

Total Antioxidant Capacity: Biochemical Aspects and Clinical Significance

 ,
, 

Abstract
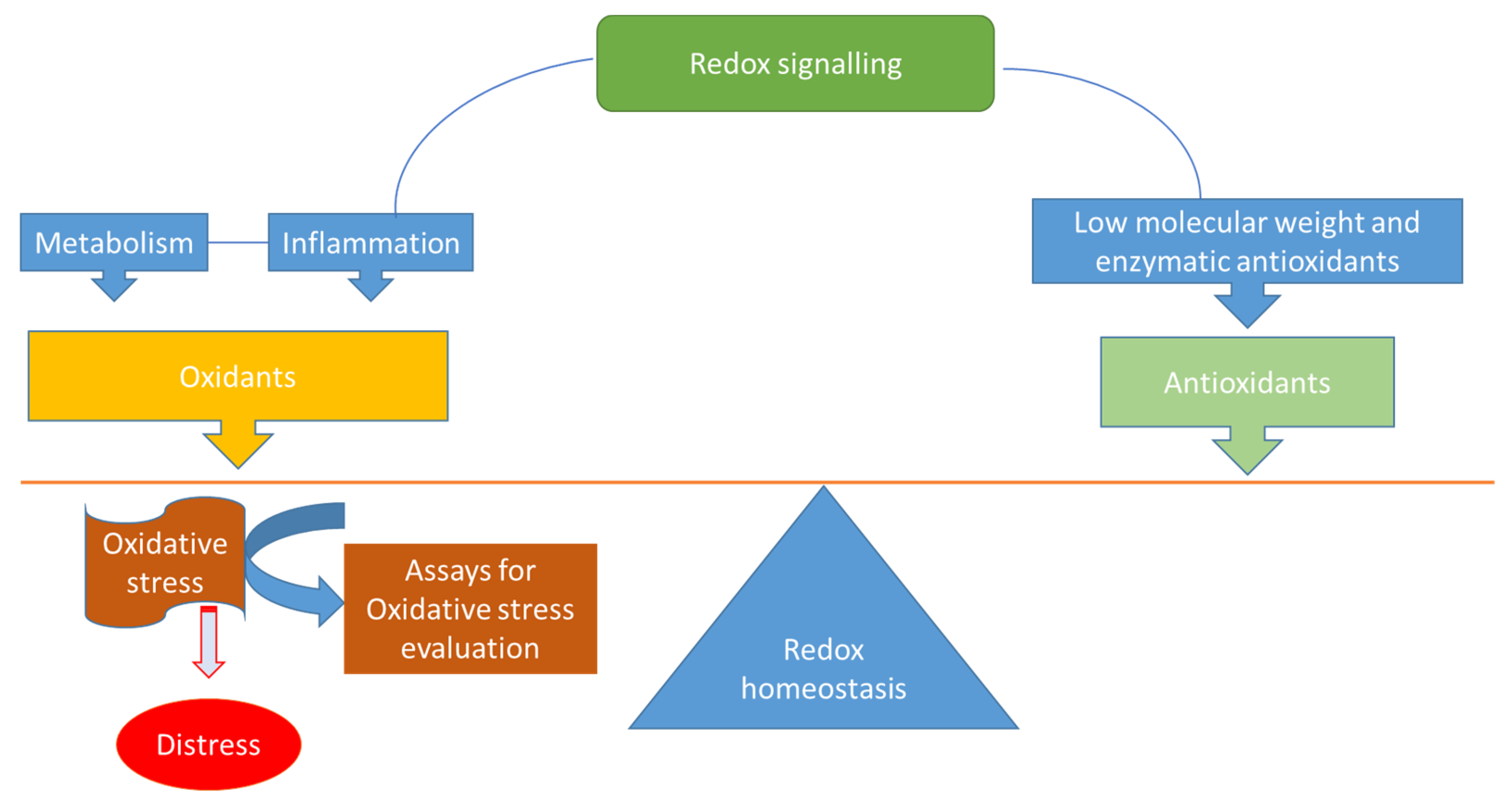
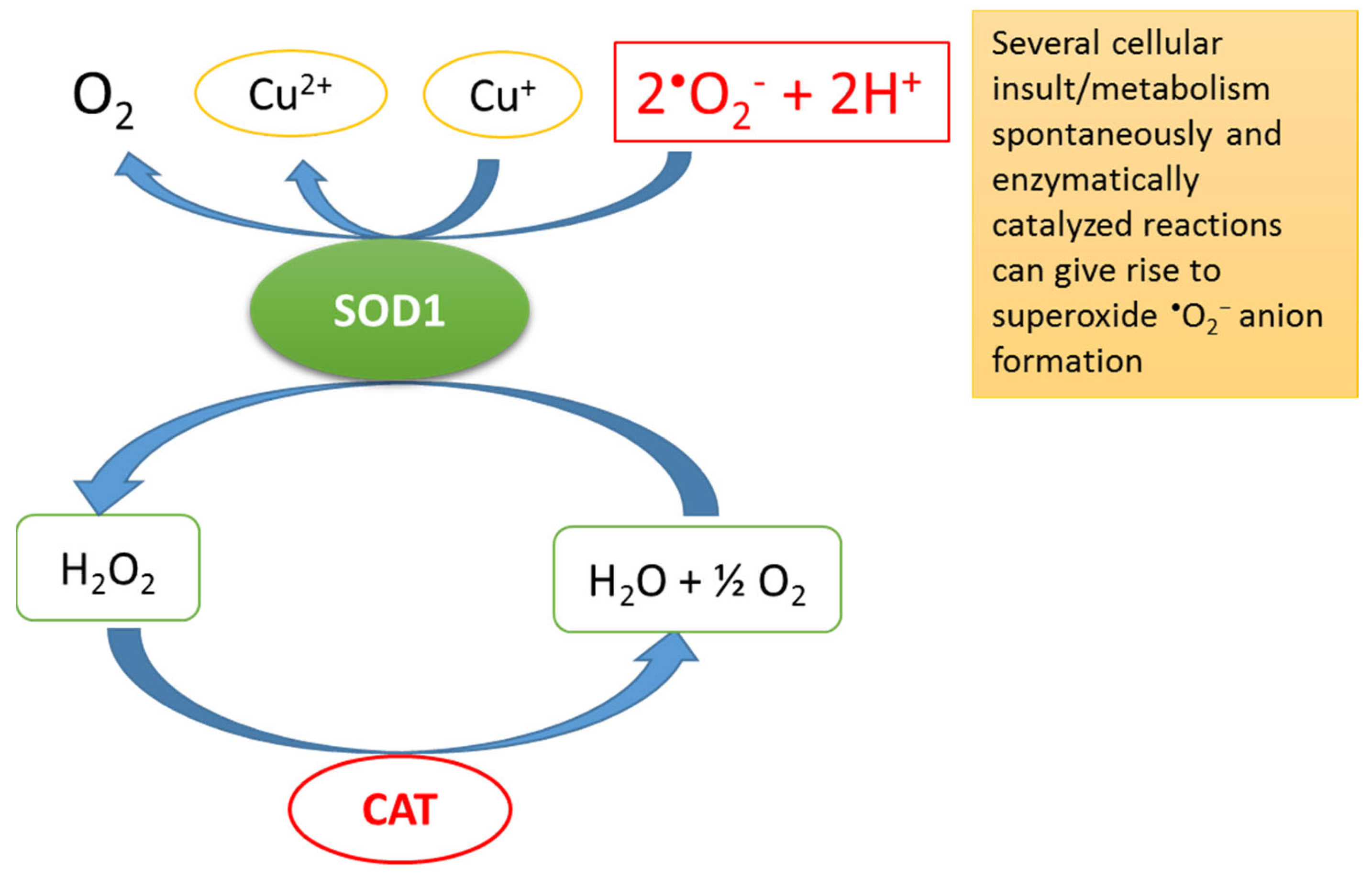
1. Redox Homeostasis, Oxidative Stress (OS) and Total Antioxidant Capacity (TAC)
2. Assays
2.1. The TEAC Assay
2.2. The ORAC Assay
2.3. The FRAP Assay
2.4. The TRAP Assay
3. Perspective on Future Clinical Implication and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Antunes, F.; Brito, P.M. Quantitative biology of hydrogen peroxide signaling. Redox Biol. 2017, 13, 1–7. [Google Scholar] [PubMed]
- Poljsak, B.; Suput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Rosa, A.C.; Corsi, D.; Cavi, N.; Bruni, N.; Dosio, F. Superoxide Dismutase Administration: A Review of Proposed Human Uses. Molecules 2021, 26, 1844. [Google Scholar] [CrossRef]
- Pillay, C.S.; Eagling, B.D.; Driscoll, S.R.; Rohwer, J.M. Quantitative measures for redox signaling. Free Radic. Biol. Med. 2016, 96, 290–303. [Google Scholar]
- Ferrari, C.K. Effects of xenobiotics on total antioxidant capacity. Interdiscip. Toxicol. 2012, 5, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Arts, M.J.; Haenen, G.R.; Voss, H.P.; Bast, A. Antioxidant capacity of reaction products limits the applicability of the Trolox Equivalent Antioxidant Capacity (TEAC) assay. Food Chem. Toxicol. 2004, 42, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.J. Total antioxidant status in plasma and body fluids. Methods Enzymol. 1994, 234, 279–293. [Google Scholar] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Leone, E.; Festa, R.; Grande, G.; Di Donna, V.; De Marinis, L.; Pontecorvi, A.; Tacchino, R.M.; Littarru, G.P.; Silvestrini, A.; et al. Evaluation of antioxidant systems (coenzyme Q10 and total antioxidant capacity) in morbid obesity before and after biliopancreatic diversion. Metab. Clin. Exp. 2008, 57, 1384–1389. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Cadenas, E. Inhibition of protein radical reactions of ferrylmyoglobin by the water-soluble analog of vitamin E, Trolox C. Arch. Biochem. Biophys. 1993, 303, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Maestre, A.B.; Hernández-Ruiz, J.; Arnao, M.B. ABTS/TAC Methodology: Main Milestones and Recent Applications. Processes 2023, 11, 185. [Google Scholar] [CrossRef]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef]
- Huang, D.; Boxin, O.U.; Prior, P.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 55, 1841–1856. [Google Scholar] [CrossRef]
- Gunawardena, H.; Silva, R.; Ranasinghe, P. Human plasma dynamically quenches the fluorescein at the initial point of oxygen radical absorption capacity (ORAC) assay. BMC Res. Notes 2019, 12, 809. [Google Scholar] [CrossRef] [PubMed]
- Price, J.A.; Sanny, C.G.; Shevlin, D. Application of manual assessment of oxygen radical absorbent capacity (ORAC) for use in high throughput assay of “total” antioxidant activity of drugs and natural products. J. Pharmacol. Toxicol. Methods 2006, 54, 56–61. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef] [PubMed]
- Fejér, J.; Kron, I.; Eliašová, A.; Gruľová, D.; Gajdošová, A.; Lancíková, V.; Hricová, A. New Mutant Amaranth Varieties as a Potential Source of Biologically Active Substances. Antioxidants 2021, 10, 1705. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Çelik, S.E. Mechanism of antioxidant capacity assays and the CUPRAC (cupric ion reducing antioxidant capacity) assay. Microchim. Acta 2008, 160, 413–419. [Google Scholar] [CrossRef]
- Marques, S.S.; Magalhães, L.M.; Tóth, I.V.; Segundo, M.A. Insights on Antioxidant Assays for Biological Samples Based on the Reduction of Copper Complexes—The Importance of Analytical Conditions. Int. J. Mol. Sci. 2014, 15, 11387–11402. [Google Scholar] [CrossRef]
- Bener, M.; Ozyürek, M.; Güçlü, K.; Apak, R. Novel optical fiber reflectometric CUPRAC sensor for total antioxidant capacity measurement of food extracts and biological samples. J. Agric. Food Chem. 2013, 61, 8381–8388. [Google Scholar] [CrossRef]
- Schlesier, K.; Harwat, M.; Bohm, V.; Bitsch, R. Assessment of antioxidant activity by using different in vitro methods. Free Radic. Res. 2002, 36, 177–187. [Google Scholar] [CrossRef]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 2. Hydrogen atom transfer (HAT)-based, mixed-mode (electron transfer (ET)/HAT), and lipid peroxidation assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef]
- Lissi, E.; Salim-Hanna, M.; Pascual, C.; del Castillo, M.D. Evaluation of total antioxidant potential (TRAP) and total antioxidant reactivity from luminol-enhanced chemiluminescence measurements. Free Radic. Biol. Med. 1995, 18, 153–158. [Google Scholar] [CrossRef]
- Kharb, S. Total free radical trapping antioxidant potential in pre-eclampsia. Int. J. Gynaecol. Obstet. 2000, 69, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Leo, F.; Rossodivita, A.N.; Di Segni, C.; Raimondo, S.; Canichella, S.; Silvestrini, A.; Miggiano, G.A.D.; Meucci, E.; Mancini, A. Frailty of obese children: Evaluation of plasma antioxidant capacity in pediatric obesity. Exp. Clin. Endocrinol. Diabetes 2016, 124, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 2, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, C.V.; Sales, M.M.; Rosa, T.S.; Lewis, J.E.; de Andrade, R.V.; Simoes, H.G. The Antioxidant Effect of Exercise: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 277–293. [Google Scholar] [CrossRef]
- Mancini, A.; Festa, R.; Di Donna, V.; Leone, E.; Littarry, G.P.; Silvestrini, A.; Meucci, E.; Pontecorvi, A. Hormones and antioxidant systems: Role of pituitary and pituitary-dependent axes. J. Endocrinol. Investig. 2010, 33, 422–433. [Google Scholar]
- Mancini, A.; Leone, E.; Festa, R.; Grande, G.; Silvestrini, A.; De Marinis, L.; Pontecorvi, A.; Maira, G.; Littarru, G.P.; Meucci, E. Effects of testosterone on antioxidant systems in male secondary hypogonadism. J. Androl. 2008, 29, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Akhigbe, R.; Ajayi, A. The impact of reactive oxygen species in the development of cardiometabolic disorders: A review. Lipids Health Dis. 2021, 20, 23. [Google Scholar]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab. J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, C.; Chaturvedi, M.M.; Agarwal, B.-B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar]
- Furukawa, S.; Fujita, T.; Shimabukuru, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar]
- Ceriello, A.; Motz, E. Is oxidative stress the pathogenetic mechanism underlying insulin-resistance, diabetes and cardiovascular disease? The common soil hypothesis revisited. Arterioscler. Thromb. Vasc. Biol. 2007, 24, 816–823. [Google Scholar] [CrossRef]
- Harasym, J.; Oledzki, R. Effect of fruit and vegetable antioxidants on total antioxidant capacity of blood plasma. Nutrition 2014, 30, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Laufs, U.; Wassmann, S.; Czech, T.; Münzel, T.; Eisenhauer, M.; Böhm, M.; Nickenig, G. Physical Inactivity Increases Oxidative Stress, Endothelial Dysfunction, and Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxid. Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Littarru, G.P.; Battino, M.; Tomasetti, M.; Mordente, A.; Santini, S.; Oradei, A.; Manto, A.; Ghirlanda, G. Metabolic implications of coenzyme Q10 in red blood cells and plasma lipoproteins. Mol. Asp. Med. 1994, 15, s67–s72. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Orlando, P.; Silvestri, S.; Marcheggiani, F.; Cirilli, I.; Ziqubu, K.; Ndevahoma, F.; et al. Tea consumption and its effects on primary and secondary prevention of coronary artery disease: Qualitative synthesis of evidence from randomized controlled trials. Clin. Nutr. ESPEN 2021, 41, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Miyake, Y.; Yamamoto, K.; Shimomura, Y.; Ochi, H.; Mori, Y.; Osawa, T. Preparation of a monoclonal anti-body to N(epsilon)-(Hexanonyl)lysine: Application to the evaluation of protective effects of flavonoid supplementation against exercise-induced oxidative stress in rat skeletal muscle. Biochem. Biophys. Res. Commun. 2000, 274, 389–393. [Google Scholar] [CrossRef]
- Kato, Y.; Wu, X.; Naito, M.; Nomura, H.; Kitamoto, N.; Osawa, T. Immunochemical detection of protein dityrosine in atherosclerotic lesion of Apo-E-deficient mice using a novel monoclonal antibody. Biochem. Biophys. Res. Commun. 2000, 275, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Yamauchi, H.; Hasui, Y.; Kurashige, J.; Ochi, H.; Yoshida, K. Quantitative determination of urinary 8-hydroxydeoxyguanosine (8-OH-dg) by using ELISA. Res. Commun. Mol. Pathol. Pharmacol. 2000, 107, 39–44. [Google Scholar]
- Mancini, A.; Bruno, C.; Vergani, E.; Guidi, F.; Meucci, E.; Angelini, F.; Meucci, E.; Silvestrini, A. Evaluation of oxidative stress effects on different macromolecules in adult growth hormone deficiency. PLoS ONE 2020, 15, e0236357. [Google Scholar] [CrossRef]
- Mancini, A.; Guidi, F.; Bruno, C.; Angelini, F.; Vergani, E.; Lanza, P.; Mordente, A.; Meucci, E.; Silvestrini, A. Can plasma antioxidants prevent DNA damage in oxidative stress condition induced by growth hormone deficiency? A pilot study. PLoS ONE 2021, 16, e0248971. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodriguez, M.A.; Mendoza-Nunez, V.M. Oxidative stress indexes for diagnosis of health and disease in humans. Oxid. Med. Cell. Longev. 2019, 25, 4128152. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Li, F.; Duan, Y.; Wen, C.; Wang, W.; Zhang, L.; Huang, R.; Yin, Y. Oxidative stress, nutritional antioxidants and beyond. Sci. China Life Sci. 2020, 63, 866–874. [Google Scholar] [CrossRef]
- Gantenbein, K.V.; Kanaka-Gantembein, C. Mediterranean diet as an antioxidant: The impact on metabolic health and overall wellbeing. Nutrients 2021, 13, 1951. [Google Scholar] [CrossRef]
- Mancini, A.; Martorana, G.E.; Magini, M.; Festa, R.; Raimondo, S.; Silvestrini, A.; Nicolotti, N.; Mordente, A.; Mele, M.C.; Miggiano, G.A.D.; et al. Oxidative stress and metabolic syndrome: Effects of a natural antioxidants enriched diet on insulin resistance. Clin. Nutr. ESPEN 2015, 10, e52–e60. [Google Scholar] [CrossRef]
- Nishi, S.K.; Viguiliouk, E.; Kendall, C.W.C.; Jenkins, D.J.A.; Hu, F.B.; Sievenpiper, J.L.; Atzeni, A.; Misra, A.; Salas-Salvadò, J. Nuts in the prevention and management of type 2 diabetes. Nutrients 2023, 15, 878. [Google Scholar] [CrossRef]
- Vergani, E.; Bruno, C.; Olivieri, G.; Brunetti, A.; Guidi, F.; Guercio, I.; Magini, M.; Miggiano, G.A.D.; Mancini, A. Differential metabolic response to various regimens of natural antioxidant enriched-diets in patients with insulin resistance. Endocr. Rev. 2019, 39 (Suppl. S1), 1417. [Google Scholar]
- Castagneto, M.; De Gaetano, A.; Mingrone, G.; Tacchino, R.; Nanni, G.; Capristo, E.; Benedetti, G.; Tataranni, P.A.; Greco, A.V. Normalization of insulin-sensitivity in the obese patients after stable weight reduction with biliopancreatic diversion. Obes. Surg. 1994, 4, 161–168. [Google Scholar] [CrossRef]
- Tacchino, R.M.; Mancini, A.; Perrelli, M.; Bianchi, A.; Giampietro, A.; Milardi, D.; Vezzosi, C.; Sacco, E.; De Marinis, L. Body composition and energy expenditure: Relationship and changes in obese subjects before and after biliopancreatic diversion. Metab. Clin. Exp. 2003, 52, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Festa, R.; Raimondo, S.; Di Segni, C.; Marchitelli, S.; Pontecorvi, A.; Tacchino, R. Effects of different bariatric surgery procedures on oligoelements and antioxidants in human obesity. In Proceedings of the Abstracts of the 94th Annual Meeting of the Endocrine Society, Houston, TX, USA, 23–26 June 2012. [Google Scholar]
- Mechanick, J.I.; Apovian, C.; Brethauer, S.; Garvey, W.T.; Joffe, A.M.; Kim, J.; Kushner, R.F.; Lindquist, R.; Pessah-Pollack, R.; Seger, J.; et al. Clinical practice guidelines for the perioperative nutrition, metabolic, and nonsurgical support of patients undergoing bariatric procedures—2019 Update. Endocr. Pract. 2019, 25, 1346–1359. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.C.; Braga, P.C.; Martins, A.D.; Silva, B.M.; Alves, M.G.; Oliveira, P.F. Antioxidants present in reproductive tract fluids and their relevance for fertility. Antioxidants 2021, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef]
- Khosrowbeygi, A.; Zarghami, N. Levels of oxidative stress biomarkers in seminal plasma and their relationship with seminal parameters. BMC Clin. Pathol. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Pahune, P.P.; Choudhari, A.R.; Muley, P.A. The total antioxidant power of semen and its correlation with the fertility potential of human male subjects. J. Clin. Diagn. Res. 2013, 7, 991–995. [Google Scholar] [CrossRef]
- Agarwal, A.; Roychoudhury, S.; Bjugstad, K.B.; Cho, C.L. Oxidation-reduction potential of semen: What is its role in the treatment of male infertility? Ther. Adv. Urol. 2016, 8, 302–318. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Meucci, E.; Milardi, D.; Giacchi, E.; Bianchi, A.; Pantano, A.L.; Mordente, A.; Martorana, G.E.; De Marinis, L. Seminal antioxidant capacity in pre- and post-operative varicocele. J. Androl. 2004, 25, 44–49. [Google Scholar] [CrossRef]
- Evans, E.P.P.; Scholten, J.T.M.; Mzyk, A.; Reyes-San-Martin, C.; Llumbet, A.E.; Hamoh, T.; Arts, E.G.J.M.; Schirhagl, R.; Cantineau, A.E.P. Male subfertility and oxidative stress. Redox Biol. 2021, 46, 102071. [Google Scholar] [CrossRef]
- Bruno, C.; Basile, U.; Vergani, E.; Napodano, C.; Oliva, A.; Gulli, F.; Meucci, E.; Silvestrini, A.; Orlando, P.; Silvestri, S.; et al. Inflammation and Oxidative Stress in Seminal Plasma: Search for Biomarkers in Diagnostic Approach to Male Infertility. J. Pers. Med. 2022, 12, 857. [Google Scholar] [CrossRef]
- Mancini, A.; Milardi, D.; Bianchi, A.; Festa, R.; Silvestrini, A.; De Marinis, L.; Pontecorvi, A.; Meucci, E. Increased Total Antioxidant Capacity in Seminal Plasma of Varicocele Patients: A Multivariate Analysis. Arch. Androl. 2007, 53, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Oliva, A.; Vergani, E.; Festa, R.; Silvestrini, A. The Dual Role of Oxidants in Male (In)fertility: Every ROSe Has a Thorn. Int. J. Mol. Sci. 2023, 24, 4994. [Google Scholar] [CrossRef] [PubMed]
- Bruno, C.; Silvestrini, A.; Calarco, R.; Favuzzi, A.M.R.; Vergani, E.; Nicolazzi, M.A.; d’Abate, C.; Meucci, E.; Mordente, A.; Landolfi, R.; et al. Anabolic Hormones Deficiencies in Heart Failure With Preserved Ejection Fraction: Prevalence and Impact on Antioxidants Levels and Myocardial Dysfunction. Front. Endocrinol. 2020, 11, 281. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antioxidant | Source | Range | Solubility |
|---|---|---|---|
| Vitamin C | Exogenous (diet) | 0.4–1.5 mg/dL | Water-soluble |
| Vitamin E | Exogenous (diet) | 7.4–23.5 mg/L | Fat-soluble |
| Vitamin A | Exogenous (diet) | 0.22–0.62 mg/L | Fat-soluble |
| Uric acid | Purine catabolism | 2.5–8 mg/dL | Water-soluble |
| GSH | GSSG reductases | 60–80 μmol/L | Water-soluble |
| Assay 1 | Probe 2 | Wavelength (λmax) | Method | Endpoint |
|---|---|---|---|---|
| TEAC | ABTS | Absorbance (734 nm) | Indirect | Lag-phase |
| ORAC | ABAP | Fluorescence (λex 495 nm) | Direct | Fixed time |
| FRAP | Fe-TPTZ | Absorbance (593 nm) | Indirect | Varies times |
| CUPRAC | Neocuproine | Absorbance (450 nm) | Indirect | Time |
| TRAP | R-PE | Fluorescence (λex 495 nm) | Direct | Lag-phase |
| DCHFT | DCHF-DA | Fluorescence (λex 502 nm) | Intracellular | Staining |
| NBT-T | NBT | Absorbance (540 nm) | Intracellular | Staining |
| DPPH-SA | DPPH | Absorbance (517 nm) | In vivo | % inhibition |
| SOD | Pyrogallol | Absorbance (420 nm) | In vivo | Fixed time |
| RP-m | K3Fe(CN)6/FeCl3 | Absorbance (700 nm) | In vitro | Time |
| TBA-m | TBA | Absorbance (552 nm) | In vitro | Time |
| DMPD-m | DMPD | Absorbance (505 nm) | In vitro | Time |
| GSH | Ellman’s reagent | Absorbance (412 nm) | In vivo | standard curve |
| CAT | H2O2 | Absorbance (240 nm) | In vivo | Time |
| LPO | MDA | Absorbance (532 nm) | In vivo | TBARS/mg protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvestrini, A.; Meucci, E.; Ricerca, B.M.; Mancini, A. Total Antioxidant Capacity: Biochemical Aspects and Clinical Significance. Int. J. Mol. Sci. 2023, 24, 10978. https://doi.org/10.3390/ijms241310978
Silvestrini A, Meucci E, Ricerca BM, Mancini A. Total Antioxidant Capacity: Biochemical Aspects and Clinical Significance. International Journal of Molecular Sciences. 2023; 24(13):10978. https://doi.org/10.3390/ijms241310978
Chicago/Turabian StyleSilvestrini, Andrea, Elisabetta Meucci, Bianca Maria Ricerca, and Antonio Mancini. 2023. "Total Antioxidant Capacity: Biochemical Aspects and Clinical Significance" International Journal of Molecular Sciences 24, no. 13: 10978. https://doi.org/10.3390/ijms241310978
APA StyleSilvestrini, A., Meucci, E., Ricerca, B. M., & Mancini, A. (2023). Total Antioxidant Capacity: Biochemical Aspects and Clinical Significance. International Journal of Molecular Sciences, 24(13), 10978. https://doi.org/10.3390/ijms241310978