


Melanocortin 1 Receptor (MC1R): Pharmacological and Therapeutic Aspects
Abstract
1. Introduction
2. Melanocortin 1 Receptor (MC1R)
2.1. Structure
2.2. Signaling Pathway
3. MC1R-Related Diseases
3.1. Melanoma
3.2. Systemic Sclerosis
3.3. Neuroinflammation
3.4. Atherosclerosis
4. Modulators
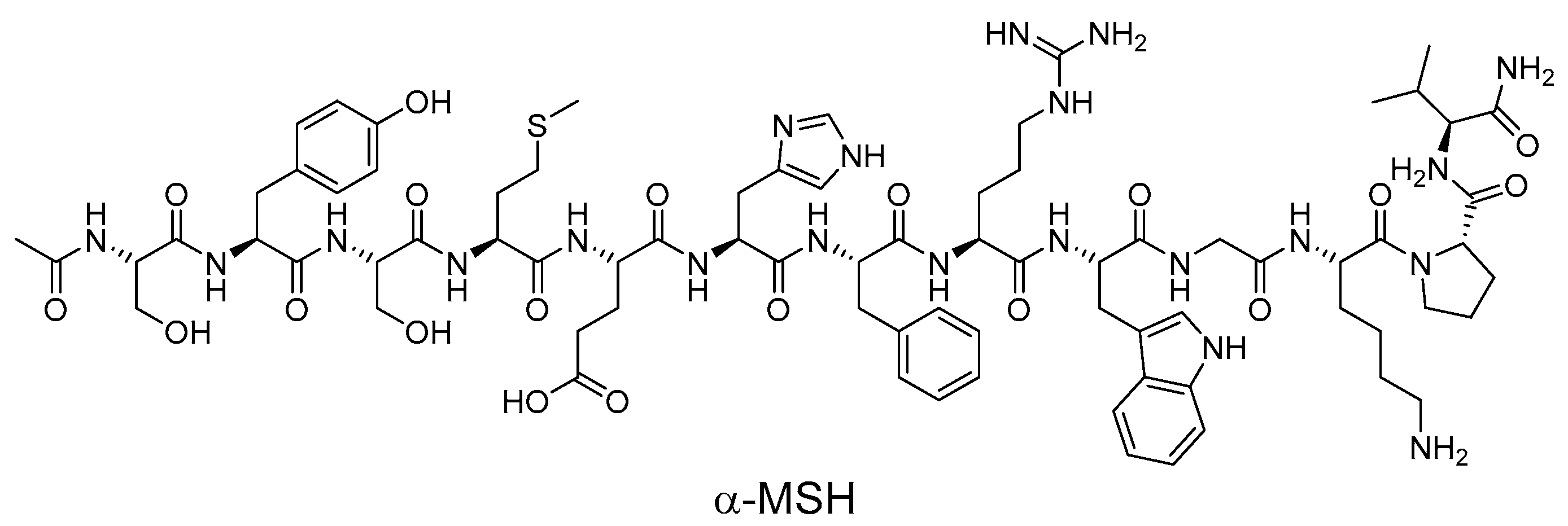
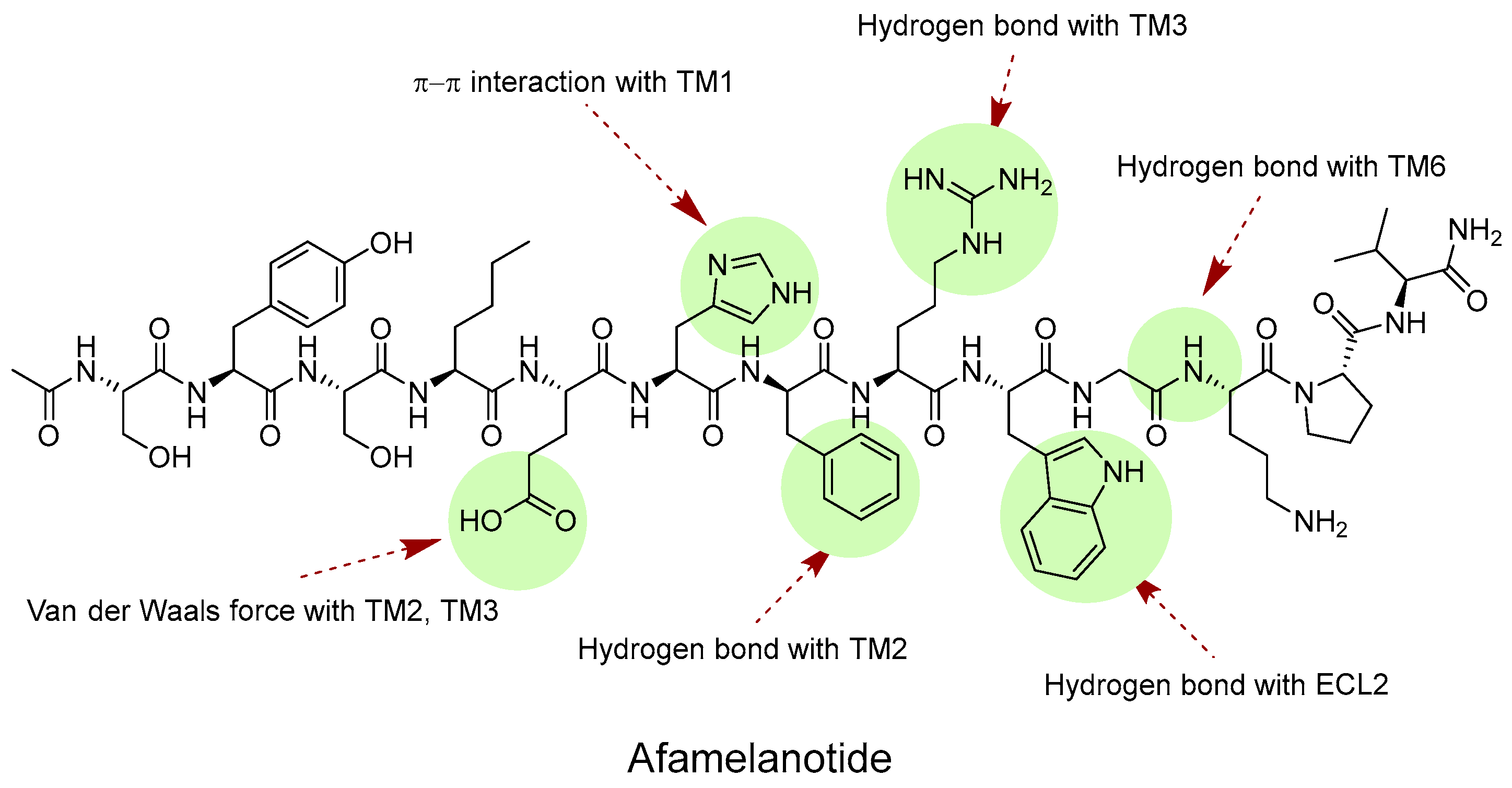
4.1. Endogenous Ligands
4.2. Synthetic Peptide Modulators
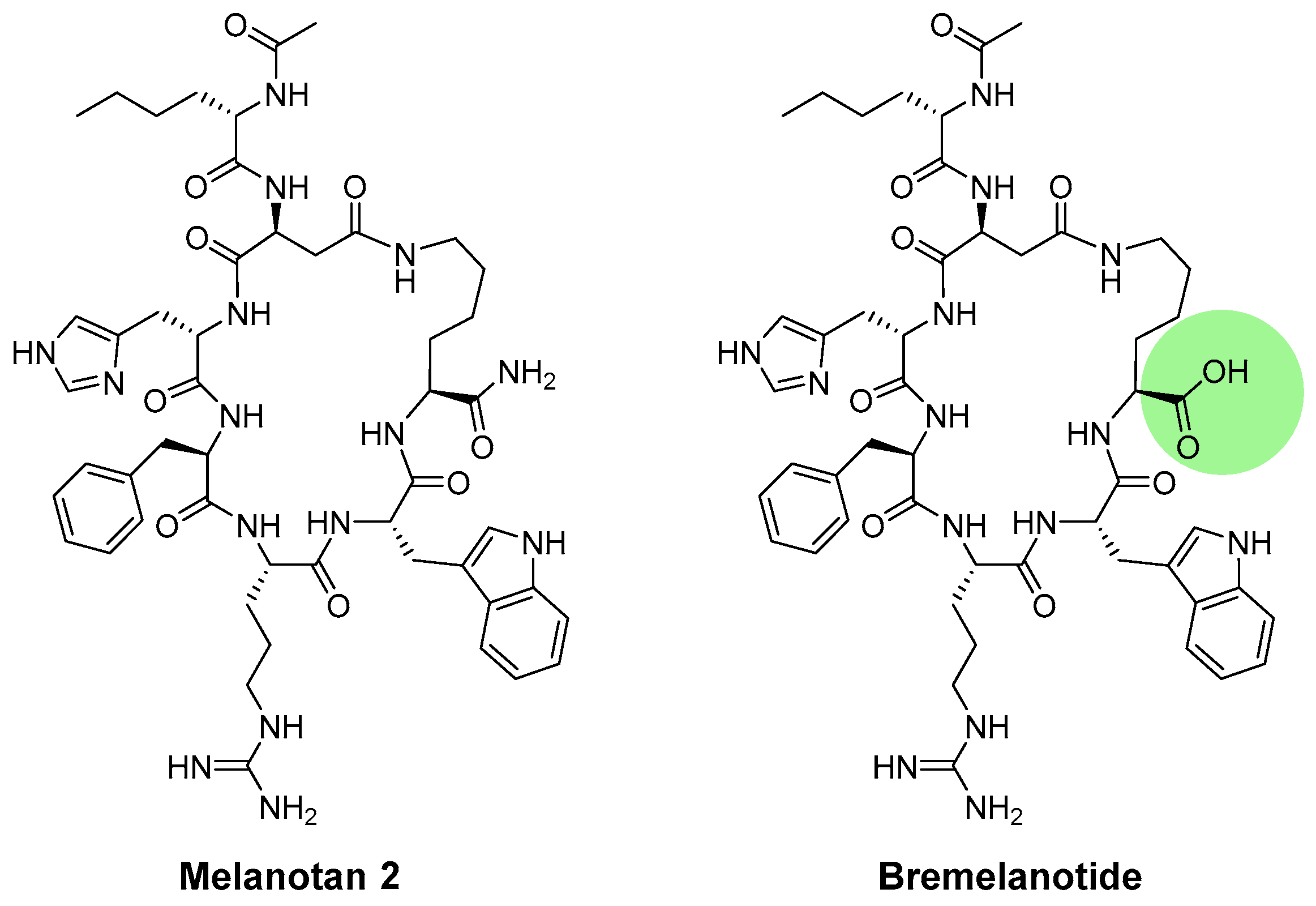
4.2.1. Melanotan I (MT-I)
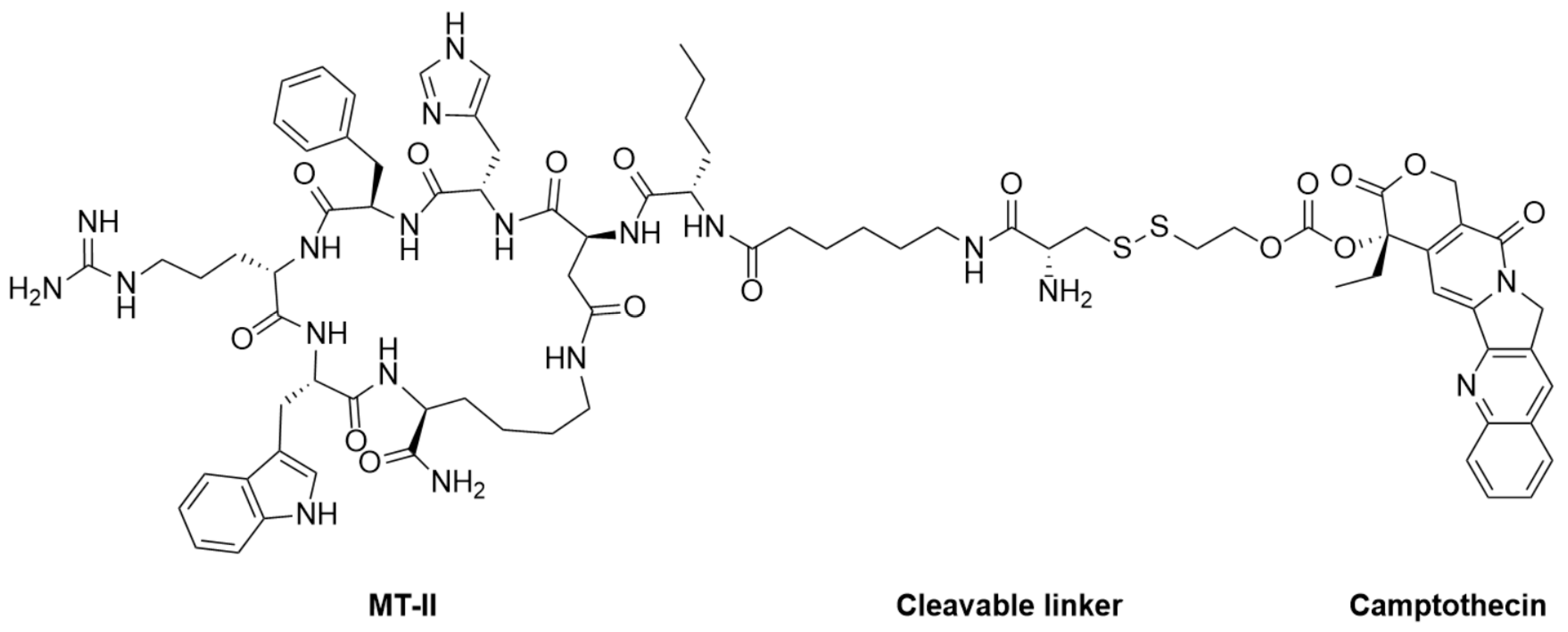
4.2.2. Melanotan II (MT-II)
4.2.3. Peptide Agonist for Vitiligo Treatment
4.3. Small Molecule Modulators
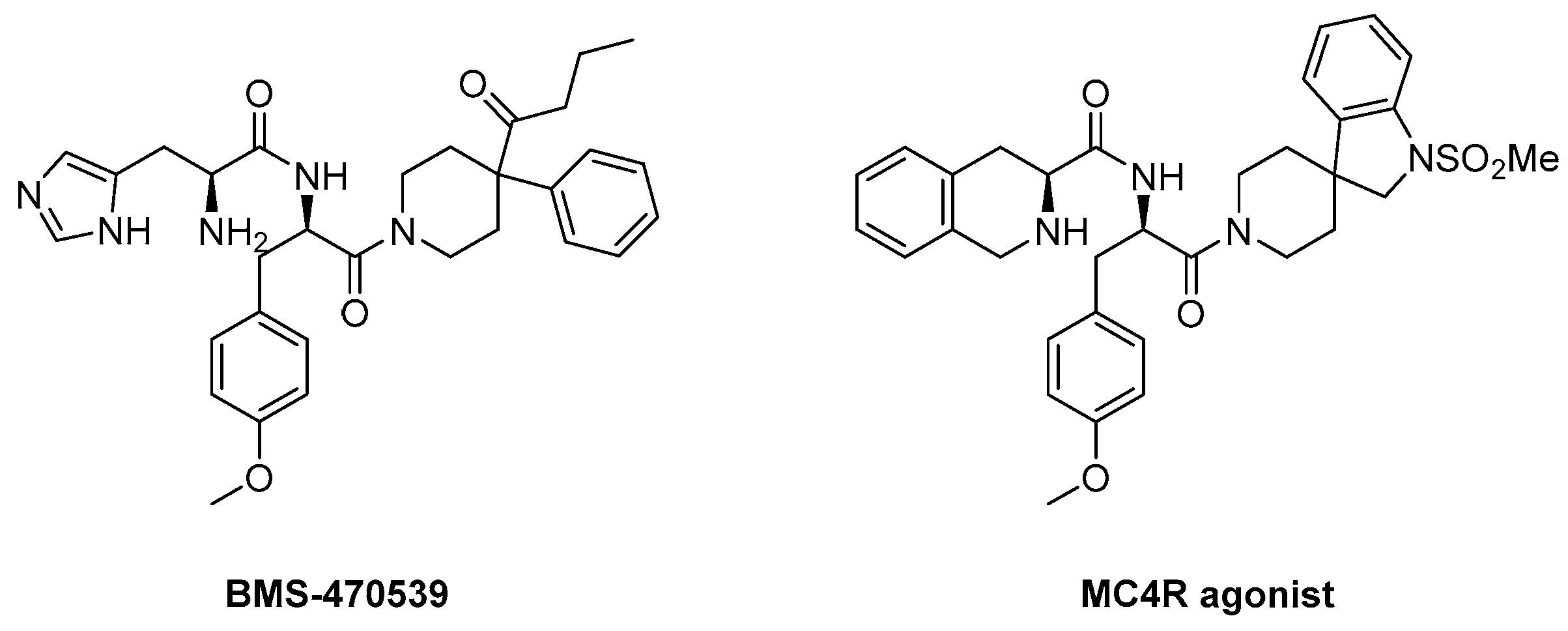
4.3.1. BMS-470539
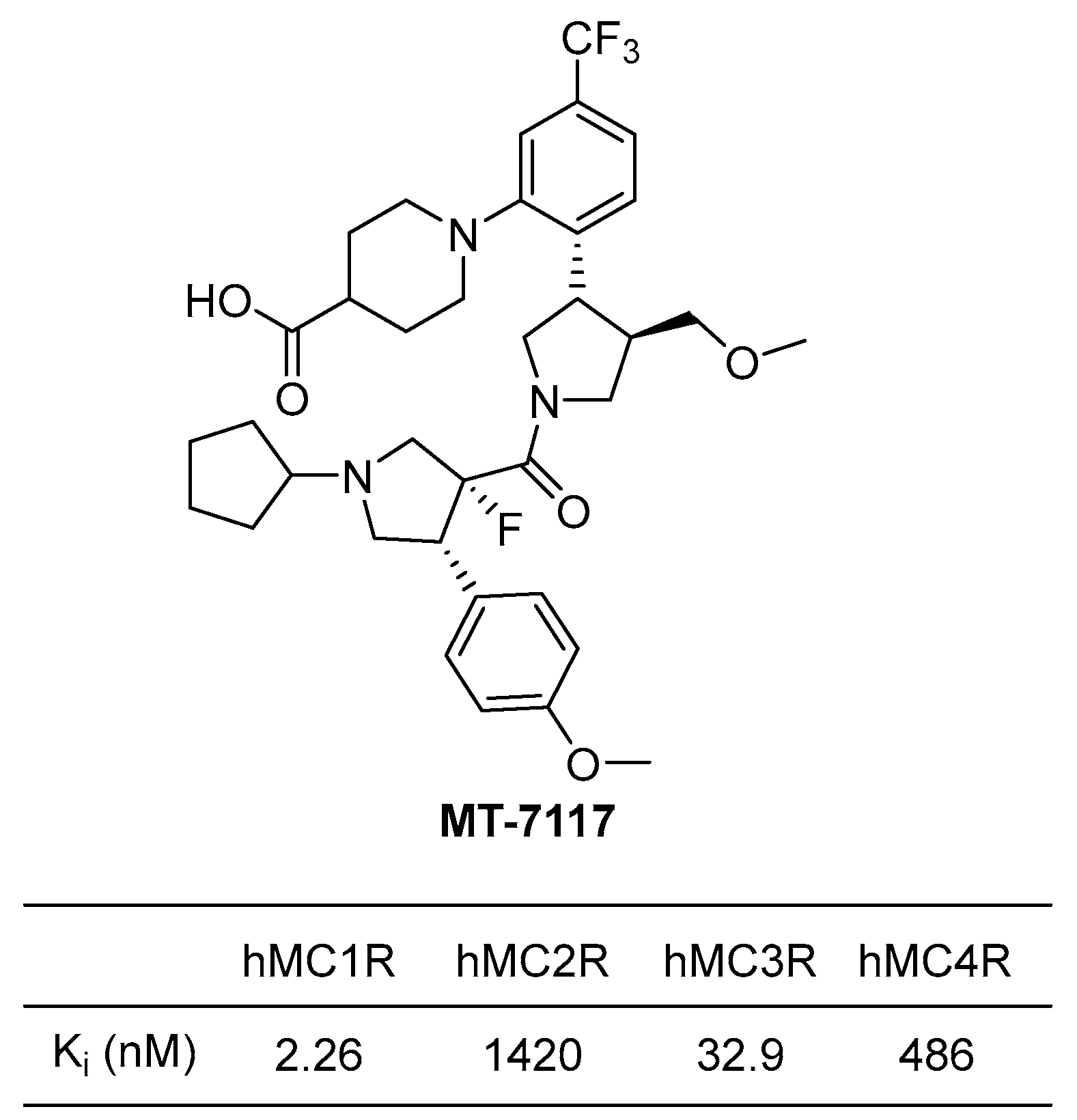
4.3.2. MT-7117 (Dersimelagon)
4.3.3. AP1189 (Resomelagon)
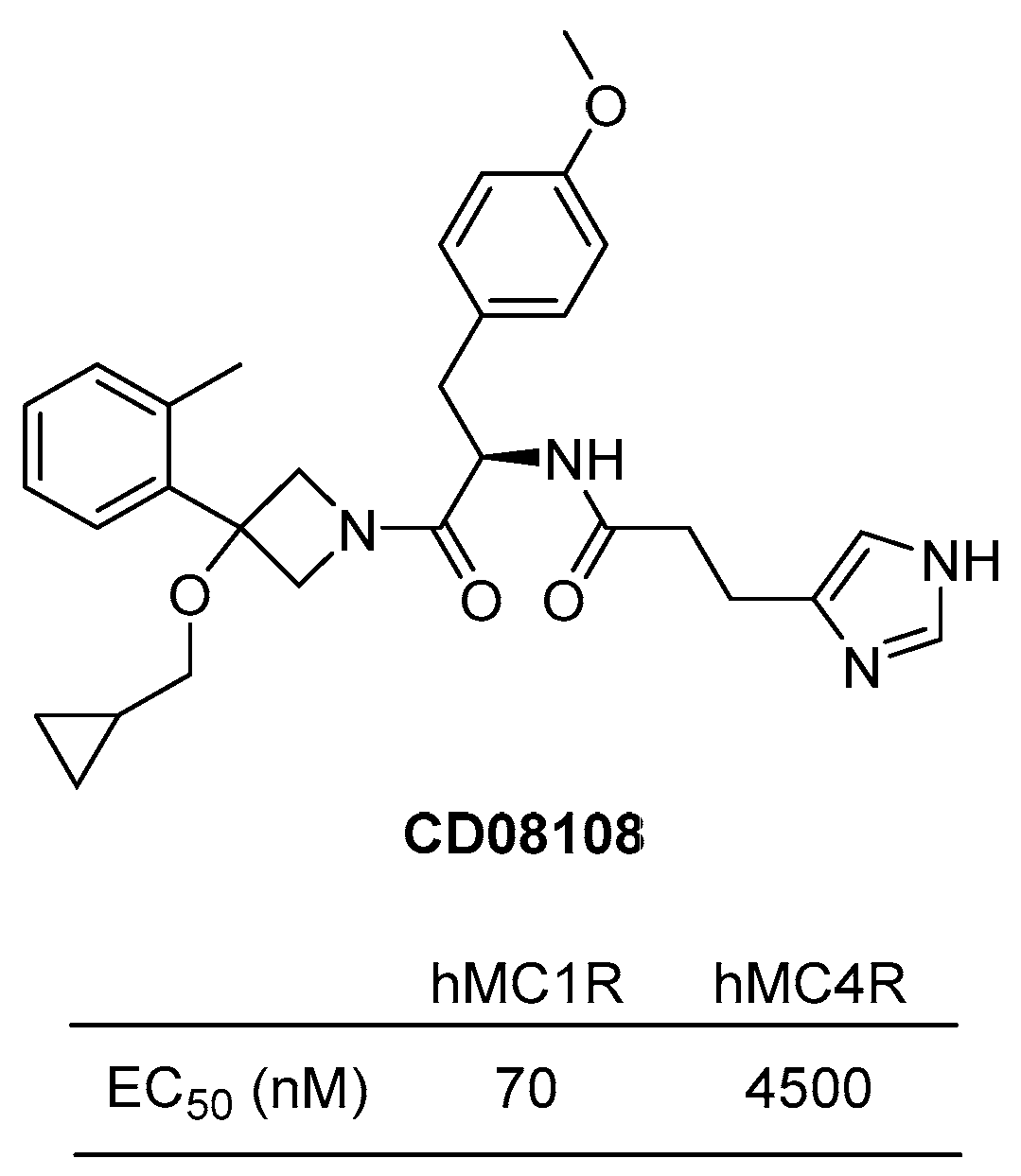
4.3.4. CD08108
4.3.5. JNJ-10229570
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cawley, N.X.; Li, Z.; Loh, Y.P. Biosynthesis, trafficking, and secretion of pro-opiomelanocortin-derived peptides. J. Mol. Endocrinol. 2016, 56, T77–T97. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.D. Studies on the Physiological Functions of the Melanocortin System. Endocr. Rev. 2006, 27, 736–749. [Google Scholar] [CrossRef]
- Gallo-Payet, N. Adrenal and extra-adrenal functions of ACTH. J. Mol. Endocrinol. 2016, 56, T135–T156. [Google Scholar] [CrossRef] [PubMed]
- Robbins, L.S.; Nadeau, J.H.; Johnson, K.R.; Kelly, M.A.; Roselli-Rehfuss, L.; Baack, E.; Mountjoy, K.G.; Cone, R.D. Pigmentation phenotypes of variant extension locus alleles result from point mutations that alter MSH receptor function. Cell 1993, 72, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Hiltz, M.E.; Lipton, J.M. Alpha-MSH peptides inhibit acute inflammation and contact sensitivity. Peptides 1990, 11, 979–982. [Google Scholar] [CrossRef]
- Cutuli, M.; Cristiani, S.; Lipton, J.M.; Catania, A. Antimicrobial effects of α-MSH peptides. J. Leukoc. Biol. 2000, 67, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.F.; Queiroz-Junior, C.M.; Montero-Melendez, T.; Werneck, S.M.; Correa, J.D.; Soriani, F.M.; Garlet, G.P.; Souza, D.G.; Teixeira, M.M.; Silva, T.A.; et al. Melanocortin agonism as a viable strategy to control alveolar bone loss induced by oral infection. FASEB J. 2016, 30, 4033–4041. [Google Scholar] [CrossRef]
- Gomez-SanMiguel, A.B.; Martin, A.I.; Nieto-Bona, M.P.; Fernandez-Galaz, C.; Villanua, M.A.; Lopez-Calderon, A. The melanocortin receptor type 3 agonist d-Trp(8)-γMSH decreases inflammation and muscle wasting in arthritic rats. J. Cachexia Sarcopenia Muscle 2016, 7, 79–89. [Google Scholar] [CrossRef]
- Gomez-SanMiguel, A.B.; Villanua, M.A.; Martin, A.I.; Lopez-Calderon, A. D-TRP(8)-γMSH Prevents the Effects of Endotoxin in Rat Skeletal Muscle Cells through TNFα/NF-KB Signalling Pathway. PLoS ONE 2016, 11, e0155645. [Google Scholar] [CrossRef]
- Kadiri, J.J.; Thapa, K.; Kaipio, K.; Cai, M.; Hruby, V.J.; Rinne, P. Melanocortin 3 receptor activation with [D-Trp8]-γ-MSH suppresses inflammation in apolipoprotein E deficient mice. Eur. J. Pharmacol. 2020, 880, 173186. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Melanocortin 1 Receptor: Structure, Function, and Regulation. Front. Genet. 2016, 7, 95. [Google Scholar] [CrossRef] [PubMed]
- Gantz, I.; Shimoto, Y.; Konda, Y.; Miwa, H.; Dickinson, C.J.; Yamada, T. Molecular cloning, expression, and characterization of a fifth melanocortin receptor. Biochem. Biophys. Res. Commun. 1994, 200, 1214–1220. [Google Scholar] [CrossRef]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef] [PubMed]
- Desarnaud, F.; Labbe, O.; Eggerickx, D.; Vassart, G.; Parmentier, M. Molecular cloning, functional expression and pharmacological characterization of a mouse melanocortin receptor gene. Biochem. J. 1994, 299 Pt 2, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Gantz, I.; Yamada, T.; Tashiro, T.; Konda, Y.; Shimoto, Y.; Miwa, H.; Trent, J.M. Mapping of the gene encoding the melanocortin-1 (α-melanocyte stimulating hormone) receptor (MC1R) to human chromosome 16q24.3 by Fluorescence in situ hybridization. Genomics 1994, 19, 394–395. [Google Scholar] [CrossRef]
- Schaible, E.V.; Steinstrasser, A.; Jahn-Eimermacher, A.; Luh, C.; Sebastiani, A.; Kornes, F.; Pieter, D.; Schafer, M.K.; Engelhard, K.; Thal, S.C. Single administration of tripeptide α-MSH(11–13) attenuates brain damage by reduced inflammation and apoptosis after experimental traumatic brain injury in mice. PLoS ONE 2013, 8, e71056. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Becher, E.; Mahnke, K.; Hartmeyer, M.; Schwarz, T.; Scholzen, T.; Luger, T.A. Evidence for the differential expression of the functional alpha-melanocyte-stimulating hormone receptor MC-1 on human monocytes. J. Immunol. 1997, 158, 3378–3384. [Google Scholar] [CrossRef]
- Siegrist, W.; Stutz, S.; Eberle, A.N. Homologous and heterologous regulation of alpha-melanocyte-stimulating hormone receptors in human and mouse melanoma cell lines. Cancer Res. 1994, 54, 2604–2610. [Google Scholar]
- Siegrist, W.; Solca, F.; Stutz, S.; Giuffre, L.; Carrel, S.; Girard, J.; Eberle, A.N. Characterization of receptors for alpha-melanocyte-stimulating hormone on human melanoma cells. Cancer Res. 1989, 49, 6352–6358. [Google Scholar]
- Ghanem, G.E.; Comunale, G.; Libert, A.; Vercammen-Grandjean, A.; Lejeune, F.J. Evidence for alpha-melanocyte-stimulating hormone (α-MSH) receptors on human malignant melanoma cells. Int. J. Cancer 1988, 41, 248–255. [Google Scholar] [CrossRef]
- Roberts, D.W.; Newton, R.A.; Beaumont, K.A.; Helen Leonard, J.; Sturm, R.A. Quantitative analysis of MC1R gene expression in human skin cell cultures. Pigment Cell Res. 2006, 19, 76–89. [Google Scholar] [CrossRef]
- Donatien, P.D.; Hunt, G.; Pieron, C.; Lunec, J.; Taieb, A.; Thody, A.J. The expression of functional MSH receptors on cultured human melanocytes. Arch. Dermatol. Res. 1992, 284, 424–426. [Google Scholar] [CrossRef]
- Sanchez-Laorden, B.L.; Sanchez-Mas, J.; Turpin, M.C.; Garcia-Borron, J.C.; Jimenez-Cervantes, C. Variant amino acids in different domains of the human melanocortin 1 receptor impair cell surface expression. Cell. Mol. Biol. 2006, 52, 39–46. [Google Scholar]
- Frandberg, P.A.; Doufexis, M.; Kapas, S.; Chhajlani, V. Cysteine residues are involved in structure and function of melanocortin 1 receptor: Substitution of a cysteine residue in transmembrane segment two converts an agonist to antagonist. Biochem. Biophys. Res. Commun. 2001, 281, 851–857. [Google Scholar] [CrossRef]
- Wallin, E.; von Heijne, G. Properties of N-terminal tails in G-protein coupled receptors: A statistical study. Protein Eng. 1995, 8, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Chhajlani, V.; Xu, X.; Blauw, J.; Sudarshi, S. Identification of ligand binding residues in extracellular loops of the melanocortin 1 receptor. Biochem. Biophys. Res. Commun. 1996, 219, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Borron, J.C.; Sanchez-Laorden, B.L.; Jimenez-Cervantes, C. Melanocortin-1 receptor structure and functional regulation. Pigment Cell Res. 2005, 18, 393–410. [Google Scholar] [CrossRef]
- Pitcher, J.A.; Freedman, N.J.; Lefkowitz, R.J. G protein-coupled receptor kinases. Annu. Rev. Biochem. 1998, 67, 653–692. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L.M.; Lefkowitz, R.J. The role of β-arrestins in the termination and transduction of G-protein-coupled receptor signals. J. Cell Sci. 2002, 115, 455–465. [Google Scholar] [CrossRef]
- Strader, C.D.; Fong, T.M.; Tota, M.R.; Underwood, D.; Dixon, R.A. Structure and function of G protein-coupled receptors. Annu. Rev. Biochem. 1994, 63, 101–132. [Google Scholar] [CrossRef]
- Qanbar, R.; Bouvier, M. Role of palmitoylation/depalmitoylation reactions in G-protein-coupled receptor function. Pharmacol. Ther. 2003, 97, 1–33. [Google Scholar] [CrossRef]
- Schulein, R.; Hermosilla, R.; Oksche, A.; Dehe, M.; Wiesner, B.; Krause, G.; Rosenthal, W. A dileucine sequence and an upstream glutamate residue in the intracellular carboxyl terminus of the vasopressin V2 receptor are essential for cell surface transport in COS.M6 cells. Mol. Pharmacol. 1998, 54, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Schwartz, T.W. Molecular mechanism of agonism and inverse agonism in the melanocortin receptors: Zn2+ as a structural and functional probe. Ann. N. Y. Acad. Sci. 2003, 994, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chen, Y.; Dai, A.; Yin, W.; Guo, J.; Yang, D.; Zhou, F.; Jiang, Y.; Wang, M.W.; Xu, H.E. Structural mechanism of calcium-mediated hormone recognition and Gβ interaction by the human melanocortin-1 receptor. Cell Res. 2021, 31, 1061–1071. [Google Scholar] [CrossRef]
- Arisi, M.; Rossi, M.; Rovati, C.; Tomasi, C.; Mori, L.; Laini, L.; Calzavara-Pinton, P.G. Clinical and dermoscopic changes of acquired melanocytic nevi of patients treated with afamelanotide. Photochem. Photobiol. Sci. 2021, 20, 315–320. [Google Scholar] [CrossRef]
- Kadekaro, A.L.; Leachman, S.; Kavanagh, R.J.; Swope, V.; Cassidy, P.; Supp, D.; Sartor, M.; Schwemberger, S.; Babcock, G.; Wakamatsu, K.; et al. Melanocortin 1 receptor genotype: An important determinant of the damage response of melanocytes to ultraviolet radiation. FASEB J. 2010, 24, 3850–3860. [Google Scholar] [CrossRef]
- Song, X.; Mosby, N.; Yang, J.; Xu, A.; Abdel-Malek, Z.; Kadekaro, A.L. α-MSH activates immediate defense responses to UV-induced oxidative stress in human melanocytes. Pigment Cell Melanoma Res. 2009, 22, 809–818. [Google Scholar] [CrossRef]
- Kokot, A.; Metze, D.; Mouchet, N.; Galibert, M.D.; Schiller, M.; Luger, T.A.; Bohm, M. α-melanocyte-stimulating hormone counteracts the suppressive effect of UVB on Nrf2 and Nrf-dependent gene expression in human skin. Endocrinology 2009, 150, 3197–3206. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.A.; Swope, V.B.; Starner, R.J.; Koikov, L.; Cassidy, P.; Leachman, S. Melanocortins and the melanocortin 1 receptor, moving translationally towards melanoma prevention. Arch Biochem. Biophys. 2014, 563, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, I.; Tada, A.; Ollmann, M.M.; Barsh, G.S.; Im, S.; Lamoreux, M.L.; Hearing, V.J.; Nordlund, J.J.; Abdel-Malek, Z.A. Agouti signaling protein inhibits melanogenesis and the response of human melanocytes to α-melanotropin. J. Investig. Dermatol. 1997, 108, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat. Rev. Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Neves, S.R.; Ram, P.T.; Iyengar, R. G protein pathways. Science 2002, 296, 1636–1639. [Google Scholar] [CrossRef]
- Herraiz, C.; Journé, F.; Abdel-Malek, Z.; Ghanem, G.; Jiménez-Cervantes, C.; García-Borrón, J.C. Signaling from the human melanocortin 1 receptor to ERK1 and ERK2 mitogen-activated protein kinases involves transactivation of cKIT. Mol. Endocrinol. 2011, 25, 138–156. [Google Scholar] [CrossRef]
- Bohm, M.; Schulte, U.; Kalden, H.; Luger, T.A. Alpha-melanocyte-stimulating hormone modulates activation of NF-κB and AP-1 and secretion of interleukin-8 in human dermal fibroblasts. Ann. N. Y. Acad. Sci. 1999, 885, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, V.; Camillieri, G.; Avitabile, T.; Leggio, G.M.; Drago, F. Effects of the COOH-terminal tripeptide α-MSH(11–13) on corneal epithelial wound healing: Role of nitric oxide. Exp. Eye Res. 2006, 83, 1366–1372. [Google Scholar] [CrossRef]
- Becher, E.; Mahnke, K.; Brzoska, T.; Kalden, D.H.; Grabbe, S.; Luger, T.A. Human peripheral blood-derived dendritic cells express functional melanocortin receptor MC-1R. Ann. N. Y. Acad. Sci. 1999, 885, 188–195. [Google Scholar] [CrossRef]
- Scholzen, T.E.; Sunderkotter, C.; Kalden, D.H.; Brzoska, T.; Fastrich, M.; Fisbeck, T.; Armstrong, C.A.; Ansel, J.C.; Luger, T.A. α-melanocyte stimulating hormone prevents lipopolysaccharide-induced vasculitis by down-regulating endothelial cell adhesion molecule expression. Endocrinology 2003, 144, 360–370. [Google Scholar] [CrossRef]
- Kalden, D.H.; Scholzen, T.; Brzoska, T.; Luger, T.A. Mechanisms of the antiinflammatory effects of α-MSH. Role of transcription factor NF-κB and adhesion molecule expression. Ann. N. Y. Acad. Sci. 1999, 885, 254–261. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Liu, X.; Jiang, Y.; Zhang, L.; Zhang, X.; Li, X.; Zhang, Y. α-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS ONE 2014, 9, e93433. [Google Scholar] [CrossRef] [PubMed]
- Galimberti, D.; Baron, P.; Meda, L.; Prat, E.; Scarpini, E.; Delgado, R.; Catania, A.; Lipton, J.M.; Scarlato, G. α-MSH peptides inhibit production of nitric oxide and tumor necrosis factor-α by microglial cells activated with β-amyloid and interferon γ. Biochem. Biophys. Res. Commun. 1999, 263, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Carlin, A.; Airaghi, L.; Demitri, M.T.; Meda, L.; Galimberti, D.; Baron, P.; Lipton, J.M.; Catania, A. Melanocortin peptides inhibit production of proinflammatory cytokines and nitric oxide by activated microglia. J. Leukoc. Biol. 1998, 63, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Taherzadeh, S.; Sharma, S.; Chhajlani, V.; Gantz, I.; Rajora, N.; Demitri, M.T.; Kelly, L.; Zhao, H.; Ichiyama, T.; Catania, A.; et al. α-MSH and its receptors in regulation of tumor necrosis factor-α production by human monocyte/macrophages. Am. J. Physiol. 1999, 276, R1289–R1294. [Google Scholar] [CrossRef]
- Ottani, A.; Giuliani, D.; Neri, L.; Calevro, A.; Canalini, F.; Vandini, E.; Cainazzo, M.M.; Ruberto, I.A.; Barbieri, A.; Rossi, R.; et al. NDP-α-MSH attenuates heart and liver responses to myocardial reperfusion via the vagus nerve and JAK/ERK/STAT signaling. Eur. J. Pharmacol. 2015, 769, 22–32. [Google Scholar] [CrossRef]
- Buggy, J.J. Binding of α-melanocyte-stimulating hormone to its G-protein-coupled receptor on B-lymphocytes activates the Jak/STAT pathway. Biochem. J. 1998, 331 Pt 1, 211–216. [Google Scholar] [CrossRef]
- Mandrika, I.; Muceniece, R.; Wikberg, J.E. Effects of melanocortin peptides on lipopolysaccharide/interferon-gamma-induced NF-kappaB DNA binding and nitric oxide production in macrophage-like RAW 264.7 cells: Evidence for dual mechanisms of action. Biochem. Pharmacol. 2001, 61, 613–621. [Google Scholar] [CrossRef]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-κB action. Cell. Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef]
- Manna, S.K.; Aggarwal, B.B. Alpha-melanocyte-stimulating hormone inhibits the nuclear transcription factor NF-kappa B activation induced by various inflammatory agents. J. Immunol. 1998, 161, 2873–2880. [Google Scholar] [CrossRef]
- Wang, W.; Guo, D.Y.; Lin, Y.J.; Tao, Y.X. Melanocortin Regulation of Inflammation. Front. Endocrinol. 2019, 10, 683. [Google Scholar] [CrossRef]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: From UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef]
- Rosenkranz, A.A.; Slastnikova, T.A.; Durymanov, M.O.; Sobolev, A.S. Malignant melanoma and melanocortin 1 receptor. Biochemistry 2013, 78, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Onfray, F.; Lopez, M.; Lundqvist, A.; Aguirre, A.; Escobar, A.; Serrano, A.; Korenblit, C.; Petersson, M.; Chhajlani, V.; Larsson, O.; et al. Tissue distribution and differential expression of melanocortin 1 receptor, a malignant melanoma marker. Br. J. Cancer 2002, 87, 414–422. [Google Scholar] [CrossRef]
- Ogbah, Z.; Puig-Butille, J.A.; Simonetta, F.; Badenas, C.; Cervera, R.; Mila, J.; Benitez, D.; Malvehy, J.; Vilella, R.; Puig, S. Molecular characterization of human cutaneous melanoma-derived cell lines. Anticancer. Res. 2012, 32, 1245–1251. [Google Scholar] [PubMed]
- Eggermont, A.M. Advances in systemic treatment of melanoma. Ann. Oncol. 2010, 21 (Suppl. S7), vii339–vii344. [Google Scholar] [CrossRef]
- Iriyama, N.; Yoshino, Y.; Yuan, B.; Horikoshi, A.; Hirabayashi, Y.; Hatta, Y.; Toyoda, H.; Takeuchi, J. Speciation of arsenic trioxide metabolites in peripheral blood and bone marrow from an acute promyelocytic leukemia patient. J. Hematol. Oncol. 2012, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Montero-Melendez, T.; Boesen, T.; Jonassen, T.E.N. Translational advances of melanocortin drugs: Integrating biology, chemistry and genetics. Semin. Immunol. 2022, 59, 101603. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, B.; Yin, C.; Liu, W.; Han, C.; Chen, B.; Liu, T.; Li, X.; Chen, X.; Li, C.; et al. Palmitoylation-dependent activation of MC1R prevents melanomagenesis. Nature 2017, 549, 399–403. [Google Scholar] [CrossRef]
- Castejon-Grinan, M.; Herraiz, C.; Olivares, C.; Jimenez-Cervantes, C.; Garcia-Borron, J.C. cAMP-independent non-pigmentary actions of variant melanocortin 1 receptor: AKT-mediated activation of protective responses to oxidative DNA damage. Oncogene 2018, 37, 3631–3646. [Google Scholar] [CrossRef]
- Zhu, C.; Li, T.; Wang, Z.; Li, Z.; Wei, J.; Han, H.; Yuan, D.; Cai, M.; Shi, J. MC1R Peptide Agonist Self-Assembles into a Hydrogel That Promotes Skin Pigmentation for Treating Vitiligo. ACS Nano 2023, 17, 8723–8733. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Wei, J.; Varga, J. Understanding fibrosis in systemic sclerosis: Shifting paradigms, emerging opportunities. Nat. Rev. Rheumatol. 2011, 8, 42–54. [Google Scholar] [CrossRef]
- Khanna, D.; Lin, C.J.F.; Furst, D.E.; Goldin, J.; Kim, G.; Kuwana, M.; Allanore, Y.; Matucci-Cerinic, M.; Distler, O.; Shima, Y.; et al. Tocilizumab in systemic sclerosis: A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2020, 8, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for Systemic Sclerosis-Associated Interstitial Lung Disease. N. Engl. J. Med. 2019, 380, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Suzuki, T.; Kawano, Y.; Kojima, S.; Miyashiro, M.; Matsumoto, A.; Kania, G.; Blyszczuk, P.; Ross, R.L.; Mulipa, P.; et al. Dersimelagon, a novel oral melanocortin 1 receptor agonist, demonstrates disease-modifying effects in preclinical models of systemic sclerosis. Arthritis Res. Ther. 2022, 24, 210. [Google Scholar] [CrossRef]
- Pimentel, V.C.; Bellé, L.P.; Pinheiro, F.V.; De Bona, K.S.; Da Luz, S.C.; Moretto, M.B. Adenosine deaminase activity, lipid peroxidation and astrocyte responses in the cerebral cortex of rats after neonatal hypoxia ischemia. Int. J. Dev. Neurosci. 2009, 27, 857–862. [Google Scholar] [CrossRef]
- McRae, A.; Gilland, E.; Bona, E.; Hagberg, H. Microglia activation after neonatal hypoxic-ischemia. Brain Res. Dev. Brain Res. 1995, 84, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Fu, S.; Liu, Y.; Luo, H.; Li, F.; Wang, Y.; Gao, M.; Cheng, Y.; Xie, Z. NDP-MSH binding melanocortin-1 receptor ameliorates neuroinflammation and BBB disruption through CREB/Nr4a1/NF-κB pathway after intracerebral hemorrhage in mice. J. Neuroinflamm. 2019, 16, 192. [Google Scholar] [CrossRef]
- Kang, L.; McIntyre, K.W.; Gillooly, K.M.; Yang, Y.; Haycock, J.; Roberts, S.; Khanna, A.; Herpin, T.F.; Yu, G.; Wu, X.; et al. A selective small molecule agonist of the melanocortin-1 receptor inhibits lipopolysaccharide-induced cytokine accumulation and leukocyte infiltration in mice. J. Leukoc. Biol. 2006, 80, 897–904. [Google Scholar] [CrossRef]
- Yu, S.; Doycheva, D.M.; Gamdzyk, M.; Yang, Y.; Lenahan, C.; Li, G.; Li, D.; Lian, L.; Tang, J.; Lu, J.; et al. Activation of MC1R with BMS-470539 attenuates neuroinflammation via cAMP/PKA/Nurr1 pathway after neonatal hypoxic-ischemic brain injury in rats. J. Neuroinflamm. 2021, 18, 26. [Google Scholar] [CrossRef]
- Rinne, P.; Rami, M.; Nuutinen, S.; Santovito, D.; van der Vorst, E.P.C.; Guillamat-Prats, R.; Lyytikainen, L.P.; Raitoharju, E.; Oksala, N.; Ring, L.; et al. Melanocortin 1 Receptor Signaling Regulates Cholesterol Transport in Macrophages. Circulation 2017, 136, 83–97. [Google Scholar] [CrossRef]
- Catania, A.; Gatti, S.; Colombo, G.; Lipton, J.M. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol. Rev. 2004, 56, 1–29. [Google Scholar] [CrossRef]
- Rinne, P.; Silvola, J.M.; Hellberg, S.; Stahle, M.; Liljenback, H.; Salomaki, H.; Koskinen, E.; Nuutinen, S.; Saukko, P.; Knuuti, J.; et al. Pharmacological activation of the melanocortin system limits plaque inflammation and ameliorates vascular dysfunction in atherosclerotic mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1346–1354. [Google Scholar] [CrossRef]
- Rinne, P.; Kadiri, J.J.; Velasco-Delgado, M.; Nuutinen, S.; Viitala, M.; Hollmen, M.; Rami, M.; Savontaus, E.; Steffens, S. Melanocortin 1 Receptor Deficiency Promotes Atherosclerosis in Apolipoprotein E−/− Mice. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.B.; Mao, Y.F.; Meng, H.B.; Tian, Y.P.; Deng, X.M. STY39, a novel alpha-melanocyte-stimulating hormone analogue, attenuates bleomycin-induced pulmonary inflammation and fibrosis in mice. Shock 2011, 35, 308–314. [Google Scholar] [CrossRef]
- Spana, C.; Taylor, A.W.; Yee, D.G.; Makhlina, M.; Yang, W.; Dodd, J. Probing the Role of Melanocortin Type 1 Receptor Agonists in Diverse Immunological Diseases. Front. Pharmacol. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Cai, W.; Srivastava, P.; Feng, D.; Lin, Y.; Vanderburg, C.R.; Xu, Y.; McLean, P.; Frosch, M.P.; Fisher, D.E.; Schwarzschild, M.A.; et al. Melanocortin 1 receptor activation protects against alpha-synuclein pathologies in models of Parkinson’s disease. Mol. Neurodegener. 2022, 17, 16. [Google Scholar] [CrossRef]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: Regulation by ultraviolet B. Biochim. Biophys. Acta 1996, 1313, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Schauer, E.; Trautinger, F.; Kock, A.; Schwarz, A.; Bhardwaj, R.; Simon, M.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Proopiomelanocortin-derived peptides are synthesized and released by human keratinocytes. J. Clin. Investig. 1994, 93, 2258–2262. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.; Scott, M.C.; Suzuki, I.; Tada, A.; Im, S.; Lamoreux, L.; Ito, S.; Barsh, G.; Hearing, V.J. The melanocortin-1 receptor is a key regulator of human cutaneous pigmentation. Pigment Cell Res. 2000, 13 (Suppl. S8), 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev. 1998, 12, 316–330. [Google Scholar] [CrossRef]
- Blanchard, S.G.; Harris, C.O.; Ittoop, O.R.; Nichols, J.S.; Parks, D.J.; Truesdale, A.T.; Wilkison, W.O. Agouti antagonism of melanocortin binding and action in the B16F10 murine melanoma cell line. Biochemistry 1995, 34, 10406–10411. [Google Scholar] [CrossRef]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A β-defensin mutation causes black coat color in domestic dogs. Science 2007, 318, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Bartels, J.; Christophers, E.; Schroder, J.M. Isolation and characterization of human μ-defensin-3, a novel human inducible peptide antibiotic. J. Biol. Chem. 2001, 276, 5707–5713. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.D.; Ollmann, M.M.; Kang, L.; Stoffel, M.; Bell, G.I.; Barsh, G.S. Structure and function of ASP, the human homolog of the mouse agouti gene. Hum. Mol. Genet. 1995, 4, 223–230. [Google Scholar] [CrossRef]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Nix, M.A.; Kaelin, C.B.; Ta, T.; Weis, A.; Morton, G.J.; Barsh, G.S.; Millhauser, G.L. Molecular and functional analysis of human β-defensin 3 action at melanocortin receptors. Chem. Biol. 2013, 20, 784–795. [Google Scholar] [CrossRef]
- Swope, V.B.; Jameson, J.A.; McFarland, K.L.; Supp, D.M.; Miller, W.E.; McGraw, D.W.; Patel, M.A.; Nix, M.A.; Millhauser, G.L.; Babcock, G.F.; et al. Defining MC1R regulation in human melanocytes by its agonist α-melanocortin and antagonists agouti signaling protein and β-defensin 3. J. Investig. Dermatol. 2012, 132, 2255–2262. [Google Scholar] [CrossRef]
- Harris, J.I.; Lerner, A.B. Amino-acid sequence of the α-melanocyte-stimulating hormone. Nature 1957, 179, 1346–1347. [Google Scholar] [CrossRef]
- Hruby, V.J.; Wilkes, B.C.; Hadley, M.E.; Al-Obeidi, F.; Sawyer, T.K.; Staples, D.J.; de Vaux, A.E.; Dym, O.; Castrucci, A.M.; Hintz, M.F.; et al. alpha-Melanotropin: The minimal active sequence in the frog skin bioassay. J. Med. Chem. 1987, 30, 2126–2130. [Google Scholar] [CrossRef]
- Sawyer, T.K.; Sanfilippo, P.J.; Hruby, V.J.; Engel, M.H.; Heward, C.B.; Burnett, J.B.; Hadley, M.E. 4-Norleucine, 7-D-phenylalanine-alpha-melanocyte-stimulating hormone: A highly potent alpha-melanotropin with ultralong biological activity. Proc. Natl. Acad. Sci. USA 1980, 77, 5754–5758. [Google Scholar] [CrossRef]
- Hadley, M.E.; Marwan, M.M.; Al-Obeidi, F.; Hruby, V.J.; Castrucci, A.M. Linear and cyclic α-melanotropin [4-10]-fragment analogues that exhibit superpotency and residual activity. Pigment Cell Res. 1989, 2, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Al-Obeidi, F.; Castrucci, A.M.; Hadley, M.E.; Hruby, V.J. Potent and prolonged acting cyclic lactam analogues of alpha-melanotropin: Design based on molecular dynamics. J. Med. Chem. 1989, 32, 2555–2561. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.; Todd, C.; Cresswell, J.E.; Thody, A.J. α-melanocyte stimulating hormone and its analogue Nle4DPhe7 alpha-MSH affect morphology, tyrosinase activity and melanogenesis in cultured human melanocytes. J. Cell Sci. 1994, 107 Pt 1, 205–211. [Google Scholar] [CrossRef]
- Hunt, G.; Kyne, S.; Ito, S.; Wakamatsu, K.; Todd, C.; Thody, A. Eumelanin and phaeomelanin contents of human epidermis and cultured melanocytes. Pigment Cell Res. 1995, 8, 202–208. [Google Scholar] [CrossRef]
- Dorr, R.T.; Dvorakova, K.; Brooks, C.; Lines, R.; Levine, N.; Schram, K.; Miketova, P.; Hruby, V.; Alberts, D.S. Increased eumelanin expression and tanning is induced by a superpotent melanotropin [Nle4-D-Phe7]-α-MSH in humans. Photochem. Photobiol. 2000, 72, 526–532. [Google Scholar] [CrossRef]
- Langendonk, J.G.; Balwani, M.; Anderson, K.E.; Bonkovsky, H.L.; Anstey, A.V.; Bissell, D.M.; Bloomer, J.; Edwards, C.; Neumann, N.J.; Parker, C.; et al. Afamelanotide for Erythropoietic Protoporphyria. N. Engl. J. Med. 2015, 373, 48–59. [Google Scholar] [CrossRef]
- Schiöth, H.B.; Chhajlani, V.; Muceniece, R.; Klusa, V.; Wikberg, J.E.S. Major pharmacological distinction of the ACTH receptor from other melanocortin receptors. Life Sci. 1996, 59, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Haskell-Luevano, C.; Nikiforovich, G.; Sharma, S.D.; Yang, Y.-K.; Dickinson, C.; Hruby, V.J.; Gantz, I. Biological and Conformational Examination of Stereochemical Modifications Using the Template Melanotropin Peptide, Ac-Nle-c[Asp-His-Phe-Arg-Trp- Ala-Lys]-NH2, on Human Melanocortin Receptors. J. Med. Chem. 1997, 40, 1738–1748. [Google Scholar] [CrossRef]
- Wessells, H.; Hruby, V.J.; Hackett, J.; Han, G.; Balse-Srinivasan, P.; Vanderah, T.W. Ac-Nle-c[Asp-His-DPhe-Arg-Trp-Lys]-NH2 induces penile erection via brain and spinal melanocortin receptors. Neuroscience 2003, 118, 755–762. [Google Scholar] [CrossRef]
- Wessells, H.; Levine, N.; Hadley, M.E.; Dorr, R.; Hruby, V. Melanocortin receptor agonists, penile erection, and sexual motivation: Human studies with Melanotan II. Int. J. Impot. Res. 2000, 12 (Suppl. S4), S74–S79. [Google Scholar] [CrossRef]
- Rossler, A.S.; Pfaus, J.G.; Kia, H.K.; Bernabe, J.; Alexandre, L.; Giuliano, F. The melanocortin agonist, melanotan II, enhances proceptive sexual behaviors in the female rat. Pharmacol. Biochem. Behav. 2006, 85, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Diamond, L.E.; Earle, D.C.; Garcia, W.D.; Spana, C. Co-administration of low doses of intranasal PT-141, a melanocortin receptor agonist, and sildenafil to men with erectile dysfunction results in an enhanced erectile response. Urology 2005, 65, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Molinoff, P.B.; Shadiack, A.M.; Earle, D.; Diamond, L.E.; Quon, C.Y. PT-141: A Melanocortin Agonist for the Treatment of Sexual Dysfunction. Ann. N. Y. Acad. Sci. 2003, 994, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Hugo, W.; Zaretsky, J.M.; Sun, L.; Song, C.; Moreno, B.H.; Hu-Lieskovan, S.; Berent-Maoz, B.; Pang, J.; Chmielowski, B.; Cherry, G.; et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell 2016, 165, 35–44. [Google Scholar] [CrossRef]
- Zhou, Y.; Mowlazadeh Haghighi, S.; Liu, Z.; Wang, L.; Hruby, V.J.; Cai, M. Development of Ligand-Drug Conjugates Targeting Melanoma through the Overexpressed Melanocortin 1 Receptor. ACS Pharmacol. Transl. Sci. 2020, 3, 921–930. [Google Scholar] [CrossRef]
- Goodwin, R.G.; Kell, W.J.; Laidler, P.; Long, C.C.; Whatley, S.D.; McKinley, M.; Badminton, M.N.; Burnett, A.K.; Williams, G.T.; Elder, G.H. Photosensitivity and acute liver injury in myeloproliferative disorder secondary to late-onset protoporphyria caused by deletion of a ferrochelatase gene in hematopoietic cells. Blood 2006, 107, 60–62. [Google Scholar] [CrossRef]
- Haskell-Luevano, C.; Rosenquist, A.; Souers, A.; Khong, K.C.; Ellman, J.A.; Cone, R.D. Compounds that activate the mouse melanocortin-1 receptor identified by screening a small molecule library based upon the β-turn. J. Med. Chem. 1999, 42, 4380–4387. [Google Scholar] [CrossRef]
- Herpin, T.F.; Yu, G.; Carlson, K.E.; Morton, G.C.; Wu, X.; Kang, L.; Tuerdi, H.; Khanna, A.; Tokarski, J.S.; Lawrence, R.M.; et al. Discovery of tyrosine-based potent and selective melanocortin-1 receptor small-molecule agonists with anti-inflammatory properties. J. Med. Chem. 2003, 46, 1123–1126. [Google Scholar] [CrossRef] [PubMed]
- Boman, A.; Jonassen, T.E.N.; Lundstedt, T. Phenyl Pyrrole Aminoguanidine Derivatives. U.S. Patent 8,058,306, 15 November 2011. [Google Scholar]
- Bouix-Peter, C. Oxazetidine Derivatives, Process for Preparing Them and Use in Human Medicine and in Cosmetics. U.S. Patent 9,353,083, 31 May 2016. [Google Scholar]
- Eisinger, M.; Li, W.H.; Anthonavage, M.; Pappas, A.; Zhang, L.; Rossetti, D.; Huang, Q.; Seiberg, M. A melanocortin receptor 1 and 5 antagonist inhibits sebaceous gland differentiation and the production of sebum-specific lipids. J. Dermatol. Sci. 2011, 63, 23–32. [Google Scholar] [CrossRef]
- Lee, S.; Ju, W.; Tin, T.D.; Kim, J.; Lee, J.S.; Park, C.H.; Kwak, S.H. Effect of BMS-470539 on lipopolysaccharide-induced neutrophil activation. Korean J. Anesthesiol. 2020, 73, 151–157. [Google Scholar] [CrossRef]
- Suzuki, T.; Kawano, Y.; Matsumoto, A.; Kondo, M.; Funayama, K.; Tanemura, S.; Miyashiro, M.; Nishi, A.; Yamada, K.; Tsuda, M.; et al. Melanogenic effect of dersimelagon (MT-7117), a novel oral melanocortin 1 receptor agonist. Ski. Health Dis. 2022, 2, e78. [Google Scholar] [CrossRef]
- Lundstedt, T.; Skottner, A.; Boman, A.; Andersson, P.; Seifert, E.; Andrianov, V. N-Phenylpyrrole Guanidine Derivatives as Melanocortin Receptor Ligands. U.S. Patent 7,442,807, 28 October 2008. [Google Scholar]
- Montero-Melendez, T.; Gobbetti, T.; Cooray, S.N.; Jonassen, T.E.; Perretti, M. Biased agonism as a novel strategy to harness the proresolving properties of melanocortin receptors without eliciting melanogenic effects. J. Immunol. 2015, 194, 3381–3388. [Google Scholar] [CrossRef] [PubMed]
- Boiteau, J.G.; Rodeville, N.; Martin, C.; Tabet, S.; Moureou, C.; Muller, F.; Jetha, J.C.; Cardinaud, I. Development of a Kilogram-Scale Synthesis of a Novel MC1R Agonist. Org. Process Res. Dev. 2015, 19, 646–651. [Google Scholar] [CrossRef]
- Zhang, L.; Li, W.H.; Anthonavage, M.; Eisinger, M. Melanocortin-5 receptor: A marker of human sebocyte differentiation. Peptides 2006, 27, 413–420. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Wu, P.P.; Ciletti, N. Sebaceous epithelial cell differentiation requires cyclic adenosine monophosphate generation. In Vitro Cell. Dev. Biol. Anim. 2002, 38, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Anthonavage, M.; Huang, Q.; Li, W.H.; Eisinger, M. Proopiomelanocortin peptides and sebogenesis. Ann. N. Y. Acad. Sci. 2003, 994, 154–161. [Google Scholar] [CrossRef]
- Zouboulis, C.C. Acne and sebaceous gland function. Clin. Dermatol. 2004, 22, 360–366. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Schagen, S.; Alestas, T. The sebocyte culture: A model to study the pathophysiology of the sebaceous gland in sebostasis, seborrhoea and acne. Arch. Dermatol. Res. 2008, 300, 397–413. [Google Scholar] [CrossRef]
- Saint-Leger, D.; Bague, A.; Lefebvre, E.; Cohen, E.; Chivot, M. A possible role for squalene in the pathogenesis of acne. II. In vivo study of squalene oxides in skin surface and intra-comedonal lipids of acne patients. Br. J. Dermatol. 1986, 114, 543–552. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| MC1R | MC3R | MC4R | MC5R | |
| Compound | EC50(nM) | EC50(nM) | EC50(nM) | EC50(nM) |
| A | 28 ± 12 | NA | 2600 ± 200 | 4400 ± 1300 |
| B | 0.19 ± 0.02 | 250 ± 170 | 2.9 ± 0.04 | 2100 ± 400 |
| C | 0.35 ± 0.07 | 2200 ± 1000 | 23 ± 12 | 3900 ± 900 |
| D | 2.5 ± 1.3 | 7000 ± 1700 | 840 ± 150 | 9400 ± 200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mun, Y.; Kim, W.; Shin, D. Melanocortin 1 Receptor (MC1R): Pharmacological and Therapeutic Aspects. Int. J. Mol. Sci. 2023, 24, 12152. https://doi.org/10.3390/ijms241512152
Mun Y, Kim W, Shin D. Melanocortin 1 Receptor (MC1R): Pharmacological and Therapeutic Aspects. International Journal of Molecular Sciences. 2023; 24(15):12152. https://doi.org/10.3390/ijms241512152
Chicago/Turabian StyleMun, Yoonwoo, Woohyun Kim, and Dongyun Shin. 2023. "Melanocortin 1 Receptor (MC1R): Pharmacological and Therapeutic Aspects" International Journal of Molecular Sciences 24, no. 15: 12152. https://doi.org/10.3390/ijms241512152
APA StyleMun, Y., Kim, W., & Shin, D. (2023). Melanocortin 1 Receptor (MC1R): Pharmacological and Therapeutic Aspects. International Journal of Molecular Sciences, 24(15), 12152. https://doi.org/10.3390/ijms241512152