Abstract
Reeta is a popular late-maturing high-yielding rice variety recommended for cultivation in the eastern Indian states. The cultivar is highly sensitive to submergence stress. Phosphorus deficiency is an additional constraint for realizing high yield. The quantitative trait loci (QTLs), Sub1, for submergence and Pup1 for low phosphorus stress tolerance along with narrow-grained trait, GW5 were introgressed into the variety from the donor parent, Swarna-Sub1 through marker-assisted breeding. In addition, phenotypic selections for higher panicle weight, grain number, and spikelet fertility were performed in each segregating generation. Foreground selection detected the 3 target QTLs in 9, 8 and 7 progenies in the BC1F1, BC2F1, and BC3F1 generation, respectively. Recurrent parent’s genome recovery was analyzed using 168 SSR polymorphic markers. The foreground analysis in 452 BC3F2 progenies showed five pyramided lines in homozygous condition for the target QTLs. No donor fragment drag was noticed in the Sub1 and GW5 QTLs carrier while a segmentwas observed in the Pup1 carrier chromosome. The developed lines were higher yielding, had submergence, and had low phosphorus stress-tolerance alongwith similar to the recipient parent in the studied morpho-quality traits. A promising pyramided line is released in the name of Reeta-Panidhan (CR Dhan 413) for the flood-prone areas of Odisha state.
1. Introduction
Rice is life for millions of farmers and rice based traders in India and many Asiatic countries. Rice cultivation for consumption provides livelihood for millions of farmers in India. Importance of rice is known to people even from its use in rituals for birth, marriage and death and other ceremonies. Rice though mainly supplies carbohydrate but compounds such as quality proteins, many minerals, vitamins, specific oils, and dietary fiber along with few useful antioxidants are available in the grain [1,2,3]. The crop is cultivated from very high elevations to a below sea level and is highly diverse in nature. Rice is cultivated in about 160 Mha in the world. The annual value from the rice is about 206 billion USD, which gives about 17% of the total crop value globally [4]. Rainfed rice is the practice in about 45% of the total rice cultivated area [5]. The production and productivity of the crop is now highly challenged by many climate change-related adverse effects [6]. The distribution pattern of monsoon rain in India during the last few decades confirms about the variability of rainfall distribution which causes instable rice production in the rainfed ecology. The variability in monsoon rainfall may invite stresses like drought, submergence, salinity, and mineral deficiencies. Few stresses are interrelated and the effects on rice yield may be very high.
The area under rainfed rice is 22 Mha, of which 90% is located in eastern region of India [7]. Flash flood is a common adverse effect faced by the farmers in this region for and suffers a huge crop loss by growing high yielding and submergence sensitive varieties [8,9]. The occurrence of flash floods is common in rainy season in India, particularly in the eastern states of the country. Low rice production in the region is mainly due to the cultivation of sensitive varieties those are affected by submergence stress. Rice variety, Reeta (CR Dhan 401) is a popular variety for the late maturity group but susceptible to submergence stress under flash flood situation. A total crop failure occurs if the crop is exposed to two weeks submergence stress due to flash flood. But, the panicle of Reeta variety gives more grains/panicle and bears heavy panicle than the variety, Swarna-Sub1. However, the grains of Reeta are bold type with more gain width than Swarna-Sub1. Varieties built-in with submergence tolerance QTL, Sub1, confers tolerance for approximately two weeks submergence stress [10]. Gene-based and closely linked markers are available for marker-assisted selection for transfer of the Sub1 QTL. This tolerance trait has been incorporated into many high yielding varieties including Swarna using QTL introgression [7,11,12,13,14,15,16]. The transfer of the target gene from an improved source to recipient variety may contribute very less undesirable genetic effects from the donor variety [17].
Phosphorous (P) is an essential major nutrient for rice plant growth, development and higher production. P-deficiency is a global problem and tentatively 5.7 billion hectares of cultivated areas are with phosphorus deficient [18]. The soil phosphorus availability is low or medium in about 80% of the districts in India [19,20]. Direct seeded rice is a common practice for rainfed ecology that faces a deficiency in phosphorus very frequently [7]. Among the major nutrients, P is the second limiting nutrient for rice cultivation. About 30 to 65 percent of total soil phosphorus is in organic and unavailable forms to the plant. Indian farmers usually neglect application of required quality of this fertilizer due to it’s high cost in our country [21]. In addition, fertilizer runoff may leads to creation of pollution in water bodies. Therefore, rice cultivation with tolerance to low soil phosphorus level is very much needed for the farmers in India and abroad. The trait is controlled by a major QTL, Pup1 which is mapped and located on chromosome 12 providing phenotypic variance of >70% tolerance to phosphorus deficiency in soil [15,22]. The full Pup1 region comprised of 68 genes of which PSTOL1 QTL is the most probable candidate gene that confers low-P stress tolerance in rice [23]. The other region within the Pup1 QTL may provide a supportive role for better phosphorus uptake Rice varieties with low soil P stress tolerance will be P efficient and will produce more at a lower input cost. Adaptation of low p-stress tolerant varieties/cultivars requires neither additional cost nor major changes in the cropping system. The Pup1 introgressed rice varieties developed by marker-assisted breeding programs showed very good results in the field trials [24,25]. Thus, popular varieties need to be pyramided with low P-tolerance QTL, Pup1 that can grow and yield well even under low P-availability.
The narrow grained rice is more preferred and a market driven trait in recent years in India. Though there is impressive growth in rice production and productivity in recent years, the demand of this type of rice is growing upward in the country. Enhancement of yield is also regulated by the grain weight and grain dimension QTLs viz., GS3, GW2, GS5, and GW5 [9]. Reduced grain width is associated with the QTL, GW5 and the effects of the QTL were consistent under multiple environments. Deletion of a nucleotide segment in the GW5 region resulted in wide-grained genotypes in most japonica and indica rice [26]. The donor parent, Swarna-Sub1 exhibits narrow grain type and carries the QTL, GW5 [5]. The variety Reeta is a late maturing variety which produces a wide-grained kernel. This investigation aims at the development of pyramided lines carrying Sub1, Pup1, and GW5 (narrow grain) QTL for submergence, low phosphorus tolerance and narrow-grained rice along with panicle traits improvement in the popular variety background, ‘Reeta’.
2. Result
2.1. Validation of the Donor and Recipient Parents for the Target Traits
The target QTLs namely Sub1, Pup1, and GW5 controlling the traits were validated for their presence by comparing the donor versus recipient parents before beginning the hybridization and selection activities. The presence of target QTLs, Sub1 for submergence tolerance, Pup1 for low phosphorus tolerance, and GW5 for yield component grain weight were confirmed in the donor parent (Supplementary Figure S1). The recipient parent, Reeta was a high-yielding popular variety but deficient in these QTLs. The gene-based and tightly linked molecular markers for Sub1, a direct marker for Pup1, and the yield component QTL, GW5, were used to validate and track the target QTL in the parental and backcross-derived lines (Supplementary Table S1; Supplementary Figure S1). A parental polymorphism survey was performed by using 728 simple sequence repeat markers covering all the chromosomes. A total of 168 polymorphic markers between the two parents were deployed for background screening (Supplementary Table S2).
2.2. Marker-Assisted Selection in BC1F1 Progenies
Reeta was hybridized with ‘Swarna-Sub1′, and 250 F1 seeds were obtained. The hybridity in F1 plants was confirmed by genotyping the hybrid plants using the Sub1 specific marker. One true F1 plant was crossed with the recipient parent, Reeta, and a total of 265 BC1F1 seeds were generated. A total of 138 backcross generation 1 seeds were raised and the remaining 127 BC1F1 seeds were stored as backup seeds. Foreground screening was performed in all the 140 BC1F1 plants using the 7 selected molecular markers for the QTLs, Sub1, Pup1, and GW5 (Figure 1).
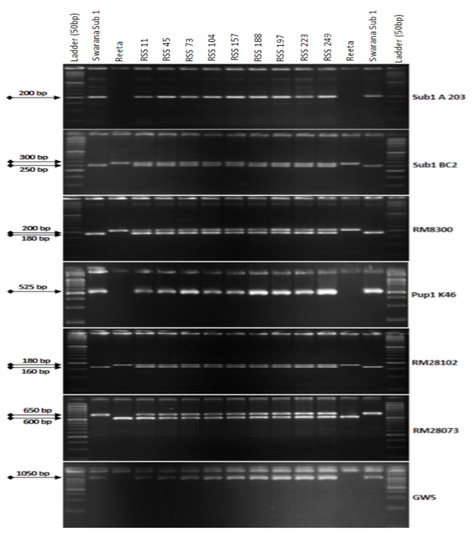
Figure 1.
PCR amplification of markers Sub1A203 and Sub1BC2 linked to submergence tolerance; Pup1K46, RM28073 and RM28102 linked to low phosphorus tolerance genes along with grain yield marker GW5 in BC1F1 progenies. L: Molecular weight marker (50 bp plus ladder) and lanes on the top of the gel indicate BC1F1 progenies (RSS: Reeta Swarna-Sub1).
The screening results from 138 BC1F1 progenies of the backcross revealed the presence of Sub1 QTL in 65 derivatives detected by the markers Sub1 A203 and Sub1BC2 (200 bp and 240 bp). Tracking of the Pup1 QTL by the markers Pup1K46, RM28073 and RM28102 showed its presence in 25 individuals out of a total of 65 derivatives with submergence tolerance. Screening for the presence of GW5 Swarna allele (narrow-grain) controlling the grain width and weight was detected in 9 individuals to carry all the 3 target traits. Background screening was performed in the 9 BC1F1 foreground-positive progenies by using 168 SSR markers. Out of these 9 individuals, the progeny carrying the maximum recipient genome content was selected for the next backcross. The recurrent parent’s genome content in those 9 progenies varied from 64.88% to 80.95% with an average value of 76.91% (Table 1). The backcross derivatives RSS45 and RSS223 showed the recurrent genome content of 80.24% and 80.95%, respectively. However, higher panicle weight, grain number and spikelet fertility were better in RSS45 than RSS223. The BC1F1 line generated from RSS45 was backcrossed with the recipient parent, Reeta, to obtain BC2F1 seeds.

Table 1.
Genotyping of backcross progenies for three QTLs and recovery of recipient parent’s genome in the foreground positive backcross progenies.
2.3. Marker-Assisted Selection in BC2F1 Generation
One hundred and thirty-five BC2F1 plants were grown in the field for selection. The 3 target QTLs were tracked by foreground selection using gene-specific and linked markers. The genotyping results of 135 BC2F1 individuals showed 64 positive progenies for Sub1 QTL. These individuals were checked for the presence/absence of the Pup1 QTL using markers Pup1K46, RM28073, and RM28102. A total of 27 positive plants carrying Pup1 QTL were identified by tracking the QTL. Those 27 plants were checked for the presence of the GW5Swarna allele (narrow-grain) using a gene-specific marker. Eight plants with all the desired QTLs were identified for further background selection (Figure 2). Background screening for recovery of the recipient parent genome in those 8 identified plants containing the 3 targets QTLs varied from 81.54% to 92.26% with an average of 89.06% (Table 1). The plant RSS45-83 showing 92.26% of the recipient variety Reeta genome content, was used for the next BC3 backcrossing.
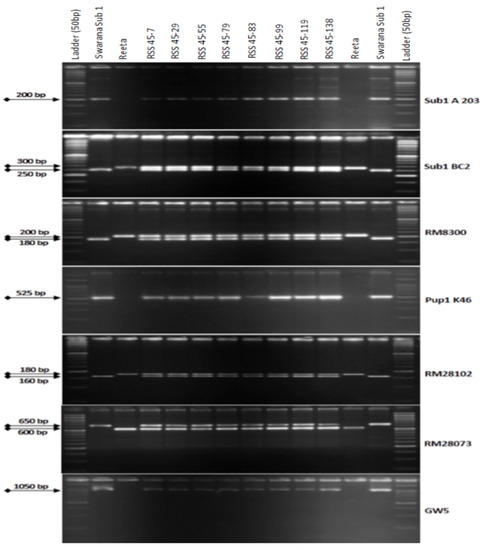
Figure 2.
PCR amplification of markers Sub1A203 and Sub1BC2 linked to submergence tolerance; Pup1K46, RM28073 and RM28102 linked to low phosphorus tolerance genes along with grain yield marker GW5 in BC2F1 progenies. L: Molecular weight marker (50 bp plus ladder) and lanes on the top of the gel indicate BC1F1 progenies (RSS: Reeta Swarna-Sub1).
2.4. Marker-Assisted Selection in BC3F1 and BC3F2 Generations
The BC3F1 seeds were generated by crossing BC2F1 plant no. RSS45-83 and the recurrent parent ‘Reeta’. A total of 156 BC3F1 seeds were generated and raised for molecular screening by foreground and background selections. The genotyping results for the target QTL, Sub1, were positive in 65 progenies. Those 65 Sub1 carrier plants were genotyped for checking the presence/absence of the Pup1 QTL. Within this population, 27 plants were positive for 3 Pup1 markers. A total of 30 positive plants were screened for GW5 QTL. This analysis identified 7 plants positive for GW5 and further genotyped for background screening (Figure 3). The background analysis using 168 SSR markers in these 7 plants detected 92.86 to 95.24% recurrent parent’s genome recovery, with an average of 94.30% (Table 1). The highest recurrent genome containing plant RSS45-83-27 was self-bred, and 22.5 g seeds were produced for further evaluation in BC3F2 generation. A total of 452 self-bred seeds were raised, and the rest seeds were kept as backup seeds. All the plants were subjected to foreground screening, of which 96 plants were identified to be homozygous for the Sub1 allele. All the Sub1-homozygous plants were screened homozygosity for Pup1 QTL carrying plant. A total of 19 plants showed the presence of Pup1 in the homozygous condition. Those plants were screened for the presence of GW5 in homozygous conditions. Finally, 5 plants were identified to carry all the 3 targets QTL in homozygous condition (Figure 4). Seeds of these five pyramided lines were used for further evaluation in the subsequent generations for various morphological and quality traits. Cluster analysis with agro-morphologic and quality traits showed distinct clusters of Reeta with 2 pyramided lines, RSS45-83-27-54 and RSS45-83-27-415 while the donor parent was placed in a separate cluster (Figure 5A). Additionally, a dendrogram was generated by using the alleles detected with the SSR markers, which grouped the developed pyramided and parental lines into two groups. Group 1 is further divided into two subgroups with the 3 pyramided lines (RSS45-83-27-54, RSS45-83-27-526, and RSS45-83-27-648) and the recipient parent in one cluster and the other 2 pyramided lines in a separate group (Figure 5B). Five pyramided lines were accommodated in cluster I along with the recipient parent ‘Reeta’, while the donor parent remained in cluster II. The backcross-derived lines in cluster I were found to form different subclusters based on the 14 agro-morphologic traits studied but were similar to the recipient parent ‘Reeta’ for the majority of the studied morphological and quality traits. The pyramided lines RSS45-83-27-54, RSS45-83-27-235, RSS45-83-27-415, RSS45-83-27-526, and RSS45-83-27-648 were almost similar in terms of genome recovery among themselves and with recipient parent ‘Reeta’ except grain size.
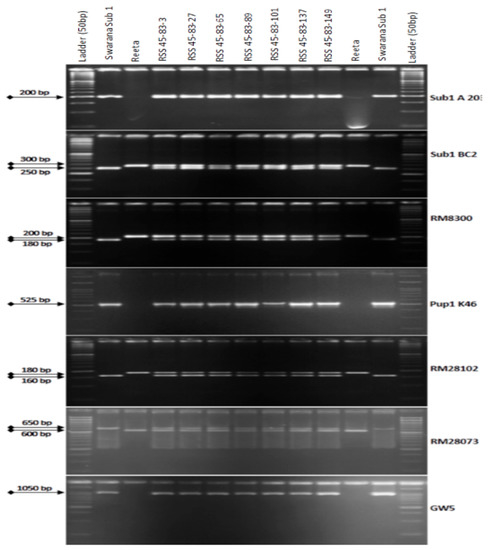
Figure 3.
PCR amplification of markers Sub1A203 and Sub1BC2 linked to submergence tolerance; Pup1K46, RM28073 and RM28102 linked to low phosphorus tolerance genes along with grain yield marker GW5 in BC1F1 progenies. L: Molecular weight marker (50 bp plus ladder) and lanes on the top of the gel indicate BC3F1 progenies (RSS: Reeta Swarna-Sub1).
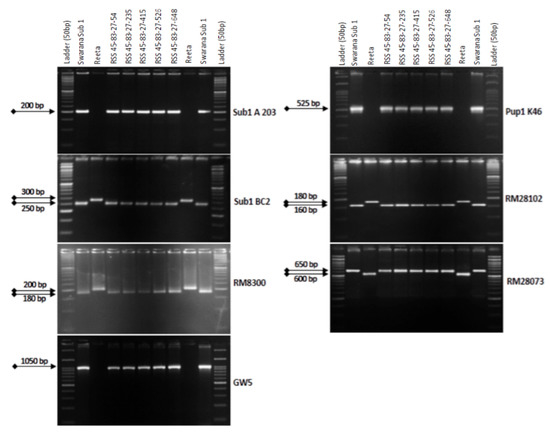
Figure 4.
PCR amplification of markers Sub1A203 and Sub1BC2 linked to submergence tolerance; Pup1K46, RM28073 and RM28102 linked to low phosphorus tolerance genes along with grain yield marker GW5 in BC1F1 progenies. L: Molecular weight marker (50 bp plus ladder) and lanes on the top of the gel indicate BC3F2 progenies (RSS: Reeta Swarna-Sub1).
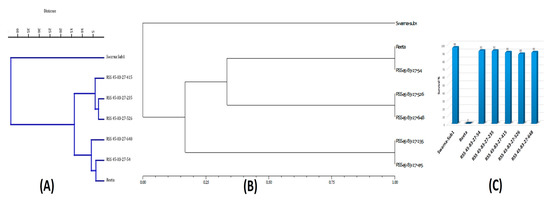
Figure 5.
Seven pyramided lines of Swarna-Sub1 and Reeta along with parents in (A) dendrogram showing relatedness based on the 14 morphologic and quality traits; (B) Dendrogram showing the genetic relationship between lines based on 7 microsatellite markers and (C) % contribution of recurrent genome in the pyramided lines.
2.5. Analysis of Genome Introgression on the Carrier Chromosomes of the Pyramided Lines
The background analysis for recipient genome recovery and genetic drag linked to the donor segments were assessed using 168 background and 7 foreground markers. The markers were carefully selected for all the chromosomes to obtain maximum coverage in background screening. The foreground analysis detected five BC3F2 pyramided lines carrying the 3 target QTLs in the homozygous condition in the progenies. The Sub1 carrier on chromosome 9 showed no linkage drag of the donor fragment on both sides of the marker RM8300 and Sub1BC2 in all five NILs (Near Isogenic Lines) (Figure 6). Also, the GW5 (narrow-grain) carrier chromosome present on the chromosome 5 showed no drag of the donor segment in all pyramided lines. However, a donor segment was detected in between the marker RM28073 and RM28102 in all the pyramided lines (Figure 6).
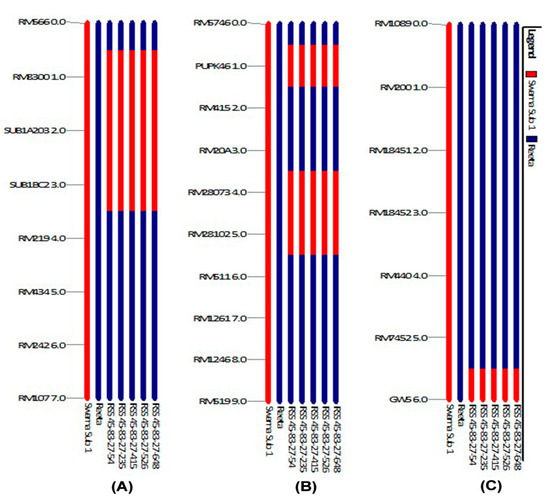
Figure 6.
Analyses of QTLs stacking in carrier chromosomes associated with submergence tolerance, low phosphorus tolerance and yield component QTLs in the 5 pyramided lines (A) Sub1 QTL on the carrier chromosome 9 (B) Pup1 and (C) GW5 (narrow-grain) yield component QTLs on the carrier chromosome 5 present in the BC3F3 progenies of Reeta/Swarna-Sub1. The numbers indicate the pyramided lines, 1. RSS45-83-27-54, 2. RSS45-83-27-235, 3. RSS45-83-27-415, 4. RSS45-83-27-526, and 5. RSS45-83-27-648.
2.6. Evaluation of the Pyramided Lines for Submergence Tolerance
Seven genotypes including five pyramided lines carrying target QTLs were evaluated under the controlled submergence screening tank for confirmation of the submergence tolerance trait in the pyramided lines. Two weeks of complete submergence stress was given to the test genotypes by exposing the materials to 1.5 m water depth after 18 days of transplanting. After one week of de-submergence, all five pyramided lines showed regeneration ability from 85 to 95% while the donor parent ‘Swarna-Sub1’ showed regeneration of 95% (Figure 7). No regeneration was found in the sensitive parent ‘Reeta’. The pyramided lines RSS45-83-27-54, RSS 45-83-27-235, RSS 45-83-27-415, and RSS45-83-27-648 showed high regeneration ability under control testing (Figure 8). The pyramided lines showed high regeneration abilities of about 90% regeneration ability.
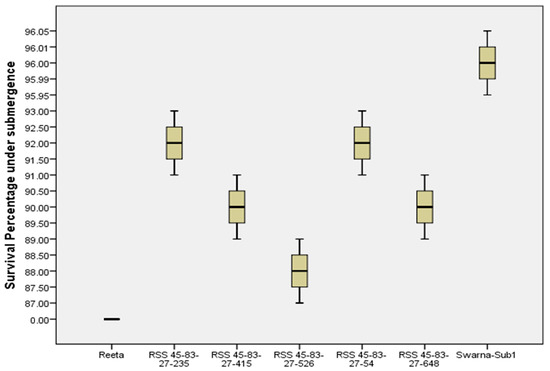
Figure 7.
Percent of plants regenerated in the parental lines Reeta and Swarna-Sub1 and their pyramided lines carrying Sub1 QTL under control screening facility after one week of de-submergence from 14 days of submergence stress.

Figure 8.
Successful introgression of Sub1 and Pup1 QTLs for stress resiliency and increased panicle yield in the flood-prone late-maturing rice using marker-assisted breeding. Photograph shows the panicles of parents, Reeta (right) and Swarna (left) along with the best pyramided lines (middle two) observed during the wet season, 2021.
2.7. Evaluation of the Pyramided Lines under Low Phosphorus Stress
Molecular screening was performed to identify better lines for efficient phosphorous uptake. Both gene-specific as well as flanking markers were used to detect the presence of Pup1 locus in the 5 pyramided lines during screening in the segregating generations (Supplementary Table S1). Swarna-Sub1 and Reeta were used as positive and negative checks. RM28073 and RM28102, the two closest markers to Pup1 QTL [24]; one gene-based marker, Pup1K46; Pup1 negative and positive checks were used in the pot study. The samples at dough stage of the plants were collected and dried in a hot air oven at 80 °C for recording of dry weight. The oven dried plant materials were chopped and grounded in a Willey mill and stored in wide-mouthed Stoppard bottles. After suitable sub-sampling, the samples were analyzed for total phosphorus by Vanadomolybdate yellow color method. The genotypes showing positive response for all the 3 markers employed were taken as positive for Pup1 locus. Based on this criteria, 5 genotypes were observed to be positive for Pup1 QTL (Figure 6). Pup1K46 being a dominant marker associated directly with the PSTOL1 gene showed the expected amplicon of 523bp in both derived and pyramided lines and the tolerant parent used in this study (Supplementary Table S2). The results from the pot study also clearly showed that the pyramided lines carrying Pup1 QTL showed better P-uptake under normal and deficient p-condition. The –ve check lacking the Pup1 QTL showed relatively less uptake of phosphorus in the P-deficient condition than normal condition (Table 2).
Table 2.
Phosphorus uptake (mg/g tissue dry weight) and grain yield of the parental lines Reeta and Swarna-Sub1 and their pyramided lines grown under normal and P-deficient soils.
2.8. Evaluation of the Pyramided Lines for Agro-Morphologic, Yield and Grain Quality Traits
The pyramided lines carrying submergence and low phosphorus tolerance along with yield component QTL in the background of the Reeta variety were evaluated for various traits during the wet seasons in 2020, 2021, and 2022. The pyramided lines were compared with both the parental varieties Reeta and Swarna-Sub1. The recipient parent ‘Reeta’ produced a pooled mean grain yield of 4.38 t/ha under typical shallow lowland conditions. The pyramided lines RSS45-83-27-54, RSS45-83-27-235, RSS45-83-27-415, RSS45-83-27-526, and RSS45-83-27-648 produced more yield than the recipient parent, Reeta (Table 3). However, the agro-morphologic traits of all of the pyramided lines were not similar to that of the parent, Reeta. The target morphologic traits controlled by the yield component QTL viz., no. of primary branches, spikelet fertility, and panicle weight finally influencing grain yield in the pyramided lines were observed to be almost similar within each subcluster (Table 3; Figure 8). Much of the grain quality and the cooking characteristics of the recipient parent such as milling (%), head rice recovery (%), gel consistency, amylose content (%), and alkali spreading value were retained in the pyramided lines similar to the recipient parent (Table 3). The placement pattern of the parents and the pyramided lines in the quadrants of the genotype-by-trait biplot diagram constructed based on 14 agro-morphologic, yield, and component traits over three years showed similarity among the pyramided lines (Supplementary Figure S2). The pyramided lines were found in the first and second quadrants along with the recipient parent ‘Reeta’. The developed lines closer to each other were almost similar in grain yield, grain quality, and the other studied parameters (Supplementary Figure S2). The best-line was evaluated in various parts of the country through All India Coordinated Rice Improvement Programme and showed superiority and stability in performance compared to the check varieties in Odisha state. The variation observed for the first principal component was 61.75%, while 15.05% was explained for the second component.
Table 3.
Agro-morphologic and grain quality traits of the pyramided lines along with parents pooled over 3 seasons.
3. Discussion
Marker-assisted backcrossing (MAB) is a technology far superior over the conventional methods of selection and breeding in precision and efficiency for improvement of user friendly crop plants. Integration of a molecular marker for selection of segregating progenies enhances the accuracy in transfer of a desired trait into a recipient variety through backcross breeding program. This technology helps in identification of recombinants exhibiting the least amount of linkage drag and thus decreases selection time. In the realm of climate change induced disaster-prone rice agro-ecosystem, MAB assumes great significance by decreasing selection times of major stress-resilient gene(s) involved and pyramiding them successfully. In the present study, rice cultivar ‘Reeta’ is a popular variety, but long maturity duration increases its chances of being inundated by flash floods that results in oxidative stress and soil P-deficiency detriment of grain yield. MAB breeding approach adopted herein has achieved a great distinction in developing pyramided-lines, which exhibited submergence and low phosphorus tolerance, and a higher yield without compromising the main features of the farmer-friendly variety. The 3 target QTLs were simultaneously transferred into it. In addition, breeding the variety duration was reduced through MAB compared to the classical backcross breeding approach. The backcrossing was continued up to 3 backcrosses and then one selfing generation were performed to transfer the target QTLs into the variety. Thus, the essential lowland features lacking in variety were improved precisely in less time duration with negligible genetic drag. Variety development through marker-assisted breeding by precise transfer of genes and with a shorter duration has been reported in rice improvement by several workers previously [27,28,29,30,31]. Our work has added a new dimension to these illustrious examples because of the successful introgression of not only stress resiliency but also concomitantly improving grain yield.
Earlier successful gene transfers and pyramiding results in rice were published in rice crop [14,28,29,30,31,32,33,34]. Further, this pyramiding study of QTLs conferring tolerance for submergence and low phosphorus stresses including improvement of yield component QTL was clearly different from earlier gene pyramiding publications. The earlier publications on gene pyramiding of bacterial blight resistance along with submergence tolerance into rice varieties, namely improved Tapaswini and improved Lalat have been reported, but the recipient varieties belonged to the mid-early maturing group and were not suitable for lowlands [35,36]. Hence, yield improvement along with submergence and low phosphorus tolerance through gene pyramiding is a novelty and typical example of gene stacking for a variety of rainfed lowland rice ecology. In contrast to the plethora of publications on the development of cultivars through pyramiding of resistance genes for insects and diseases in rice [29,30,31,32,33,34,35,36,37], the present research targets the development stress tolerance like submergence and P-deficiency tolerance. The pyramided lines carrying the Pup1 locus showed not much difference in P-uptake in the plant tissue under normal and deficient phosphorus in the pots. The –ve check showed higher P-uptake under normal P-level but low uptake under deficient conditions (Table 2). Such a result confirms expectations of differential response between the low P-tolerant and P-susceptible varieties. There are instances of a similar correlation between the Pup1 allele expression and P-uptake in studies reported earlier [20,22].
Majority of the crop breeding strategies for biotic and abiotic stress tolerance are based on a single resistant gene introgression into plants and they are short-lived. By contrast, pyramidization of multiple resistant genes into a single plant from different sources is durable and ensures stability. Besides, the success of trait correction is achieved in the shortest period of time for agricultural sustainability, as cited in previous publications [14,32,33,34,35,36,37]. However, in the present work, the developed pyramided lines carrying the target QTLs viz., Sub1+ Pup1 + GW5 along with recipient parents’ genome content of >95% were clearly different from what was reported previously. It is possible that an undesirable drag from the donor genome may come in progenies as additional unlinked loci in the backcross generations [31]. In our investigation, such effects were detected but from the elite donor source into pyramided lines while transferring the Sub1, Pup1 and GW5 QTLs into the Reeta variety background. The linkage drags are depicted in the graphical representation of genotyping data on the chromosome carrying the target QTLs (Figure 7). A very low linkage drag was noticed while transferring 3 target QTLs because the donor parent was an elite variety. In previous studies also low linkage drag was noticed while using elite donor sources and using more background markers [29,30,34]. The donor parent used in this study was Swarna-Sub1 which is a popular variety, and therefore, the expected drag may not show any undesirable effects in the pyramided lines (Figure 7). Similar findings were also reported by many workers which recommend the use of an improved variety as the donor that results in less or no undesirable drag compared to the wild and landraces sources [28,29,30,35,36,37,38,39,40].
In fact, there were a few pyramided lines showing similarity to the recipient parent in many traits. The dendrogram generated based on the traits indicated grouping of the pyramided and parental lines into mainly 2 clusters with similarity within the clusters (Figure 6A). The pyramided and recipient parents were found in quadrants I and II in the biplot diagram drawn based on 14 studied morpho-quality traits, showing minor variations among the pyramided lines (Supplementary Figure S2). Evaluation of the pyramided lines for yield and grain quality traits showed higher yields in pyramided lines viz., RSS45-83-27-54, RSS 45-83-27-235, RSS 45-83-27-415, RSS 45-83-27-526, and RSS45-83-27-648 than the recipient parent, Reeta (Table 3). The transfer of traits and achieving similar or better yield in the pyramided lines were also reported earlier in some gene-pyramiding publications [1,28,29,30,35,36,37,38,39,40].
In our work, the pyramided lines were closer to each other with the recipient parent, Reeta in the trait-biplot diagram, while the donor parent was far away and placed in another quadrant. Therefore, the similarity of the pyramided and recipient lines was quite similar and no linkage drag was observed from the donor parent in the transfer of the target genes. In addition, the yield, quality, and other morphological traits in a few pyramided lines were better than the recipient parent (Table 3). It is clear from the genotyping results of background analysis that there was an accelerated recovery of the recipient parent’s genome in a few pyramided lines than the expected value in the backcross generations. Also, it revealed that the transfer of Sub1, Pup1 and GW5 QTLs into one genetic background may not show antagonistic effects for yield and related traits.
4. Materials and Methods
4.1. Plant Materials and Breeding Program
In the breeding program, Swarna-Sub1variety bearing Sub1, Pup1, and GW5 QTLs, with agronomic traits submergence tolerance, low P (phosphorus) deficiency-tolerance, and grain size respectively, was used as the donor male parent. The recipient parent, Reeta (CR Dhan 401) was a high-yielding variety, but vulnerable to submergence stress. The varieties were obtained from the gene bank of the ICAR-National Rice Research Institute, Cuttack, India, and grown in the screening tanks and crossed in the wet season of 2017 as per the scheme depicted for marker-assisted breeding (Figure 9). According to the protocol, one true F1 plant was hybridized with the recipient parent during the dry season of 2018 to generate BC1F1 generation seeds. True hybridity was checked using the direct Sub1-marker, Sub1A203, and a co-dominant marker, RM8300. The BC1F1 seeds were grown, and the progenies were screened for the target genes for submergence tolerance (Sub1), P-deficiency tolerance (Pup1), and grain size (GW5) by using the established molecular markers (Supplementary Table S1). A selection was performed to get the Swarn-Sub1 grain width (GW5). In the background selection, progenies of the BC1F1 generation carrying the 3 target QTLs were screened using the polymorphic markers. Among the foreground positive progenies, the lines containing highest genome of the recurrent parent was hybridized with the recipient parent, Reeta to get BC2F1 seeds. BC2F1 seeds were harvested during the dry season of 2019. The background analysis of the BC3F1 progenies was performed during the same season. The BC3F1 plant population containing the highest recipient genome content along with two major target QTLs Sub1 and Pup1 were self-bred during the wet season of 2019. The BC3F2 progenies were genotyped to search for the presence of homozygosity for the two major target QTLs and the recipient parent’s GW5 QTL during the dry season of 2020. Foreground positive plants exhibiting higher panicle weight, grain number and panicle branching were also checked in each segregating generation. Phenotyping of these QTLs was performed and evaluated during the wet seasons in 2020 and repeated in the years 2021, and 2022.
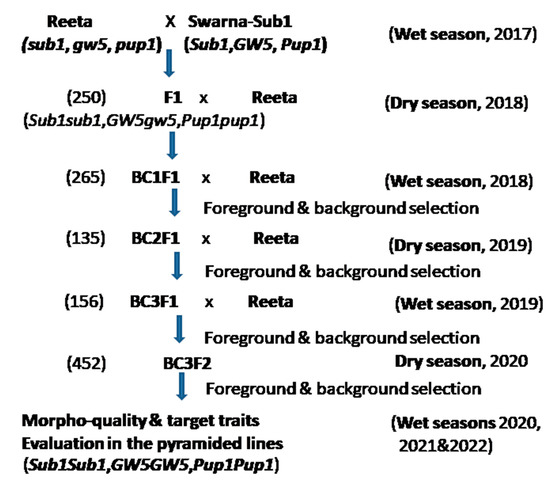
Figure 9.
The marker-assisted breeding scheme used for the development of pyramided lines for screening of submergence, low phosphorus stress tolerance, and panicle yield component QTLs. The numerical figures within parentheses indicate the number of hybrids/lines raised in the generation.
4.2. Genomic DNA Isolation, Polymerase Chain Reaction, and Marker Analysis
Genomic DNA content was isolated following the standard extraction protocol [41]. PCR reaction was performed following the procedure used in our previous publications [3,42,43]. The information regarding chromosome number, position, and sequence of the primers used in the polymerase chain reaction are presented in Supplementary Table S1. Seven gene-specific and tightly linked markers for the two target QTLs and four recipient QTLs were used in foreground selection (Supplementary Table S1). The markers used in this study were taken from earlier publications [6,9,17,20,21,25,26]. A total of 728 SSR markers available in public domain were used for the study of polymorphism between the two parents. The polymorphic markers detected were used for background selection (Supplementary Table S2). Agarose gel electrophoresis was used to segregate the amplification products obtained from PCR reactions. The images were recorded in a gel documentation system (SynGene, Cambridge, UK). Data analysis and dendrogram construction were performed following the standard publications [44,45,46]. Graphical Geno Types (GGT) Version 2.0 software was used to construct the genome recovery graph of the recipient parent in the pyramided lines based on the SSR marker data [47].
4.3. Screening for Submergence Tolerance
The BC3F4 generation pyramided lines and parents were transplanted (3 weeks old seedlings) in the screening tank of ICAR-NRRI, Cuttack, during the wet seasons in 2020 and 2021. The screening trial was laid out in a randomized complete block design (RBD) with three replications/entries accommodating a population size of 66 plants/entry. The experiment materials were transplanted at a spacing of 15 × 20 cm2 by providing three rows/entry. Two weeks of complete submergence stress was given by exposing the materials to 1.5 m water depth after 18 days of transplanting. De-submergence was performed just after the completion of the 14-day stress period, and subsequently, regeneration was assessed one week after de-submergence. The data recording and scoring of the genotypes were performed following the standard procedures [17].
4.4. Phenotyping for Phosphorus Uptake
The pyramided and parental lines were evaluated under deficient and normal soil phosphorous in the pot experiment. The experiment was laid out in a randomized block design with 3 replications for both phosphorus levels. In the deficient condition, the P-content of the collected low-P upland soil was 6.5 mg/kg while the P-content in the normal soil was 13.3 mg/kg after the enrichment of single super phosphate fertilizer. Five genotypes observed to be homozygous for Sub1, Pup1, and GW5 QTLs along with their two parents were grown in the pots. Nitrogen was applied in three equal splits viz., basal, active tillering, and panicle initiation. A full dose of phosphorus and potassium was applied as a basal application. Seeds were direct seeded in thepots and after germination thinned later to maintain two seedlings per pot. The samples at dough stage of the plants were collected and dried in a hot air oven at 80°C for recording of dry weight. The oven-dried plant materialswere chopped and ground in a Willey mill and stored in wide-mouthed Stoppard bottles. After suitable sub-sampling, the samples were analyzed for total phosphorus. One gram of powdered sample was taken and subjected to triacid acid (nitric acid (ISOCHEM): sulphuric acid (ISOCHEM): perchloric acid (SDFCL) of 3:2:1 ratio) digestion. The solution was filtered and the volume was made up to 100 mL using distilled water. Five ml of the triacid extract was pipetted out into a 25 mL volumetric flask. Five ml of Barton’s reagent was added and the volume was made up to mark with distilled water. The development of the yellow color was observed after 30 min and the intensity of color was measured in a photoelectric colorimeter using a blue filter (470 nm) after adjusting the transmittance of the meter to 100 with a blank. The color was stable for 24 h. The concentration of phosphorus in the solution was deduced from the standard curve from which the percentage of phosphorus content of the sample was calculated.
4.5. Evaluation of the Pyramided Lines
The seedlings of 25-day-old pyramided Reeta background lines carrying Sub1, Pup1, and yield QTLs were transplanted along with the parents during the wet seasons in 2020, 2021, and 2022. A plot size of 12 m2 was provided for each entry, with 40 plants per row, at a spacing of 15 × 20 cm2, and planted in RBD with three replications in the research farm of NRRI, Cuttack. The data for ten plants for morpho-quality traits viz., plant height, panicles/plant, panicle weight (g), number of filled grains, total spikelets, number of primary branches, secondary branches, and number of tertiary branches per panicle, grain length, grain breadth, 1000-grain weight, milling (%), head rice recovery (%), and amylose content (%) from each entry and replications were recorded. Plot yield and days to 50% flowering were recorded on a whole plot basis. The standard protocols for head rice recovery and gel consistency [48] were adopted. Amylose content in the grains of the pyramided and parental lines was estimated following the standard procedure [49].
4.6. Statistical Analysis
The recorded morpho-quality traits of the pyramided and parental lines were analyzed using SAS 2008, version 9.2 [50]. The Principal Component Analysis (PCA) for the pyramided and parental lines was performed by using multivariate analysis (Past Software version 4.03) data of the 15 morphological traits. A scatter plot was generated by using two major components: Principal Component 1 (PC1) and Principal Component 2 (PC2). The Eigen value and percentage of variance were generated by the interaction of a variance–covariance matrix. The interaction between all morphological traits was depicted through a biplot graph in the matrix. All the plots and results of PCA were generated as per the standard procedure followed in earlier publications [51,52,53,54].
5. Conclusions
The near-isogenic lines, RSS45-83-27-54, RSS 45-83-27-235, RSS 45-83-27-415, RSS 45-83-27-526, and RSS45-83-27-648 carrying Sub1, Pup1 and GW5 were submergence and low-P stress tolerant with higher yield than the recipient parent, Reeta. The higher yield recorded from the pyramided lines might be due to the accumulation of additional yield and stress tolerance QTLs. In addition, no yield penalty happened due to the interaction of these QTLs in the pyramided background. The grain quality and the cooking characteristics of the recipient parent such as milling %, head rice recovery %, gel consistency, amylose content (%), and alkali spreading value along with yielding ability were retained in a few pyramided lines. Hence, the elite pyramided lines in the background of the popular variety ‘Reeta’ may serve as potential donors of QTLs possessing Sub1 + Pup1 + GW5 in future breeding programs. Based on our innovative research, a promising near-isogenic line has been released in the name of Reeta-Panidhan (CR Dhan 413) for cultivation in the flood-prone areas of Odisha state. This study established the application of marker-assisted selection for transferring abiotic stress tolerance and for enhancing yield in rice.
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241310708/s1.
Author Contributions
S.R.B., A.M. (Arpita Moharana), E.P., A.B., S.M. and A.M. (Ankita Mishra), performed the genotyping work; S.P.M., J.M., P.S., R.R.K.R., D.R.P. and S.K.P. contributed in hybridization and phenotypic selection. A.B., S.P.M., C.R.S. and S.M. recorded the phenotypic data. S.K.P. contributed to the planning, designing, screening, evaluation, supervising and writing of the article. V.P.B. and S.D. contributed to data analysis and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The Institute’s internal funding (Project 1.6) was used for completion of the research work.
Data Availability Statement
The original contributions presented in the study are included in the article.
Acknowledgments
The authors acknowledge the support of Head, Crop Improvement Division, ICAR-National Rice Research Institute for providing all the necessary facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Bharati, B.; Chatopadhyay, K.; Singh, S.; Dash, P.; Reddy, J.N. Association mapping reveals multiple QTLs for grain protein content in rice useful for biofortification. Mol. Genet. Genom. 2019, 294, 963–983. [Google Scholar] [CrossRef] [PubMed]
- Sanghamitra, P.; Nanda, N.; Barik, S.R.; Sahoo, S.; Pandit, E.; Bastia, R.; Bagchi, T.B.; Pradhan, S.K. Genetic structure and molecular markers-trait association for physiological traits related to seed vigour in rice. Plant Gene 2021, 28, 100338. [Google Scholar] [CrossRef]
- Bastia, R.; Pandit, E.; Sanghamitra, P.; Barik, S.; Nayak, D.K.; Sahoo, A.; Moharana, A.; Meher, J.; Dash, P.K.; Raj, R. Association Mapping for Quantitative Trait Loci Controlling Superoxide Dismutase, Flavonoids, Anthocyanins, Carotenoids, γ-Oryzanol and Antioxidant Activity in Rice. Agronomy 2022, 12, 3036. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Rice Market Monitor; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017; Volume 20, pp. 1–38. [Google Scholar]
- Dar, M.H.; Waza, S.A.; Shukla, S.; Zaidi, N.W.; Nayak, S.; Hossain, M.; Singh, U.S. Drought tolerant rice for ensuring food security in Eastern India. Sustainability 2020, 12, 2214. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Barik, S.R.; Sahoo, J.; Pandit, E.; Nayak, D.K.; Pani, D.R.; Anandan, A. Comparison of Sub1 markers and their combinations for submergence tolerance and analysis of adaptation strategies of rice in rainfed lowland ecology. C. R. Biol. 2015, 338, 650–659. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Chakraborti, M.; Chakraborty, K.; Behera, L.; Meher, J.; Subudhi, H.N.; Mishra, S.K.; Pandit, E.; Reddy, J.N. Genetic Improvement of Rainfed Shallow-Lowland Rice for Higher Yield and Climate Resilience. In Rice Research for Enhancing Productivity, Profitability and Climate Resilience; Pathak, H., Nayak, A.K., Jena, M., Singh, O.N., Samal, P., Sharma, S.G., Eds.; ICAR-National Rice Research Institute: Cuttack, India, 2018; pp. 107–121. Available online: https://icar-nrri.in/wp-content/uploads/2019/02/Rice_Research_book_nrri.pdf (accessed on 7 February 2018).
- Mohapatra, S.; Panda, A.K.; Bastia, A.K.; Mukherjee, A.K.; Sanghamitra, P.; Meher, J.; Mohanty, S.P.; Pradhan, S.K. Development of Submergence-Tolerant, Bacterial Blight-Resistant, and High-Yielding Near Isogenic Lines of Popular Variety, ‘Swarna’ Through Marker-Assisted Breeding Approach. Front. Plant Sci. 2021, 12, 672618. [Google Scholar] [CrossRef]
- Pandit, E.; Pawar, S.; Barik, S.R.; Mohanty, S.P.; Meher, J.; Pradhan, S.K. Marker-Assisted Backcross Breeding for Improvement of Submergence Tolerance and Grain Yield in the Popular Rice Variety ‘Maudamani’. Agronomy 2021, 11, 1263. [Google Scholar] [CrossRef]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 1996, 2, 219–224. [Google Scholar] [CrossRef]
- Iftekharuddaula, K.M.; Newaz, M.A.; Salam, M.A.; Ahmed, H.U.; Mahbub, M.A.A.; Septiningsih, E.M.; Collard, B.C.Y.; Sanchez, D.L.; Pamplona, A.M.; Mackill, D.J. Rapid and high-precision marker assisted backcrossing to introgress the SUB1 QTL into BR11, the rainfed lowland rice mega variety of Bangladesh. Euphytica 2011, 178, 83–97. [Google Scholar] [CrossRef]
- Manivong, P.; Korinsak, S.; Korinsak, S.; Siangliw, J.L.; Vanavichit, A.; Toojinda, T. Marker-assisted selection to improve submergence tolerance, blast resistance and strong fragrance in glutinous rice. Genom. Genet. 2014, 7, 110–122. [Google Scholar]
- Khush, G.S.; Mackill, D.J.; Sidhu, G.S. Breeding Rice for Resistance to Bacterial Leaf Blight; IRRI: Manila, Philippines, 1989; pp. 207–217. [Google Scholar]
- Pradhan, S.K.; Barik, S.R.; Nayak, D.K.; Pradhan, A.; Pandit, E.; Nayak, P.; Das, S.R.; Pathak, H. Genetics, Molecular Mechanisms and Deployment of Bacterial Blight Resistance Genes in Rice. Crit. Rev. Plant Sci. 2020, 39, 360–385. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Pandit, E.; Barik, S.R.; Mohanty, S.P.; Nayak, D.K.; Sah, R.P.; Behera, L.; Sanghamitra, P.; Bose, L.K.; Das, S.R. Climate-Smart Rice Breeding: Progress and Challenges for the Rain-fed ecologies in India. In Advances in Rice Breeding: Stress Tolerance, Climate Resilience, Quality and High Yield; ICAR-NRRI: Cuttack, India, 2021; pp. 144–162. [Google Scholar]
- Das, S.R.; Collard, B.C.Y.; Pradhan, S.K. Breeding for climate resilient varieties suitable for rainfed rice ecologies. In Recent Innovations and Emerging Technologies for Transforming Rice Farming; ICAR-NRRI: Cuttack, India, 2023. [Google Scholar]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Baksh, S.Y.; Mukherjee, A.K.; Mohanty, S.P. Development of flash-flood tolerant and durable bacterial blight resistant versions of mega rice variety ‘Swarna’ through marker-assisted backcross breeding. Sci. Rep. 2019, 9, 12810. [Google Scholar] [CrossRef] [PubMed]
- Batjes, N.H. A world dataset of derived soil properties by FAO–UNESCO soil unit for global modelling. Soil Use Manag. 1997, 13, 9–16. [Google Scholar] [CrossRef]
- Motsara, M.R. Available nitrogen, phosphorus and potassium status of Indian soils as depicted by soil fertility maps. Ferilizer News 2002, 47, 15–21. [Google Scholar]
- Pandit, E.; Panda, R.K.; Pani, D.R.; Chandra, R.; Singh, S.; Pradhan, S.K. Molecular marker and phenotypic analyses for low phosphorus stress tolerance in cultivars and landraces of upland rice under irrigated and drought situations. Indian J. Genet. Plant Breed. 2018, 78, 59–68. [Google Scholar] [CrossRef]
- Pandit, E.; Sahoo, A.; Panda, R.K.; Mohanty, D.P.; Pani, D.R.; Anandan, A.; Pradhan, S.K. Survey of rice cultivars and landraces of upland ecology for phosphorus uptake 1 (Pup1) QTL using linked and gene specific molecular markers. Oryza 2016, 53, 1–9. [Google Scholar]
- Wissuwa, M.; Wegner, J.; Ae, N.; Yano, M. Substitution mapping of Pup1: A major QTL increasing phosphorus uptake of rice from a phosphorus deficient soil. Theor. Appl. Genet. 2002, 105, 890–897. [Google Scholar] [CrossRef]
- Gamuyao, R.; Chin, J.; Tanaka, J.; Pesaresi, P.; Catausan, S.; Dalid, C.; Loedin, I.; Mendoza, E.; Wissuwa, M.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–539. [Google Scholar] [CrossRef]
- Heuer, S.; Lu, X.; Chin, J.H.; Tanaka, J.P.; Kanamori, H.; Matsumoto, T.; De Leon, T.; Ulat, V.J.; Ismail, A.M.; Yano, M. Comparative sequence analyses of the major quantitative trait locus phosphorus uptake 1 (Pup1) reveal a complex genetic structure. Plant Biotechnol. J. 2009, 7, 456–457. [Google Scholar] [CrossRef]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, M.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing Rice with High Yield under Phosphorus Deficiency: Pup1 Sequence to Application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef]
- Weng, J.; Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X.; et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008, 18, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Behera, L.; Sekhar, S.; Mohanty, S.; Devanna, B.N.; Parameswaran, C.; Pradhan, S.K. Genomics and other omics approaches for rice improvement. In Advances in Rice Breeding: Stress Tolerance, Climate Resilience, Quality and High Yield; ICAR-NRRI: Cuttack, India, 2021; pp. 369–426. [Google Scholar]
- Pradhan, S.K.; Nayak, D.K.; Mohanty, S.; Behera, L.; Barik, S.R.; Pandit, E.; Lenka, S. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna. Rice 2015, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, K.C.; Pandit, E.; Mohanty, S.P.; Moharana, A.; Sanghamitra, P.; Meher, J.; Jena, B.K.; Dash, P.K.; Behera, L.; Mohapatra, P.M.; et al. Development of Broad Spectrum and Durable Bacterial Blight Resistant Variety through Pyramiding of Four Resistance Genes in Rice. Agronomy 2022, 12, 1903. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, N.S.; Sarma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2018, 160, 411–422. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Nayak, D.K.; Pandit, E.; Behera, L.; Anandan, A.; Mukherjee, A.K.; Lenka, S.; Barik, D.P. Incorporation of bacterial blight resistance genes into lowland rice cultivar through marker-assisted backcross breeding. Phytopathology 2016, 106, 710–718. [Google Scholar] [CrossRef]
- Singh, S.; Sidhu, J.S.; Huang, N.; Vikal, Y.; Li, Z.; Brar, D.S.; Dhaliwal, H.S.; Khush, G.S. Pyramiding three bacterial blight resistance genes(xa-5, xa-13 and Xa-21) using marker-assisted selection into indica rice cultivar PR-106. Theor. Appl. Genet. 2001, 102, 1011–1015. [Google Scholar] [CrossRef]
- Angeles-Shim, R.B.; Reyes, V.P.; del Valle, M.M.; Lapis, R.S.; Shim, J.; Sunohara, H.; Doi, K. Marker-assisted introgression of quantitative resistance gene pi21 confers broad spectrum resistance to rice blast. Rice Sci. 2020, 27, 113–123. [Google Scholar] [CrossRef]
- Nayak, D.K.; Pandit, E.; Mohanty, S.; Barik, D.P.; Pradhan, S.K. Marker-assisted selection in back cross progenies for transfer of bacterial leaf blight resistance genes into a popular lowland rice cultivar. Oryza 2015, 52, 163–172. [Google Scholar]
- Das, G.; Rao, G.J.N. Molecular marker assisted gene stacking for biotic and abiotic stress resistance genes in an elite rice cultivar. Front. Plant Sci. 2015, 6, 698. [Google Scholar] [CrossRef]
- Das, G.; Rao, G.J.N.; Varier, V.; Prakash, A.; Dokku, P. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef]
- Pradhan, K.C.; Barik, S.R.; Mohapatra, S.; Nayak, D.K.; Pandit, E.; Jena, B.K.; Sangeeta, S.; Pradhan, A.; Samal, A.; Meher, J.; et al. Incorporation of Two Bacterial Blight Resistance Genes into the Popular Rice Variety, Ranidhan through Marker-Assisted Breeding. Agriculture 2022, 12, 1287. [Google Scholar] [CrossRef]
- Mohapatra, S.; Barik, S.R.; Dash, P.K.; Lenka, D.; Pradhan, K.C.; Reshmi Raj, K.R.; Mohanty, S.P.; Mohanty, M.R.; Sahoo, A.; Jena, B.K.; et al. Molecular Breeding for Incorporation of Submergence Tolerance and Durable Bacterial Blight Resistance into the Popular Rice Variety ‘Ranidhan’. Biomolecules 2023, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.K.; Nayak, D.K.; Pandit, E.; Barik, S.R.; Mohanty, S.P.; Anandan, A.; Reddy, J.N. Characterization of morpho-quality traits and validation of bacterial blight resistance in pyramided rice genotypes under various hot spots of India. Aust. J. Crop Sci. 2015, 9, 127–134. [Google Scholar]
- Behera, L.; Parameswaran, C.; Anandan, A.; Sanghamitra, P.; Pradhan, S.K.; Jena, M.; Umakanta, N.; Dash, S.K.; Swain, P.; Sahu, R.K.; et al. Development of genomic resources for rice improvement. In Rice Research for Enhancing Productivity, Profitability and Climate Resilience; Pathak, H., Nayak, A.K., Jena, M., Singh, O.N., Samal, P., Sharma, S.G., Eds.; ICAR-National Rice Research Institute: Cuttack, India, 2018; pp. 197–223. Available online: https://icar-nrri.in/wp-content/uploads/2019/02/Rice_Research_book_nrri.pdf (accessed on 24 March 2023).
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA mini preparation: Version II. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Barik, S.R.; Pandit, E.; Mohanty, S.P.; Nayak, D.K.; Pradhan, S.K. Genetic mapping of physiological traits associated with terminal stage drought tolerance in rice. BMC Genet. 2020, 21, 76. [Google Scholar] [CrossRef]
- Pandit, E.; Panda, R.K.; Sahoo, A.; Pani, D.R.; Pradhan, S.K. Genetic relationship and structure analysis of root growth angle for improvement of drought avoidance in early and mid-early maturing rice genotypes. Rice Sci. 2020, 27, 124–132. [Google Scholar] [CrossRef]
- Sanghamitra, P.; Barik, S.R.; Bastia, R.; Mohanty, S.P.; Pandit, E.; Behera, A.; Mishra, J.; Kumar, G.; Pradhan, S.K. Detection of Genomic Regions Controlling the Antioxidant Enzymes, Phenolic Content, and Antioxidant Activities in Rice Grain through Association Mapping. Plants 2022, 11, 1463. [Google Scholar] [CrossRef]
- Barik, S.R.; Pandit, E.; Pradhan, S.K.; Singh, S.; Swain, P.; Mohapatra, T. QTL mapping for relative water content trait at reproductive stage drought stress in rice. Indian J. Genet. 2018, 78, 401–408. [Google Scholar]
- Pawar, S.; Pandit, E.; Mohanty, I.C.; Saha, D.; Pradhan, S.K. Population genetic structure and association mapping for iron toxicity tolerance in rice. PLoS ONE 2021, 16, e0246232. [Google Scholar] [CrossRef]
- Van Berloo, R. GGT: Software for display of graphical genotypes. J. Hered. 1999, 90, 328–330. [Google Scholar] [CrossRef]
- Cagampang, G.B.; Perez, C.M.; Juliano, B.O. A gel consistency test for eating quality in rice. J. Sci. Food Agric. 1973, 24, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. Rice quality screening with the Rapid ViscoAnalyser. Appl. Rapid Visco Anal. 1996, 19–24. [Google Scholar]
- SAS Institute Inc. SAS® 9.2 Language Reference: Concepts, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2010; pp. 626–978. [Google Scholar]
- Pandit, E.; Tasleem, S.; Nayak, D.K.; Barik, S.R.; Mohanty, D.P.; Das, S.; Pradhan, S.K. Genome-wide association mapping reveals multiple QTLs governing tolerance response for seedling stage chilling stress in indica rice. Front. Plant Sci. 2017, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Naveenkumar, R.; Barik, S.R.; Mohanty, S.P.; Nayak, D.K.; Ghritlahre, S.K.; Rao, D.S.; Reddy, J.N.; et al. Linkage disequilibrium mapping for grain Fe and Zn enhancing QTLs useful for nutrient dense rice breeding. BMC Plant Biol. 2020, 20, 57. [Google Scholar] [CrossRef] [PubMed]
- Nayak, D.K.; Sahoo, S.; Barik, S.R.; Sanghamitra, P.; Sangeeta, S.; Pandit, E.; Reshmi Raj, K.R.; Basak, N.; Pradhan, S.K. Association mapping for protein, total soluble sugars, starch, amylose and chlorophyll content in rice. BMC Plant Biol. 2022, 22, 620. [Google Scholar] [CrossRef]
- Mohanty, S.P.; Nayak, D.K.; Sanghamitra, P.; Barik, S.R.; Pandit, E.; Behera, A.; Pani, D.R.; Mohapatra, S.; Reshmi Raj, K.R.; Pradhan, K.C.; et al. Mapping the Genomic Regions Controlling Germination Rate and Early Seedling Growth Parameters in Rice. Genes 2023, 14, 902. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).