Abstract
All the cells of an organism contain the same genome. However, each cell expresses only a minor fraction of its potential and, in particular, the genes encoding the proteins necessary for basal metabolism and the proteins responsible for its specific phenotype. The ability to use only the right and necessary genes involved in specific functions depends on the structural organization of the nuclear chromatin, which in turn depends on the epigenetic history of each cell, which is stored in the form of a collection of DNA and protein modifications. Among these modifications, DNA methylation and many kinds of post-translational modifications of histones play a key role in organizing the complex indexing of usable genes. In addition, non-canonical histone proteins (also known as histone variants), the synthesis of which is not directly linked with DNA replication, are used to mark specific regions of the genome. Here, we will discuss the role of the H3.3 histone variant, with particular attention to its loading into chromatin in the mammalian nervous system, both in physiological and pathological conditions. Indeed, chromatin modifications that mark cell memory seem to be of special importance for the cells involved in the complex processes of learning and memory.
1. Introduction
Since Gurdon’s experiments [1], based on the transplantation into enucleated oocytes of nuclei purified from somatic cells, it became clear that during differentiation, cells do not lose DNA and thus maintain a nucleus with intact potential to generate an entire organism. Despite the presence of the entire genome, however, each specialized cell expresses only a very small percentage of its genome. The reason for specific gene selection lies in the tridimensional organization of chromatin, which is a complex of DNA and proteins. Among these latter molecules, the most represented are histones, i.e., basic proteins that are highly conserved in evolution, which interact with DNA, allowing condensation of the nuclear genome in the very small volume of eukaryotic nuclei. The first level of chromatin organization is the nucleosome, in which about 147 base pairs (bp) of DNA are wrapped around a protein octamer formed by two molecules of each of the histones H2A, H2B, H3, and H4 (called “core” histones”). A fifth histone, the “linker” H1 histone then seals together the points at which DNA enters and exits the nucleosome. Interaction among H1 molecules allows the formation of more condensed DNA fibers [2,3,4,5,6,7,8].
Most importantly, the chromatin structure differs at the level of different genes that, depending on their structural organization, can be transcriptionally repressed or active, and this is why different cells are able to express different and specific genes, as well as a family of common genes involved in basic metabolism. Thus, specific properties and the behavior of somatic cells do not depend on changes in the genotype but on the specific arrangement of chromatin, according to the concept of “epigenetics” proposed by Waddington many decades ago [9], which is now widely accepted [4,10].
The chromatin structure is highly dynamic and changes in different cells during development and differentiation; it also changes in terminally differentiated cells in response to specific inducing factors, such as, for example, thyroid or steroid hormones [11]. At least three connected mechanisms are known to induce structural rearrangements of chromatin (Figure 1): (i) post-translational modification (PTM) of histone proteins [10,12] together with DNA methylation [10,13,14]; (ii) the activity of ATP-dependent complexes that are able to induce modifications in the structure/position of nucleosomes [15], and, finally, (iii) the synthesis and incorporation of histone variants into chromatin [7,16,17,18].
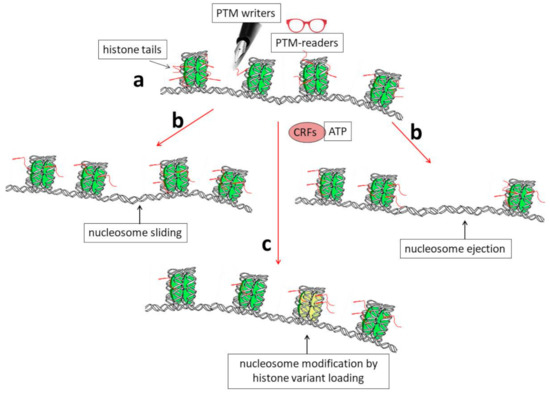
Figure 1.
Schematic representation showing the biochemical mechanisms that allow dynamic modifications of chromatin organization. (a) Histone tails protrude from nucleosomes (red segments) and can be the targets of enzymes (PTM writers) that are able to introduce into them post-translational modifications (PTMs). These modifications can, in turn, directly affect DNA–histone interactions and/or allow chromatin structural modifications through the binding of other specific factors (PTM readers). (b) A modification of the nucleosome position by sliding or nucleosome ejection can be catalyzed by ATP-dependent chromatin-remodeling factors (CRFs). (c) Finally, the incorporation of histone variants into chromatin, such as H3.3, can induce structural and functional modifications of nucleosomes (shown as the yellow nucleosome core particle in the figure).
Herein, we will mainly focus on the latter mechanism (Figure 1, pathway c) and, in particular, on the specific involvement of the H3 histone variant, known as H3.3 core histone, in the epigenetic regulation of gene expression in the nervous system during maturation, as well as in the acquisition of complex functions such as learning and memory. We also discuss the involvement of H3.3 in brain cancer.
2. General Properties of Genes Encoding Histone Variants
Although highly conserved in evolution, and thus among different species, histone proteins exist under different isoforms in the same species. These isoforms seem to have different effects on chromatin structure and then on gene expression.
Interestingly, genes encoding the main histone species differ both in structure and expression from those encoding variants synthesized only in specific moments of cell differentiation. In particular, the main histone species (also known as replication-dependent or canonical histones) are synthesized exclusively during the S phase of the cell cycle when DNA is replicated. The corresponding genes are highly repeated and very often clustered; moreover, they do not contain introns, and the corresponding mRNAs are not poly-adenylated [19,20,21]. All these features are related to the necessity of having mRNAs that are immediately available for translation and then for degradation. On the other hand, genes encoding constitutive/variant histones (also known as replication-independent or non-canonical histones) are similar to all the other genes in that they are mostly unique genes that are transcribed, independently of DNA replication, into mRNAs that can contain introns and are polyadenylated [22]. These genes should be regulated during differentiation in order to produce proteins that are able to bind specific regions of chromatin, thus allowing activation/repression of specific genes.
Many years ago, our group showed that two histone variants, i.e., the linker histone H1.0 and the core histone H3.3, are specifically expressed in the rat brain during brain maturation [23]. Interestingly, a combination of run-on experiments on isolated nuclei and transcription inhibition using actinomycin D demonstrated that the two genes have an “open” structure and that H1.0 and H3.3 histone synthesis in the central nervous system (CNS) is largely regulated at the post-transcriptional level [24]. Indeed, we identified a group of proteins that are able to bind their mRNAs and cloned a couple of them (CSD-C2/PIPPin and LPI/PEP-19) [25,26,27,28,29]. Notably, in a very recent and interesting paper focused on the effects of hunger on neuronal histone modifications and the life span of the Drosophila fruit fly, the effects of a diet containing low amounts of branched-chain amino acids (BCAAs) were analyzed. In particular, the authors found that total histone H3 abundance decreased in flies fed a low BCAA diet, while H3 mRNA increased [30]. This observation suggested that post-transcriptional events might be of importance for general H3 metabolism. Moreover, the paper also reported that canonical H3 is evicted from chromatin and replaced with H3.3 [30].
It is also worth noting that two genes encoding the H3.3 histone are present in mammals: H3.3A and H3.3B (also called H3F3A and H3F3B), which are located on different chromosomes. While the distribution of exons and introns, as well as promoters and other regulatory regions, are different in the two genes, the corresponding proteins are identical [31] and also highly conserved in evolution (see Figure 2). Thus, it is highly probable that the existence of the two genes is not important for having two proteins with different activities but, instead, because it offers the possibility to regulate the genes (and the related mRNAs) independently and/or with different mechanisms [32]. In particular, it was suggested that the two genes may have cell type-specific expression [33], although their overall activity in different tissues was reported to be quite similar [34].
Actually, the H3.3 protein is not so different with respect to the canonical H3.1 and H3.2 isoforms: indeed, it differs by only five and four amino acids, respectively, from them [35,36,37]. However, these amino acids are, for example, sufficient to allow H3.3 to interact with specific histone chaperones, such as the Death domain-associated protein (DAXX), the alpha-thalassemia/mental retardation X-linked protein (ATRX) complex, and the histone regulator A (Hira)/calcineurin-binding protein 1 (Cabin 1)/ubinuclein1 (Ubn1) complex, involved in its loading on chromatin [7,38,39,40,41]. The H3.3 interaction with these chaperones is determinant for its deposition on specific regions of the genome. It was shown, for example, that a mutation of the ATRX complex leads, as a consequence, to variation in the deposition of H3.3 and chromatin accessibility in association with an alteration in gene expression [42].
The H3.3 variant is indeed very often bound to active chromatin and regulates transcription. In 2002, Ahmad and Henikoff reported that in Drosophila cells, H3.3 is the only H3 species deposited in chromatin in a replication-independent way, and they suggested that this event might be responsible for the activation of genes previously silenced because of histone PTMs [43]. As a confirmation of its gene-activating function, the H3.3 histone was also found at the level of active enhancers [44,45,46].
It was also reported that some chromatin remodeling complexes, such as the SWItch/Sucrose Non-Fermentable (SWI/SNF) complex and, in particular, its subunit “T-rich interactive domain-containing protein 1A” (ARAD1A) are required for maintaining the H3.3 histone at the level of regulatory sequences, among which are the so-called super-enhancers [47]. On the other hand, by interacting with the ATRX/DAXX chaperones, H3.3 might also be loaded on pericentric heterochromatin and telomeres [38,48,49]. This event, together with lysine 56 (H3.3K56) acetylation, seems necessary for chromosome segregation in mammals. Indeed, it was shown that cell lines carrying the mutation K56R increase cell death and modify cell morphology [50].
Recently, a brain-specific function of the chromodomain-helicase-DNA binding protein 1 (CHD1) which is a member of the SWI/SNF family of chromatin remodeling complexes was reported in Drosophila. CHD1 is indeed involved in the loading of H3.3 in the fly brain, where it seems to contribute to the regulation of genes that control the homeostasis of hunger and satiety signals [51]. On the other hand, as a demonstration of the wide range of tissues and functions in which H3.3 histone is probably involved, it was also found to be essential for the chromatin transitions that accompany Drosophila male germline maturation [52].
Further work is required to understand how H3.3, which, as mentioned above, is not so different from the canonical H3 species, can stimulate transcription. Evidently, its sequence should contain features that are able to attract, directly or indirectly, the transcriptional apparatus to the genes to which it is bound. Interestingly, for example, the H3.3 amino-terminal tail contains a serine residue (S31) that is not present in the other H3 species (which contain, instead, an alanine at that position). Moreover, this serine can be phosphorylated, and it has been suggested that this might represent a feature determinant for preferential transcription [53]. It should be noted that S31 is actually S32 in the original amino acid sequence of the H3.3 protein (Figure 2); however, in the mature protein, it becomes S31 because the initiator methionine is immediately cleaved during translation ([54] and references therein). Notably, it was recently demonstrated that S31 phosphorylation can also modify the accessibility of regulatory factors at telomeres during replication, thus stabilizing heterochromatin probably by influencing the activity of histone lysine demethylase 4B (KDM4B) [55].
One possible link between H3.3 modification and gene expression was suggested by Martire and co-workers [56]. Using mouse embryonal stem cells (mESCs), these authors showed that the cells missing histone H3.3 cannot normally acetylate the enhancers that are activated during differentiation and, more specifically, show a reduction in the acetylation of the H3 histone at lysine 27. The normal ability to regulate acetylation would depend on the stimulation of p300 acetyltransferase by phosphorylation at specific sites of the H3.3 histone variant [56,57].

Figure 2.
Alignment of H3.3 histones in different mammalian species. NCBI reference sequences reported: Homo sapiens (NP_005315.1); Pan troglodytes (NP_001267377.1); Mus musculus (NP_032237.1); Rattus norvegicus (NP_446437.1); Equus caballus (XP_023499762.1); Loxodonta africana (XP_010595256.1); Sus scrofa (NP_999095.1); Bos taurus (NP_001014411.1). Alignment of the shown sequences was completed using the Bioedit sequence alignment editor [58].
3. The H3.3 Variant in Brain Development and Maturation as Well as in the Processes of Learning and Memory
As mentioned above, all the cells of the same organism have the same genome. However, thanks to rearrangements in the structural organization of chromatin, each cell expresses only the genes necessary for basal metabolism, as well as the genes involved in its tissue-specific functions. The cell-specific overall organization of chromatin, in turn, derives from the specific history of that cell. In other words, we can state that the nuclear organization of the genome is a form of cell memory, since, at any moment, it reflects the experiences that a cell has had in its life. This epigenetic memory is built up with the specific combination and location of the different histone variants and with their specific post-translational modifications (PTMs) [59,60], thus creating a sort of “histone code” [12,61] that is superimposed to the genetic code. In the case of the H3 histone isoforms, Hake and Allis proposed the “H3 barcode hypothesis”, which suggests the existence of chromatin indexing based on the specific distribution of canonical and non-canonical H3 species, as well as on their PTMs [62]. Notably, it was also reported that H3.3 accumulates with age in most somatic tissues of mice and that this accumulation correlates with a general age-dependent change in H3 methylation [63]. This observation might be the most important since, in spite of the large body of evidence showing the involvement of H3.3 in gene transcription, it still remains not completely clear how the presence of this histone variant in chromatin can drive the activation of specific genes. Thus, it is highly probable that the H3.3 effect is also due to its specific interaction with PTM writers and/or PTM readers.
Interestingly, many chromatin-specific marks should be transmitted to the daughter cells during each cycle of DNA replication and cell division [64]; if the marks are lost, specific gene expression and, hence, central aspects of cell phenotype are equally lost. It was shown, for example, that the canonical isoforms of the H3 histone, as well as their epigenetic marks, are transferred from the mother to the daughter cells [64,65,66,67]. Similarly, H3.3-containing nucleosomes can be replicated as such during cell division, thus contributing to the epigenetic memory of the cells [68,69,70]. The importance of the events that allow transgenerational transmission of these marks is also underlined by the fact that in vertebrates, loss of chromatin assembly factor 1 (CAF1), the main chaperone involved in the DNA replication-dependent deposition of canonical H3 histones, blocks embryo development [71,72]. On the other hand, the non-canonical H3.3 histone seems to be specifically loaded at chromatin sites where nucleosomes should face rapid turnover, thus allowing active transcription [37,73,74]. Indeed, the position of H3.3 in chromatin often coincides with that of RNA polymerase II (RNAPII) [58].
As for other cells, epigenetic marks, such as histone acetylation and DNA methylation, accumulate in neuronal chromatin during cellular responses to received internal and/or environmental stimuli [75,76,77,78,79]. As a whole, chromatin DNA and protein modifications are fundamental in the control of neural progenitor (NPC) development and differentiation; for example, trimethylation of lysine 27 in the H3 histone (H3K27me3, a repressive mark) should be removed by the Jumonji C domain-containing 3 (Jmjd3) demethylase in order to allow embryonic stem cells (ESCs) to give rise to NPCs [80,81,82]. Similar events are also linked to neuronal migration [82,83]. Interestingly, in the epigenetic control of gene expression during brain maturation, microRNAs seem to also be involved, thanks to cross-talk with other epigenetic regulators. In particular, the identification of astrocyte-enriched microRNAs has been reported, and their possible role in NPC differentiation into glial cells has been suggested [84].
It is also worth noting that, recently, a new kind of histone PTM was described: modification by monoamine neurotransmitters, such as dopamine and serotonin [85,86,87,88]. Some of these modifications seem to have H3 histones as specific targets [87,88]. For example, it was reported that serotonylation involves glutamine 5 of H3 histones and that the presence of this modified H3Q5 can, in turn, stabilize H3K4 methylation [89]. While further studies are necessary to fully understand the mechanisms, functions, and regulation of histone monoamination, it is already clear that this kind of PTM can have a powerful effect on the nervous system, where neurotransmitters, such as dopamine, noradrenaline, or serotonin, are highly represented. Notably, it is known that, in addition to the monoamines stored in synaptic vesicles and ready for neurotransmission, a significant number of them are not in vesicles but are present in the neuronal soma and even in the cell nucleus. Moreover, these molecules were involved in development well before the appearance of nerve cells [88,90,91]. Notably, it was also reported that histone H3 dopaminylation can be involved in the response to abuse drugs, such as cocaine [92,93].
In addition to PTMs, loading of the H3.3 histone variant onto chromatin seems to be required for neuronal differentiation and, in particular, for activation of mature neuron-specific genes [94,95]. For example, during mouse lens fiber cell differentiation, high concentrations of H3.3 were observed at the level of transcribed regions in chromatin, like those including genes encoding crystallins, gap junction components, and intermediate filaments [96]. Actually, H3.3, together with an H2A variant known as H2A.Z, contributes to opening chromatin structure to transcription factors, by increasing accessibility of the upstream regulatory sequences present in the genes to which they are bound [57,97,98]. However, the precise mechanisms underlying how H3.3 causes chromatin opening are not completely clear. For example, using a genomic analysis on mouse embryonic stem cells in which H3.3 knockout had been induced, it was found that there is a general deregulation of promoter activity at the level of expressed genes associated with a reduction in transcription factor binding and a consequent reduction in RNA polymerase II at the transcription start sites [57].
Now, if we accept the principle that nuclear chromatin organization confers to each cell a specific phenotype because it represents the epigenetic memory of the history of that cell [99], then we have to admit that it will be even more so, and lifelong, for cells that are responsible for the macroscopic ability to learn and remember, namely neurons and probably also some glial cells such as astrocytes [100]. Indeed, epigenetic changes in chromatin, such as post-translational modifications of histones and, in particular, acetylation, are involved in the phenomenon of long-term potentiation (LTP), which is in the stabilization of memory [82,94,101,102,103]. For example, it was found that H3 and H4 histone acetylation at the level of the promoter for the gene encoding the brain-derived neurotrophic factor (BDNF) is fundamental for increasing BDNF expression in the rat hippocampus and for allowing LTP [82].
In addition to a variety of histone PTMs, the H3.3 histone is also enriched on active chromatin related to memory storing [104], and it seems to be fundamental for maintaining neuronal activity in the mouse hippocampus, as an H3.3 decrease can alter long-term memory [94]. Again, the H3.3 histone shares its ability to act as a regulator of hippocampal memory with another histone variant known as H2A1, the localization of which was found to dynamically change during the learning process [105].
As mentioned above, the H3.3 histone also has a role to play at certain heterochromatic regions, such as telomeres and pericentric repeats, where it is trimethylated at the level of its lysine 9 (H3.3K9me3), a modification bound to its ability to stabilize heterochromatin [49]. Recently, it was reported that phosphorylation at the level of its serine 31, one of the residues for which it differs from H3.1 and H3.2, stabilizes heterochromatin by inhibiting lysine-specific demethylase 4B (KDM4B), which could otherwise demethylate H3K9/K36, thus damaging the heterochromatic structure [55].
The importance of the H3.3 histone in brain development was confirmed with the discovery and analysis of different neurodevelopmental disorders. For example, mutations in the H3.3A/H3.3B genes have been indicated as new candidates for microcephaly, and intellectual disability, as well as in a novel neurodevelopmental disorder now known as Bryant-Li-Bhoj syndrome [32,106,107]. Indeed, experiments in which H3.3 expression was silenced with RNA interference demonstrated profound alterations in both the structural and functional organization of the brain, with clear effects on the mechanisms underlying memory processes [94,95]. Moreover, stage-dependent deletion of the two H3.3-encoding genes demonstrated that synthesis of H3.3 is required in the first postnatal days of rat life, thus indicating the importance of de novo accumulation of this variant at the beginning of brain development [108]. In this phase, it was clearly shown that genes involved in the proliferative stage of brain development are silenced, while a collection of genes are activated in order to drive neuronal differentiation, with even different genes involved in the specification of different layers of the developing cortex [108,109,110,111].
4. Histone Post-Translational Modifications and the H3.3 Variant in Neurodegenerative Diseases
Some studies have also shown that PTMs of histones can contribute to the pathogenesis of neurodegenerative diseases, such as Parkinson’s disease (PD) and Huntington’s disease (HD). In an experimental model of PD, exposure of dopaminergic cells (N27 cells) from the rat midbrain to pesticides, i.e., neurotoxic substances, induced hyperacetylation of the core histones H3/H4 and the subsequent death of dopaminergic neurons. The inhibition of hyperacetylation was neuroprotective, preventing N27 cell apoptosis [112]. Notably, it was suggested that histone hyperacetylation in PD may be a consequence of mitochondrial damage, a dysfunction-enhancing progression in PD. Indeed, in both ex vivo and in vivo neurodegenerative models of PD, induced perturbation of the mitochondria, reduced ATP production, and altered intracellular activity of proteasomes induced the hyper-activation of histone acetyltransferase (HAT) and inhibition of histone deacetylase (HDAC). This resulted in H3.3 lysine 27 hyperacetylation (H3.3K27). This epigenetic alteration directly induced the death of dopaminergic neurons by affecting gene transcription. Also, in post-mortem brains from patients with PD, histone extracts from substantia nigra lysates showed a significant nuclear accumulation of H3K27 acetylation in comparison to age-matched controls [113]. Reduced trimethylation of histone H3 lysine 4 (H3K4) on promoters of down-regulated genes was also evidenced in mice and human models of Huntington’s disease (HD). Moreover, loss of the enzyme known as little imaginal discs (Lid), a trimethyl-H3K4 demethylating enzyme, reduces neurodegeneration in Drosophila models of HD [114]. Alteration of H3K4 methylation was also demonstrated by Dong and collaborators using human post-mortem tissues [115]. However, modulation of histone methylation and demethylation may have opposite effects on neurodegeneration in HD. Indeed, reduced methylation of H3K27 for mutations of the polycomb repressive complex 2 (PRC2), which is responsible for the methylation of H3K27, promoted neurodegenerative processes, while inhibition of the activity of the H3K27 demethylase Utx (ubiquitously transcribed tetratricopeptide repeat, X chromosome) reduced pathological modifications in HD [116].
Interestingly, it was also suggested that mutant huntingtin, which accumulates in HD, can interact with HAT enzymes, inducing a decrease in their activity [60,117]. This observation is of special importance when we consider that histones should be acetylated in order to be degraded [118], and that epigenetic modifications of cell activity also include histone substitution at specific loci in chromatin. On the other hand, it is also important to consider that histone acetylation requires, as a substrate, Acetyl-CoA produced in the mitochondria, and this clearly means that, in general terms, metabolic dysfunctions can also hamper histone PTMs [119].
5. The H3.3 Variant in Cancer
Notably, alterations of histone sequences and PTMs play a role in cancer [21,120]. As discussed above, chromatin modification and accessibility largely depend on enzymatic systems that introduce PTMs, on histone variant loading into chromatin, and on factors that are able to read such modifications, thus activating transcription. Obviously, given the H3.3 transcription activating function, it does not come as a surprise that it is found mutated in many tumors [121,122,123]. In some cases, it also promotes metastasis, favoring all the mechanisms that underly the basis of this phenomenon. For example, the genes for epithelial–mesenchymal transition (EMT), like SRY-box transcription factor 9 (Sox9) and Snail, are stimulated in certain cancers [123], also thanks to the stabilization of the Hira complex [124].
Interestingly, a variety of somatic mutations have been discovered in histone genes [21,125]. It is worth noting that, independent of the number of mutated histone genes, the effect of mutation is dominant in inducing transformation, very often in children and adolescents [21,126,127]. Actually, there are features that clearly distinguish adult from pediatric gliomas, and one of them is the mutation of proteins related to the chromatin structure [128].
Now, cancer cells are clearly characterized by modification of their metabolism: in particular, they show an increase in lipid metabolism, thanks to which they can produce higher amounts of membranes in order to proliferate. Synthesis of higher amounts of lipids, in turn, requires reduced nicotinamide adenine dinucleotide phosphate (NADPH). Indeed, it was recently demonstrated the connection between breast cancer metastasis and the H3.3-driven upregulation of NADK enzyme that, in turn, favors NAPDH production, thus sustaining proliferation [129]. Another metabolic property of cancer cells is a high methionine requirement, especially for the ability of the adenosyl-derivative of methionine (S-Adenosyl methionine, SAM) to be used as a cofactor in both DNA and histone methylation [130].
A mutation of both H3.3 and H3.1 histones has been found in diffuse intrinsic pontine glioma (DIPG), a very aggressive tumor with a median overall survival of 8–11 months. This kind of tumor belongs to the category of diffuse midline gliomas (DMG) and primarily occurs in children. The lysine to methionine mutation at position 27 (K27M) of the H3.3 histone, in particular, is often associated with TP53 and ATRX mutations [131]. These latter mutations have been mainly found in midline brain structures, such as the thalamus, pons, and brainstem, where they associate with the alteration of the normal post-translational modification pattern of the H3 histone. The following variation in gene expression often leads to tumor development, even though it is not yet clear how these events are connected. One interesting finding is that the H3.3 K27M mutation differs from H3.1 K27M both for the association to secondary mutation and for the age of tumor onset [127,132,133,134]. Comparing two cell lines, one of which bore K27M mutation, Lewis and colleagues demonstrated that the mutated cell line presents a specific region of accessible chromatin, containing genes involved in neurogenesis and neuronal development, like the Achaete–Scute Family BHLH transcription factor 1 (ASCL1) and NeuroD [133].
One of the suggested H3.3 interactors is polycomb repressive complex 2 (PRC2), a methyltransferase involved in the control of gene expression and, more precisely, in the silencing of the promoters of specific genes by the trimethylation of lysine 27 of the H3 histones. In particular, in vitro studies showed binding between PRC2 and the mutated H3.3 [135,136]. The effect of the interaction between the H3.3 K27M mutated protein and the PRC2 complex is inhibition in the activity of the complex itself: the enhancer of the zeste homolog 2 (EZH2) component, a methyltransferase, stops methylating the other H3.3 histone residues in adjacent nucleosomes, causing general hypomethylation [31,136]. On the other hand, the interaction has not been proven in vivo, and the localization of the two proteins does not seem to coincide in the nucleus [137].
Another frequent mutation found in histone H3.3 in gliomas is the glycine to arginine (or valine) variation at position 34 (G34R/V). It has been suggested that this mutation affects the activity of the enzymes that modify the status of lysine 36 methylation, but a clear mechanism deriving from the original defect is not yet known. Differently from the K27M mutation, which can affect both the H3.3 and H3.1 histones, the G34R mutation is specific to the variant H3.3 [127].
As already mentioned, ATRX is a partner of the DAXX chaperone and forms with it a complex that deposits H3.3 on chromatin. A mutation to the components of this complex is frequent in different cancers and is associated with the pathway called alternative lengthening of telomeres (ALT) [138].
In many pediatric high-grade gliomas, ATRX is mutated, and, in the majority of cases, the evidenced mutation is associated with either the H3.3 K27M or H3.3 G34R/V mutation [132].
Moreover, the G34R/V mutation causes a reduced capacity of DNA repair in patients suffering from pHGG (high-grade glioma), leading to an enhanced susceptibility to DNA damage, also from radiotherapy [139].
In the alveolar rhabdomyosarcoma, H3.3 is overexpressed both in vivo and in cell lines: in this latter system, knockdown of the H3F3A gene reduces the ability of the cell to migrate, suggesting that the H3.3 variant regulates genes involved in motility, like melanoma cell adhesion molecule (MCAM, also known as CD146) [140].
Mutations of the H3.3 histone were also described in bone and cartilage cancers, such as the giant cell tumor (GCT) of bone [141]. One H3.3 mutation, changing lysine at position 36 with methionine (K36M), is associated with chondroblastoma, another tumor occurring especially in childhood [142]. The mutated H3.3 protein seems to block the activity of the multiple myeloma SET domain (MMSET) and SET domain containing 2 (SETD2) methyltransferases, altering the methylation landscape and gene expression and thus leading to neoplastic transformation [143].
6. Conclusions and Perspectives
In conclusion, all the cells of an organism are able to store memory of their past experiences in the form of specific modifications to chromatin structural organization. On the other hand, nerve cells not only store information related to the steps that allowed their development and differentiation, but they also store information related to events that determine macroscopic phenomena known as learning and memory. However, at the molecular level, the basal mechanisms that allow cognitive acquisition are very similar to the ones used by all cells to remember their history. Indeed, DNA methylation, histone PTMs of many kinds, and histone variant loading into chromatin constitute a collection of marks that is able to index the different genes to be transcribed. All these events, as discussed, are of central importance both for the general and specific structural organization of genes, and they also have an effect because of the existing interactions among different marks, as well as the binding of further factors (the readers), which are able to recognize the marks. Most importantly, long-term potentiation relies on these kinds of events, and it is induced mostly in the periphery of the nerve cells, as it depends on the signals that arrive through a variety of mechanisms, such as wiring (synaptic) and volume transmission (probably also mediated by extracellular vesicles) [100]. Therefore, we propose that a central aspect of all these events is the regulation of peripheral translation of mRNAs that encode proteins, which are then able to reach the nucleus, inducing adaptive modifications of nuclear chromatin. Pre-localization of mRNA is indeed an important aspect of adaptation in the nervous system, and the proteins that are able to bind mRNAs and transport/localize them play a key role both in neurodevelopment and the functions of the adult brain [144]. Moreover, RBPs also interact with non-coding RNAs, which are also part of the network that is able to regulate cell phenotype at the epigenetic level [144]. As a demonstration of the central epigenetic importance of RBPs in the nervous system functions, we finally cite the fact that, in almost all the neurodegenerative pathologies, alteration to the function/localization of RBPs has been evidenced [144].
Author Contributions
Conceptualization: I.D.L.; artworks: I.D.L. and C.M.D.L.; writing, and editing: all the authors. All authors have read and agreed to the published version of the manuscript.
Funding
The authors did not receive any external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The cited data are from bibliography and are thus all available in the literature.
Acknowledgments
The authors are supported by the Università degli Studi di Palermo (University of Palermo), Palermo, Italy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gurdon, J.B.; Elsdale, T.R.; Fischberg, M. Sexually mature individuals of Xenopus laevis from the transplantation of single somatic nuclei. Nature 1958, 182, 64–65. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. Chromatin structure: A repeating unit of histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Felsenfeld, G. A brief history of epigenetics. Cold Spring Harb. Perspect. Biol. 2014, 6, a018200. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin accessibility and the regulatory epigenome. Nat. Rev. Genet. 2019, 20, 207–220. [Google Scholar] [CrossRef]
- Sokolova, V.; Sarkar, S.; Tan, D. Histone variants and chromatin structure, update of advances. Comput. Struct. Biotechnol. J. 2022, 21, 299–311. [Google Scholar] [CrossRef]
- Hergeth, S.P.; Schneider, R. The H1 linker histones: Multifunctional proteins beyond the nucleosomal core particle. EMBO Rep. 2015, 16, 1439–1453. [Google Scholar] [CrossRef]
- Waddington, C.H. The epigenotype. 1942. Int. J. Epidemiol. 2012, 41, 10–13. [Google Scholar] [CrossRef]
- Kim, U.; Lee, D.-S. Epigenetic Regulations in Mammalian Cells: Roles and Profiling Techniques. Mol. Cells 2023, 46, 86–98. [Google Scholar] [CrossRef]
- Kato, S.; Yokoyama, A.; Fujiki, R. Nuclear receptor coregulators merge transcriptional coregulation with epigenetic regulation. Trends Biochem. Sci. 2011, 36, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, G.; Qian, J. Transcription factors as readers and effectors of DNA methylation. Nat. Rev. Genet. 2016, 17, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Hajkova, P.; Ecker, J.R. Dynamic DNA methylation: In the right place at the right time. Science 2018, 361, 1336–1340. [Google Scholar] [CrossRef]
- Jiang, D.; Li, T.; Guo, C.; Tang, T.S.; Liu, H. Small molecule modulators of chromatin remodeling: From neurodevelopment to neurodegeneration. Cell Biosci. 2023, 13, 10. [Google Scholar] [CrossRef]
- Shindo, Y.; Brown, M.G.; Amodeo, A.A. Versatile roles for histones in early development. Curr. Opin. Cell Biol. 2022, 75, 102069. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, W.J.; Zhang, Q.; Su, Y.H.; Tang, L.P. The dynamics of chromatin states mediated by epigenetic modifications during somatic cell reprogramming. Front. Cell. Dev. Biol. 2023, 11, 1097780. [Google Scholar] [CrossRef]
- Simon, L.; Probst, A.V. Maintenance and dynamic reprogramming of chromatin organization during development. Plant J. 2023. [Google Scholar] [CrossRef]
- Marzluff, W.F.; Gongidi, P.; Woods, K.R.; Jin, J.; Maltais, L.J. The human and mouse replication-dependent histone genes. Genomics 2002, 80, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Marzluff, W.F.; Wagner, E.J.; Duronio, R.J. Metabolism and regulation of canonical histone mRNAs: Life without a poly(A) tail. Nat. Rev. Genet. 2008, 9, 843–854. [Google Scholar] [CrossRef]
- Amatori, S.; Tavolaro, S.; Gambardella, S.; Fanelli, M. The dark side of histones: Genomic organization and role of oncohistones in cancer. Clin. Epigenetics 2021, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Talbert, P.B.; Henikoff, S. Histone variants at a glance. J. Cell Sci. 2021, 134, jcs244749. [Google Scholar] [CrossRef] [PubMed]
- Castiglia, D.; Cestelli, A.; Scaturro, M.; Nastasi, T.; Di Liegro, I. H1.0 and H3.3B mRNA levels in developing rat brain. Neurochem. Res. 1994, 19, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Scaturro, M.; Cestelli, A.; Castiglia, D.; Nastasi, T.; Di Liegro, I. Posttranscriptional Regulation of H1.0 and H3.3B histone genes in differentiating rat cortical neurons. Neurochem. Res. 1995, 20, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Castiglia, D.; Scaturro, M.; Nastasi, T.; Cestelli, A.; Di Liegro, I. PIPPin, a putative RNA-binding protein specifically expressed in the rat brain. Biochem. Biophys. Res. Commun. 1996, 218, 390–394. [Google Scholar] [CrossRef]
- Scaturro, M.; Nastasi, T.; Raimondi, L.; Bellafiore, M.; Cestelli, A.; Di Liegro, I. H1(0) RNA-binding proteins specifically expressed in the rat brain. J. Biol. Chem. 1998, 273, 22788–22791. [Google Scholar] [CrossRef]
- Nastasi, T.; Scaturro, M.; Bellafiore, M.; Raimondi, L.; Beccari, S.; Cestelli, A.; Di Liegro, I. PIPPin is a brain-specific protein that contains a cold-shock domain and binds specifically to H1 degrees and H3.3 mRNAs. J. Biol Chem. 1999, 274, 24087–24093. [Google Scholar] [CrossRef]
- Sala, A.; Scaturro, M.; Proia, P.; Schiera, G.; Balistreri, E.; Aflalo-Rattenbach, R.; Créau, N.; Di Liegro, I. Cloning of a rat-specific long PCP4/PEP19 isoform. Int. J. Mol. Med. 2007, 19, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Saladino, P.; Di Liegro, C.M.; Proia, P.; Sala, A.; Schiera, G.; Lo Cicero, A.; Di Liegro, I. RNA-binding activity of the rat calmodulin-binding PEP-19 protein and of the long PEP-19 isoform. Int. J. Mol. Med. 2012, 29, 141–145. [Google Scholar] [CrossRef]
- Weaver, K.J.; Holt, R.A.; Henry, E.; Pletcher, S.D. Effects of hunger on neuronal histone modifications slow aging in Drosophila. Science 2023, 380, 625–632. [Google Scholar] [CrossRef]
- Kumar, V.C.; Pai, R. Genes of the month: H3.3 histone genes: H3F3A and H3F3B. J. Clin. Pathol. 2021, 74, 753–758. [Google Scholar] [CrossRef]
- Bryant, L.; Sangree, A.; Clark, K.; Bhoj, E. Histone 3.3-related chromatinopathy: Missense variants throughout H3-3A and H3-3B cause a range of functional consequences across species. Hum. Genet. 2023. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Cervantes, V.; Yee, J.Q.; Klein, R.H.; Knoepfler, P.S. A knockout-first model of H3f3a gene targeting leads to developmental lethality. Genesis 2023, 61, e23507. [Google Scholar] [CrossRef] [PubMed]
- Bachu, M.; Tamura, T.; Chen, C.; Narain, A.; Nehru, V.; Sarai, N.; Ghosh, S.B.; Ghosh, A.; Kavarthapu, R.; Dufau, M.L.; et al. A versatile mouse model of epitope-tagged histone H3.3 to study epigenome dynamics. J. Biol. Chem. 2019, 294, 1904–1914. [Google Scholar] [CrossRef]
- Elsaesser, S.J.; Goldberg, A.D.; Allis, C.D. New functions for an old variant: No substitute for histone H3.3. Curr. Opin. Genet. Dev. 2010, 20, 110–117. [Google Scholar] [CrossRef]
- Filipescu, D.; Szenker, E.; Almouzni, G. Developmental roles of histone H3 variants and their chaperones. Trends Genet. 2013, 29, 630–640. [Google Scholar] [CrossRef]
- Delaney, K.; Almouzni, G. Transcription-coupled H3.3 recycling: A link with chromatin states. Semin. Cell Dev. Biol. 2023, 135, 13–23. [Google Scholar] [CrossRef]
- Lewis, P.W.; Elsaesser, S.J.; Noh, K.M.; Stadler, S.C.; Allis, C.D. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl Acad. Sci. USA 2010, 107, 14075–14080. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Banaszynski, L.A.; Noh, K.M.; Lewis, P.W.; Elsaesser, S.J.; Stadler, S.; Dewell, S.; Law, M.; Guo, X.; Li, X.; et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 2010, 140, 678–691. [Google Scholar] [CrossRef]
- Smith, R.; Susor, A.; Ming, H.; Tait, J.; Conti, M.; Jiang, Z.; Lin, C.-J. The H3.3 chaperone Hira complex orchestrates oocyte developmental competence. Development 2022, 149, dev200044. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Xiong, C.; Chen, J.; Wang, L.; Wen, Z.; Yu, J.; Chen, P.; Xu, Y.; Jin, J.; et al. HIRA complex presets transcriptional potential through coordinating depositions of the histone variants H3.3 and H2A.Z on the poised genes in mESCs. Nucleic Acids Res. 2022, 50, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Truch, J.; Downes, D.J.; Scott, C.; Gür, E.R.; Telenius, J.M.; Repapi, E.; Schwessinger, R.; Gosden, M.; Brown, J.M.; Taylor, S.; et al. The chromatin remodeller ATRX facilitates diverse nuclear processes, in a stochastic manner, in both heterochromatin and euchromatin. Nat. Commun. 2022, 13, 3485. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Henikoff, S. The histone Variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell 2002, 9, 1191–1200. [Google Scholar] [CrossRef]
- Chen, P.; Zhao, J.; Wang, Y.; Wang, M.; Long, H.; Liang, D.; Huang, L.; Wen, Z.; Li, W.; Li, X.; et al. H3.3 actively marks enhancers and primes gene transcription via opening higher ordered chromatin. Genes Dev. 2013, 27, 2109–2124. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A.M.; Gomez-Rodriguez, M.; Mieczkowski, J.; Tolstorukov, M.Y.; Kundu, S.; Sadreyev, R.I.; Jansen, L.E.; Kingston, R.E. Enhancer regions show high histone H3.3 turnover that changes during differentiation. Elife 2016, 5, e15316. [Google Scholar] [CrossRef]
- Shi, L.; Wen, H.; Shi, X. The histone variant H3.3 in transcriptional regulation and human disease. J. Mol. Biol. 2017, 429, 1934–1945. [Google Scholar] [CrossRef] [PubMed]
- Reske, J.J.; Wilson, M.R.; Armistead, B.; Harkins, S.; Perez, C.; Hrit, J.; Adams, M.; Rothbart, S.B.; Missmer, S.A.; Fazleabas, A.T.; et al. ARID1A-dependent maintenance of H3.3 is required for repressive CHD4-ZMYND8 chromatin interactions at super-enhancers. BMC Biol. 2022, 20, 209. [Google Scholar] [CrossRef]
- Wong, L.H.; Ren, H.; Williams, E.; McGhie, J.; Ahn, S.; Sim, M.; Tam, A.; Earle, E.; Anderson, M.A.; Mann, J.; et al. Histone H3.3 incorporation provides a unique and functionally essential telomeric chromatin in embryonic stem cells. Genome Res. 2009, 19, 404–414. [Google Scholar] [CrossRef]
- Udugama, M.; Chang, F.T.M.; Chan, F.L.; Tang, M.C.; Pickett, H.A.; McGhie, J.D.R.; Mayne, L.; Collas, P.; Mann, J.R.; Wong, L.H. Histone variant H3.3 provides the heterochromatic H3 lysine 9 tri-methylation mark at telomeres. Nucleic Acids Res. 2015, 43, 10227–10237. [Google Scholar] [CrossRef]
- Fang, L.; Chen, D.; Zhang, J.; Li, H.; Bradford, B.; Jin, C. Potential functions of histone H3.3 lysine 56 acetylation in mammals. Epigenetics 2022, 17, 498–517. [Google Scholar] [CrossRef]
- Schoberleitner, I.; Mertens, B.; Bauer, I.; Lusser, A. Regulation of sensory perception and motor abilities by brain-specific action of chromatin remodelling factor CHD1. Front. Mol. Neurosci. 2022, 15, 840966. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Schwartz, B.E.; Goldstein, S.; Ahmad, K. Transcriptional and developmental functions of the H3.3 histone variant in Drosophila. Curr. Biol. 2009, 19, 1816–1820. [Google Scholar] [CrossRef]
- Armache, A.; Yang, S.; Martínez de Paz, A.; Robbins, L.E.; Durmaz, C.; Cheong, J.Q.; Ravishankar, A.; Daman, A.W.; Ahimovic, D.J.; Klevorn, T.; et al. Histone H3.3 phosphorylation amplifies stimulation-induced transcription. Nature 2020, 583, 852–857. [Google Scholar] [CrossRef]
- Chen, J.; Horton, J.; Sagum, C.; Zhou, J.; Cheng, X.; Bedford, M.T. Histone H3 N-terminal mimicry drives a novel network of methyl-effector interactions. Biochem. J. 2021, 478, 1943–1958. [Google Scholar] [CrossRef] [PubMed]
- Udugama, M.; Vinod, B.; Chan, F.L.; Hii, L.; Garvie, A.; Collas, P.; Kalitsis, P.; Steer, D.; Das, P.P.; Tripathi, P.; et al. Histone H3.3 phosphorylation promotes heterochromatin formation by inhibiting H3K9/K36 histone demethylase. Nucleic Acids Res. 2022, 50, 4500–4514. [Google Scholar] [CrossRef]
- Martire, S.; Gogate, A.A.; Whitmill, A.; Tafessu, A.; Nguyen, J.; Teng, Y.C.; Tastemel, M.; Banaszynski, L.A. Phosphorylation of histone H3.3 at serine 31 promotes p300 activity and enhancer acetylation. Nat. Genet. 2019, 51, 941–946. [Google Scholar] [CrossRef]
- Tafessu, A.; O’Hara, R.; Martire, S.; Dube, A.L.; Saha, P.; Gant, V.U.; Banaszynski, L.A. H3.3 contributes to chromatin accessibility and transcription factor binding at promoter-proximal regulatory elements in embryonic stem cells. Genome Biol. 2023, 24, 25. [Google Scholar] [CrossRef] [PubMed]
- Wollmann, H.; Holec, S.; Alden, K.; Clarke, N.D.; Jacques, P.E.; Berger, F. Dynamic deposition of histone variant H3.3 accompanies developmental remodeling of the Arabidopsis transcriptome. PLoS Genet. 2012, 8, e1002658. [Google Scholar] [CrossRef]
- Duan, G.; Walther, D. The roles of post-translational modifications in the context of protein interaction networks. PLoS Comput. Biol. 2015, 11, e1004049. [Google Scholar] [CrossRef]
- Kabir, F.; Atkinson, R.; Cook, A.L.; Phipps, A.J.; King, A.E. The role of altered protein acetylation in neurodegenerative disease. Front. Aging Neurosci. 2023, 14, 1025473. [Google Scholar] [CrossRef]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef]
- Hake, S.B.; Allis, C.D. Histone H3 variants and their potential role in indexing mammalian genomes: The "H3 barcode hypothesis". Proc. Natl. Acad. Sci. USA 2006, 103, 6428–6435. [Google Scholar] [CrossRef]
- Tvardovskiy, A.; Schwämmie, V.; Kempf, S.J.; Rogowska-Wrzesinska, A.; Jensen, O.N. Accumulation of histone variant H3.3 with age is associated with profound changes in the histone methylation landscape. Nucleic Acids Res. 2017, 45, 9272–9289. [Google Scholar] [CrossRef] [PubMed]
- Loppin, B.; Berger, F. Histone variants: The nexus of developmental decisions and epigenetic memory. Annu. Rev. Genet. 2020, 54, 121–149. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Long, C.; Chen, X.; Huang, C.; Chen, S.; Zhu, B. Partitioning of histone H3-H4 tetramers during DNA replication-dependent chromatin assembly. Science 2010, 328, 94–98. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, Z.; Xu, M.; Li, Y.; Li, Z.; Ma, Y.; Cai, T.; Zhu, B. H3.3-H4 tetramer splitting events feature cell-type specific enhancers. PLoS Genet. 2013, 9, e1003558. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, A.T. The fork in the road: Histone partitioning during DNA replication. Genes 2015, 6, 353–371. [Google Scholar] [CrossRef] [PubMed]
- Ng, R.K.; Gurdon, J.B. Epigenetic inheritance of cell differentiation status. Cell Cycle 2008, 7, 1173–1177. [Google Scholar] [CrossRef] [PubMed]
- Ng, R.K.; Gurdon, J.B. Epigenetic memory of an active gene state depends on histone H3.3 incorporation into chromatin in the absence of transcription. Nat. Cell Biol. 2008, 10, 102–109. [Google Scholar] [CrossRef]
- Xu, X.; Duan, S.; Hua, X.; Li, Z.; He, R.; Zhang, Z. Stable inheritance of H3.3-containing nucleosomes during mitotic cell division. Nat. Commun. 2022, 13, 2514. [Google Scholar] [CrossRef]
- Smith, S.; Stillman, B. Purification and characterization of CAF-I, a human cell factor required for chromatin assembly during DNA replication in vitro. Cell 1989, 58, 15–25. [Google Scholar] [CrossRef]
- Shibahara, K.; Stillman, B. Replication-dependent marking of DNA by PCNA facilitates CAF-1-coupled inheritance of chromatin. Cell 1999, 96, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.-M.; Georgiou, A.; Szutorisz, H.; Maia e Silvia, A.; Pombo, A.; Barahona, I.; Dargelos, E.; Canzonetta, C.; Dillon, N. Variant histone H3.3 marks promoters of transcriptionally active genes during mammalian cell division. EMBO Rep. 2005, 6, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Kraushaar, D.C.; Jin, W.; Maunakea, A.; Abraham, B.; Ha, M.; Zhao, K. Genome-wide incorporation dynamics reveal distinct categories of turnover for the histone variant H3.3. Genome Biol. 2013, 14, R121. [Google Scholar] [CrossRef]
- Borrelli, E.; Nestler, E.J.; Allis, C.D.; Sassone-Corsi, P. Decoding the epigenetic language of neuronal plasticity. Neuron 2008, 60, 961–974. [Google Scholar] [CrossRef]
- Peixoto, L.; Abel, T. The role of histone acetylation in memory formation and cognitive impairments. Neuropsychopharmacology 2013, 38, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, D.A.; Chan, U.; Chen, L.F.; West, A.E. Chromatin regulation of neuronal maturation and plasticity. Trends Neurosci. 2018, 41, 311–324. [Google Scholar] [CrossRef]
- Nord, A.S.; West, A.E. Neurobiological functions of transcriptional enhancers. Nat. Neurosci. 2020, 23, 5–14. [Google Scholar] [CrossRef]
- Giacoman-Lozano, M.; Meléndez-Ramirez, C.; Martinez-Ledesma, E.; Cuevas-Diaz Duran, R.; Velasco, I. Epigenetics of neuronal differentiation: Spotlight on enhancers. Front. Cell Dev. Biol. 2022, 10, 1001701. [Google Scholar] [CrossRef]
- Burgold, T.; Spreafico, F.; De Santa, F.; Totaro, M.G.; Prosperini, E.; Natoli, G.; Testa, G. The histone H3 lysine 27-specific demethylase Jmjd3 is required for neural commitment. PLoS ONE 2008, 3, e3034. [Google Scholar] [CrossRef]
- Desai, D.; Khanna, A.; Pethe, P. PRC1 catalytic unit RING1B regulates early neural differentiation of human pluripotent stem cells. Exp. Cell Res. 2020, 396, 112294. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Chen, H.; Wang, H.; Wang, L. The Histone Modifications of Neuronal Plasticity. Neural Plast. 2021, 2021, 6690523. [Google Scholar] [CrossRef] [PubMed]
- Di Meglio, T.; Kratochwil, C.F.; Vilain, N.; Loche, A.; Vitobello, A.; Yonehara, K.; Hrycaj, S.M.; Roska, B.; Peters, A.H.; Eichmann, A.; et al. Ezh2 orchestrates topographic migration and connectivity of mouse precerebellar neurons. Science 2013, 339, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Cacci, E.; Negri, R.; Biagioni, S.; Lupo, G. Histone Methylation and microRNA-dependent Regulation of Epigenetic Activities in Neural Progenitor Self-Renewal and Differentiation. Curr. Top. Med. Chem. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bader, M. Serotonylation: Serotonin signalling and epigenetics. Front. Mol. Neurosci. 2019, 12, 288. [Google Scholar] [CrossRef]
- Farrelly, L.A.; Thompson, R.E.; Zhao, S.; Lepack, A.E.; Lyu, Y.; Bhanu, N.V.; Zhang, B.; Loh, Y.E.; Ramakrishnan, A.; Vadodaria, K.C.; et al. Histone serotonylation is a permissive modification that enhances TFIID binding to H3K4me3. Nature 2019, 567, 535–539. [Google Scholar] [CrossRef]
- Al-Kachak, A.; Maze, I. Post-translational modifications of histone proteins by monoamine neurotransmitters. Curr. Opin. Chem. Biol. 2023, 74, 102302. [Google Scholar] [CrossRef]
- Zhao, S.; Chuh, K.N.; Zhang, B.; Dul, B.E.; Thompson, R.E.; Farrelly, L.A.; Liu, X.; Xu, N.; Xue, Y.; Roeder, R.G.; et al. Histone H3Q5 serotonylation stabilizes H3K4 methylation and potentiates its readout. Proc. Natl. Acad. Sci. USA 2021, 118, e2016742118. [Google Scholar] [CrossRef]
- Azmitia, E.C. Modern views on an ancient chemical: Serotonin effects on cell proliferation, maturation, and apoptosis. Brain Res. Bull. 2001, 56, 413–424. [Google Scholar] [CrossRef]
- Fukumoto, T.; Kema, I.P.; Levin, M. Serotonin signaling is a very early step in patterning of the left-right axis in chick and frog embryos. Curr. Biol. 2005, 15, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Lepack, A.E.; Werner, C.T.; Stewart, A.F.; Fulton, S.L.; Zhong, P.; Farrelly, L.A.; Smith, A.C.W.; Ramakrishnan, A.; Lyu, Y.; Bastle, R.M.; et al. Dopaminylation of histone H3 in ventral tegmental area regulates cocaine seeking. Science 2020, 368, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.F.; Lepack, A.E.; Fulton, S.L.; Safovich, P.; Maze, I. Histone H3 dopaminylation in nucleus accumbens, but not medial prefrontal cortex, contributes to cocaine-seeking following prolonged abstinence. Mol. Cell. Neurosci. 2023, 125, 103824. [Google Scholar] [CrossRef] [PubMed]
- Maze, I.; Wenderski, W.; Noh, K.M.; Bagot, R.C.; Tzavaras, N.; Purushothaman, I.; Elsässer, S.J.; Guo, Y.; Ionete, C.; Hurd, Y.L.; et al. Critical role of histone turnover in neuronal transcription and plasticity. Neuron 2015, 87, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Jiao, J. 2017. Histone variant H3.3 orchestrates neural stem cell differentiation in the developing brain. Cell Death Differ. 2017, 24, 1548–1563. [Google Scholar] [CrossRef]
- Chang, W.; Zhao, Y.; Rayee, D.; Xie, Q.; Suzuki, M.; Zheng, D.; Cvekl, A. Dynamic changes in whole genome DNA methylation, chromatin and gene expression during mouse lens differentiation. Epigenetics Chromatin 2023, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Zang, C.; Wei, G.; Cui, K.; Peng, W.; Zhao, K.; Felsenfeld, G. H3.3/H2A.Z double variant-containing nucleosomes mark ‘nucleosome-free regions’ of active promoters and other regulatory regions in the human genome. Nat. Genet. 2009, 41, 941–945. [Google Scholar] [CrossRef]
- Martire, S.; Banaszynski, L.A. The roles of histone variants in fine-tuning chromatin organization and function. Nat. Rev. Mol. Cell Biol. 2020, 21, 522–541. [Google Scholar] [CrossRef]
- Kamada, R.; Yang, W.; Zhang, Y.; Patel, M.C.; Yang, Y.; Ouda, R.; Dey, A.; Wakabayashi, Y.; Sakaguchi, K.; Fujita, T.; et al. Interferon stimulation creates chromatin marks and establishes transcriptional memory. Proc. Natl. Acad. Sci. USA 2018, 115, E9162–E9171. [Google Scholar] [CrossRef]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Cell-to-Cell Communication in Learning and Memory: From Neuro- and Glio-Transmission to Information Exchange Mediated by Extracellular Vesicles. Int. J. Mol. Sci. 2019, 21, 266. [Google Scholar] [CrossRef]
- Levenson, J.M.; O’Riordan, K.J.; Brown, K.D.; Trinh, M.A.; Molfese, D.L.; Sweatt, J.D. Regulation of histone acetylation during memory formation in the hippocampus. J. Biol. Chem. 2004, 279, 40545–40559. [Google Scholar] [CrossRef]
- Ronan, J.L.; Wu, W.; Crabtree, G.R. From neural development to cognition: Unexpected roles for chromatin. Nat. Rev. Genet. 2013, 14, 347–359. [Google Scholar] [CrossRef]
- Bousiges, O.; Vasconcelos, A.P.; Neidl, R.; Cosquer, B.; Herbeaux, K.; Panteleeva, I.; Loeffler, J.P.; Cassel, J.C.; Boutillier, A.L. Spatial memory consolidation is associated with induction of several lysine-acetyltransferase (histone acetyltransferase) expression levels and H2B/H4 acetylation-dependent transcriptional events in the rat hippocampus. Neuropsychopharmacology 2010, 35, 2521–2537. [Google Scholar] [CrossRef]
- McNally, A.G.; Poplawski, S.G.; Mayweather, B.A.; White, K.M.; Abel, T. Characterization of a Novel Chromatin Sorting Tool Reveals Importance of Histone Variant H3.3 in Contextual Fear Memory and Motor Learning. Front. Mol. Neurosci. 2016, 9, 11. [Google Scholar] [CrossRef]
- Singh, G.; Stefanelli, G.; Narkaj, K.; Brimble, M.A.; Creighton, S.D.; McLean, T.A.B.; Hall, M.; Mitchnick, K.A.; Zakaria, J.; Phung, T.; et al. Histone macroH2A1 is a stronger regulator of hippocampal transcription and memory than macroH2A2 in mice. Commun. Biol. 2022, 5, 482. [Google Scholar] [CrossRef]
- Maver, A.; Čuturilo, G.; Ruml, S.J.; Peterlin, B. Clinical next generation sequencing reveals an H3F3A gene as a new potential gene candidate for microcephaly associated with severe developmental delay, intellectual disability and growth retardation. Balk. J. Med. Genet. 2019, 22, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Okur, V.; Chen, Z.; Vossaert, L.; Peacock, S.; Rosenfeld, J.; Zhao, L.; Du, H.; Calamaro, E.; Gerard, A.; Zhao, S.; et al. De novo variants in H3-3A and H3-3B are associated with neurodevelopmental delay, dysmorphic features, and structural brain abnormalities. NPJ Genom Med. 2021, 6, 104. [Google Scholar] [CrossRef]
- Funk, O.H.; Qalieh, Y.; Doyle, D.Z.; Lam, M.M.; Kwan, K.Y. Postmitotic accumulation of histone variant H3.3 in new cortical neurons establishes neuronal chromatin, transcriptome, and identity. Proc. Natl. Acad. Sci. USA 2022, 119, e2116956119. [Google Scholar] [CrossRef] [PubMed]
- Bedogni, F.; Hodge, R.D.; Elsen, G.E.; Nelson, B.R.; Daza, R.A.; Beyer, R.P.; Bammler, T.K.; Rubenstein, J.L.; Hevner, R.F. Tbr1 regulates regional and laminar identity of postmitotic neurons in developing neocortex. Proc. Natl. Acad. Sci. USA 2010, 107, 13129–13134. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Kwan, K.Y.; Shim, S.; Lam, M.M.; Shin, Y.; Xu, X.; Zhu, Y.; Li, M.; Sestan, N. TBR1 directly represses Fezf2 to control the laminar origin and development of the corticospinal tract. Proc. Natl. Acad. Sci. USA 2011, 108, 3041–3046. [Google Scholar] [CrossRef] [PubMed]
- Tsyporin, J.; Tastad, D.; Ma, X.; Nehme, A.; Finn, T.; Huebner, L.; Liu, G.; Gallardo, D.; Makhamreh, A.; Roberts, J.M.; et al. Transcriptional repression by FEZF2 restricts alternative identities of cortical projection neurons. Cell Rep. 2021, 35, 109269. [Google Scholar] [CrossRef]
- Song, C.; Kanthasamy, A.; Anantharam, V.; Sun, F.; Kanthasamy, A.G. Environmental neurotoxic pesticide increases histone acetylation to promote apoptosis in dopaminergic neuronal cells: Relevance to epigenetic mechanisms of neurodegeneration. Mol Pharmacol. 2010, 77, 621–632. [Google Scholar] [CrossRef]
- Huang, M.; Lou, D.; Charli, A.; Kong, D.; Jin, H.; Zenitsky, G.; Anantharam, V.; Kanthasamy, A.; Wang, Z.; Kanthasamy, A.G. Mitochondrial dysfunction-induced H3K27 hyperacetylation perturbs enhancers in Parkinson’s disease. JCI Insight. 2021, 6, e138088. [Google Scholar] [CrossRef] [PubMed]
- Vashishtha, M.; Ng, C.W.; Yildirim, F.; Gipson, T.A.; Kratter, I.H.; Bodai, L.; Song, W.; Lau, A.; Labadorf, A.; Vogel-Ciernia, A.; et al. Targeting H3K4 trimethylation in Huntington disease. Proc. Natl. Acad. Sci. USA 2013, 110, E3027-36. [Google Scholar] [CrossRef]
- Dong, X.; Tsuji, J.; Labadorf, A.; Roussos, P.; Chen, J.F.; Myers, R.H.; Akbarian, S.; Weng, Z. The Role of H3K4me3 in Transcriptional Regulation Is Altered in Huntington’s Disease. PLoS ONE 2015, 10, e0144398. [Google Scholar] [CrossRef]
- Song, W.; Zsindely, N.; Faragó, A.; Marsh, J.L.; Bodai, L. Systematic genetic interaction studies identify histone demethylase Utx as potential target for ameliorating Huntington’s disease. Hum. Mol. Genet. 2018, 27, 649–666, Erratum. Hum Mol Genet.2018, 27, 759. [Google Scholar] [CrossRef] [PubMed]
- Rouaux, C.; Loeffler, J.P.; Boutillier, A.L. Targeting CREB-binding protein (CBP) loss of function as a therapeutic strategy in neurological disorders. Biochem. Pharmacol. 2004, 68, 1157–1164. [Google Scholar] [CrossRef]
- Qian, M.X.; Pang, Y.; Liu, C.H.; Haratake, K.; Du, B.Y.; Ji, D.Y.; Wang, G.F.; Zhu, Q.Q.; Song, W.; Yu, Y.; et al. Acetylation-mediated proteasomal degradation of core histones during DNA repair and spermatogenesis. Cell 2013, 153, 1012–1024. [Google Scholar] [CrossRef] [PubMed]
- Peleg, S.; Feller, C.; Ladurner, A.G.; Imhof, A. The Metabolic Impact on Histone Acetylation and Transcription in Ageing. Trends Biochem. Sci. 2016, 41, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Koldobskiy, M.A.; Göndör, A. Epigenetic modulators, modifiers and mediators in cancer aetiology and progression. Nat. Rev. Genet. 2016, 17, 284–299. [Google Scholar] [CrossRef]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.Y.; Jones, D.T.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.A.; Tönjes, M.; et al. Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, E.; Bae, M.; Kim, S.; Park, J.B.; Yoo, H.; Choi, J.K.; Kim, Y.-J.; Lee, S.-H.; Kim, I.-H. Histone variant H3F3A promotes lung cancer cell migration through intronic regulation. Nat. Commun. 2016, 7, 12914. [Google Scholar] [CrossRef]
- Gomes, A.P.; Ilter, D.; Low, V.; Rosenzweig, A.; Shen, Z.J.; Schild, T.; Rivas, M.A.; Er, E.E.; McNally, D.R.; Mutvei, A.P.; et al. Dynamic Incorporation of Histone H3 Variants into Chromatin Is Essential for Acquisition of Aggressive Traits and Metastatic Colonization. Cancer Cell. 2019, 36, 402–417.e13. [Google Scholar] [CrossRef]
- Huang, X.; Liu, J.; Ma, Q. Prohibitin participates in the HIRA complex to promote cell metastasis in breast cancer cell lines. FEBS Open Bio. 2020, 10, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Broniscer, A.; McEachron, T.A.; Lu, C.; Paugh, B.S.; Becksfort, J.; Qu, C.; Ding, L.; Huether, R.; Parker, M.; et al. St. Jude Children’s Research Hospital–Washington University Pediatric Cancer Genome Project. Somatic histone H3 alterations in pediatric diffuse intrinsic pontine gliomas and non-brainstem glioblastomas. Nat. Genet. 2012, 44, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, F.; Helin, K. Oncohistones: Drivers of pediatric cancers. Genes Dev. 2017, 31, 2313–2324. [Google Scholar] [CrossRef]
- Voon, H.P.J.; Wong, L.H. Chromatin mutations in pediatric high grade gliomas. Front. Oncol. 2023, 12, 1104129. [Google Scholar] [CrossRef]
- Sturm, D.; Bender, S.; Jones, D.T.; Lichter, P.; Grill, J.; Becher, O.; Hawkins, C.; Majewski, J.; Jones, C.; Costello, J.F.; et al. Paediatric and adult glioblastoma: Multiform (epi)genomic culprits emerge. Nat. Rev Cancer. 2014, 14, 92–107. [Google Scholar] [CrossRef]
- Ilter, D.; Drapela, S.; Schild, T.; Ward, N.P.; Adhikari, E.; Low, V.; Asara, J.; Oskarsson, T.; Lau, E.K.; DeNicola, G.M.; et al. NADK-mediated de novo NADP(H) synthesis is a metabolic adaptation essential for breast cancer metastasis. Redox Biol. 2023, 61, 102627. [Google Scholar] [CrossRef]
- Montalbano, S.; Raboni, S.; Sidoli, S.; Mozzarelli, A.; Bettati, S.; Buschini, A. Post-translational modifications of histone variants in the absence and presence of a methionine-depleting enzyme in normal and cancer cells. Cancers 2023, 15, 527. [Google Scholar] [CrossRef]
- Yadav, R.K.; Jablonowski, C.M.; Fernandez, A.G.; Lowe, B.R.; Henry, R.A.; Finkelstein, D.; Barnum, K.J.; Pidoux, A.L.; Kuo, Y.M.; Huang, J.; et al. Histone H3G34R mutation causes replication stress, homologous recombination defects and genomic instability in S. pombe. eLife 2017, 6, e27406. [Google Scholar] [CrossRef] [PubMed]
- Mackay, A.; Burford, A.; Carvalho, D.; Izquierdo, E.; Fazal-Salom, J.; Taylor, K.R.; Bjerke, L.; Clarke, M.; Vinci, M.; Nandhabalan, M.; et al. Integrated Molecular Meta-Analysis of 1,000 Pediatric High-Grade and Diffuse Intrinsic Pontine Glioma. Cancer Cell. 2017, 32, 520–537.e5. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.A.; Klein, R.H.; Kelly, C.; Yee, J.; Knoepfler, P.S. Histone H3.3 K27M chromatin functions implicate a network of neurodevelopmental factors including ASCL1 and NEUROD1 in DIPG. Epigenetics Chromatin. 2022, 15, 18. [Google Scholar] [CrossRef]
- Dalle Ore, C.; Coleman-Abadi, C.; Gupta, N.; Mueller, S. Advances and Clinical Trials Update in the Treatment of Diffuse Intrinsic Pontine Gliomas. Pediatr Neurosurg. 2023. [Google Scholar] [CrossRef]
- Lewis, P.W.; Müller, M.M.; Koletsky, M.S.; Cordero, F.; Lin, S.; Banaszynski, L.A.; Garcia, B.A.; Muir, T.W.; Becher, O.J.; Allis, C.D. Inhibition of PRC2 activity by a gain-of-function H3 mutation found in pediatric glioblastoma. Science 2013, 340, 857–861. [Google Scholar] [CrossRef]
- Justin, N.; Zhang, Y.; Tarricone, C.; Martin, S.R.; Chen, S.; Underwood, E.; De Marco, V.; Haire, L.F.; Walker, P.A.; Reinberg, D.; et al. Structural basis of oncogenic histone H3K27M inhibition of human polycomb repressive complex 2. Nat. Commun. 2016, 7, 11316. [Google Scholar] [CrossRef]
- Sarthy, J.F.; Meers, M.P.; Janssens, D.H.; Henikoff, J.G.; Feldman, H.; Paddison, P.J.; Lockwood, C.M.; Vitanza, N.A.; Olson, J.M.; Ahmad, K.; et al. Histone deposition pathways determine the chromatin landscapes of H3.1 and H3.3 K27M oncohistones. eLife 2020, 9, e61090. [Google Scholar] [CrossRef]
- Sieverling, L.; Hong, C.; Koser, S.D.; Ginsbach, P.; Kleinheinz, K.; Hutter, B.; Braun, D.M.; Cortés-Ciriano, I.; Xi, R.; Kabbe, R.; et al. Genomic footprints of activated telomere maintenance mechanisms in cancer. Nat. Commun. 2020, 11, 733. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Banerjee, K.; Mujeeb, A.A.; Hartlage, C.S.; Núñez, F.M.; Núñez, F.J.; Alghamri, M.S.; Kadiyala, P.; Carney, S.; Barissi, M.N.; et al. H3.3-G34 mutations impair DNA repair and promote cGAS/STING-mediated immune responses in pediatric high-grade glioma models. J. Clin Invest. 2022, 132, e154229. [Google Scholar] [CrossRef] [PubMed]
- Karthik, N.; Lee, J.J.H.; Soon, J.L.J.; Chiu, H.Y.; Loh, A.H.P.; Ong, D.S.T.; Tam, W.L.; Taneja, R. Histone variant H3.3 promotes metastasis in alveolar rhabdomyosarcoma. J. Pathol. 2023, 259, 342–356. [Google Scholar] [CrossRef]
- Cottone, L.; Ligammari, L.; Lee, H.M.; Knowles, H.J.; Henderson, S.; Bianco, S.; Davies, C.; Strauss, S.; Amary, F.; Leite, A.P.; et al. Aberrant paracrine signalling for bone remodelling underlies the mutant histone-driven giant cell tumour of bone. Cell Death Differ. 2022, 29, 2459–2471. [Google Scholar] [CrossRef] [PubMed]
- Behjati, S.; Tarpey, P.S.; Presneau, N.; Scheipl, S.; Pillay, N.; Van Loo, P.; Wedge, D.C.; Cooke, S.L.; Gundem, G.; Davies, H.; et al. Distinct H3F3A and H3F3B driver mutations define chondroblastoma and giant cell tumor of bone. Nat. Genet. 2013, 45, 1479–1482. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.; Gan, H.; Lee, J.H.; Han, J.; Wang, Z.; Riester, S.M.; Jin, L.; Chen, J.; Zhou, H.; Wang, J.; et al. The histone H3.3K36M mutation reprograms the epigenome of chondroblastomas. Science 2016, 352, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Di Liegro, C.M.; Schiera, G.; Schirò, G.; Di Liegro, I. RNA-Binding Proteins as Epigenetic Regulators of Brain Functions and Their Involvement in Neurodegeneration. Int. J. Mol. Sci. 2022, 23, 14622. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).