
Trapa bispinosa Roxb. Pericarp Extract Exerts 5α-Reductase Inhibitory Activity in Castrated Benign Prostatic Hyperplasia Model Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
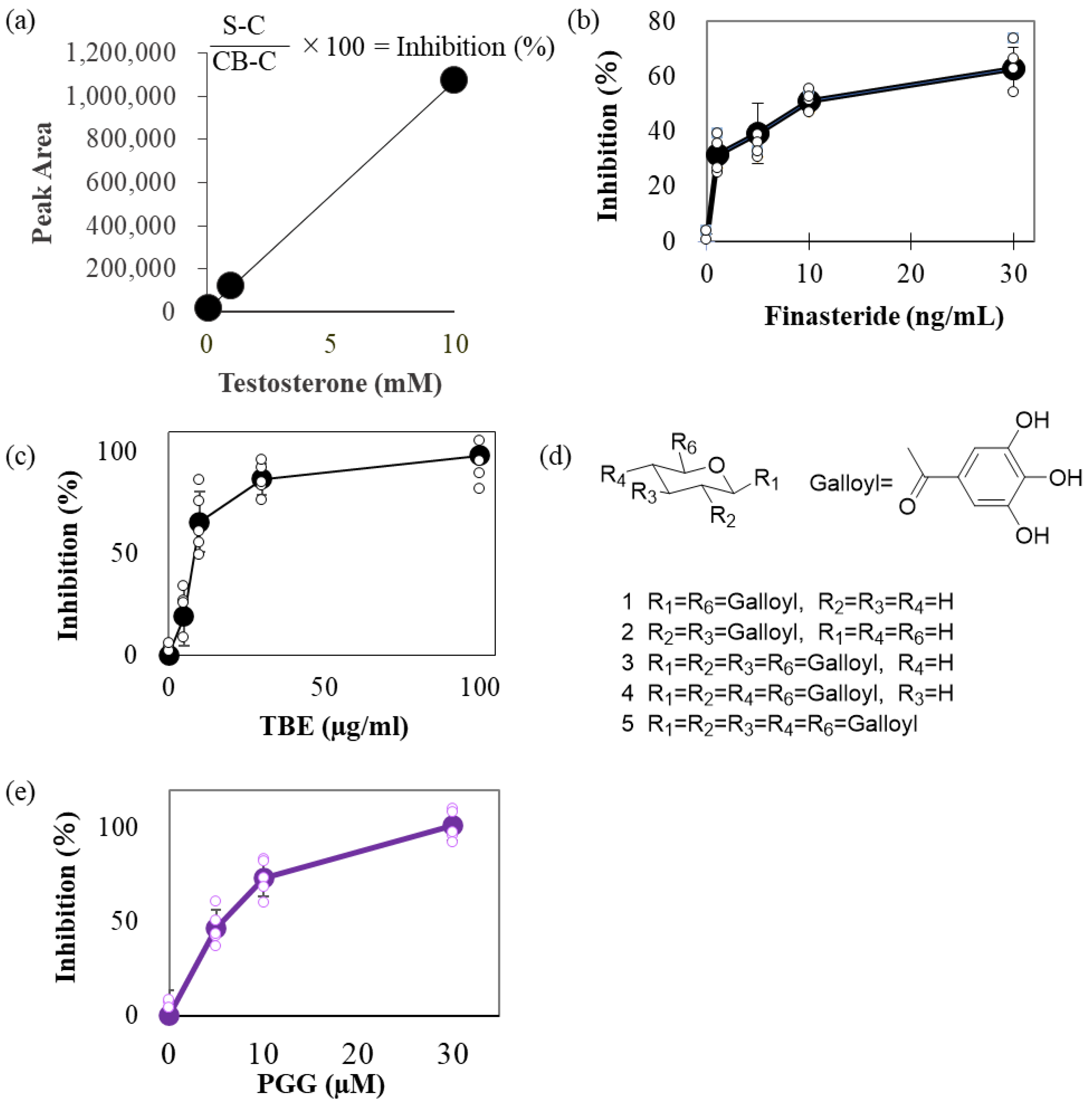
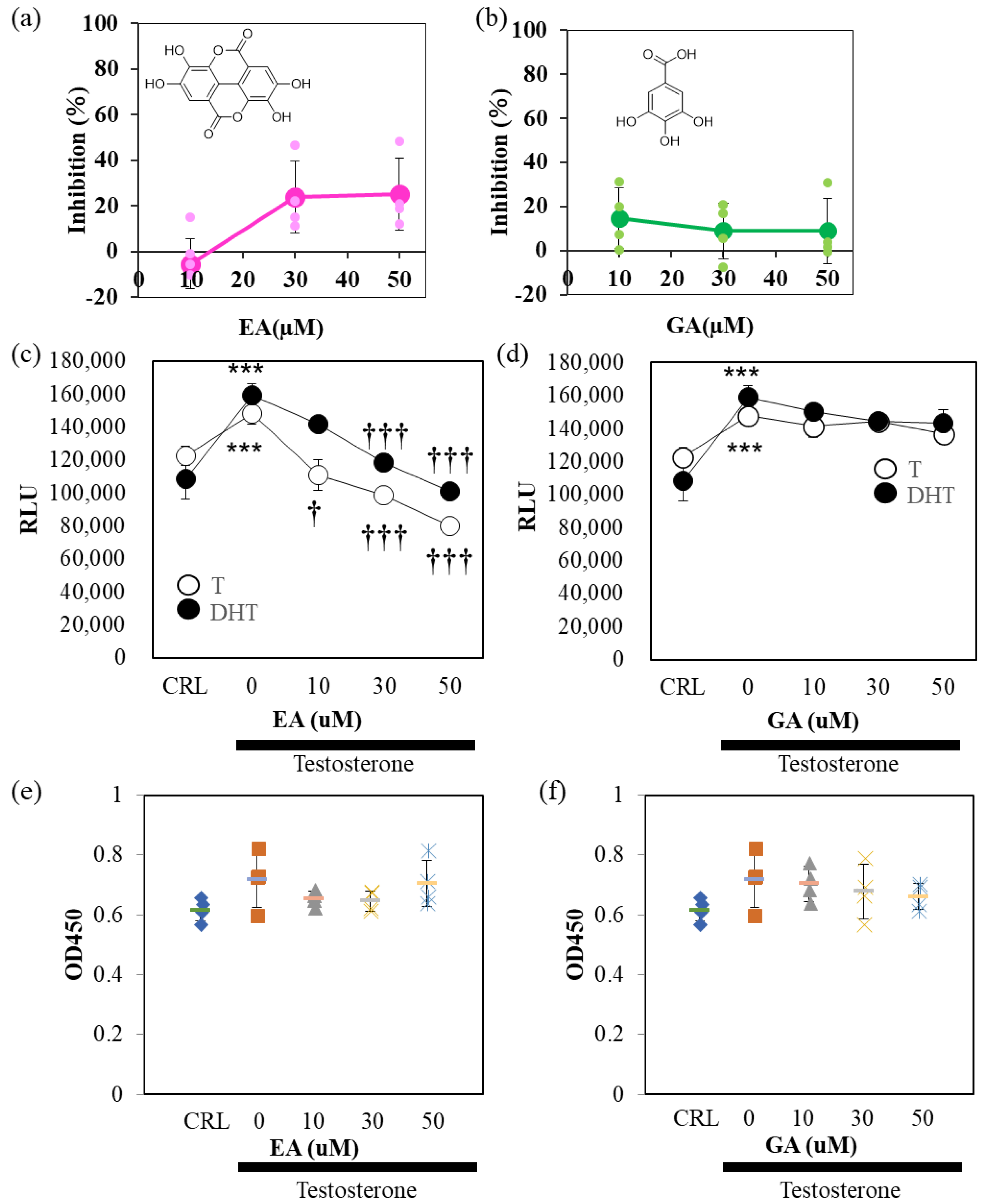
2.1. TBE Inhibited 5αR Activity
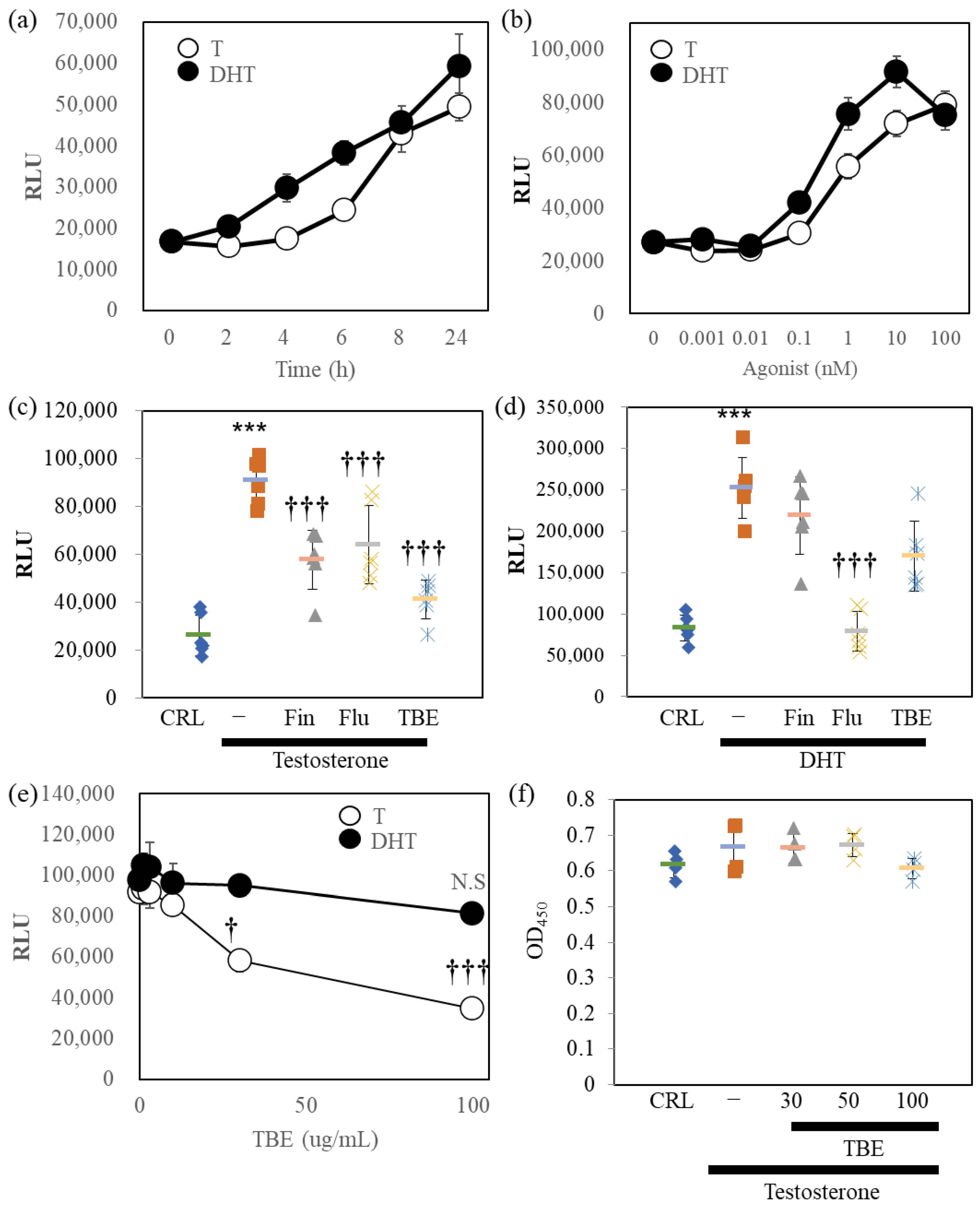
2.2. TBE Inhibited AR-Luc Reporter Induced by Testosterone
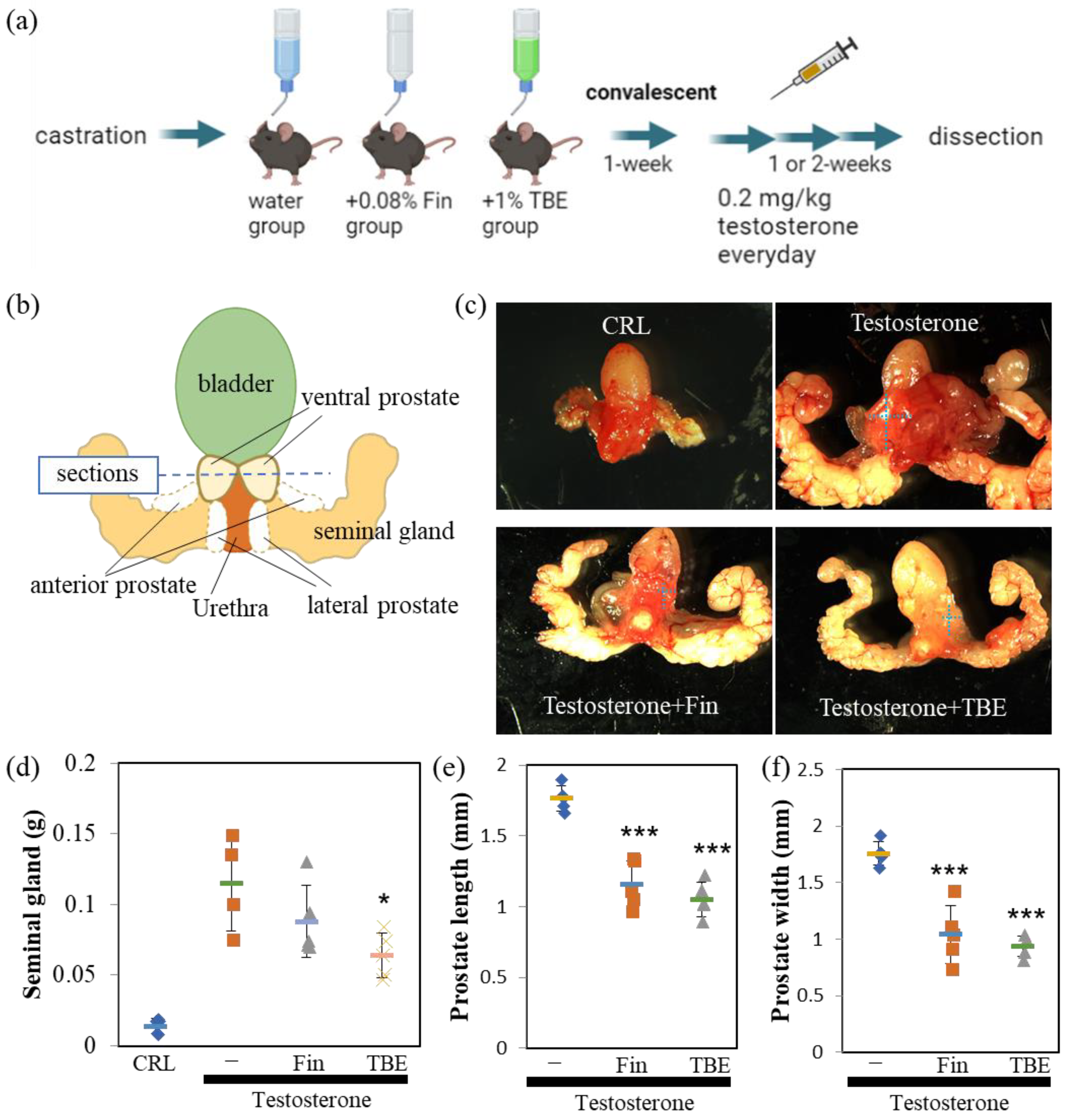
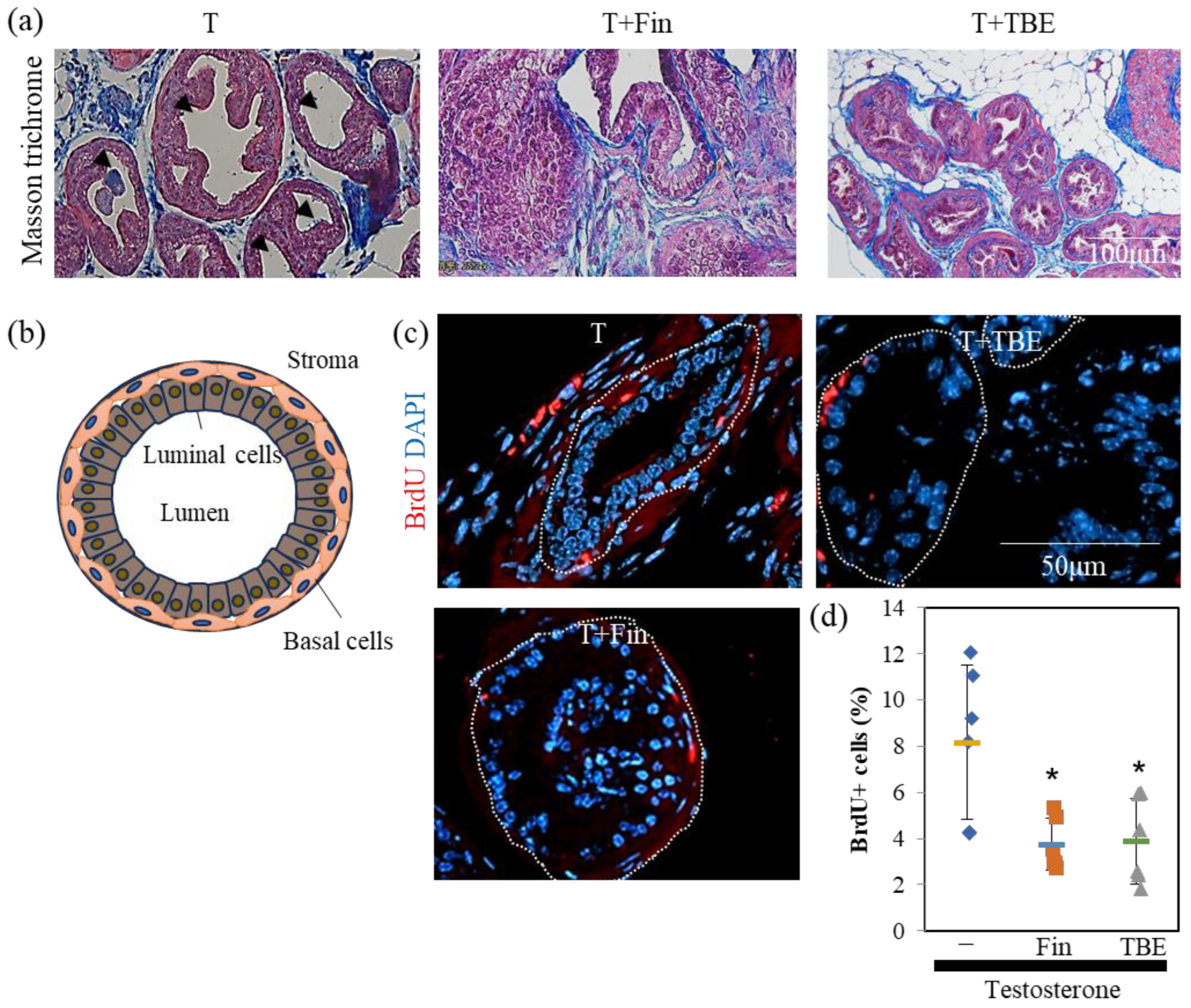
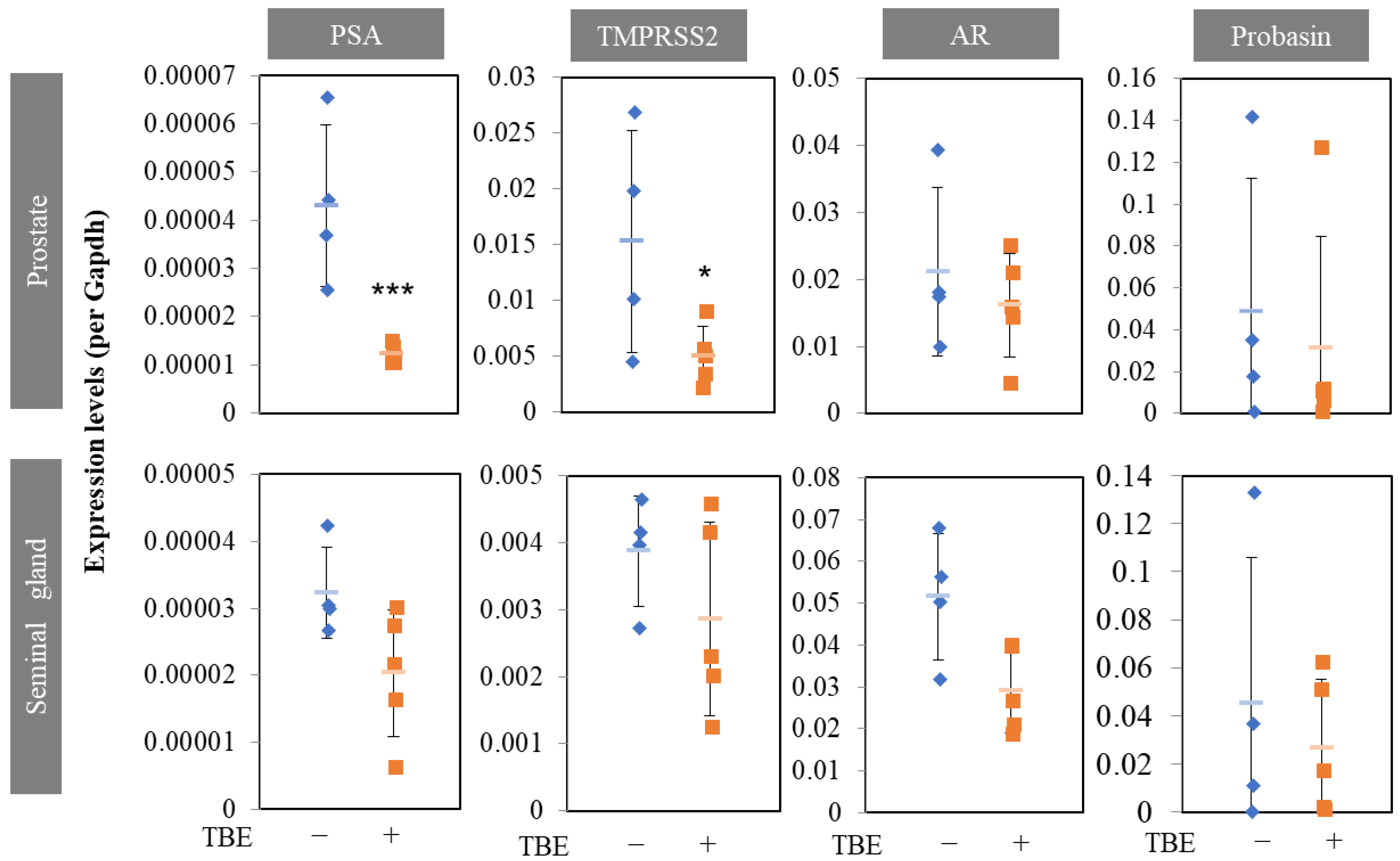
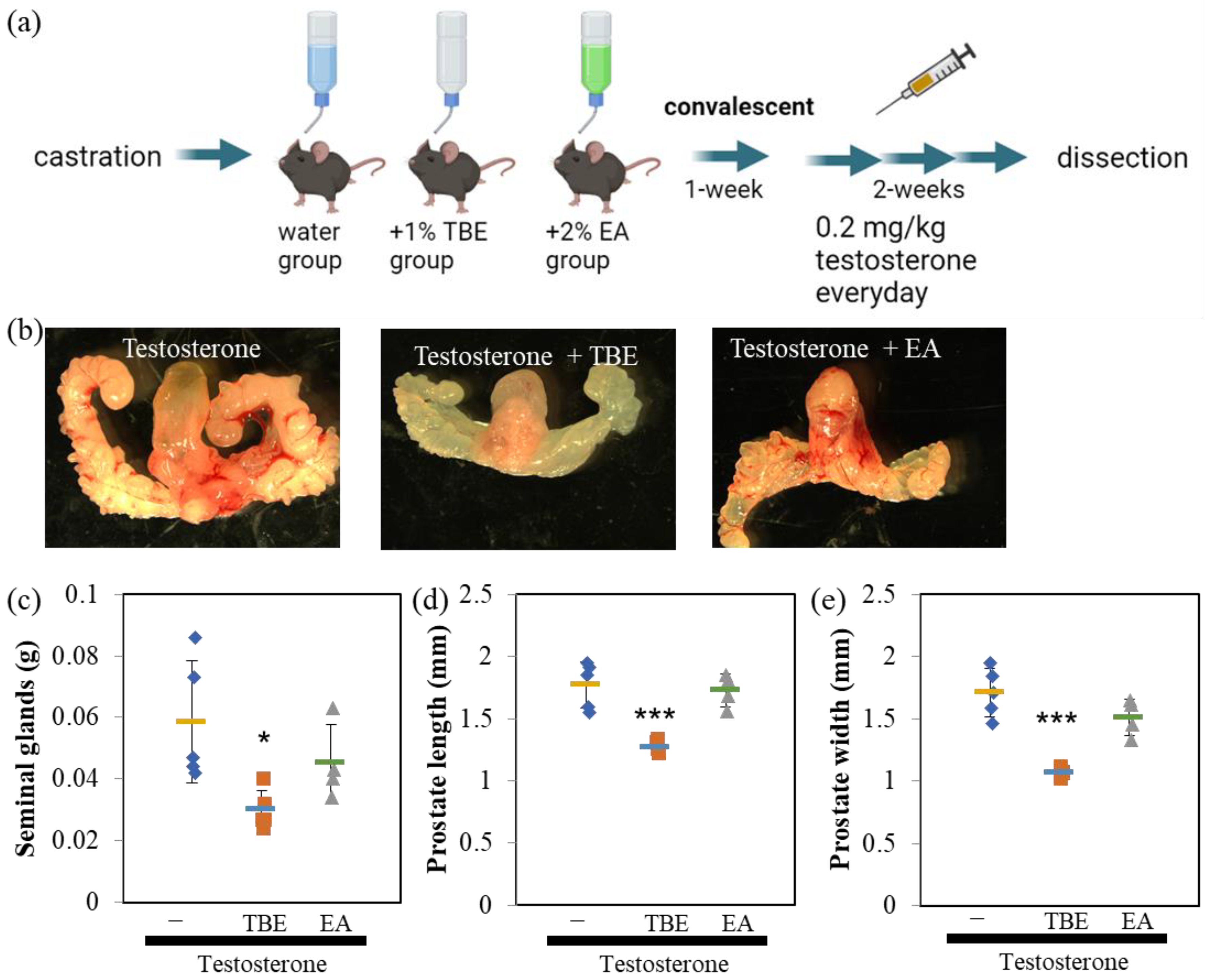
2.3. TBE Ameliorated Testosterone-Induced Prostatic Gland Formation and Increased Proliferation in Castrated BPH Mice
2.4. Ellagic Acid Inhibited 5αR Both In Vitro and In Vivo
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. High-Performance Liquid Chromatography (HPLC) Analysis
4.3. Reporter Cells and Cell Viability Test
4.4. Animal Experiments
4.5. Histology and Immunohistochemical Analysis
4.6. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (QPCR)
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iwaoka, Y.; Suzuki, S.; Kato, N.; Hayakawa, C.; Kawabe, S.; Ganeko, N.; Uemura, T.; Ito, H. Characterization and Identification of Bioactive Polyphenols in the Trapa bispinosa Roxb. Pericarp Extract. Molecules 2021, 26, 5802. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Gouvinhas, I.; Rocha, J.; Barros, A.I.R.N.A. Phytochemical and antioxidant analysis of medicinal and food plants towards bioactive food and pharmaceutical resources. Sci. Rep. 2021, 11, 10041. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Ikeoka, M.; Kondo, S. Skin-related enzyme inhibitory activity by hydrolyzable polyphenols in water chestnut (Trapa natans) husk. Biosci. Biotechnol. Biochem. 2021, 85, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Adkar, P.; Dongare, A.; Ambavade, S.; Bhaskar, V.H. Trapa bispinosa Roxb.: A Review on Nutritional and Pharmacological Aspects. Adv. Pharmacol. Sci. 2014, 2014, 959830. [Google Scholar] [CrossRef] [PubMed]
- Youness, R.A.; Kamel, R.; Elkasabgy, N.A.; Shao, P.; Farag, M.A. Recent Advances in Tannic Acid (Gallotannin) Anticancer Activities and Drug Delivery Systems for Efficacy Improvement; A Comprehensive Review. Molecules 2021, 26, 1486. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative Stress, Antioxidant Capabilities, and Bioavailability: Ellagic Acid or Urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef]
- Behrendt, P.; Perin, P.; Menzel, N.; Banda, D.; Pfaender, S.; Alves, M.P.; Thiel, V.; Meuleman, P.; Colpitts, C.C.; Schang, L.M.; et al. Pentagalloylglucose, a highly bioavailable polyphenolic compound present in Cortex moutan, efficiently blocks hepatitis C virus entry. Antivir. Res. 2017, 147, 19–28. [Google Scholar] [CrossRef]
- Takeshita, S.; Ishioka, Y.; Yagi, M.; Uemura, T.; Yamada, M.; Yonei, Y. The effects of water chestnut (Trapa bispinosa Roxb.) on the inhibition of glycometabolism and the improvement in postprandial blood glucose levels in humans. Glycative Stress Res. 2016, 3, 124–132. [Google Scholar]
- Csikós, E.; Horváth, A.; Ács, K.; Papp, N.; Balázs, V.L.; Dolenc, M.S.; Kenda, M.; Kočevar Glavač, N.; Nagy, M.; Protti, M.; et al. Treatment of Benign Prostatic Hyperplasia by Natural Drugs. Molecules 2021, 26, 7141. [Google Scholar] [CrossRef]
- Jin, R.; Forbes, C.; Miller, N.L.; Strand, D.; Case, T.; Cates, J.M.; Kim, H.H.; Wages, P.; Porter, N.A.; Mantione, K.M.; et al. Glucocorticoids are induced while dihydrotestosterone levels are suppressed in 5-alpha reductase inhibitor treated human benign prostate hyperplasia patients. Prostate 2022, 82, 1378–1388. [Google Scholar] [CrossRef]
- Kim, J.; Coetzee, G.A. Prostate specific antigen gene regulation by androgen receptor. J. Cell. Biochem. 2004, 93, 233–241. [Google Scholar] [CrossRef]
- Clinckemalie, L.; Spans, L.; Dubois, V.; Laurent, M.; Helsen, C.; Joniau, S.; Claessens, F. Androgen regulation of the TMPRSS2 gene and the effect of a SNP in an androgen response element. Mol. Endocrinol. 2013, 27, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Koseki, J.; Matsumoto, T.; Matsubara, Y.; Tsuchiya, K.; Mizuhara, Y.; Sekiguchi, K.; Nishimura, H.; Watanabe, J.; Kaneko, A.; Hattori, T.; et al. Inhibition of Rat 5α-Reductase Activity and Testosterone-Induced Sebum Synthesis in Hamster Sebocytes by an Extract of Quercus acutissima Cortex. Evid. Based Complement. Altern. Med. 2015, 2015, 853846. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Ho, C.T.; Lin, J.K. Theaflavin-3,3’-digallate and penta-O-galloyl-β-D-glucose inhibit rat liver microsomal 5α-reductase activity and the expression of androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 2004, 25, 1109–1118. [Google Scholar] [CrossRef]
- Joo, K.J.; Sung, W.S.; Park, S.H.; Yang, W.J.; Kim, T.H. Comparison of α-blocker monotherapy and α-blocker plus 5α-reductase inhibitor combination therapy based on prostate volume for treatment of benign prostatic hyperplasia. J. Int. Med. Res. 2012, 40, 899–908. [Google Scholar] [CrossRef]
- Mizokami, A.; Koh, E.; Izumi, K.; Narimoto, K.; Takeda, M.; Honma, S.; Dai, J.; Keller, E.T.; Namiki, M. Prostate cancer stromal cells and LNCaP cells coordinately activate the androgen receptor through synthesis of testosterone and dihydrotestosterone from dehydroepiandrosterone. Endocr. Relat. Cancer 2009, 16, 1139–1155. [Google Scholar] [CrossRef]
- Keil, K.P.; Mehta, V.; Abler, L.L.; Joshi, P.S.; Schmitz, C.T.; Vezina, C.M. Visualization and quantification of mouse prostate development by in situ hybridization. Differentiation 2012, 84, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Vezina, C.M.; Isaacson, D.; Ricke, W.A.; Timms, B.G.; Cao, M.; Franco, O.; Baskin, L.S. Development of the human prostate. Differentiation 2018, 103, 24–45. [Google Scholar] [CrossRef]
- Srivilai, J.; Minale, G.; Scholfield, C.N.; Ingkaninan, K. Discovery of Natural Steroid 5 Alpha-Reductase Inhibitors. Assay Drug Dev. Technol. 2019, 17, 44–57. [Google Scholar] [CrossRef]
- Azizi, A.; Mumin, N.H.; Shafqat, N. Phytochemicals with Anti 5-alpha-reductase Activity: A Prospective for Prostate Cancer Treatment. F1000Research 2021, 10, 221. [Google Scholar] [CrossRef]
- Chen, F.; Knecht, K.; Birzin, E.; Fisher, J.; Wilkinson, H.; Mojena, M.; Moreno, C.T.; Schmidt, A.; Harada, S.; Freedman, L.P.; et al. Direct agonist/antagonist functions of dehydroepiandrosterone. Endocrinology 2005, 146, 4568–4576. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.G.; Ko, E.B.; Choi, K.C. Gallic acid, a phenolic acid, hinders the progression of prostate cancer by inhibition of histone deacetylase 1 and 2 expression. J. Nutr. Biochem. 2020, 84, 108444. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.T.; Altuwaijri, S.; Ricke, W.A.; Huang, S.P.; Yeh, S.; Zhang, C.; Niu, Y.; Tsai, M.Y.; Chang, C. Increased prostate cell proliferation and loss of cell differentiation in mice lacking prostate epithelial androgen receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 12679–12684. [Google Scholar] [CrossRef] [PubMed]
- Yeung, L.H.Y.; Read, J.T.; Sorenson, P.; Nelson, C.C.; Jia, W.; Rennie, P.S. Identification and characterization of a prostate-specific androgen-independent protein-binding site in the probasin promoter. Biochem. J. 2003, 371, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y.; et al. Impact of Gallic Acid on Gut Health: Focus on the Gut Microbiome, Immune Response, and Mechanisms of Action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef]
- Takeshita, S.; Yagi, M.; Uemura, T.; Yamada, M.; Yonei, Y. Peel extract of water chestnut (Trapa bispinosa Roxb.) inhibits glycation, degradesα-dicarbonyl compound, and breaks advanced glycation end product crosslinks. Glycative Stress Res. 2015, 2, 72–79. [Google Scholar]
- Morikawa, T.; Luo, F.; Manse, Y.; Sugita, H.; Saeki, S.; Chaipech, S.; Pongpiriyadacha, Y.; Muraoka, O.; Ninomiya, K. Geranylated Coumarins from Thai Medicinal Plant Mammea siamensis with Testosterone 5α-Reductase Inhibitory Activity. Front. Chem. 2020, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Sonneveld, E.; Jansen, H.J.; Riteco, J.A.; Brouwer, A.; van der Burg, B. Development of androgen- and estrogen-responsive bioassays, members of a panel of human cell line-based highly selective steroid-responsive bioassays. Toxicol. Sci. 2005, 83, 136–148. [Google Scholar] [CrossRef]
- Natsume, C.; Aoki, N.; Aoyama, T.; Senda, K.; Matsui, M.; Ikegami, A.; Tanaka, K.; Azuma, Y.T.; Fujita, T. Fucoxanthin Ameliorates Atopic Dermatitis Symptoms by Regulating Keratinocytes and Regulatory Innate Lymphoid Cells. Int. J. Mol. Sci. 2020, 21, 2180. [Google Scholar] [CrossRef]
- Lofgren, J.L.; Erdman, S.E.; Hewes, C.; Wong, C.; King, R.; Chavarria, T.E.; Discua, A.R.; Fox, J.G.; Maurer, K.J. Castration eliminates conspecific aggression in group-housed CD1 male surveillance mice (Mus musculus). J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 594–599. [Google Scholar]
- Azuma, Y.T.; Fujita, T.; Izawa, T.; Hirota, K.; Nishiyama, K.; Ikegami, A.; Aoyama, T.; Ike, M.; Ushikai, Y.; Kuwamura, M.; et al. IL-19 Contributes to the Development of Nonalcoholic Steatohepatitis by Altering Lipid Metabolism. Cells 2021, 10, 3513. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujita, T.; Aoyama, T.; Uemura, T.; Takeshita, S.; Yamasaki, T.; Heijou, H.; Morimoto, K. Trapa bispinosa Roxb. Pericarp Extract Exerts 5α-Reductase Inhibitory Activity in Castrated Benign Prostatic Hyperplasia Model Mice. Int. J. Mol. Sci. 2023, 24, 11765. https://doi.org/10.3390/ijms241411765
Fujita T, Aoyama T, Uemura T, Takeshita S, Yamasaki T, Heijou H, Morimoto K. Trapa bispinosa Roxb. Pericarp Extract Exerts 5α-Reductase Inhibitory Activity in Castrated Benign Prostatic Hyperplasia Model Mice. International Journal of Molecular Sciences. 2023; 24(14):11765. https://doi.org/10.3390/ijms241411765
Chicago/Turabian StyleFujita, Takashi, Tomoko Aoyama, Tomohiro Uemura, Shouko Takeshita, Takuto Yamasaki, Hiroko Heijou, and Koji Morimoto. 2023. "Trapa bispinosa Roxb. Pericarp Extract Exerts 5α-Reductase Inhibitory Activity in Castrated Benign Prostatic Hyperplasia Model Mice" International Journal of Molecular Sciences 24, no. 14: 11765. https://doi.org/10.3390/ijms241411765
APA StyleFujita, T., Aoyama, T., Uemura, T., Takeshita, S., Yamasaki, T., Heijou, H., & Morimoto, K. (2023). Trapa bispinosa Roxb. Pericarp Extract Exerts 5α-Reductase Inhibitory Activity in Castrated Benign Prostatic Hyperplasia Model Mice. International Journal of Molecular Sciences, 24(14), 11765. https://doi.org/10.3390/ijms241411765