
Cryopreservation Induces Acetylation of Metabolism-Related Proteins in Boar Sperm

 ,
,
Abstract
1. Introduction
2. Results
2.1. Differentially Expressed Acetylated Proteins and Modified Sites during Sperm Cryopreservation
2.2. GO and KEGG Analysis Indicating the Role of Modified Proteins in Energy Metabolism
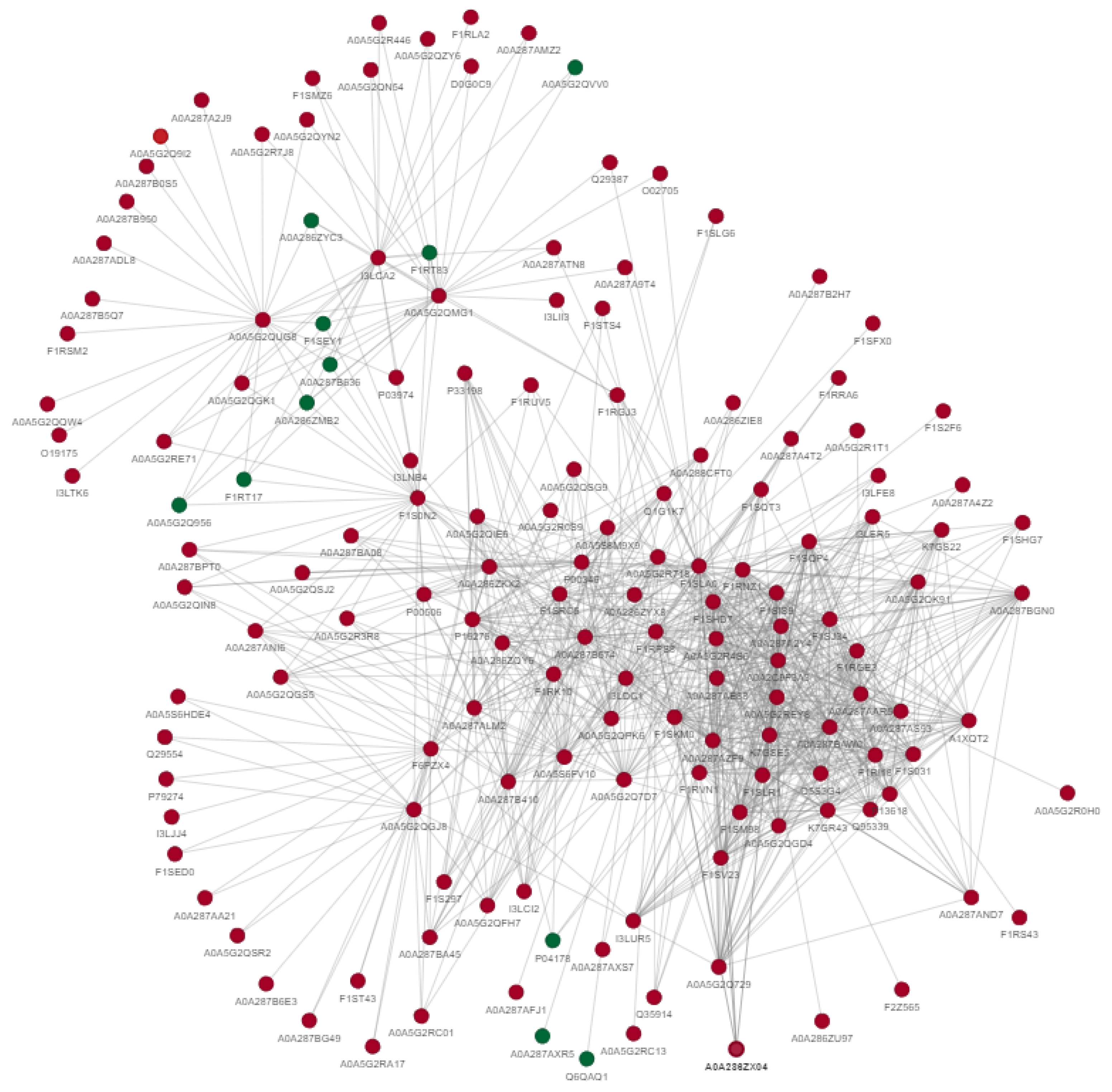
2.3. Protein–Protein Interaction
2.4. mRNA and Protein Expression of IDH2, MDH2, LDHC and SIRT5
2.5. Knockdown of SIRT5 (siSIRT5) Affects Sperm Motility, Acrosomal Integrity and Mitochondrial Membrane Potential (MMP)
3. Discussion
4. Materials and Methods
4.1. Semen Collection and Processing
4.2. RNA Extraction and Quantitative PCR
4.3. Protein Extraction and Western Blotting
4.4. Enzymatic Hydrolysis
4.5. Modification Site Enrichment and Mass Spectrometry Analysis (LC-MS)
4.6. Protein Interaction, Functional Enrichment and Cluster Analysis
4.7. Electrotransfection and Detection of Boar Sperm Quality Parameters
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeste, M.; Rodríguez-Gil, J.E.; Bonet, S. Artificial insemination with frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Flores, E.; Ramió-Lluch, L.; Bucci, D.; Fernández-Novell, J.M.; Peña, A.; Rodríguez-Gil, J.E. Freezing-thawing induces alterations in histone H1-DNA binding and the breaking of protein-DNA disulfide bonds in boar sperm. Theriogenology 2011, 76, 1450–1464. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.C.; Briggs, E.F.; Burnett, R.H.; Contreras-Correa, Z.E.; Duvic, M.A.; Dysart, L.M.; Gilmore, A.A.; Messman, R.D.; Reid, D.; Rasit Ugur, M.; et al. Harnessing the value of reproductive hormones in cattle production with considerations to animal welfare and human health. J. Anim. Sci. 2022, 100, skac177. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Recent Advances in Boar Sperm Cryopreservation: State of the Art and Current Perspectives. Reprod. Domest. Anim. 2015, 50 (Suppl. 2), 71–79. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.J.; Kim, Y.J. Changes in sperm membrane and ROS following cryopreservation of liquid boar semen stored at 15 °C. Anim. Reprod. Sci. 2011, 124, 118–124. [Google Scholar] [CrossRef]
- Ali, M.A.; Wang, Y.; Qin, Z.; Yuan, X.; Zhang, Y.; Zeng, C. Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility. Genes 2021, 12, 488. [Google Scholar] [CrossRef]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. Biomed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef]
- Martins, A.; Agarwal, A.; Henkel, R. Sperm Cryopreservation. In Vitro Fertilization; Springer: Berlin/Heidelberg, Germany, 2019; pp. 625–642. [Google Scholar] [CrossRef]
- Cerolini, S.; Maldjian, A.; Surai, P.; Noble, R. Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage. Anim. Reprod. Sci. 2000, 58, 99–111. [Google Scholar] [CrossRef]
- Bailey, J.L.; Lessard, C.; Jacques, J.; Brèque, C.; Dobrinski, I.; Zeng, W.; Galantino-Homer, H.L. Cryopreservation of boar semen and its future importance to the industry. Theriogenology 2008, 70, 1251–1259. [Google Scholar] [CrossRef]
- Simonik, O.; Bubenickova, F.; Tumova, L.; Frolikova, M.; Sur, V.P.; Beran, J.; Havlikova, K.; Hackerova, L.; Spevakova, D.; Komrskova, K.; et al. Boar Sperm Cryopreservation Improvement Using Semen Extender Modification by Dextran and Pentaisomaltose. Animals 2022, 12, 868. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Y.; Ali, M.A.; Zhang, J.; Wang, W.; Huang, Y.; Luo, B.; Zhang, H.; Qin, Z.; Zhang, Y.; et al. Comparative Analysis of piRNA Profiles Helps to Elucidate Cryoinjury Between Giant Panda and Boar Sperm During Cryopreservation. Front. Vet. Sci. 2021, 8, 635013. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, A.; Zhou, H.; Sun, W.; Ding, R.; Li, X.; Liu, J.; Zhou, Y.; Chen, X.; Ding, F.; Yang, L.; et al. Cryopreservation Induces Alterations of miRNA and mRNA Fragment Profiles of Bull Sperm. Front. Genet. 2020, 11, 419. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.; Jodar, M.; Oliva, R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef]
- Guimarães, D.B.; Barros, T.B.; van Tilburg, M.F.; Martins, J.A.M.; Moura, A.A.; Moreno, F.B.; Monteiro-Moreira, A.C.; Moreira, R.A.; Toniolli, R. Sperm membrane proteins associated with the boar semen cryopreservation. Anim. Reprod. Sci. 2017, 183, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Maciel, V.L., Jr.; Tamashiro, L.K.; Bertolla, R.P. Post-translational modifications of seminal proteins and their importance in male fertility potential. Expert Rev. Proteom. 2019, 16, 941–950. [Google Scholar] [CrossRef]
- Khosravizadeh, Z.; Khodamoradi, K.; Rashidi, Z.; Jahromi, M.; Shiri, E.; Salehi, E.; Talebi, A. Sperm cryopreservation and DNA methylation: Possible implications for ART success and the health of offspring. J. Assist. Reprod. Genet. 2022, 39, 1815–1824. [Google Scholar] [CrossRef]
- Qin, Z.; Wang, W.; Ali, M.A.; Wang, Y.; Zhang, Y.; Zhang, M.; Zhou, G.; Yang, J.; Zeng, C.J.B.G. Transcriptome-wide m6A profiling reveals mRNA post-transcriptional modification of boar sperm during cryopreservation. BMC Genom. 2021, 22, 588. [Google Scholar] [CrossRef]
- Parrilla, I.; Perez-Patiño, C.; Li, J.; Barranco, I.; Padilla, L.; Rodriguez-Martinez, H.; Martinez, E.A.; Roca, J. Boar semen proteomics and sperm preservation. Theriogenology 2019, 137, 23–29. [Google Scholar] [CrossRef]
- Purdy, P.H. Ubiquitination and its influence in boar sperm physiology and cryopreservation. Theriogenology 2008, 70, 818–826. [Google Scholar] [CrossRef]
- Kuo, Y.W.; Li, S.H.; Maeda, K.; Gadella, B.M.; Tsai, P.S. Roles of the reproductive tract in modifications of the sperm membrane surface. J. Reprod. Dev. 2016, 62, 337–343. [Google Scholar] [CrossRef]
- Samanta, L.; Swain, N.; Ayaz, A.; Venugopal, V.; Agarwal, A. Post-Translational Modifications in sperm Proteome: The Chemistry of Proteome diversifications in the Pathophysiology of male factor infertility. Biochim. Biophys. Acta 2016, 1860, 1450–1465. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.; Park, S.; Oh, S.; Choi, J.; Kim, E.-K.; Youn, H.-D.; Cho, E.-J. Histone acylation marks respond to metabolic perturbations and enable cellular adaptation. Exp. Mol. Med. 2020, 52, 2005–2019. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tu, B.P. Acetyl-CoA and the regulation of metabolism: Mechanisms and consequences. Curr. Opin. Cell Biol. 2015, 33, 125–131. [Google Scholar] [CrossRef]
- Moreno-Sánchez, R.; Gallardo-Pérez, J.C.; Pacheco-Velazquez, S.C.; Robledo-Cadena, D.X.; Rodríguez-Enríquez, S.; Encalada, R.; Saavedra, E.; Marín-Hernández, Á. Regulatory role of acetylation on enzyme activity and fluxes of energy metabolism pathways. Biochim. Biophys. Acta. Gen. Subj. 2021, 1865, 130021. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xia, S.; Zhang, R.; Li, Y.; Famulare, C.A.; Fan, H.; Wu, R.; Wang, M.; Zhu, A.C.; Elf, S.E.; et al. Lysine acetylation restricts mutant IDH2 activity to optimize transformation in AML cells. Mol. Cell 2021, 81, 3833–3847.e11. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.; Sun, R.; Tao, X.; Wang, X.; Kang, Q.; Wang, H.; Zhang, L.; Liu, P.; Zhang, J.; et al. SIRT5 deficiency suppresses mitochondrial ATP production and promotes AMPK activation in response to energy stress. PLoS ONE 2019, 14, e0211796. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, L.; Nakamura, A.; Someya, S.; Miyakawa, T.; Tanokura, M. Studies on the regulatory mechanism of isocitrate dehydrogenase 2 using acetylation mimics. Sci. Rep. 2017, 7, 9785. [Google Scholar] [CrossRef]
- Smolková, K.; Špačková, J.; Gotvaldová, K.; Dvořák, A.; Křenková, A.; Hubálek, M.; Holendová, B.; Vítek, L.; Ježek, P. SIRT3 and GCN5L regulation of NADP+- and NADPH-driven reactions of mitochondrial isocitrate dehydrogenase IDH2. Sci. Rep. 2020, 10, 8677. [Google Scholar] [CrossRef]
- Tamura, I.; Fujimura, T.; Doi-Tanaka, Y.; Takagi, H.; Shirafuta, Y.; Kajimura, T.; Mihara, Y.; Maekawa, R.; Taketani, T.; Sato, S.; et al. The essential glucose transporter GLUT1 is epigenetically upregulated by C/EBPβ and WT1 during decidualization of the endometrium. J. Biol. Chem. 2021, 297, 101150. [Google Scholar] [CrossRef]
- Bringman-Rodenbarger, L.R.; Guo, A.H.; Lyssiotis, C.A.; Lombard, D.B. Emerging Roles for SIRT5 in Metabolism and Cancer. Antioxid. Redox Signal. 2018, 28, 677–690. [Google Scholar] [CrossRef]
- Yadav, N.; Bhagat, S.; Singh, S. Surface modification of metal oxide nanoparticles to realize biological applications. In Reference Module in Materials Science and Materials Engineering; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Samanta, L.; Agarwal, A. Functional analysis of differentially expressed acetylated spermatozoal proteins in infertile men with unilateral and bilateral varicocele. Int. J. Mol. Sci. 2020, 21, 3155. [Google Scholar] [CrossRef]
- Salehi, M.; Mahdavi, A.H.; Sharafi, M.; Shahverdi, A.J.T. Cryopreservation of rooster semen: Evidence for the epigenetic modifications of thawed sperm. Theriogenology 2020, 142, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ren, L.; Chang, Z.; Zhao, Y.; Zhang, Y.; Xia, D.; Zhao, R.; He, B. Lysine acetylation participates in boar spermatozoa motility and acrosome status regulation under different glucose conditions. Theriogenology 2021, 159, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M. Chapter Five—Site-Specific Quantification of Lysine Acetylation Using Isotopic Labeling. In Methods in Enzymology; Shukla, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 586, pp. 85–95. [Google Scholar]
- Sun, G.; Jiang, M.; Zhou, T.; Guo, Y.; Cui, Y.; Guo, X.; Sha, J. Insights into the lysine acetylproteome of human sperm. J. Proteom. 2014, 109, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Zhuang, X.J.; Wei, Y.M.; Zhang, M.; Lu, S.S.; Lu, Y.; Yang, X.G.; Lu, K.H. Comparison of Mitochondrial Function in Boar and Bull Spermatozoa Throughout Cryopreservation Based on JC-1 Staining. Cryo Lett. 2017, 38, 75–79. [Google Scholar]
- Sun, W.; Jiang, S.; Su, J.; Zhang, J.; Bao, X.; Ding, R.; Shi, P.; Li, S.; Wu, C.; Zhao, G.; et al. The effects of cryopreservation on the acrosome structure, enzyme activity, motility, and fertility of bovine, ovine, and goat sperm. Anim. Reprod. 2021, 17, e20200219. [Google Scholar] [CrossRef]
- Valcarce, D.G.; Cartón-García, F.; Herráez, M.P.; Robles, V. Effect of cryopreservation on human sperm messenger RNAs crucial for fertilization and early embryo development. Cryobiology 2013, 67, 84–90. [Google Scholar] [CrossRef]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A preliminary study on epigenetic changes during boar spermatozoa cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef]
- Kläver, R.; Bleiziffer, A.; Redmann, K.; Mallidis, C.; Kliesch, S.; Gromoll, J. Routine cryopreservation of spermatozoa is safe--evidence from the DNA methylation pattern of nine spermatozoa genes. J. Assist. Reprod. Genet. 2012, 29, 943–950. [Google Scholar] [CrossRef]
- Bowker, Z.; Goldstein, S.; Breitbart, H. Protein acetylation protects sperm from spontaneous acrosome reaction. Theriogenology 2022, 191, 231–238. [Google Scholar] [CrossRef]
- Fu, J.; Li, Y.; Wang, L.; Zhen, L.; Yang, Q.; Li, P.; Li, X. Bovine serum albumin and skim-milk improve boar sperm motility by enhancing energy metabolism and protein modifications during liquid storage at 17 °C. Theriogenology 2017, 102, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Guan, X.; Jia, X.; Li, H.; Chen, R.; Lu, Y. In-depth Profiling and Quantification of the Lysine Acetylome in Hepatocellular Carcinoma with a Trapped Ion Mobility Mass Spectrometer. Mol. Cell. Proteom. MCP 2022, 21, 100255. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, U.; Galindo, A.N.; Thevenet, J.; Hermant, A.; Bermont, F.; Lassueur, S.; Domingo, J.S.; Kussmann, M.; Dayon, L.; Wiederkehr, A. Mitochondrial lysine deacetylation promotes energy metabolism and calcium signaling in insulin-secreting cells. FASEB J. 2019, 33, 4660–4674. [Google Scholar] [CrossRef]
- Naik, A.; Decock, J. Targeting of lactate dehydrogenase C dysregulates the cell cycle and sensitizes breast cancer cells to DNA damage response targeted therapy. Mol. Oncol. 2022, 16, 885–903. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, Z.; Xu, J.; Wu, H.; Cai, S.; He, Y. The prognostic value of lactate dehydrogenase levels in colorectal cancer: A meta-analysis. BMC Cancer 2016, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, Y.; Lin, Y.; Lan, F.; Cui, Z. Cancer-testis antigen lactate dehydrogenase C4 as a novel biomarker of male infertility and cancer. Front. Oncol. 2022, 12, 936767. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Diao, H.; Wang, C.; Lin, Y.; Yu, F.; Lu, H.; Xu, W.; Li, Z.; Shi, H.; Zhao, S.; et al. Acetylproteomic Analysis Reveals Functional Implications of Lysine Acetylation in Human Spermatozoa (sperm)*[S]. Mol. Cell. Proteom. 2015, 14, 1009–1023. [Google Scholar] [CrossRef]
- Goldberg, E. The sperm-specific form of lactate dehydrogenase is required for fertility and is an attractive target for male contraception (a review). Biol. Reprod. 2021, 104, 521–526. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin Inhibits Formation of Mitochondrial Permeability Transition Pores and Improves Oxidative Phosphorylation of Frozen-Thawed Ram Sperm. Front. Endocrinol. 2020, 10, 896. [Google Scholar] [CrossRef]
- Lambhate, S.; Bhattacharjee, D.; Jain, N. APC/C CDH1 ubiquitinates IDH2 contributing to ROS increase in mitosis. Cell. Signal. 2021, 86, 110087. [Google Scholar] [CrossRef]
- Bisht, S.; Dada, R. Oxidative stress: Major executioner in disease pathology, role in sperm DNA damage and preventive strategies. Front. Biosci. 2017, 9, 420–447. [Google Scholar]
- Wang, X.; Yin, L.; Wen, Y.; Yuan, S. Mitochondrial regulation during male germ cell development. Cell. Mol. Life Sci. 2022, 79, 91. [Google Scholar] [CrossRef]
- Novak, S.; Ruiz-Sánchez, A.; Dixon, W.T.; Foxcroft, G.R.; Dyck, M.K. Seminal plasma proteins as potential markers of relative fertility in boars. J. Androl. 2010, 31, 188–200. [Google Scholar] [CrossRef]
- Pérez-Patiño, C.; Parrilla, I.; Barranco, I.; Vergara-Barberán, M.; Simó-Alfonso, E.F.; Herrero-Martínez, J.M.; Rodriguez-Martínez, H.; Martínez, E.A.; Roca, J. New In-Depth Analytical Approach of the Porcine Seminal Plasma Proteome Reveals Potential Fertility Biomarkers. J. Proteome Res. 2018, 17, 1065–1076. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Roberts, K.P. Seminal plasma proteins inhibit in vitro- and cooling-induced capacitation in boar spermatozoa. Reprod. Fertil. Dev. 2010, 22, 893–900. [Google Scholar] [CrossRef]
- Sutovsky, P.; Kerns, K.; Zigo, M.; Zuidema, D. Boar semen improvement through sperm capacitation management, with emphasis on zinc ion homeostasis. Theriogenology 2019, 137, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, H.; Hu, C.; Hao, H.; Zhang, J.; Li, K.; Zhao, X.; Qin, T.; Zhao, K.; Zhu, H.; et al. Identification of differentially expressed proteins in fresh and frozen-thawed boar spermatozoa by iTRAQ-coupled 2D LC-MS/MS. Reproduction 2014, 147, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Jobim, M.I.M.; Oberst, E.R.; Salbego, C.G.; Souza, D.O.; Wald, V.B.; Tramontina, F.; Mattos, R.C. Two-dimensional polyacrylamide gel electrophoresis of bovine seminal plasma proteins and their relation with semen freezability. Theriogenology 2004, 61, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Killian, G.J.; Chapman, D.A.; Rogowski, L.A. Fertility-associated proteins in Holstein bull seminal plasma. Biol. Reprod. 1993, 49, 1202–1207. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, H.; Wang, Y.; Zhang, Z.; Wu, Q. Comprehensive metabolic profiles of seminal plasma with different forms of male infertility and their correlation with sperm parameters. J. Pharm. Biomed. Anal. 2020, 177, 112888. [Google Scholar] [CrossRef]
- Iida-Norita, R.; Miyata, H.; Kaneda, Y.; Emori, C.; Noda, T.; Nakagawa, T.; Matzuk, M.M.; Ikawa, M. Generation of humanized LDHC knock-in mice as a tool to assess human LDHC-targeting contraceptive drugs. Andrology 2022, 11, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.-S.; Rahman, M.S.; Lee, J.-S.; Yoon, S.-J.; Park, Y.-J.; Pang, M.-G. Discovery of Predictive Biomarkers for Litter Size in Boar Spermatozoa. Mol. Cell. Proteom. 2015, 14, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, F.; La Vignera, S.; Cannarella, R.; Mongioì, L.M.; Garofalo, V.; Leanza, C.; Marino, M.; Calogero, A.E.; Condorelli, R.A. Obesity and Male Reproduction: Do Sirtuins Play a Role? Int. J. Mol. Sci. 2022, 23, 973. [Google Scholar] [CrossRef] [PubMed]
- Bello, J.H.; Khan, M.J.; Amir, S.; Kakakhel, H.G.; Tahir, F.; Sultan, S.; Raza, S.Q.; Mamoulakis, C.; Zachariou, A.; Tsatsakis, A.; et al. Dysregulation of mitochondrial sirtuin genes is associated with human male infertility. Andrologia 2022, 54, e14274. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Cantó, C. Mitochondrial Post-translational Modifications and Metabolic Control: Sirtuins and Beyond. Curr. Diabetes Rev. 2017, 13, 338–351. [Google Scholar] [CrossRef]
- Ketema, E.B.; Lopaschuk, G.D. Post-translational Acetylation Control of Cardiac Energy Metabolism. Front. Cardiovasc. Med. 2021, 8, 723996. [Google Scholar] [CrossRef]
- Peng, Y.; Liu, H.; Liu, J.; Long, J. Post-translational modifications on mitochondrial metabolic enzymes in cancer. Free Radic. Biol. Med. 2022, 179, 11–23. [Google Scholar] [CrossRef]
- Wiese, E.; Hitosugi, T. Tyrosine Kinase Signaling in Cancer Metabolism: PKM2 Paradox in the Warburg Effect. Front. Cell Dev. Biol. 2018, 6, 79. [Google Scholar] [CrossRef]
- Chen, J.; Cao, L.; Li, Z.; Li, Y. SIRT1 promotes GLUT1 expression and bladder cancer progression via regulation of glucose uptake. Hum. Cell 2019, 32, 193–201. [Google Scholar] [CrossRef]
- Dai, M.; Wang, L.; Yang, J.; Chen, J.; Dou, X.; Chen, R.; Ge, Y.; Lin, Y. LDHA as a regulator of T cell fate and its mechanisms in disease. Biomed. Pharmacother. 2023, 158, 114164. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, R.; Priyadarshana, C.; Tajima, A.; Travis, A.J.; Asano, A. Localisation and function of glucose transporter GLUT1 in chicken (Gallus gallus domesticus) spermatozoa: Relationship between ATP production pathways and flagellar motility. Reprod. Fertil. Dev. 2020, 32, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liang, K.; Chang, Y.; Ran, M.; Zhang, Y.; Ali, M.A.; Dai, D.; Qazi, I.H.; Zhang, M.; Zhou, G.; et al. miR-26a is Involved in Glycometabolism and Affects Boar Sperm Viability by Targeting PDHX. Cells 2020, 9, 146. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Mei, C.; Chen, Y.; Guo, L.; Yu, Y.; Huang, D. C-type natriuretic peptide regulates sperm capacitation by the cGMP/PKG signalling pathway via Ca(2+) influx and tyrosine phosphorylation. Reprod. Biomed. Online 2019, 38, 289–299. [Google Scholar] [CrossRef]
- Gómez-Torres, M.J.; Medrano, L.; Romero, A.; Fernández-Colom, P.J.; Aizpurúa, J. Effectiveness of human spermatozoa biomarkers as indicators of structural damage during cryopreservation. Cryobiology 2017, 78, 90–94. [Google Scholar] [CrossRef]
- Grevengoed, T.J.; Klett, E.L.; Coleman, R.A. Acyl-CoA metabolism and partitioning. Annu. Rev. Nutr. 2014, 34, 1–30. [Google Scholar] [CrossRef]
- Vucic, D.; Dixit, V.M.; Wertz, I.E. Ubiquitylation in apoptosis: A post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 2011, 12, 439–452. [Google Scholar] [CrossRef]
- Yi, C.H.; Pan, H.; Seebacher, J.; Jang, I.H.; Hyberts, S.G.; Heffron, G.J.; Vander Heiden, M.G.; Yang, R.; Li, F.; Locasale, J.W.; et al. Metabolic regulation of protein N-alpha-acetylation by Bcl-xL promotes cell survival. Cell 2011, 146, 607–620. [Google Scholar] [CrossRef]
- Ran, M.-X.; Zhou, Y.-M.; Liang, K.; Wang, W.-C.; Zhang, Y.; Zhang, M.; Yang, J.-D.; Zhou, G.-B.; Wu, K.; Wang, C.-D.; et al. Comparative Analysis of MicroRNA and mRNA Profiles of Sperm with Different Freeze Tolerance Capacities in Boar (Sus scrofa) and Giant Panda (Ailuropoda melanoleuca). Biomolecules 2019, 9, 432. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, X.; Ali, M.A.; Qin, Z.; Zhang, Y.; Zeng, C. piR-121380 Is Involved in Cryo-Capacitation and Regulates Post-Thawed Boar Sperm Quality Through Phosphorylation of ERK2 via Targeting PTPN7. Front. Cell Dev. Biol. 2022, 9, 792994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xuan, Z.; Li, H.-X.; Cui, Q.-W.; Jing, Y.; Wang, G.-L. Delivery of CatSper2 siRNA into rat sperms by electroporation repressed Ca2+ influx during sperm hyperactivation. Agric. Sci. China 2011, 10, 1958–1967. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Spectrums | Matched Spectrums | Peptides | Modified Peptides | Identified Proteins | Quantifiable Proteins | Identified Sites | Quantifiable Sites |
|---|---|---|---|---|---|---|---|
| 155,547 | 33,829 | 7640 | 4567 | 1440 | 977 | 4705 | 2764 |
| Protein | Acetylation Sites | Fold Change (Fts/Fs) | Acetylation Status | Function |
|---|---|---|---|---|
| PC | 999, 261, 79 | 33.147 | Up | TCA cycle, metabolic pathways |
| ENO1 | 239, 335, 71, 228, 233, 420 | 5.487 | Up | Glycolysis, metabolic pathways |
| PGAM2 | 100 | 0.807 | Down | Glycolysis, metabolic pathways |
| GAPDHS | 49, 286, 147, 130, 115, 124 | 12.66 | Up | Glycolysis, spermatogenesis, metabolic pathway |
| SIRT5 | 50, 150, 161 | 11.483 | Up | Metabolic pathways, oxidation–reduction pathways, TCA cycle |
| IDH2 | 241, 244, 124, 232, 168 | 13.745 | Up | Carbohydrate metabolism, organic acid metabolism, TCA cycle |
| MDH2 | 185, 328, 329, 335, 78, 301 | 21.913 | Up | Metabolic pathways, TCA cycle |
| LDHC | 228, 232, 81, 224, 217 | 9.568 | Up | Metabolic pathways |
| GPX4 | 58, 132, 145, 172 | 36.776 | Up | Glutathione metabolism, metabolic pathways |
| GPD2 | 607, 95, 410, 198, 114 | 15.156 | Up | Carbohydrate metabolism |
| Gene | Sequence (5′-3′) | Tm (°C) | Size (bp) |
|---|---|---|---|
| GAPDH | F: ACCCAGAAGACTGTGGATGG R: CATGGCCTCCAAGGAGTAAG | 60.07 | 346 |
| IDH2 | F: GGGCCTGCAAGAACTACGAT R: CAGGGGACCCTGCAATGAC | 60.11 | 191 |
| MDH2 | F: TCCCGTAACACACAGACAGC R: GCGTTGGTGTTGAACAGGTC | 59.97 | 117 |
| LDHC | F: CCTTCACTGCTCACGTTTGG R: CGCCAATCCCTTTTCACCAC | 58.53 | 218 |
| SIRT5 | F: CACCCCTCCAGCTTACCAAG R: TGTGTCCATCCTGTAATGTCGG | 60.04 | 325 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.A.; Qin, Z.; Dou, S.; Huang, A.; Wang, Y.; Yuan, X.; Zhang, Y.; Ni, Q.; Azmat, R.; Zeng, C. Cryopreservation Induces Acetylation of Metabolism-Related Proteins in Boar Sperm. Int. J. Mol. Sci. 2023, 24, 10983. https://doi.org/10.3390/ijms241310983
Ali MA, Qin Z, Dou S, Huang A, Wang Y, Yuan X, Zhang Y, Ni Q, Azmat R, Zeng C. Cryopreservation Induces Acetylation of Metabolism-Related Proteins in Boar Sperm. International Journal of Molecular Sciences. 2023; 24(13):10983. https://doi.org/10.3390/ijms241310983
Chicago/Turabian StyleAli, Malik Ahsan, Ziyue Qin, Shan Dou, Anqi Huang, Yihan Wang, Xiang Yuan, Yan Zhang, Qingyong Ni, Rameesha Azmat, and Changjun Zeng. 2023. "Cryopreservation Induces Acetylation of Metabolism-Related Proteins in Boar Sperm" International Journal of Molecular Sciences 24, no. 13: 10983. https://doi.org/10.3390/ijms241310983
APA StyleAli, M. A., Qin, Z., Dou, S., Huang, A., Wang, Y., Yuan, X., Zhang, Y., Ni, Q., Azmat, R., & Zeng, C. (2023). Cryopreservation Induces Acetylation of Metabolism-Related Proteins in Boar Sperm. International Journal of Molecular Sciences, 24(13), 10983. https://doi.org/10.3390/ijms241310983