

A Short-Term Model of Colitis-Associated Colorectal Cancer That Suggests Initial Tumor Development and the Characteristics of Cancer Stem Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
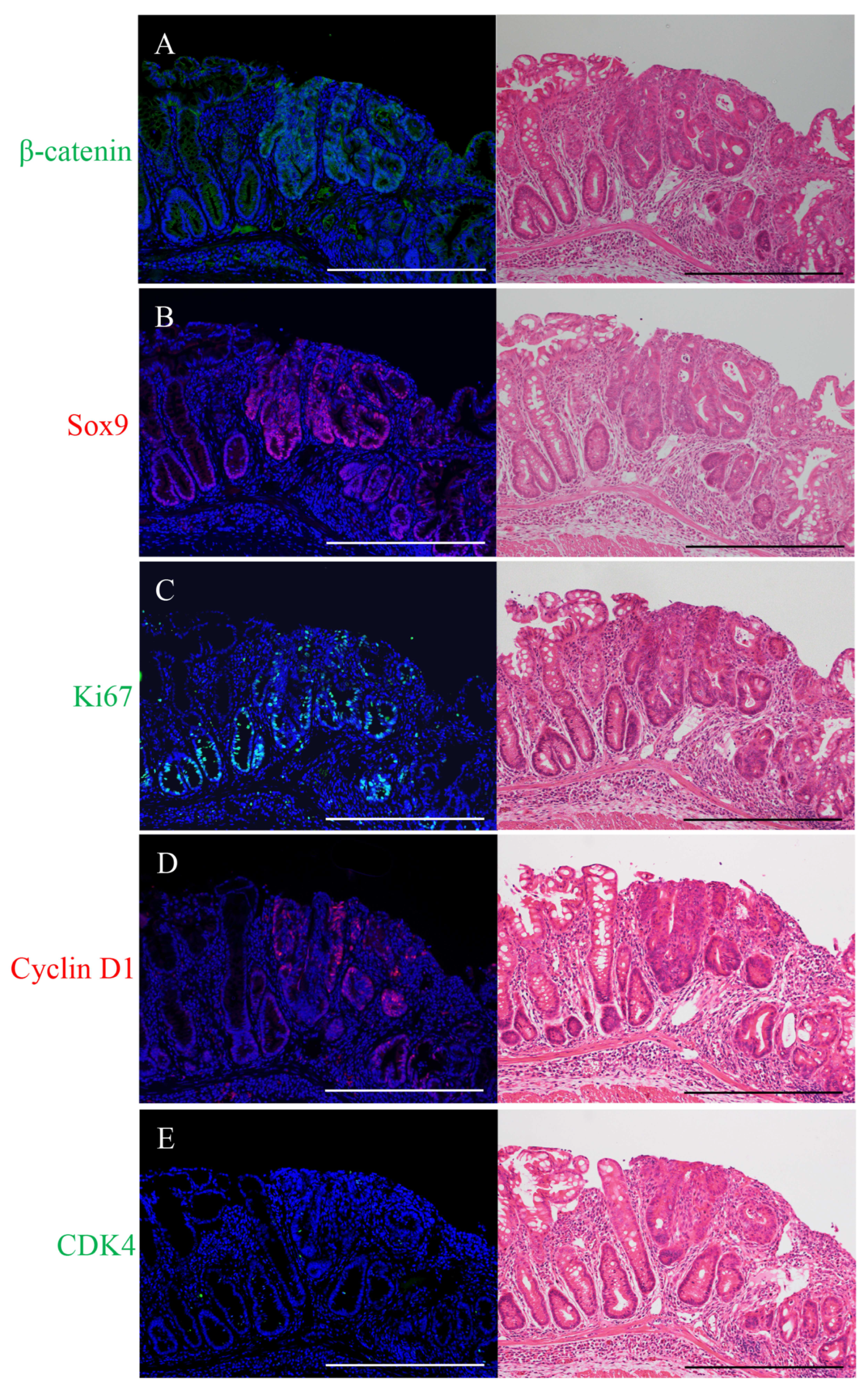
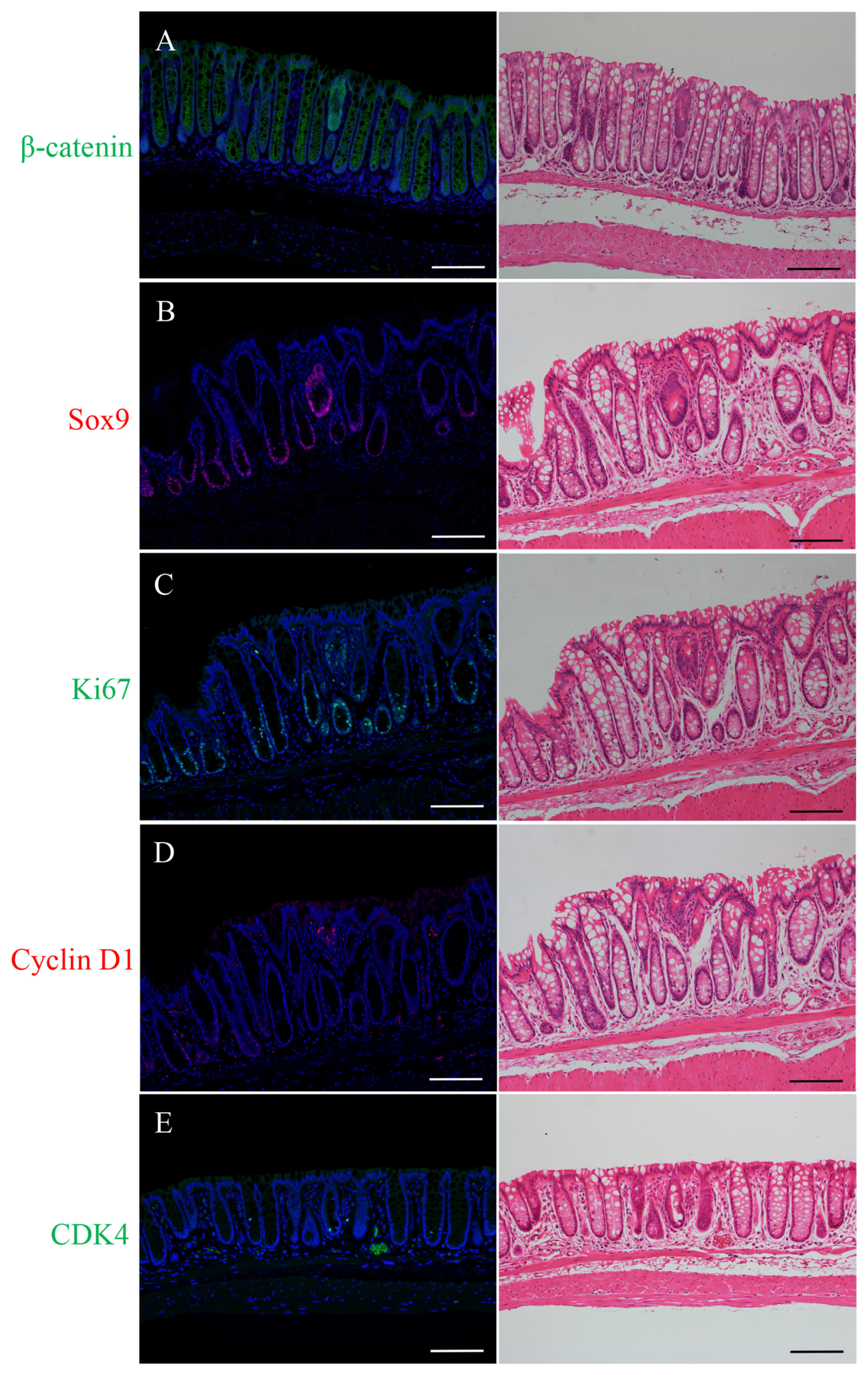
2.1. Immunofluorescence Staining for β-Catenin, Sox9, Ki67, Cyclin D1, or CDK4 and H&E Staining in the Micro-Neoplastic Lesions of the AOM/DSS Mice
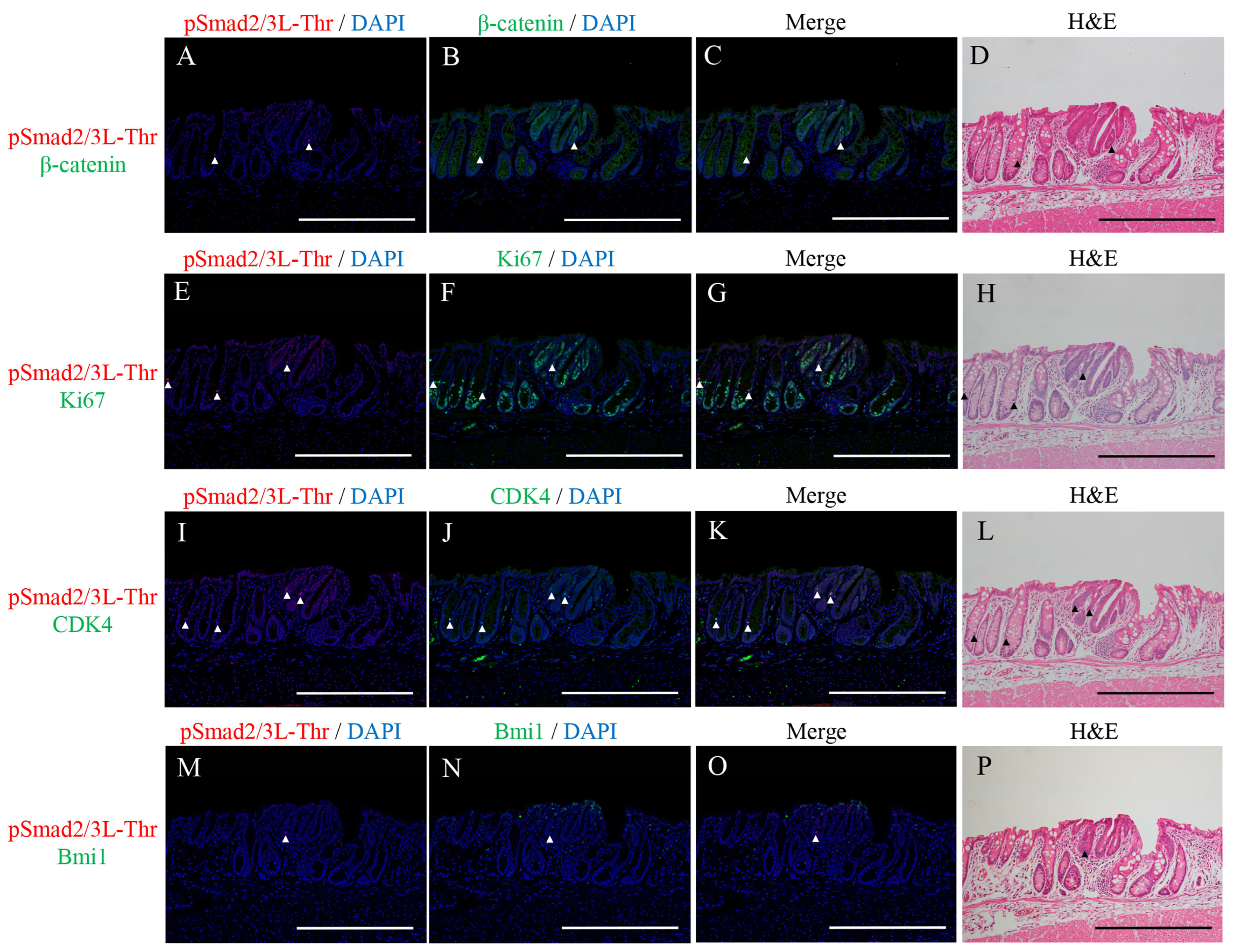
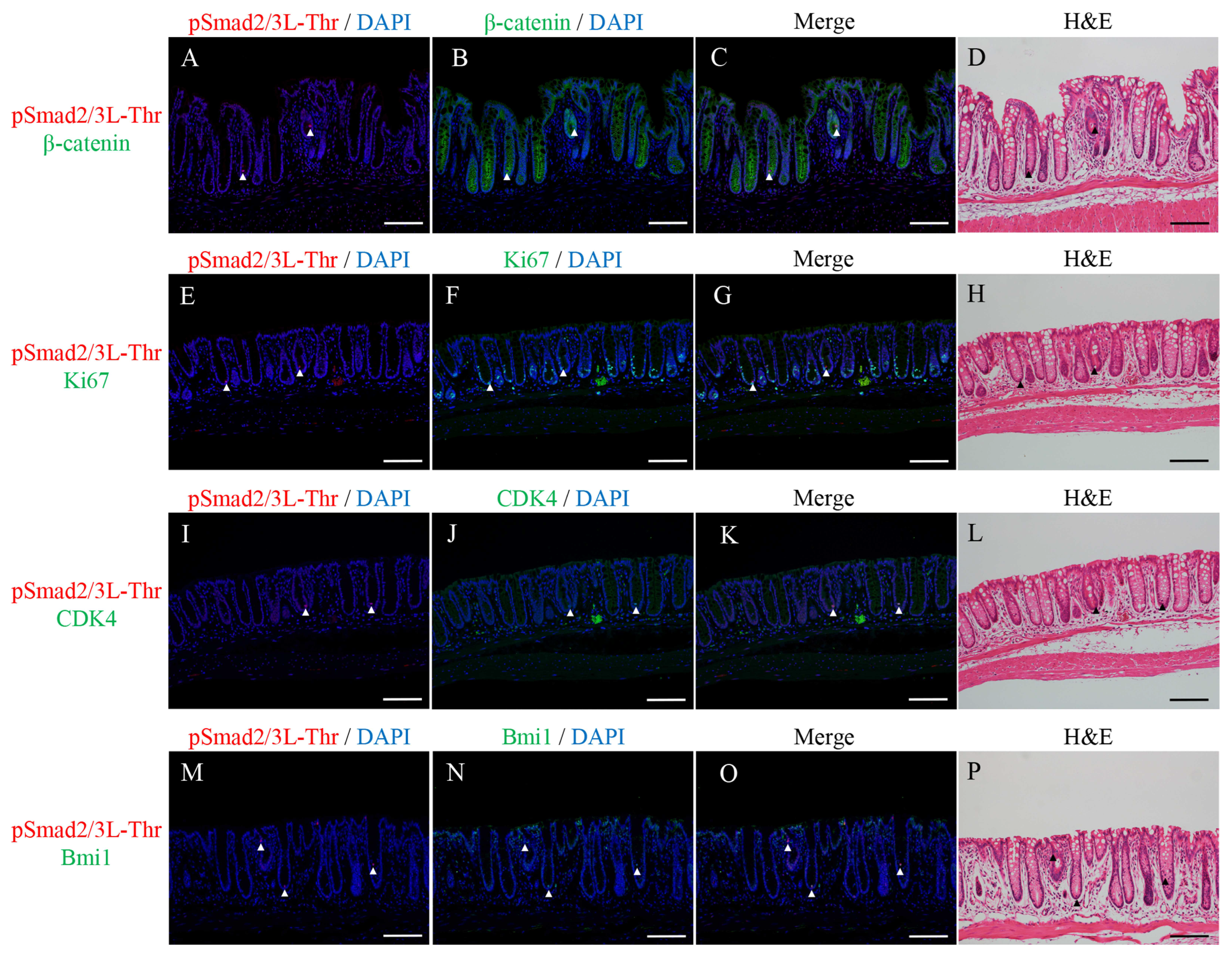
2.2. Double Immunofluorescence Staining for pSmad2/3L-Thr with β-Catenin, Ki67, CDK4, or Bmi1 and H&E Staining in the Micro-Neoplastic Lesions of the AOM/DSS Mice
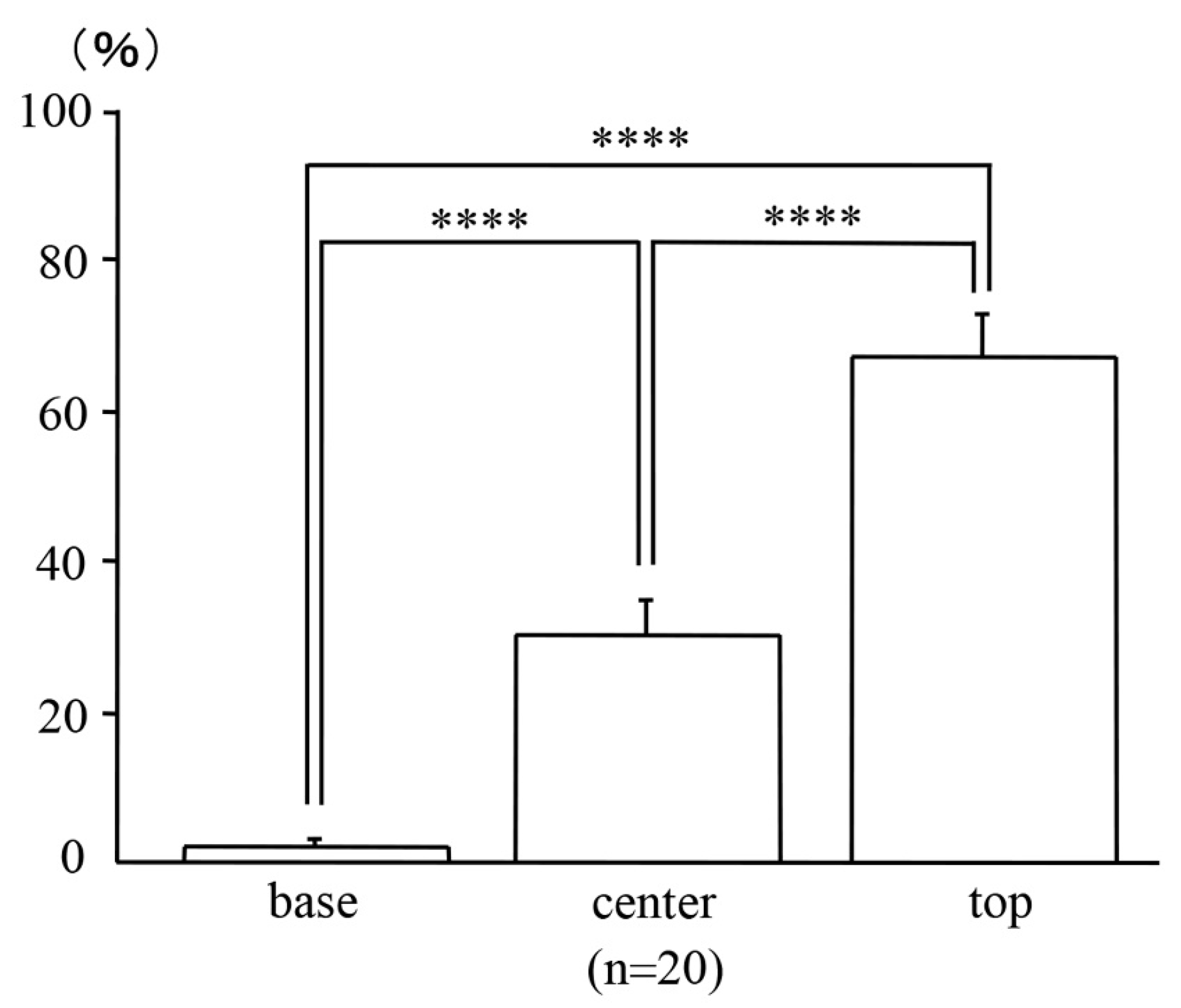
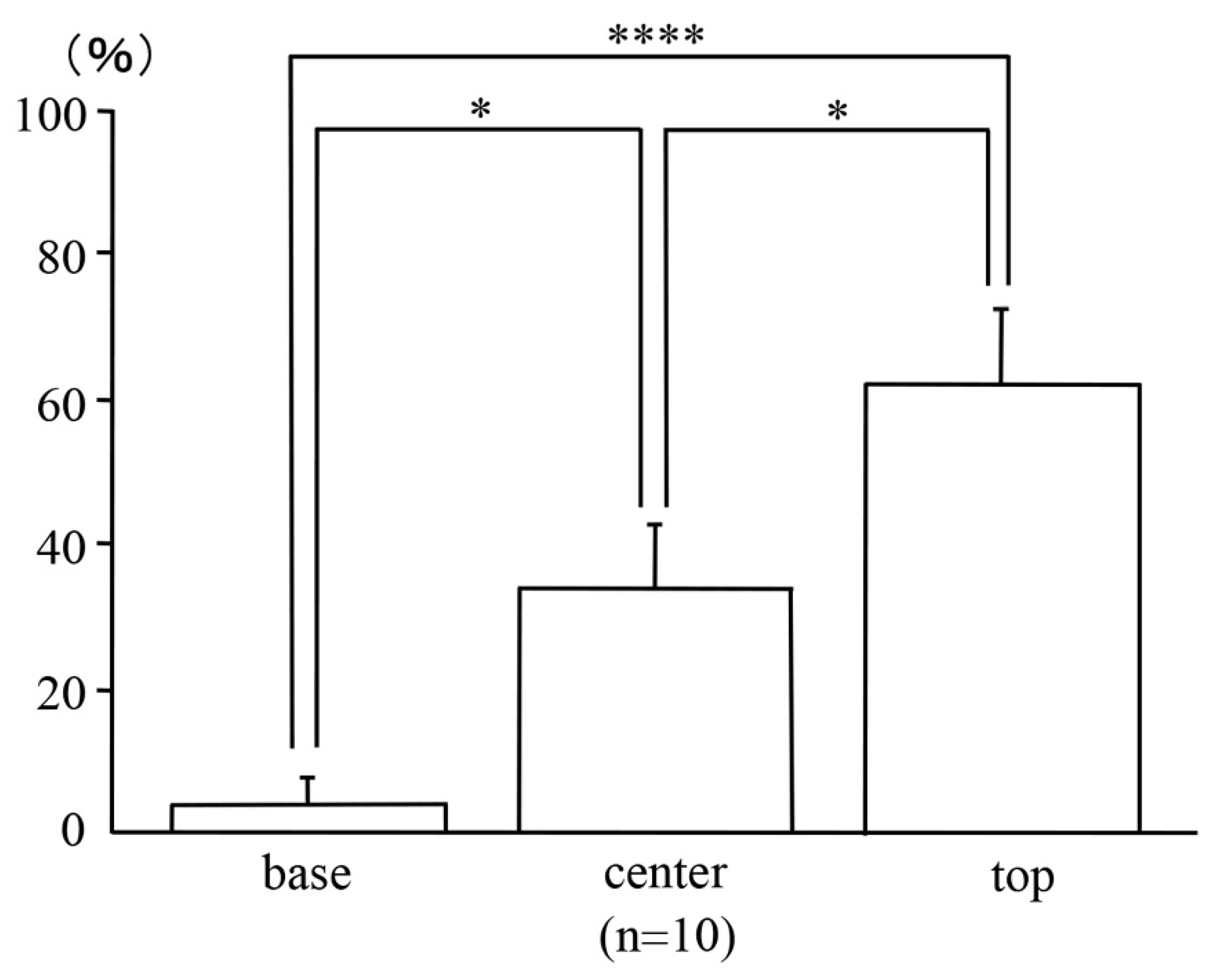
2.3. Locations of the Micro-Neoplastic Lesions in the AOM/DSS Mice within the Mucosae
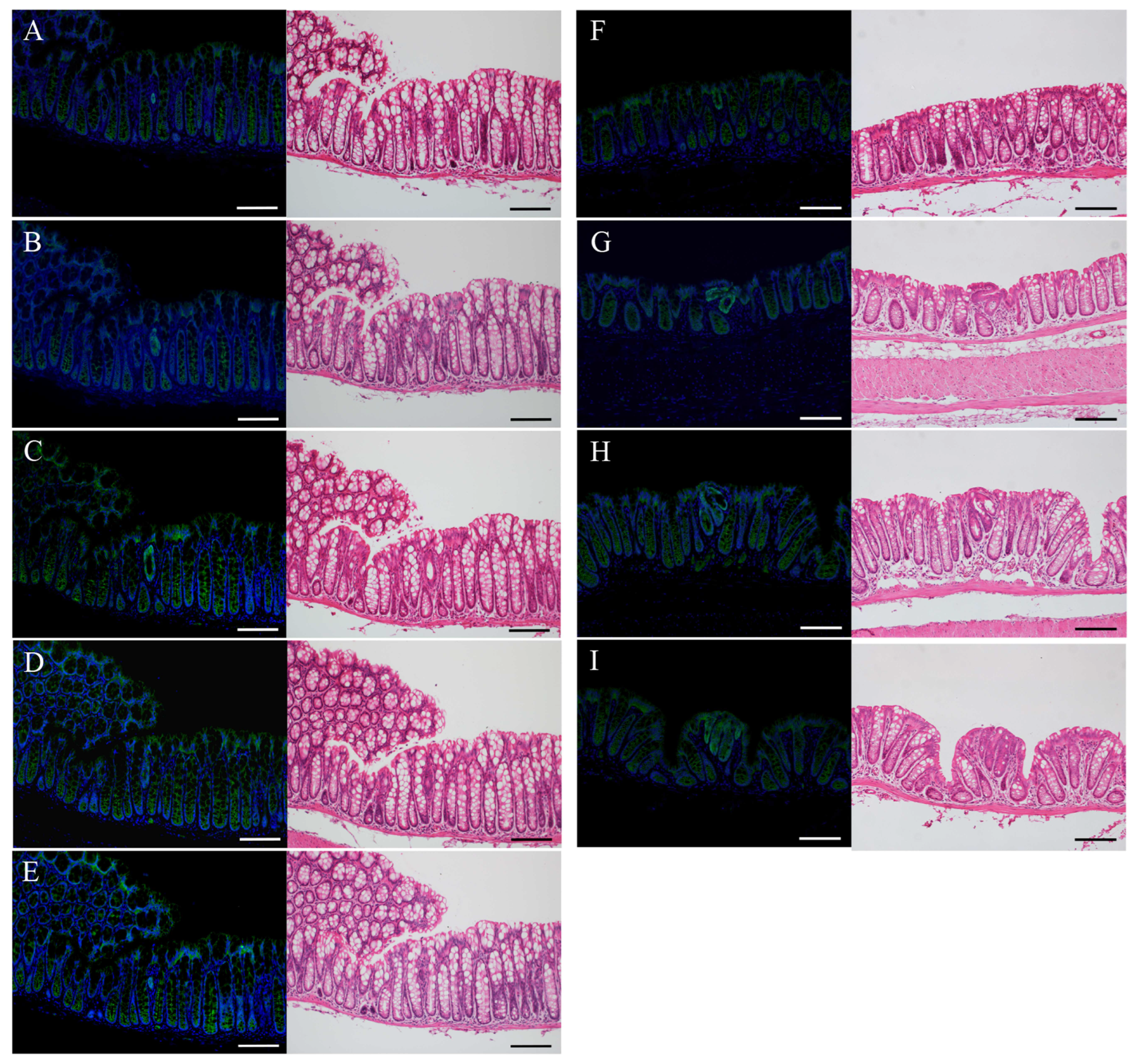
2.4. Immunofluorescence Staining for β-Catenin and H&E Staining in the Uni-, Bi-, or Tri-Cryptal Neoplastic and Micro-Neoplastic Lesions of the AOM/DSS Mice
2.5. Immunofluorescence Staining for β-Catenin, Sox9, Ki67, Cyclin D1, or CDK4 and H&E Staining in the Uni-Cryptal Neoplastic Lesions of the AOM/DSS Mice
2.6. Double Immunofluorescence Staining for pSmad2/3L-Thr with β-Catenin, Ki67, CDK4, or Bmi1 and H&E Staining in the Uni-Cryptal Neoplastic Lesions of the AOM/DSS Mice
2.7. Locations of the Uni-Cryptal Neoplastic Lesions in the AOM/DSS Mice within the Mucosae
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Chemicals
4.3. Experimental Design
4.4. Histopathological Analysis
4.5. Domain-Specific Antibodies against the Phosphorylated Smad2 and Smad3
4.6. Immunohistochemistry
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Ekbom, A.; Helmick, C.; Zack, M.; Adami, H.O. Ulcerative colitis and colorectal cancer. A population-based study. N. Engl. J. Med. 1990, 323, 1228–1233. [Google Scholar] [CrossRef] [PubMed]
- Seril, D.N.; Liao, J.; Yang, G.Y.; Yang, C.S. Oxidative stress and ulcerative colitis-associated carcinogenesis: Studies in humans and animal models. Carcinogenesis 2003, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Development of an inflammation-associated colorectal cancer model and its application for research on carcinogenesis and chemoprevention. Int. J. Inflammat. 2012, 2012, 658786. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Colorectal carcinogenesis: Review of human and experimental animal studies. J. Carcinogen. 2009, 8, 5. [Google Scholar] [CrossRef]
- Campbell, L.L.; Polyak, K. Breast tumor heterogeneity: Cancer stem cells or clonal evolution? Cell Cycle 2007, 6, 2332–2338. [Google Scholar] [CrossRef]
- Clarke, M.F.; Dick, J.E.; Dirks, P.B.; Eaves, C.J.; Jamieson, C.H.; Jones, D.L.; Visvader, J.; Weissman, I.L.; Wahl, G.M. Cancer stem cells--perspectives on current status and future directions: Aacr workshop on cancer stem cells. Cancer Res. 2006, 66, 9339–9344. [Google Scholar] [CrossRef]
- Barker, N.; Ridgway, R.A.; van Es, J.H.; van de Wetering, M.; Begthel, H.; van den Born, M.; Danenberg, E.; Clarke, A.R.; Sansom, O.J.; Clevers, H. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 2009, 457, 608–611. [Google Scholar] [CrossRef]
- Vicente-Duenas, C.; Gutierrez de Diego, J.; Rodriguez, F.D.; Jimenez, R.; Cobaleda, C. The role of cellular plasticity in cancer development. Curr. Med. Chem. 2009, 16, 3676–3685. [Google Scholar] [CrossRef]
- Shih, I.M.; Wang, T.L.; Traverso, G.; Romans, K.; Hamilton, S.R.; Ben-Sasson, S.; Kinzler, K.W.; Vogelstein, B. Top-down morphogenesis of colorectal tumors. Proc. Nat. Acad. Sci. USA 2001, 98, 2640–2645. [Google Scholar] [CrossRef] [PubMed]
- Kukitsu, T.; Takayama, T.; Miyanishi, K.; Nobuoka, A.; Katsuki, S.; Sato, Y.; Takimoto, R.; Matsunaga, T.; Kato, J.; Sonoda, T.; et al. Aberrant crypt foci as precursors of the dysplasia-carcinoma sequence in patients with ulcerative colitis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Yoshitani, S.; Takashima, S.; Okumura, A.; Murakami, A.; Hosokawa, M. Ligands for peroxisome proliferator-activated receptors alpha and gamma inhibit chemically induced colitis and formation of aberrant crypt foci in rats. Cancer Res. 2001, 61, 2424–2428. [Google Scholar] [PubMed]
- Whiteley, L.O.; Hudson, L., Jr.; Pretlow, T.P. Aberrant crypt foci in the colonic mucosa of rats treated with a genotoxic and nongenotoxic colon carcinogen. Toxicol. Pathol. 1996, 24, 681–689. [Google Scholar] [CrossRef]
- Schwitalla, S.; Fingerle, A.A.; Cammareri, P.; Nebelsiek, T.; Goktuna, S.I.; Ziegler, P.K.; Canli, O.; Heijmans, J.; Huels, D.J.; Moreaux, G.; et al. Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem-cell-like properties. Cell 2013, 152, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. Tgf-beta signalling from cell membrane to nucleus through smad proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef]
- Massague, J. Tgf-beta signal transduction. Ann. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef]
- Wrana, J.L. Crossing smads. Sci. STKE Signal Transduct. Knowl. Environ. 2000, 2000, re1. [Google Scholar] [CrossRef]
- Kretzschmar, M.; Doody, J.; Timokhina, I.; Massague, J. A mechanism of repression of tgfbeta/smad signaling by oncogenic ras. Genes Dev. 1999, 13, 804–816. [Google Scholar] [CrossRef]
- Matsuura, I.; Denissova, N.G.; Wang, G.; He, D.; Long, J.; Liu, F. Cyclin-dependent kinases regulate the antiproliferative function of smads. Nature 2004, 430, 226–231. [Google Scholar] [CrossRef]
- Mori, S.; Matsuzaki, K.; Yoshida, K.; Furukawa, F.; Tahashi, Y.; Yamagata, H.; Sekimoto, G.; Seki, T.; Matsui, H.; Nishizawa, M.; et al. Tgf-beta and hgf transmit the signals through jnk-dependent smad2/3 phosphorylation at the linker regions. Oncogene 2004, 23, 7416–7429. [Google Scholar] [CrossRef] [PubMed]
- Tarasewicz, E.; Jeruss, J.S. Phospho-specific smad3 signaling: Impact on breast oncogenesis. Cell Cycle 2012, 11, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Smad3 phosphoisoform-mediated signaling during sporadic human colorectal carcinogenesis. Histol. Histopathol. 2006, 21, 645–662. [Google Scholar] [PubMed]
- Matsuzaki, K.; Kitano, C.; Murata, M.; Sekimoto, G.; Yoshida, K.; Uemura, Y.; Seki, T.; Taketani, S.; Fujisawa, J.; Okazaki, K. Smad2 and smad3 phosphorylated at both linker and cooh-terminal regions transmit malignant tgf-beta signal in later stages of human colorectal cancer. Cancer Res. 2009, 69, 5321–5330. [Google Scholar] [CrossRef]
- Sapkota, G.; Knockaert, M.; Alarcon, C.; Montalvo, E.; Brivanlou, A.H.; Massague, J. Dephosphorylation of the linker regions of smad1 and smad2/3 by small c-terminal domain phosphatases has distinct outcomes for bone morphogenetic protein and transforming growth factor-beta pathways. J. Biol. Chem. 2006, 281, 40412–40419. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and smad-independent pathways in tgf-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Kishimoto, M.; Fukui, T.; Suzuki, R.; Takahashi, Y.; Sumimoto, K.; Okazaki, T.; Sakao, M.; Sakaguchi, Y.; Yoshida, K.; Uchida, K.; et al. Phosphorylation of smad2/3 at specific linker threonine indicates slow-cycling intestinal stem-like cells before reentry to cell cycle. Digest. Dis. Sci. 2015, 60, 362–374. [Google Scholar] [CrossRef]
- Suzuki, R.; Fukui, T.; Kishimoto, M.; Miyamoto, S.; Takahashi, Y.; Takeo, M.; Mitsuyama, T.; Sakaguchi, Y.; Uchida, K.; Nishio, A.; et al. Smad2/3 linker phosphorylation is a possible marker of cancer stem cells and correlates with carcinogenesis in a mouse model of colitis-associated colorectal cancer. J. Crohn’s Colitis 2015, 9, 565–574. [Google Scholar] [CrossRef]
- Tanimura, Y.; Fukui, T.; Horitani, S.; Matsumoto, Y.; Miyamoto, S.; Suzuki, R.; Tanaka, T.; Tomiyama, T.; Ikeura, T.; Ando, Y.; et al. Long-term model of colitis-associated colorectal cancer suggests tumor spread mechanism and nature of cancer stem cells. Oncol. Lett. 2021, 21, 7. [Google Scholar] [CrossRef]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Okayasu, I.; Yamada, M.; Mikami, T.; Yoshida, T.; Kanno, J.; Ohkusa, T. Dysplasia and carcinoma development in a repeated dextran sulfate sodium-induced colitis model. J. Gastroenterol. Hepatol. 2002, 17, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett. 1995, 93, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P.; Good, C.K. The significance of aberrant crypt foci in understanding the pathogenesis of colon cancer. Toxicol. Lett. 2000, 112–113, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yoshimi, N.; Hirose, Y.; Kawabata, K.; Matsunaga, K.; Shimizu, M.; Hara, A.; Mori, H. Frequent beta-catenin gene mutations and accumulations of the protein in the putative preneoplastic lesions lacking macroscopic aberrant crypt foci appearance, in rat colon carcinogenesis. Cancer Res. 2000, 60, 3323–3327. [Google Scholar] [PubMed]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Invest. J. Tech. Methods Pathol. 1993, 69, 238–249. [Google Scholar]
- Marchal Bressenot, A.; Riddell, R.H.; Boulagnon-Rombi, C.; Reinisch, W.; Danese, S.; Schreiber, S.; Peyrin-Biroulet, L. Review article: The histological assessment of disease activity in ulcerative colitis. Aliment. Pharmacol. Ther. 2015, 42, 957–967. [Google Scholar] [CrossRef]
- Fukui, T.; Kishimoto, M.; Nakajima, A.; Yamashina, M.; Nakayama, S.; Kusuda, T.; Sakaguchi, Y.; Yoshida, K.; Uchida, K.; Nishio, A.; et al. The specific linker phosphorylation of smad2/3 indicates epithelial stem cells in stomach; particularly increasing in mucosae of helicobacter-associated gastritis. J. Gastroenterol. 2011, 46, 456–468. [Google Scholar] [CrossRef]
- Takahashi, Y.; Fukui, T.; Kishimoto, M.; Suzuki, R.; Mitsuyama, T.; Sumimoto, K.; Okazaki, T.; Sakao, M.; Sakaguchi, Y.; Yoshida, K.; et al. Phosphorylation of smad2/3 at the specific linker threonine residue indicates slow-cycling esophageal stem-like cells before re-entry to the cell cycle. Dis. Esoph. Off. J. Int. Soc. Dis. Esophagus 2016, 29, 107–115. [Google Scholar] [CrossRef]
- Espersen, M.L.; Olsen, J.; Linnemann, D.; Hogdall, E.; Troelsen, J.T. Clinical implications of intestinal stem cell markers in colorectal cancer. Clin. Colorect. Cancer 2015, 14, 63–71. [Google Scholar] [CrossRef]
- Srinivasan, T.; Walters, J.; Bu, P.; Than, E.B.; Tung, K.L.; Chen, K.Y.; Panarelli, N.; Milsom, J.; Augenlicht, L.; Lipkin, S.M.; et al. Notch signaling regulates asymmetric cell fate of fast- and slow-cycling colon cancer-initiating cells. Cancer Res. 2016, 76, 3411–3421. [Google Scholar] [CrossRef]
- Suzuki, R.; Kohno, H.; Sugie, S.; Tanaka, T. Sequential observations on the occurrence of preneoplastic and neoplastic lesions in mouse colon treated with azoxymethane and dextran sodium sulfate. Cancer Sci. 2004, 95, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M. Morphogenesis of chemically induced neoplasms of the colon and small intestine in rats. Lab. Investig. J. Tech. Methods Pathol. 1974, 30, 505–513. [Google Scholar]
- Murata, M.; Matsuzaki, K.; Yoshida, K.; Sekimoto, G.; Tahashi, Y.; Mori, S.; Uemura, Y.; Sakaida, N.; Fujisawa, J.; Seki, T.; et al. Hepatitis b virus x protein shifts human hepatic transforming growth factor (tgf)-beta signaling from tumor suppression to oncogenesis in early chronic hepatitis b. Hepatology 2009, 49, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Sekimoto, G.; Matsuzaki, K.; Yoshida, K.; Mori, S.; Murata, M.; Seki, T.; Matsui, H.; Fujisawa, J.; Okazaki, K. Reversible smad-dependent signaling between tumor suppression and oncogenesis. Cancer Res. 2007, 67, 5090–5096. [Google Scholar] [CrossRef]
- Furukawa, F.; Matsuzaki, K.; Mori, S.; Tahashi, Y.; Yoshida, K.; Sugano, Y.; Yamagata, H.; Matsushita, M.; Seki, T.; Inagaki, Y.; et al. P38 mapk mediates fibrogenic signal through smad3 phosphorylation in rat myofibroblasts. Hepatology 2003, 38, 879–889. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, Y.; Fukui, T.; Horitani, S.; Tanimura, Y.; Suzuki, R.; Tomiyama, T.; Honzawa, Y.; Tahara, T.; Okazaki, K.; Naganuma, M. A Short-Term Model of Colitis-Associated Colorectal Cancer That Suggests Initial Tumor Development and the Characteristics of Cancer Stem Cells. Int. J. Mol. Sci. 2023, 24, 11697. https://doi.org/10.3390/ijms241411697
Matsumoto Y, Fukui T, Horitani S, Tanimura Y, Suzuki R, Tomiyama T, Honzawa Y, Tahara T, Okazaki K, Naganuma M. A Short-Term Model of Colitis-Associated Colorectal Cancer That Suggests Initial Tumor Development and the Characteristics of Cancer Stem Cells. International Journal of Molecular Sciences. 2023; 24(14):11697. https://doi.org/10.3390/ijms241411697
Chicago/Turabian StyleMatsumoto, Yasushi, Toshiro Fukui, Shunsuke Horitani, Yuji Tanimura, Ryo Suzuki, Takashi Tomiyama, Yusuke Honzawa, Tomomitsu Tahara, Kazuichi Okazaki, and Makoto Naganuma. 2023. "A Short-Term Model of Colitis-Associated Colorectal Cancer That Suggests Initial Tumor Development and the Characteristics of Cancer Stem Cells" International Journal of Molecular Sciences 24, no. 14: 11697. https://doi.org/10.3390/ijms241411697
APA StyleMatsumoto, Y., Fukui, T., Horitani, S., Tanimura, Y., Suzuki, R., Tomiyama, T., Honzawa, Y., Tahara, T., Okazaki, K., & Naganuma, M. (2023). A Short-Term Model of Colitis-Associated Colorectal Cancer That Suggests Initial Tumor Development and the Characteristics of Cancer Stem Cells. International Journal of Molecular Sciences, 24(14), 11697. https://doi.org/10.3390/ijms241411697