
Mechanism of Androgen-Independent Stromal Proliferation in Benign Prostatic Hyperplasia

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Histological Features of BPH
3. Stromal-Epithelial Interaction in BPH
4. Stromal Proliferation in BPH
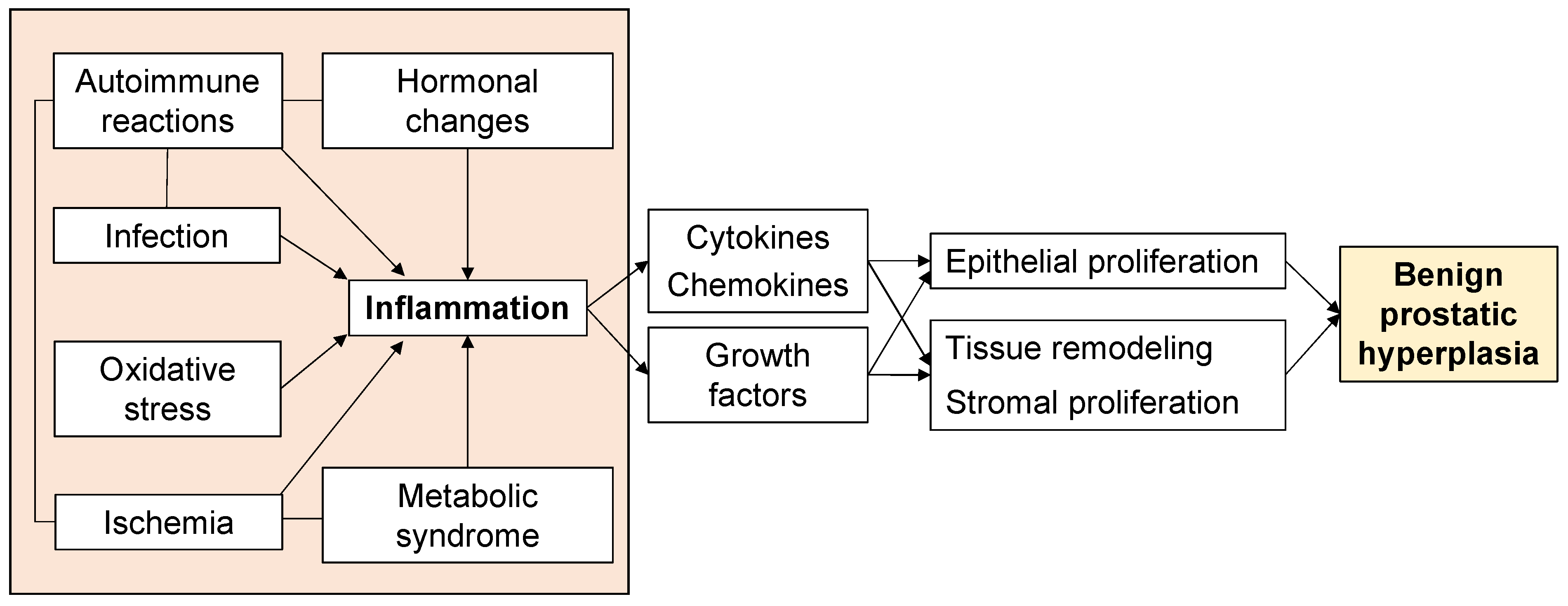
5. Inflammation in BPH Proliferation
6. Ischemia in BPH Proliferation
7. Oxidative Stress in BPH Proliferation
8. Metabolic Syndrome in BPH Proliferation
9. Infection in BPH Proliferation
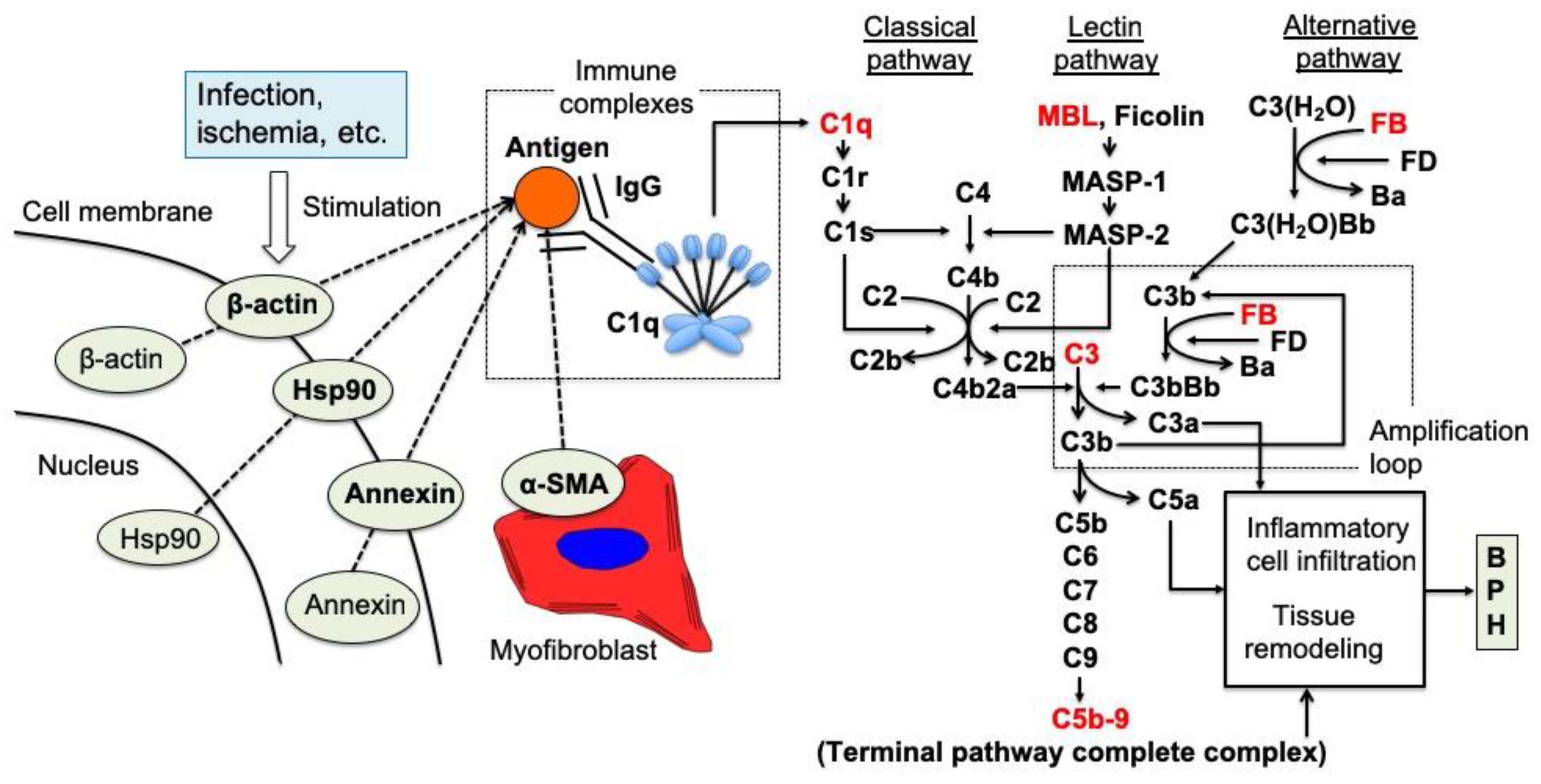
10. Autoimmune Reactions in BPH Proliferation
11. Myofibroblasts as Reactive Stroma in BPH Proliferation
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Djavan, B.; Margreiter, M.; Dianat, S.S. An algorithm for medical management in male lower urinary tract symptoms. Curr. Opin. Urol. 2011, 21, 5–12. [Google Scholar] [CrossRef] [PubMed]
- McNeal, J.E. Regional morphology and pathology of the prostate. Am. J. Clin. Pathol. 1968, 49, 347–357. [Google Scholar] [CrossRef] [PubMed]
- McNeal, J.E. The prostate and prostatic urethra: A morphologic synthesis. J. Urol. 1972, 107, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- McNeal, J.E. Origin and evolution of benign prostatic enlargement. Investig. Urol. 1978, 15, 340–345. [Google Scholar]
- McNeal, J.E. The zonal anatomy of the prostate. Prostate 1981, 2, 35–49. [Google Scholar] [CrossRef]
- McNeal, J.E. Anatomy of the prostate and morphogenesis of BPH. Prog. Clin. Biol. Res. 1984, 145, 27–53. [Google Scholar]
- Berry, S.J.; Coffey, D.S.; Walsh, P.C.; Ewing, L.L. The development of human benign prostatic hyperplasia with age. J. Urol. 1984, 132, 474–479. [Google Scholar] [CrossRef]
- Isaacs, J.T. Etiology of benign prostatic hyperplasia. Eur. Urol. 1994, 25 (Suppl. 1), 6–9. [Google Scholar] [CrossRef]
- McNeal, J. Pathology of benign prostatic hyperplasia. Insight into etiology. Urol. Clin. N. Am. 1990, 17, 477–486. [Google Scholar] [CrossRef]
- Isaacs, J.T. Prostate stem cells and benign prostatic hyperplasia. Prostate 2008, 68, 1025–1034. [Google Scholar] [CrossRef]
- Claus, S.; Wrenger, M.; Senge, T.; Schulze, H. Immunohistochemical determination of age related proliferation rates in normal and benign hyperplastic human prostates. Urol. Res. 1993, 21, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.; Thornhill, J.; Gaffney, E. The connective tissue framework in the normal prostate, BPH and prostate cancer: Analysis by scanning electron microscopy after cellular digestion. Urol. Res. 2000, 28, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Roehrborn, C.G. Benign prostatic hyperplasia: An overview. Rev. Urol. 2005, 7 (Suppl. 9), S3–S14. [Google Scholar]
- Tubaro, A.; De Nunzio, C.; Puccini, F.; Presicce, F. The evolving picture of lower urinary tract symptom management. Eur. Urol. 2015, 67, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Roehrborn, C.G.; Boyle, P.; Nickel, J.C.; Hoefner, K.; Andriole, G. Efficacy and safety of a dual inhibitor of 5-alpha-reductase types 1 and 2 (dutasteride) in men with benign prostatic hyperplasia. Urology 2002, 60, 434–441. [Google Scholar] [CrossRef]
- McConnell, J.D.; Roehrborn, C.G.; Bautista, O.M.; Andriole, G.L., Jr.; Dixon, C.M.; Kusek, J.W.; Lepor, H.; McVary, K.T.; Nyberg, L.M., Jr.; Clarke, H.S.; et al. The long-term effect of doxazosin, finasteride, and combination therapy on the clinical progression of benign prostatic hyperplasia. N. Engl. J. Med. 2003, 349, 2387–2398. [Google Scholar] [CrossRef]
- Price, H.; McNeal, J.E.; Stamey, T.A. Evolving patterns of tissue composition in benign prostatic hyperplasia as a function of specimen size. Hum. Pathol. 1990, 21, 578–585. [Google Scholar] [CrossRef]
- Franks, L.M. Benign nodular hyperplasia of the prostate; a review. Ann. R. Coll. Surg. Engl. 1953, 14, 92–106. [Google Scholar]
- Tong, Y.; Zhou, R.Y. Review of the Roles and Interaction of Androgen and Inflammation in Benign Prostatic Hyperplasia. Mediat. Inflamm. 2020, 2020, 7958316. [Google Scholar] [CrossRef]
- Lee, K.L.; Peehl, D.M. Molecular and cellular pathogenesis of benign prostatic hyperplasia. J. Urol. 2004, 172, 1784–1791. [Google Scholar] [CrossRef]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20 (Suppl. 3), S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R. Epithelio-mesenchymal interactions in primordial gland structures which become responsive to androgenic stimulation. Anat. Rec. 1972, 172, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Cooke, P.S.; Kurita, T. Role of stromal-epithelial interactions in hormonal responses. Arch. Histol. Cytol. 2004, 67, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Lung, B. The possible influence of temporal factors in androgenic responsiveness of urogenital tissue recombinants from wild-type and androgen-insensitive (Tfm) mice. J. Exp. Zool. 1978, 205, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, N.A.; Chytil, A.; Plieth, D.; Gorska, A.E.; Dumont, N.; Shappell, S.; Washington, M.K.; Neilson, E.G.; Moses, H.L. TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science 2004, 303, 848–851. [Google Scholar] [CrossRef]
- Franco, O.E.; Jiang, M.; Strand, D.W.; Peacock, J.; Fernandez, S.; Jackson, R.S., 2nd; Revelo, M.P.; Bhowmick, N.A.; Hayward, S.W. Altered TGF-β signaling in a subpopulation of human stromal cells promotes prostatic carcinogenesis. Cancer. Res. 2011, 71, 1272–1281. [Google Scholar] [CrossRef]
- Memarzadeh, S.; Xin, L.; Mulholland, D.J.; Mansukhani, A.; Wu, H.; Teitell, M.A.; Witte, O.N. Enhanced paracrine FGF10 expression promotes formation of multifocal prostate adenocarcinoma and an increase in epithelial androgen receptor. Cancer Cell. 2007, 12, 572–585. [Google Scholar] [CrossRef]
- Cunha, G.R.; Hayward, S.W.; Dahiya, R.; Foster, B.A. Smooth muscle-epithelial interactions in normal and neoplastic prostatic development. Acta. Anat. 1996, 155, 63–72. [Google Scholar] [CrossRef]
- Hayward, S.W.; Haughney, P.C.; Rosen, M.A.; Greulich, K.M.; Weier, H.U.; Dahiya, R.; Cunha, G.R. Interactions between adult human prostatic epithelium and rat urogenital sinus mesenchyme in a tissue recombination model. Differentiation 1998, 63, 131–140. [Google Scholar] [CrossRef]
- Hayward, S.W.; Baskin, L.S.; Haughney, P.C.; Foster, B.A.; Cunha, A.R.; Dahiya, R.; Prins, G.S.; Cunha, G.R. Stromal development in the ventral prostate, anterior prostate and seminal vesicle of the rat. Acta. Anat. 1996, 155, 94–103. [Google Scholar] [CrossRef]
- Cunha, G.R.; Battle, E.; Young, P.; Brody, J.; Donjacour, A.; Hayashi, N.; Kinbara, H. Role of epithelial-mesenchymal interactions in the differentiation and spatial organization of visceral smooth muscle. Epithel. Cell. Biol. 1992, 1, 76–83. [Google Scholar]
- Liu, D.; Liu, J.; Li, Y.; Liu, H.; Hassan, H.M.; He, W.; Li, M.; Zhou, Y.; Fu, X.; Zhan, J.; et al. Upregulated bone morphogenetic protein 5 enhances proliferation and epithelial-mesenchymal transition process in benign prostatic hyperplasia via BMP/Smad signaling pathway. Prostate 2021, 81, 1435–1449. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.C.; Joyner, A.L. Hedgehog signaling in prostate epithelial-mesenchymal growth regulation. Dev. Biol. 2015, 400, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.; Bushman, W. Hedgehog signaling in the prostate. J. Urol. 2007, 177, 832–838. [Google Scholar] [CrossRef]
- Vezina, C.M.; Bushman, A.W. Hedgehog signaling in prostate growth and benign prostate hyperplasia. Curr. Urol. Rep. 2007, 8, 275–280. [Google Scholar] [CrossRef]
- Gonnissen, A.; Isebaert, S.; Haustermans, K. Hedgehog signaling in prostate cancer and its therapeutic implication. Int. J. Mol. Sci. 2013, 14, 13979–14007. [Google Scholar] [CrossRef]
- Wang, X.; Lin, W.J.; Izumi, K.; Jiang, Q.; Lai, K.P.; Xu, D.; Fang, L.Y.; Lu, T.; Li, L.; Xia, S.; et al. Increased infiltrated macrophages in benign prostatic hyperplasia (BPH): Role of stromal androgen receptor in macrophage-induced prostate stromal cell proliferation. J. Biol. Chem. 2012, 287, 18376–18385. [Google Scholar] [CrossRef]
- Nicholson, T.M.; Ricke, W.A. Androgens and estrogens in benign prostatic hyperplasia: Past, present and future. Differentiation 2011, 82, 184–199. [Google Scholar] [CrossRef]
- Vignozzi, L.; Rastrelli, G.; Corona, G.; Gacci, M.; Forti, G.; Maggi, M. Benign prostatic hyperplasia: A new metabolic disease? J. Endocrinol. Investig. 2014, 37, 313–322. [Google Scholar] [CrossRef]
- Madersbacher, S.; Sampson, N.; Culig, Z. Pathophysiology of Benign Prostatic Hyperplasia and Benign Prostatic Enlargement: A Mini-Review. Gerontology 2019, 65, 458–464. [Google Scholar] [CrossRef]
- Kamijo, T.; Sato, S.; Kitamura, T. Effect of cernitin pollen-extract on experimental nonbacterial prostatitis in rats. Prostate 2001, 49, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Mori, F.; Oda, N.; Sakuragi, M.; Sakakibara, F.; Kiniwa, M.; Miyoshi, K. New histopathological experimental model for benign prostatic hyperplasia: Stromal hyperplasia in rats. J. Urol. 2009, 181, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Tanji, R.; Onagi, A.; Honda-Takinami, R.; Matsuoka, K.; Hoshi, S.; Sato, Y.; Akaihata, H.; Haga, N.; Kojima, Y. Morphological change and characteristics of myofibroblasts during the growth process of benign prostatic hyperplasia. Int. J. Urol. 2020, 27, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Satoh, Y.; Akaihata, H.; Hiraki, H.; Ogawa, S.; Haga, N.; Ishibashi, K.; Aikawa, K.; Kojima, Y. Molecular classification of benign prostatic hyperplasia: A gene expression profiling study in a rat model. Int. J. Urol. 2016, 23, 599–612. [Google Scholar] [CrossRef]
- Kramer, G.; Marberger, M. Could inflammation be a key component in the progression of benign prostatic hyperplasia? Curr. Opin. Urol. 2006, 16, 25–29. [Google Scholar] [CrossRef]
- Kramer, G.; Mitteregger, D.; Marberger, M. Is benign prostatic hyperplasia (BPH) an immune inflammatory disease? Eur. Urol. 2007, 51, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Nickel, J.C. Inflammation and benign prostatic hyperplasia. Urol. Clin. N. Am. 2008, 35, 109–115. [Google Scholar] [CrossRef]
- Sciarra, A.; Mariotti, G.; Salciccia, S.; Autran Gomez, A.; Monti, S.; Toscano, V.; Di Silverio, F. Prostate growth and inflammation. J. Steroid. Biochem. Mol. Biol. 2008, 108, 254–260. [Google Scholar] [CrossRef]
- Untergasser, G.; Madersbacher, S.; Berger, P. Benign prostatic hyperplasia: Age-related tissue-remodeling. Exp. Gerontol. 2005, 40, 121–128. [Google Scholar] [CrossRef]
- Donnell, R.F. Benign prostate hyperplasia: A review of the year’s progress from bench to clinic. Curr. Opin. Urol. 2011, 21, 22–26. [Google Scholar] [CrossRef]
- Emberton, M.; Fitzpatrick, J.M.; Garcia-Losa, M.; Qizilbash, N.; Djavan, B. Progression of benign prostatic hyperplasia: Systematic review of the placebo arms of clinical trials. BJU Int. 2008, 102, 981–986. [Google Scholar] [CrossRef] [PubMed]
- De Nunzio, C.; Presicce, F.; Tubaro, A. Inflammatory mediators in the development and progression of benign prostatic hyperplasia. Nat. Rev. Urol. 2016, 13, 613–626. [Google Scholar] [CrossRef]
- Nickel, J.C.; Roehrborn, C.G.; O’Leary, M.P.; Bostwick, D.G.; Somerville, M.C.; Rittmaster, R.S. The relationship between prostate inflammation and lower urinary tract symptoms: Examination of baseline data from the REDUCE trial. Eur. Urol. 2008, 54, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Robert, G.; Descazeaud, A.; Nicolaïew, N.; Terry, S.; Sirab, N.; Vacherot, F.; Maillé, P.; Allory, Y.; de la Taille, A. Inflammation in benign prostatic hyperplasia: A 282 patients’ immunohistochemical analysis. Prostate 2009, 69, 1774–1780. [Google Scholar] [CrossRef]
- Pyun, J.H.; Kang, S.G.; Kang, S.H.; Cheon, J.; Kim, J.J.; Lee, J.G. Efficacy of holmium laser enucleation of the prostate (HoLEP) in men with bladder outlet obstruction (BOO) and non-neurogenic bladder dysfunction. Kaohsiung J. Med. Sci. 2017, 33, 458–463. [Google Scholar] [CrossRef]
- De Nunzio, C.; Kramer, G.; Marberger, M.; Montironi, R.; Nelson, W.; Schröder, F.; Sciarra, A.; Tubaro, A. The controversial relationship between benign prostatic hyperplasia and prostate cancer: The role of inflammation. Eur. Urol. 2011, 60, 106–117. [Google Scholar] [CrossRef]
- Palapattu, G.S.; Sutcliffe, S.; Bastian, P.J.; Platz, E.A.; De Marzo, A.M.; Isaacs, W.B.; Nelson, W.G. Prostate carcinogenesis and inflammation: Emerging insights. Carcinogenesis 2005, 26, 1170–1181. [Google Scholar] [CrossRef]
- Macoska, J.A. Chemokines and BPH/LUTS. Differentiation 2011, 82, 253–260. [Google Scholar] [CrossRef]
- McLaren, I.D.; Jerde, T.J.; Bushman, W. Role of interleukins, IGF and stem cells in BPH. Differentiation 2011, 82, 237–243. [Google Scholar] [CrossRef]
- Quillard, T.; Charreau, B. Impact of notch signaling on inflammatory responses in cardiovascular disorders. Int. J. Mol. Sci. 2013, 14, 6863–6888. [Google Scholar] [CrossRef]
- Hahn, A.M.; Myers, J.D.; McFarland, E.K.; Lee, S.; Jerde, T.J. Interleukin-driven insulin-like growth factor promotes prostatic inflammatory hyperplasia. J. Pharm. Exp. 2014, 351, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Edeling, M.; Ragi, G.; Huang, S.; Pavenstädt, H.; Susztak, K. Developmental signalling pathways in renal fibrosis: The roles of Notch, Wnt and Hedgehog. Nat. Rev. Nephrol. 2016, 12, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Fazio, C.; Ricciardiello, L. Inflammation and Notch signaling: A crosstalk with opposite effects on tumorigenesis. Cell. Death Dis. 2016, 7, e2515. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Smith, S.; Hu, X. Role of Notch signaling in regulating innate immunity and inflammation in health and disease. Protein Cell. 2016, 7, 159–174. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Madonna, R.; Melino, G.; Caruso, C. The emerging role of Notch pathway in ageing: Focus on the related mechanisms in age-related diseases. Ageing Res. Rev. 2016, 29, 50–65. [Google Scholar] [CrossRef]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef]
- Kohnen, P.W.; Drach, G.W. Patterns of inflammation in prostatic hyperplasia: A histologic and bacteriologic study. J. Urol. 1979, 121, 755–760. [Google Scholar] [CrossRef]
- Steiner, G.E.; Stix, U.; Handisurya, A.; Willheim, M.; Haitel, A.; Reithmayr, F.; Paikl, D.; Ecker, R.C.; Hrachowitz, K.; Kramer, G.; et al. Cytokine expression pattern in benign prostatic hyperplasia infiltrating T cells and impact of lymphocytic infiltration on cytokine mRNA profile in prostatic tissue. Lab. Investig. 2003, 83, 1131–1146. [Google Scholar] [CrossRef]
- Chughtai, B.; Lee, R.; Te, A.; Kaplan, S. Role of inflammation in benign prostatic hyperplasia. Rev. Urol. 2011, 13, 147–150. [Google Scholar]
- Morelli, A.; Sarchielli, E.; Comeglio, P.; Filippi, S.; Mancina, R.; Gacci, M.; Vignozzi, L.; Carini, M.; Vannelli, G.B.; Maggi, M. Phosphodiesterase type 5 expression in human and rat lower urinary tract tissues and the effect of tadalafil on prostate gland oxygenation in spontaneously hypertensive rats. J. Sex. Med. 2011, 8, 2746–2760. [Google Scholar] [CrossRef]
- Vignozzi, L.; Gacci, M.; Cellai, I.; Morelli, A.; Maneschi, E.; Comeglio, P.; Santi, R.; Filippi, S.; Sebastianelli, A.; Nesi, G.; et al. PDE5 inhibitors blunt inflammation in human BPH: A potential mechanism of action for PDE5 inhibitors in LUTS. Prostate 2013, 73, 1391–1402. [Google Scholar] [CrossRef]
- Saito, M.; Tsounapi, P.; Oikawa, R.; Shimizu, S.; Honda, M.; Sejima, T.; Kinoshita, Y.; Tomita, S. Prostatic ischemia induces ventral prostatic hyperplasia in the SHR; possible mechanism of development of BPH. Sci. Rep. 2014, 4, 3822. [Google Scholar] [CrossRef] [PubMed]
- Haga, N.; Akaihata, H.; Hata, J.; Hiraki, H.; Honda, R.; Tanji, R.; Onagi, A.; Koguchi, T.; Hoshi, S.; Ogawa, S.; et al. The association between local arteriosclerosis of the prostatic arteries and chronic inflammation in human benign prostatic enlargement. Prostate 2019, 79, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Yamashita, S.; Hayashi, N.; Goto, T.; Koyama, J.; Sato, T.; Shimada, S.; Kawasaki, Y.; Izumi, H.; Kawamorita, N.; et al. Phosphodiesterase type 5 inhibitor attenuates chronic ischemia-induced prostatic hyperplasia in a rat model. Prostate 2019, 79, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef]
- Vital, P.; Castro, P.; Ittmann, M. Oxidative stress promotes benign prostatic hyperplasia. Prostate 2016, 76, 58–67. [Google Scholar] [CrossRef]
- Minciullo, P.L.; Inferrera, A.; Navarra, M.; Calapai, G.; Magno, C.; Gangemi, S. Oxidative stress in benign prostatic hyperplasia: A systematic review. Urol. Int. 2015, 94, 249–254. [Google Scholar] [CrossRef]
- Ahmad, M.; Suhail, N.; Mansoor, T.; Banu, N.; Ahmad, S. Evaluation of oxidative stress and DNA damage in benign prostatic hyperplasia patients and comparison with controls. Indian J. Clin. Biochem. 2012, 27, 385–388. [Google Scholar] [CrossRef]
- Aryal, M.; Pandeya, A.; Gautam, N.; Baral, N.; Lamsal, M.; Majhi, S.; Chandra, L.; Pandit, R.; Das, B.K. Oxidative stress in benign prostate hyperplasia. Nepal Med. Coll. J. 2007, 9, 222–224. [Google Scholar]
- Kullisaar, T.; Türk, S.; Punab, M.; Mändar, R. Oxidative stress--cause or consequence of male genital tract disorders? Prostate 2012, 72, 977–983. [Google Scholar] [CrossRef] [PubMed]
- De Nunzio, C.; Aronson, W.; Freedland, S.J.; Giovannucci, E.; Parsons, J.K. The correlation between metabolic syndrome and prostatic diseases. Eur. Urol. 2012, 61, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.K.; Carter, H.B.; Partin, A.W.; Windham, B.G.; Metter, E.J.; Ferrucci, L.; Landis, P.; Platz, E.A. Metabolic factors associated with benign prostatic hyperplasia. J. Clin. Endocrinol. Metab. 2006, 91, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Draznin, B. Mitogenic action of insulin: Friend, foe or ‘frenemy’? Diabetologia 2010, 53, 229–233. [Google Scholar] [CrossRef]
- McKeehan, W.L.; Adams, P.S.; Rosser, M.P. Direct mitogenic effects of insulin, epidermal growth factor, glucocorticoid, cholera toxin, unknown pituitary factors and possibly prolactin, but not androgen, on normal rat prostate epithelial cells in serum-free, primary cell culture. Cancer. Res. 1984, 44, 1998–2010. [Google Scholar]
- Cannarella, R.; Condorelli, R.A.; Barbagallo, F.; La Vignera, S.; Calogero, A.E. Endocrinology of the Aging Prostate: Current Concepts. Front. Endocrinol. 2021, 12, 554078. [Google Scholar] [CrossRef]
- Du, Y.; Long, Q.; Shi, Y.; Liu, X.; Li, X.; Zeng, J.; Gong, Y.; Li, L.; Wang, X.; He, D. Insulin-like growth factor binding protein-3 mediates interleukin-24-induced apoptosis through inhibition of the mTOR pathway in prostate cancer. Oncol. Rep. 2015, 34, 2273–2281. [Google Scholar] [CrossRef]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells. 2020, 9, 1261. [Google Scholar] [CrossRef]
- Chen, I.H.; Tong, Y.C.; Cheng, J.T. Metabolic syndrome decreases tissue perfusion and induces glandular hyperplasia in the fructose-fed rat prostate. Neurourol. Urodyn. 2012, 31, 600–604. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, S.; Li, Y.; Zhou, Y.; Liu, D.; Liu, J.; DiSanto, M.E.; Zhang, X. Simvastatin Improves Benign Prostatic Hyperplasia: Role of Peroxisome-Proliferator-Activated Receptor-γ and Classic WNT/β-Catenin Pathway. Int. J. Mol. Sci. 2023, 24, 4911. [Google Scholar] [CrossRef]
- Sutcliffe, S.; Giovannucci, E.; Alderete, J.F.; Chang, T.H.; Gaydos, C.A.; Zenilman, J.M.; De Marzo, A.M.; Willett, W.C.; Platz, E.A. Plasma antibodies against Trichomonas vaginalis and subsequent risk of prostate cancer. Cancer. Epidemiol. Biomark. Prev. 2006, 15, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Urisman, A.; Molinaro, R.J.; Fischer, N.; Plummer, S.J.; Casey, G.; Klein, E.A.; Malathi, K.; Magi-Galluzzi, C.; Tubbs, R.R.; Ganem, D.; et al. Identification of a novel Gammaretrovirus in prostate tumors of patients homozygous for R462Q RNASEL variant. PLoS Pathog. 2006, 2, e25. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Machida, T.; Matsuoka, K.; Hoshi, S.; Akaihata, H.; Hiraki, H.; Suzuki, T.; Ogawa, S.; Kataoka, M.; Haga, N.; et al. Complement activation by autoantigen recognition in the growth process of benign prostatic hyperplasia. Sci. Rep. 2019, 9, 20357. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, S.S.; Han, I.H.; Sim, S.; Ahn, M.H.; Ryu, J.S. Proliferation of Prostate Stromal Cell Induced by Benign Prostatic Hyperplasia Epithelial Cell Stimulated With Trichomonas vaginalis via Crosstalk With Mast Cell. Prostate 2016, 76, 1431–1444. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Han, I.H.; Kim, Y.S.; Noh, C.S.; Ryu, J.S. Proliferation of prostate epithelia induced by IL-6 from stroma reacted with Trichomonas vaginalis. Parasite Immunol. 2018, 40, e12531. [Google Scholar] [CrossRef]
- Porter, C.M.; Shrestha, E.; Peiffer, L.B.; Sfanos, K.S. The microbiome in prostate inflammation and prostate cancer. Prostate Cancer Prostatic Dis. 2018, 21, 345–354. [Google Scholar] [CrossRef]
- Takezawa, K.; Fujita, K.; Matsushita, M.; Motooka, D.; Hatano, K.; Banno, E.; Shimizu, N.; Takao, T.; Takada, S.; Okada, K.; et al. The Firmicutes/Bacteroidetes ratio of the human gut microbiota is associated with prostate enlargement. Prostate 2021, 81, 1287–1293. [Google Scholar] [CrossRef]
- Fang, C.; Wu, L.; Zhu, C.; Xie, W.Z.; Hu, H.; Zeng, X.T. A potential therapeutic strategy for prostatic disease by targeting the oral microbiome. Med. Res. Rev. 2021, 41, 1812–1834. [Google Scholar] [CrossRef]
- Estemalik, J.; Demko, C.; Bissada, N.F.; Joshi, N.; Bodner, D.; Shankar, E.; Gupta, S. Simultaneous Detection of Oral Pathogens in Subgingival Plaque and Prostatic Fluid of Men With Periodontal and Prostatic Diseases. J. Periodontol. 2017, 88, 823–829. [Google Scholar] [CrossRef]
- Honda-Takinami, R.; Hata, J.; Matsuoka, K.; Hoshi, S.; Koguchi, T.; Sato, Y.; Akaihata, H.; Kataoka, M.; Ogawa, S.; Nishiyama, K.; et al. Association between the presence of bacteria in prostate tissue and histopathology in biopsies from men not complaining of lower urinary tract symptoms. Fukushima J. Med. Sci. 2022, 68, 161–167. [Google Scholar] [CrossRef]
- Okada, K.; Takezawa, K.; Tsujimura, G.; Imanaka, T.; Kuribayashi, S.; Ueda, N.; Hatano, K.; Fukuhara, S.; Kiuchi, H.; Fujita, K.; et al. Localization and potential role of prostate microbiota. Front. Cell. Infect. Microbiol. 2022, 12, 1048319. [Google Scholar] [CrossRef] [PubMed]
- Magri, V.; Boltri, M.; Cai, T.; Colombo, R.; Cuzzocrea, S.; De Visschere, P.; Giuberti, R.; Granatieri, C.M.; Latino, M.A.; Larganà, G.; et al. Multidisciplinary approach to prostatitis. Arch. Ital. Urol. 2019, 90, 227–248. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.B.; Macejko, A.; Taylor, A.; Nadler, R.B. Chronic prostatitis: Management strategies. Drugs 2009, 69, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Fibbi, B.; Amuchastegui, S.; Cossetti, C.; Aquilano, F.; Laverny, G.; Gacci, M.; Crescioli, C.; Maggi, M.; Adorini, L. Human benign prostatic hyperplasia stromal cells as inducers and targets of chronic immuno-mediated inflammation. J. Immunol. 2009, 182, 4056–4064. [Google Scholar] [CrossRef]
- Taguchi, O.; Nishizuka, Y. Self tolerance and localized autoimmunity. Mouse models of autoimmune disease that suggest tissue-specific suppressor T cells are involved in self tolerance. J. Exp. Med. 1987, 165, 146–156. [Google Scholar] [CrossRef]
- Ablin, R.J.; Gonder, M.J.; Soanes, W.A. Localization of immunoglobulins in human prostatic tissue. J. Immunol. 1971, 107, 603–604. [Google Scholar] [CrossRef]
- Klyushnenkova, E.N.; Ponniah, S.; Rodriguez, A.; Kodak, J.; Mann, D.L.; Langerman, A.; Nishimura, M.I.; Alexander, R.B. CD4 and CD8 T-lymphocyte recognition of prostate specific antigen in granulomatous prostatitis. J. Immunother. 2004, 27, 136–146. [Google Scholar] [CrossRef]
- Klyushnenkova, E.N.; Link, J.; Oberle, W.T.; Kodak, J.; Rich, C.; Vandenbark, A.A.; Alexander, R.B. Identification of HLA-DRB1*1501-restricted T-cell epitopes from prostate-specific antigen. Clin. Cancer Res. 2005, 11, 2853–2861. [Google Scholar] [CrossRef]
- Liedtke, V.; Rose, L.; Hiemann, R.; Nasser, A.; Rödiger, S.; Bonaventura, A.; Winkler, L.; Sowa, M.; Stöckle, M.; Schierack, P.; et al. Over-Expression of LEDGF/p75 in HEp-2 Cells Enhances Autoimmune IgG Response in Patients with Benign Prostatic Hyperplasia-A Novel Diagnostic Approach with Therapeutic Consequence? Int. J. Mol. Sci. 2023, 24, 6166. [Google Scholar] [CrossRef]
- Roumenina, L.T.; Daugan, M.V.; Noé, R.; Petitprez, F.; Vano, Y.A.; Sanchez-Salas, R.; Becht, E.; Meilleroux, J.; Clec’h, B.L.; Giraldo, N.A.; et al. Tumor Cells Hijack Macrophage-Produced Complement C1q to Promote Tumor Growth. Cancer. Immunol. Res. 2019, 7, 1091–1105. [Google Scholar] [CrossRef]
- Vickman, R.E.; Aaron-Brooks, L.; Zhang, R.; Lanman, N.A.; Lapin, B.; Gil, V.; Greenberg, M.; Sasaki, T.; Cresswell, G.M.; Broman, M.M.; et al. TNF is a potential therapeutic target to suppress prostatic inflammation and hyperplasia in autoimmune disease. Nat. Commun. 2022, 13, 2133. [Google Scholar] [CrossRef] [PubMed]
- Sampson, N.; Berger, P.; Zenzmaier, C. Therapeutic targeting of redox signaling in myofibroblast differentiation and age-related fibrotic disease. Oxid. Med. Cell. Longev. 2012, 2012, 458276. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.W.; Mifflin, R.C.; Valentich, J.D.; Crowe, S.E.; Saada, J.I.; West, A.B. Myofibroblasts. I. Paracrine cells important in health and disease. Am. J. Physiol. 1999, 277, C1–C9. [Google Scholar] [CrossRef]
- Tuxhorn, J.A.; Ayala, G.E.; Smith, M.J.; Smith, V.C.; Dang, T.D.; Rowley, D.R. Reactive stroma in human prostate cancer: Induction of myofibroblast phenotype and extracellular matrix remodeling. Clin. Cancer. Res. 2002, 8, 2912–2923. [Google Scholar] [PubMed]
- Rowley, D.R. What might a stromal response mean to prostate cancer progression? Cancer. Metastasis Rev. 1998, 17, 411–419. [Google Scholar] [CrossRef]
- Schauer, I.G.; Ressler, S.J.; Tuxhorn, J.A.; Dang, T.D.; Rowley, D.R. Elevated epithelial expression of interleukin-8 correlates with myofibroblast reactive stroma in benign prostatic hyperplasia. Urology 2008, 72, 205–213. [Google Scholar] [CrossRef]
- Schauer, I.G.; Ressler, S.J.; Rowley, D.R. Keratinocyte-derived chemokine induces prostate epithelial hyperplasia and reactive stroma in a novel transgenic mouse model. Prostate 2009, 69, 373–384. [Google Scholar] [CrossRef]
- Zenzmaier, C.; Sampson, N.; Plas, E.; Berger, P. Dickkopf-related protein 3 promotes pathogenic stromal remodeling in benign prostatic hyperplasia and prostate cancer. Prostate 2013, 73, 1441–1452. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hata, J.; Harigane, Y.; Matsuoka, K.; Akaihata, H.; Yaginuma, K.; Meguro, S.; Hoshi, S.; Sato, Y.; Ogawa, S.; Uemura, M.; et al. Mechanism of Androgen-Independent Stromal Proliferation in Benign Prostatic Hyperplasia. Int. J. Mol. Sci. 2023, 24, 11634. https://doi.org/10.3390/ijms241411634
Hata J, Harigane Y, Matsuoka K, Akaihata H, Yaginuma K, Meguro S, Hoshi S, Sato Y, Ogawa S, Uemura M, et al. Mechanism of Androgen-Independent Stromal Proliferation in Benign Prostatic Hyperplasia. International Journal of Molecular Sciences. 2023; 24(14):11634. https://doi.org/10.3390/ijms241411634
Chicago/Turabian StyleHata, Junya, Yuki Harigane, Kanako Matsuoka, Hidenori Akaihata, Kei Yaginuma, Satoru Meguro, Seiji Hoshi, Yuichi Sato, Soichiro Ogawa, Motohide Uemura, and et al. 2023. "Mechanism of Androgen-Independent Stromal Proliferation in Benign Prostatic Hyperplasia" International Journal of Molecular Sciences 24, no. 14: 11634. https://doi.org/10.3390/ijms241411634
APA StyleHata, J., Harigane, Y., Matsuoka, K., Akaihata, H., Yaginuma, K., Meguro, S., Hoshi, S., Sato, Y., Ogawa, S., Uemura, M., & Kojima, Y. (2023). Mechanism of Androgen-Independent Stromal Proliferation in Benign Prostatic Hyperplasia. International Journal of Molecular Sciences, 24(14), 11634. https://doi.org/10.3390/ijms241411634