
Bortezomib Increased Vascular Permeability by Decreasing Cell–Cell Junction Molecules in Human Pulmonary Microvascular Endothelial Cells
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
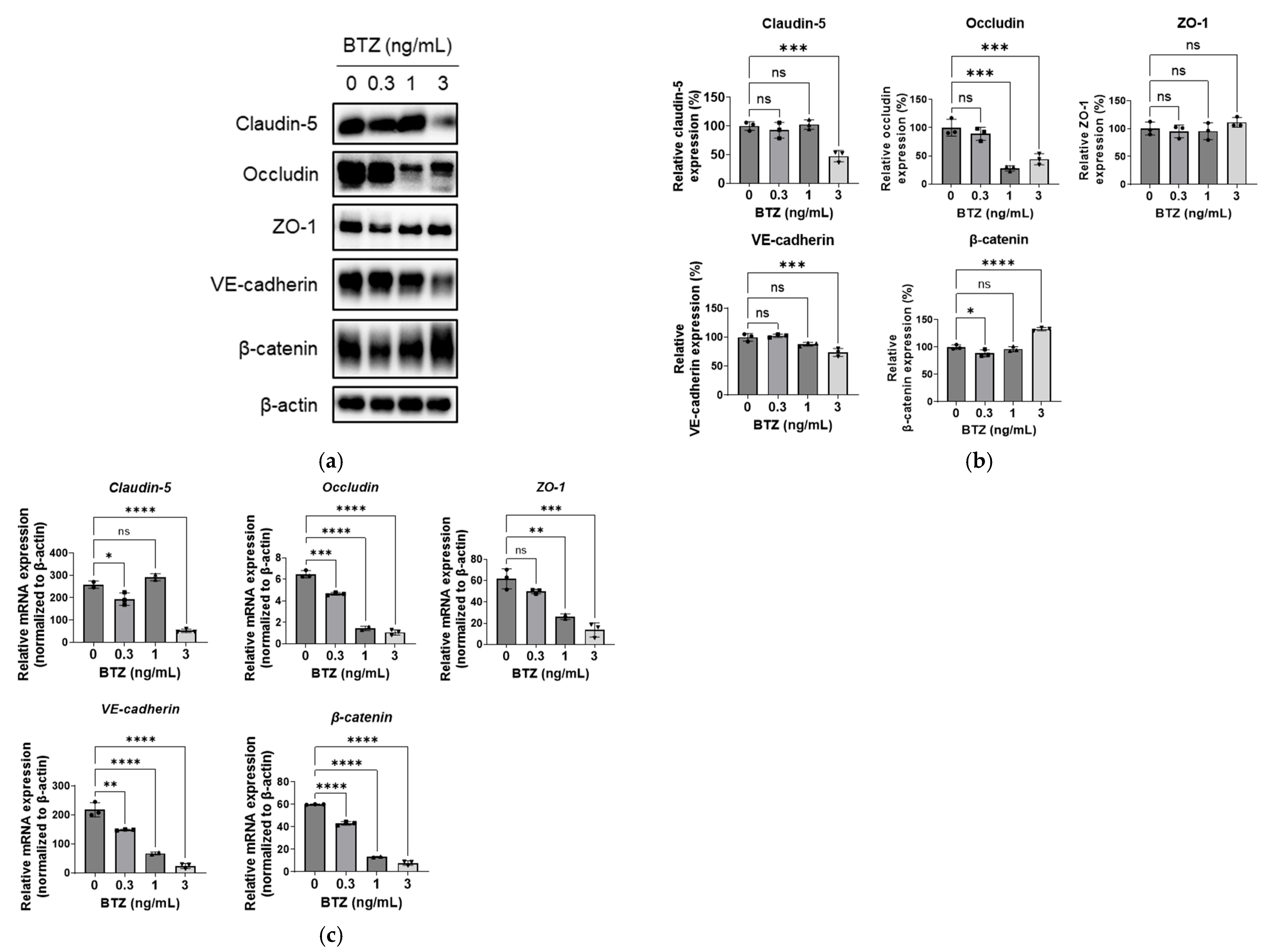
2. Results
2.1. Cytotoxicity of BTZ towards HPMEC Monolayers
2.2. Effect of BTZ on the Permeability of HPMEC Monolayers
2.3. Effect of BTZ on Adhesion Molecule Expression in HPMECs
3. Discussion
4. Materials and Methods
4.1. Culture of HPMECs
4.2. Viability Assay
4.3. Giemsa Staining
4.4. Permeability Assay
4.5. Western Blotting
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kane, R.C.; Bross, P.F.; Farrell, A.T.; Pazdur, R. Velcade: U.S. FDA approval for the treatment of multiple myeloma progressing on prior therapy. Oncologist 2003, 8, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J.; Karin, M. Nuclear factor-kappaB: A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Adams, J. Proteasome inhibition in cancer: Development of PS-341. Semin. Oncol. 2001, 28, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Hideshima, T.; Richardson, P.; Chauhan, D.; Palombella, V.J.; Elliott, P.J.; Adams, J.; Anderson, K.C. The proteasome inhibitor PS-341 inhibits growth, induces apoptosis, and overcomes drug resistance in human multiple myeloma cells. Cancer Res. 2001, 61, 3071–3076. [Google Scholar] [PubMed]
- Pancheri, E.; Guglielmi, V.; Wilczynski, G.M.; Malatesta, M.; Tonin, P.; Tomelleri, G.; Nowis, D.; Vattemi, G. Non-Hematologic Toxicity of Bortezomib in Multiple Myeloma: The Neuromuscular and Cardiovascular Adverse Effects. Cancers 2020, 12, 2540. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.E.; Batra, R.B.; Ascensao, J.L.; Schechter, G.P. Severe pulmonary complication after bortezomib treatment for multiple myeloma. Blood 2006, 108, 1113. [Google Scholar] [CrossRef] [PubMed]
- Miyakoshi, S.; Kami, M.; Yuji, K.; Matsumura, T.; Takatoku, M.; Sasaki, M.; Narimatsu, H.; Fujii, T.; Kawabata, M.; Taniguchi, S.; et al. Severe pulmonary complications in Japanese patients after bortezomib treatment for refractory multiple myeloma. Blood 2006, 107, 3492–3494. [Google Scholar] [CrossRef]
- Chew, E.; Filshie, R.; Wei, A. Development of fatal bortezomib induced acute lung injury despite concurrent therapy with high-dose dexamethasone. Leuk. Lymphoma 2007, 48, 212–213. [Google Scholar] [CrossRef]
- Kharel, P.; Uprety, D.; Chandra, A.B.; Hu, Y.; Belur, A.A.; Dhakal, A. Bortezomib-Induced Pulmonary Toxicity: A Case Report and Review of Literature. Case Rep. Med. 2018, 2018, 2913124. [Google Scholar] [CrossRef]
- Duek, A.; Feldberg, E.; Haran, M.; Berrebi, A. Pulmonary fibrosis in a myeloma patient on bortezomib treatment. A new severe adverse effect of a new drug. Am. J. Hematol. 2007, 82, 502–503. [Google Scholar] [CrossRef]
- Mukai, H.; Ohyashiki, K.; Katoh, T.; Kusumoto, M.; Gemma, A.; Sakai, H.; Sugiyama, Y.; Hatake, K.; Fukuda, Y.; Kudoh, S. Lung injury associated with bortezomib therapy in Japan. Rinsho Ketsueki 2011, 52, 1859–1869. [Google Scholar]
- Matsumoto, T.; Takamatsu, Y.; Moriyama, H.; Terada, K.; Mori, M.; Ono, K.; Migita, K.; Hara, S. Bortezomib enhances G-CSF-induced hematopoietic stem cell mobilization by decreasing CXCL12 levels and increasing vascular permeability. Exp. Hematol. 2021, 97, 21–31. [Google Scholar] [CrossRef]
- Papandreou, C.N.; Daliani, D.D.; Nix, D.; Yang, H.; Madden, T.; Wang, X.; Pien, C.S.; Millikan, R.E.; Tu, S.M.; Pagliaro, L.; et al. Phase I trial of the proteasome inhibitor bortezomib in patients with advanced solid tumors with observations in androgen-independent prostate cancer. J. Clin. Oncol. 2004, 22, 2108–2121. [Google Scholar] [CrossRef]
- Moreau, P.; Coiteux, V.; Hulin, C.; Leleu, X.; van de Velde, H.; Acharya, M.; Harousseau, J.L. Prospective comparison of subcutaneous versus intravenous administration of bortezomib in patients with multiple myeloma. Haematologica 2008, 93, 1908–1911. [Google Scholar] [CrossRef]
- Leal, T.B.; Remick, S.C.; Takimoto, C.H.; Ramanathan, R.K.; Davies, A.; Egorin, M.J.; Hamilton, A.; LoRusso, P.A.; Shibata, S.; Lenz, H.J.; et al. Dose-escalating and pharmacological study of bortezomib in adult cancer patients with impaired renal function: A National Cancer Institute Organ Dysfunction Working Group Study. Cancer Chemother. Pharmacol. 2011, 68, 1439–1447. [Google Scholar] [CrossRef]
- Takata, F.; Dohgu, S.; Yamauchi, A.; Matsumoto, J.; Machida, T.; Fujishita, K.; Shibata, K.; Shinozaki, Y.; Sato, K.; Kataoka, Y.; et al. In vitro blood-brain barrier models using brain capillary endothelial cells isolated from neonatal and adult rats retain age-related barrier properties. PLoS ONE 2013, 8, e55166. [Google Scholar] [CrossRef]
- Venkatakrishnan, K.; Rader, M.; Ramanathan, R.K.; Ramalingam, S.; Chen, E.; Riordan, W.; Trepicchio, W.; Cooper, M.; Karol, M.; von Moltke, L.; et al. Effect of the CYP3A inhibitor ketoconazole on the pharmacokinetics and pharmacodynamics of bortezomib in patients with advanced solid tumors: A prospective, multicenter, open-label, randomized, two-way crossover drug-drug interaction study. Clin. Ther. 2009, 31 Pt 2, 2444–2458. [Google Scholar] [CrossRef]
- Shasby, D.M. Cell-cell adhesion in lung endothelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L593–L607. [Google Scholar] [CrossRef]
- Tamura, D.; Arao, T.; Tanaka, K.; Kaneda, H.; Matsumoto, K.; Kudo, K.; Aomatsu, K.; Fujita, Y.; Watanabe, T.; Saijo, N.; et al. Bortezomib potentially inhibits cellular growth of vascular endothelial cells through suppression of G2/M transition. Cancer Sci. 2010, 101, 1403–1408. [Google Scholar] [CrossRef]
- Uttamsingh, V.; Lu, C.; Miwa, G.; Gan, L.S. Relative contributions of the five major human cytochromes P450, 1A2, 2C9, 2C19, 2D6, and 3A4, to the hepatic metabolism of the proteasome inhibitor bortezomib. Drug Metab. Dispos. 2005, 33, 1723–1728. [Google Scholar] [CrossRef]
- Fatunde, O.A.; Brown, S.A. The Role of CYP450 Drug Metabolism in Precision Cardio-Oncology. Int. J. Mol. Sci. 2020, 21, 604. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016, 15, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Vanlandewijck, M.; He, L.; Mae, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Lavina, B.; Gouveia, L.; et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature 2018, 554, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Castro Dias, M.; Coisne, C.; Lazarevic, I.; Baden, P.; Hata, M.; Iwamoto, N.; Francisco, D.M.F.; Vanlandewijck, M.; He, L.; Baier, F.A.; et al. Claudin-3-deficient C57BL/6J mice display intact brain barriers. Sci. Rep. 2019, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Hashimoto, Y.; Shirakura, K.; Okada, Y.; Hirayama, R.; Iwashita, Y.; Nishino, I.; Ago, Y.; Takeda, H.; Kuniyasu, H.; et al. Safety and efficacy of an anti-claudin-5 monoclonal antibody to increase blood-brain barrier permeability for drug delivery to the brain in a non-human primate. J. Control. Release 2021, 336, 105–111. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Zhou, W.; Hamauchi, K.; Shirakura, K.; Doi, T.; Yagi, K.; Sawasaki, T.; Okada, Y.; Kondoh, M.; Takeda, H. Engineered membrane protein antigens successfully induce antibodies against extracellular regions of claudin-5. Sci. Rep. 2018, 8, 8383. [Google Scholar] [CrossRef]
- Clark, P.R.; Kim, R.K.; Pober, J.S.; Kluger, M.S. Tumor necrosis factor disrupts claudin-5 endothelial tight junction barriers in two distinct NF-kappaB-dependent phases. PLoS ONE 2015, 10, e0120075. [Google Scholar] [CrossRef]
- Ding, Y.X.; Eerduna, G.W.; Duan, S.J.; Li, T.; Liu, R.X.; Zhang, L.M.; Wang, T.; Fu, F.H. Escin ameliorates the impairments of neurological function and blood brain barrier by inhibiting systemic inflammation in intracerebral hemorrhagic mice. Exp. Neurol. 2021, 337, 113554. [Google Scholar] [CrossRef]
- Ibrahim, S.; Zhu, X.; Luo, X.; Feng, Y.; Wang, J. PIK3R3 regulates ZO-1 expression through the NF-kB pathway in inflammatory bowel disease. Int. Immunopharmacol. 2020, 85, 106610. [Google Scholar] [CrossRef]
- Colas-Algora, N.; Garcia-Weber, D.; Cacho-Navas, C.; Barroso, S.; Caballero, A.; Ribas, C.; Correas, I.; Millan, J. Compensatory increase of VE-cadherin expression through ETS1 regulates endothelial barrier function in response to TNFalpha. Cell. Mol. Life Sci. 2020, 77, 2125–2140. [Google Scholar] [CrossRef]
- Aberle, H.; Bauer, A.; Stappert, J.; Kispert, A.; Kemler, R. beta-catenin is a target for the ubiquitin-proteasome pathway. EMBO J. 1997, 16, 3797–3804. [Google Scholar] [CrossRef]
- Qiang, Y.W.; Hu, B.; Chen, Y.; Zhong, Y.; Shi, B.; Barlogie, B.; Shaughnessy, J.D., Jr. Bortezomib induces osteoblast differentiation via Wnt-independent activation of beta-catenin/TCF signaling. Blood 2009, 113, 4319–4330. [Google Scholar] [CrossRef]
- Guo, M.; Breslin, J.W.; Wu, M.H.; Gottardi, C.J.; Yuan, S.Y. VE-cadherin and beta-catenin binding dynamics during histamine-induced endothelial hyperpermeability. Am. J. Physiol. Cell. Physiol. 2008, 294, C977–C984. [Google Scholar] [CrossRef]
- Prakoso, A.T.; Basri, H.; Adanta, D.; Yani, I.; Ammarullah, M.I.; Akbar, I.; Ghazali, F.A.; Syahrom, A.; Kamarul, T. The Effect of Tortuosity on Permeability of Porous Scaffold. Biomedicines 2023, 11, 427. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Afif, I.Y.; Maula, M.I.; Winarni, T.I.; Tauviqirrahman, M.; Akbar, I.; Basri, H.; van der Heide, E.; Jamari, J. Tresca Stress Simulation of Metal-on-Metal Total Hip Arthroplasty during Normal Walking Activity. Materials 2021, 14, 7554. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Hartono, R.; Supriyono, T.; Santoso, G.; Sugiharto, S.; Permana, M.S. Polycrystalline Diamond as a Potential Material for the Hard-on-Hard Bearing of Total Hip Prosthesis: Von Mises Stress Analysis. Biomedicines 2023, 11, 951. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Santoso, G.; Sugiharto, S.; Supriyono, T.; Wibowo, D.B.; Kurdi, O.; Tauviqirrahman, M.; Jamari, J. Minimizing Risk of Failure from Ceramic-on-Ceramic Total Hip Prosthesis by Selecting Ceramic Materials Based on Tresca Stress. Sustainability 2022, 14, 13413. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Santoso, G.; Sugiharto, S.; Supriyono, T.; Kurdi, O.; Tauviqirrahman, M.; Winarni, T.I.; Jamari, J. Tresca stress study of CoCrMo-on-CoCrMo bearings based on body mass index using 2D computational model. J. Tribol. 2022, 33, 31–38. [Google Scholar]
- Salaha, Z.F.M.; Ammarullah, M.I.; Abdullah, N.; Aziz, A.U.A.; Gan, H.S.; Abdullah, A.H.; Abdul Kadir, M.R.; Ramlee, M.H. Biomechanical Effects of the Porous Structure of Gyroid and Voronoi Hip Implants: A Finite Element Analysis Using an Experimentally Validated Model. Materials 2023, 16, 3298. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, T.; Matsumoto, J.; Matsushita, Y.; Arimura, M.; Aono, K.; Aoki, M.; Terada, K.; Mori, M.; Haramaki, Y.; Imatoh, T.; et al. Bortezomib Increased Vascular Permeability by Decreasing Cell–Cell Junction Molecules in Human Pulmonary Microvascular Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 10842. https://doi.org/10.3390/ijms241310842
Matsumoto T, Matsumoto J, Matsushita Y, Arimura M, Aono K, Aoki M, Terada K, Mori M, Haramaki Y, Imatoh T, et al. Bortezomib Increased Vascular Permeability by Decreasing Cell–Cell Junction Molecules in Human Pulmonary Microvascular Endothelial Cells. International Journal of Molecular Sciences. 2023; 24(13):10842. https://doi.org/10.3390/ijms241310842
Chicago/Turabian StyleMatsumoto, Taichi, Junichi Matsumoto, Yuka Matsushita, Moeno Arimura, Kentaro Aono, Mikiko Aoki, Kazuki Terada, Masayoshi Mori, Yutaka Haramaki, Takuya Imatoh, and et al. 2023. "Bortezomib Increased Vascular Permeability by Decreasing Cell–Cell Junction Molecules in Human Pulmonary Microvascular Endothelial Cells" International Journal of Molecular Sciences 24, no. 13: 10842. https://doi.org/10.3390/ijms241310842
APA StyleMatsumoto, T., Matsumoto, J., Matsushita, Y., Arimura, M., Aono, K., Aoki, M., Terada, K., Mori, M., Haramaki, Y., Imatoh, T., Yamauchi, A., & Migita, K. (2023). Bortezomib Increased Vascular Permeability by Decreasing Cell–Cell Junction Molecules in Human Pulmonary Microvascular Endothelial Cells. International Journal of Molecular Sciences, 24(13), 10842. https://doi.org/10.3390/ijms241310842