.H. Insulin-like androgenic gland hormone 1 (IAG1) regulates sexual differentiation in a hermaphrodite shrimp through feedback to neuroendocrine factors. Gen. Comp. Endocrinol. 2021, 303, 113706. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.Y.; Ye, H.H.; Liu, A.; Zhu, Z.H. RNAi reveals role of insulin-like androgenic gland hormone 2 (IAG2) in sexual differentiation and growth in hermaphrodite shrimp. Front. Mar. Sci. 2021, 8, 666763. [Google Scholar] [CrossRef]
- Levy, T.; Tamone, S.L.; Manor, R.; Aflalo, E.D.; Sklarz, M.Y.; Chalifa-Caspi, V.; Sagi, A. The IAG-Switch and further transcriptomic insights into sexual differentiation of a protandric shrimp. Front. Mar. Sci. 2020, 7, 587454. [Google Scholar] [CrossRef]
- Huang, X.D.; Zhang, H.; He, M.X. Comparative and evolutionary analysis of the interleukin 17 gene family in invertebrates. PLoS ONE 2015, 10, e0132802. [Google Scholar] [CrossRef]
- Terajima, D.; Yamada, S.; Uchino, R.; Ikawa, S.; Ikeda, M.; Shida, K.; Arai, Y.; Wang, H.-G.; Satoh, N.; Satake, M. Identification and sequence of seventy-nine new transcripts expressed in hemocytes of Ciona intestinalis, three of which may be involved in characteristic cell-cell communication. DNA Res. 2003, 10, 203–212. [Google Scholar] [CrossRef][Green Version]
- Gaffen, S.L. Structure and signalling in the IL-17 receptor family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef]
- Gu, C.F.; Wu, L.; Li, X.X. IL-17 family: Cytokines, receptors and signaling. Cytokine 2013, 64, 477–485. [Google Scholar] [CrossRef]
- Amatya, N.; Garg, A.V.; Gaffen, S.L. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef]
- Tsutsui, S.; Nakamura, O.; Watanabe, T. Lamprey (Lethenteron japonicum) IL-17 upregulated by LPS-stimulation in the skin cells. Immunogenetics 2007, 59, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Du, L.Y.; Qin, L.; Wang, X.Y.; Zhang, A.Y.; Wei, H.; Zhou, H. Characterization of grass carp (Ctenopharyngodon idella) IL-17D: Molecular cloning, functional implication and signal transduction. Dev. Comp. Immunol. 2014, 42, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.S.; Zhang, H.; Zhang, R.; Li, H.; Wang, W.L.; Wang, L.L.; Wang, H.; Qiu, L.M.; Song, L.S. CgIL17-5, an ancient inflammatory cytokine in Crassostrea gigas exhibiting the heterogeneity functions compared with vertebrate interleukin17 molecules. Dev. Comp. Immunol. 2015, 53, 339–348. [Google Scholar] [CrossRef]
- Liu, H.; Cheung, K.C.; Chu, K.H. Cell Structure and Seasonal Changes of the Androgenic Gland of the Mud Crab Scylla paramamosain (Decapoda: Portunidae). Zool. Stud. 2008, 47, 720–732. [Google Scholar]
- Gong, J.; Ye, H.H.; Xie, Y.J.; Yang, Y.A.; Huang, H.Y.; Li, S.J.; Zeng, C.S. Ecdysone receptor in the mud crab Scylla paramamosain: A possible role in promoting ovarian development. J. Endocrinol. 2015, 224, 273–287. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Huang, L.; Liu, A.; Jiang, Q.; Huang, H.; Ye, H. Identification of a Putative CFSH Receptor Inhibiting IAG Expression in Crabs. Int. J. Mol. Sci. 2023, 24, 12240. https://doi.org/10.3390/ijms241512240
Liu F, Huang L, Liu A, Jiang Q, Huang H, Ye H. Identification of a Putative CFSH Receptor Inhibiting IAG Expression in Crabs. International Journal of Molecular Sciences. 2023; 24(15):12240. https://doi.org/10.3390/ijms241512240
Chicago/Turabian StyleLiu, Fang, Lin Huang, An Liu, Qingling Jiang, Huiyang Huang, and Haihui Ye. 2023. "Identification of a Putative CFSH Receptor Inhibiting IAG Expression in Crabs" International Journal of Molecular Sciences 24, no. 15: 12240. https://doi.org/10.3390/ijms241512240
APA StyleLiu, F., Huang, L., Liu, A., Jiang, Q., Huang, H., & Ye, H. (2023). Identification of a Putative CFSH Receptor Inhibiting IAG Expression in Crabs. International Journal of Molecular Sciences, 24(15), 12240. https://doi.org/10.3390/ijms241512240