
Repurposing Simvastatin in Parkinson’s Disease Model: Protection Is throughout Modulation of the Neuro-Inflammatory Response in the Substantia nigra

 and
and 
Abstract
1. Introduction
2. Results
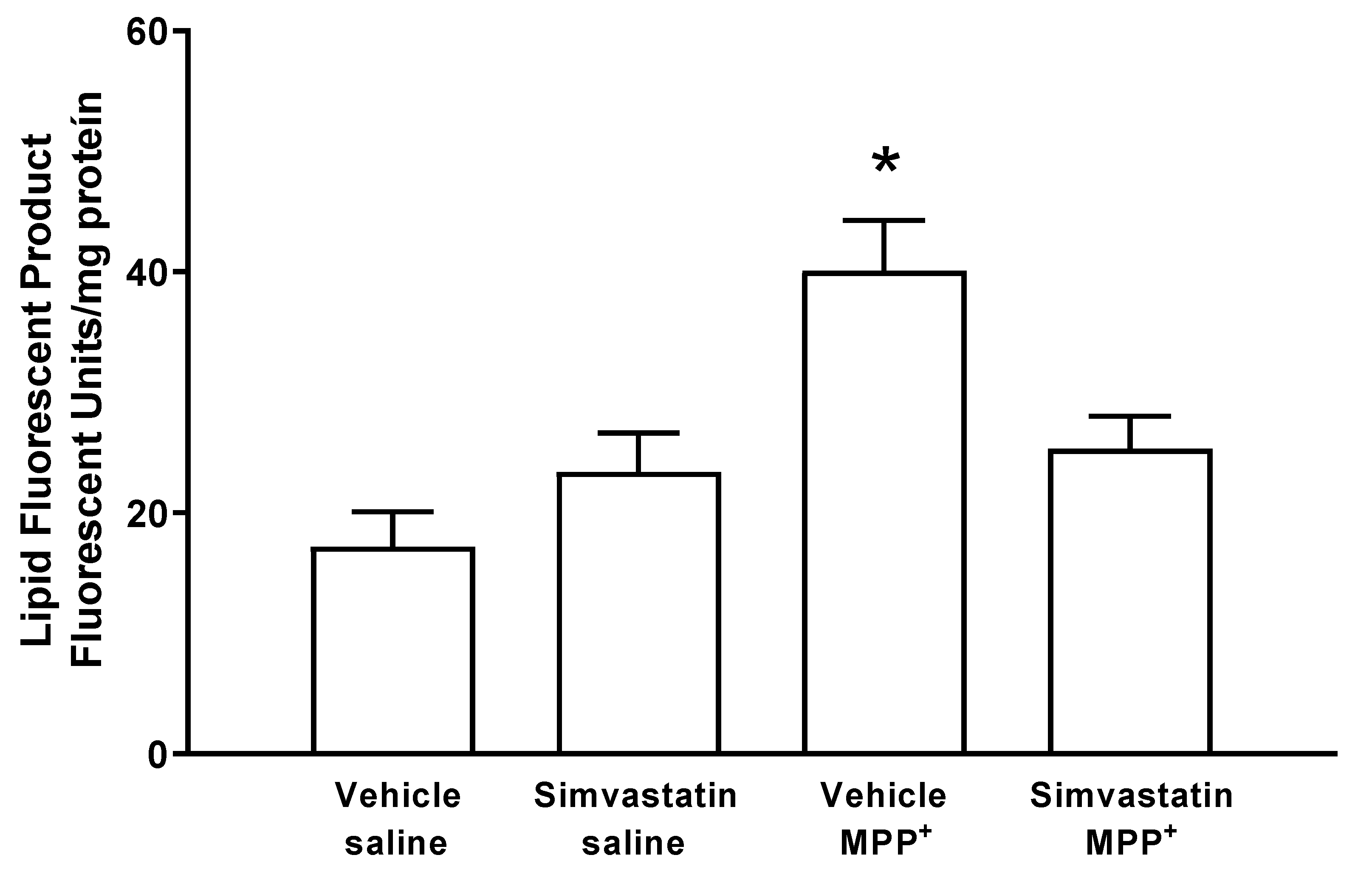
2.1. Fluorescent Lipidic Products
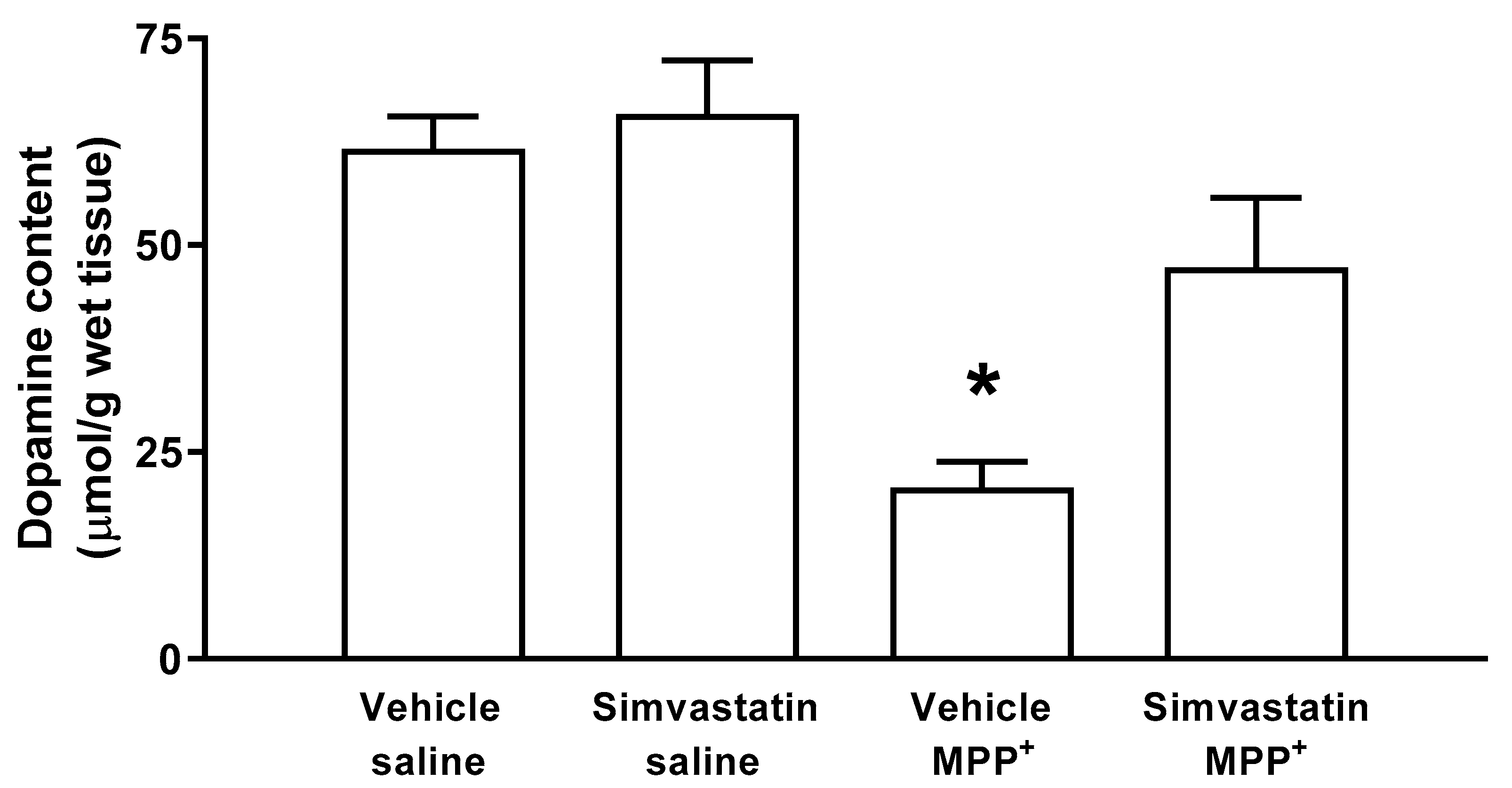
2.2. DOPAMINE (DA) Content in Striatal Tissue
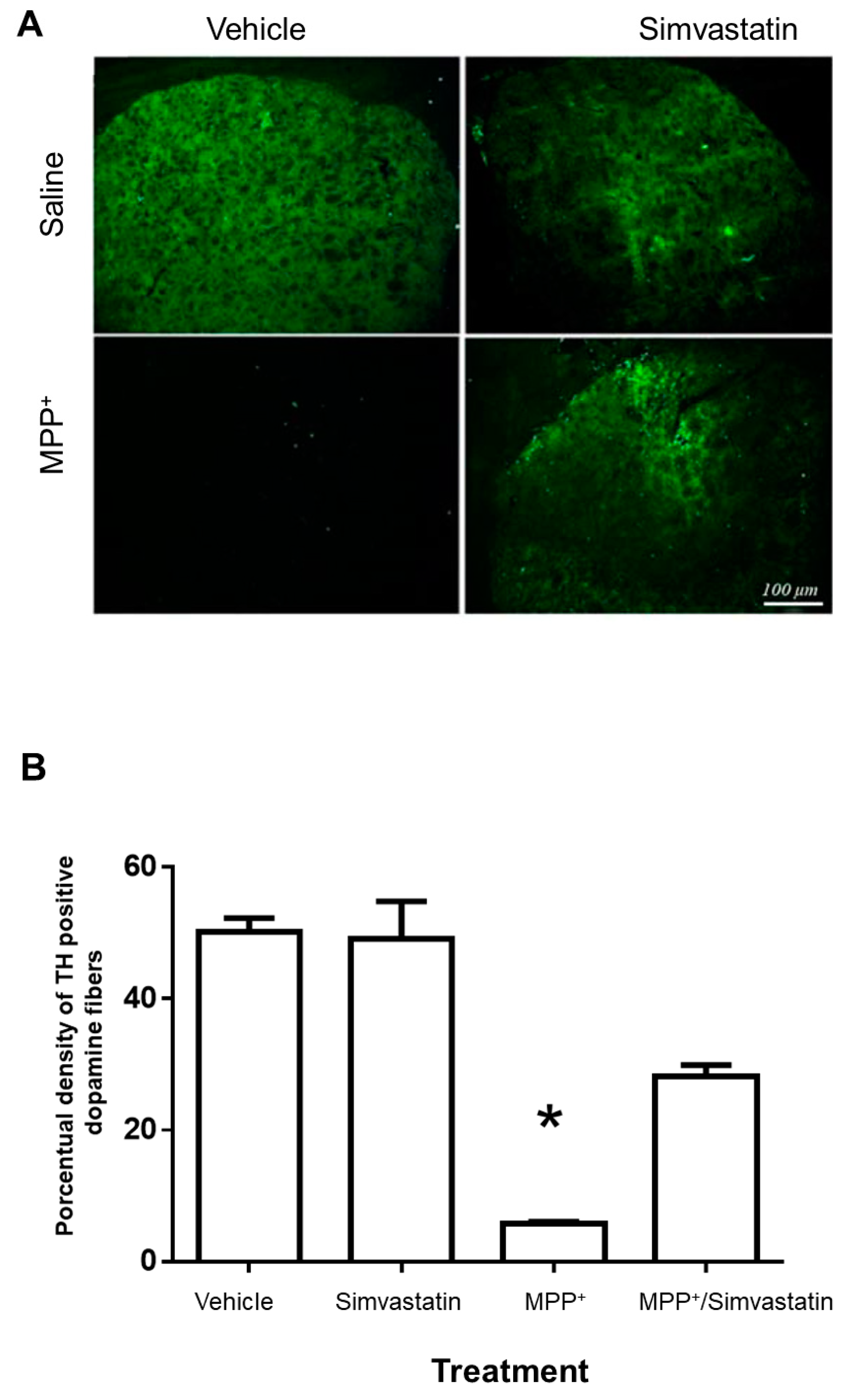
2.3. Neurodegenerative Damage
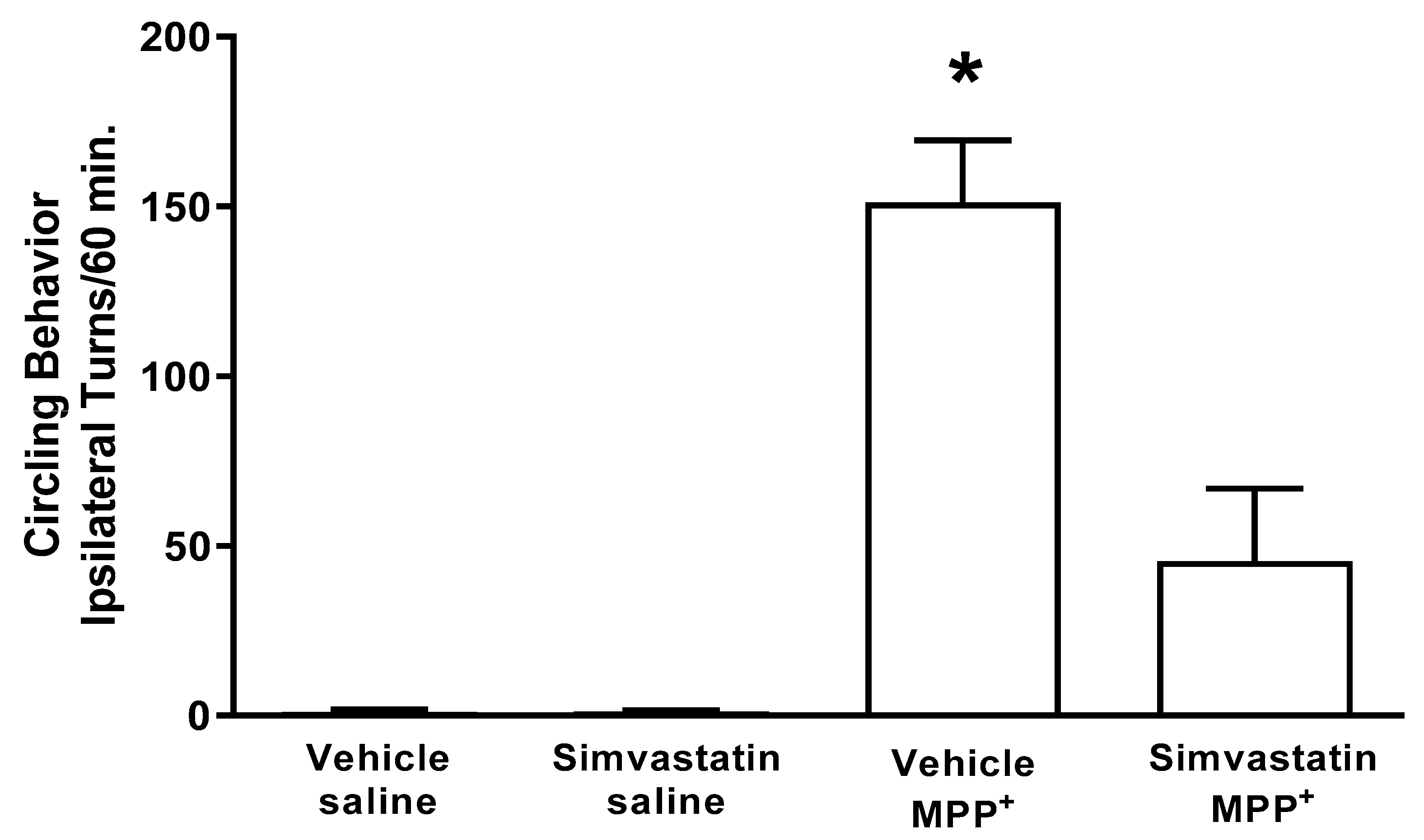
2.4. Circling Behavior
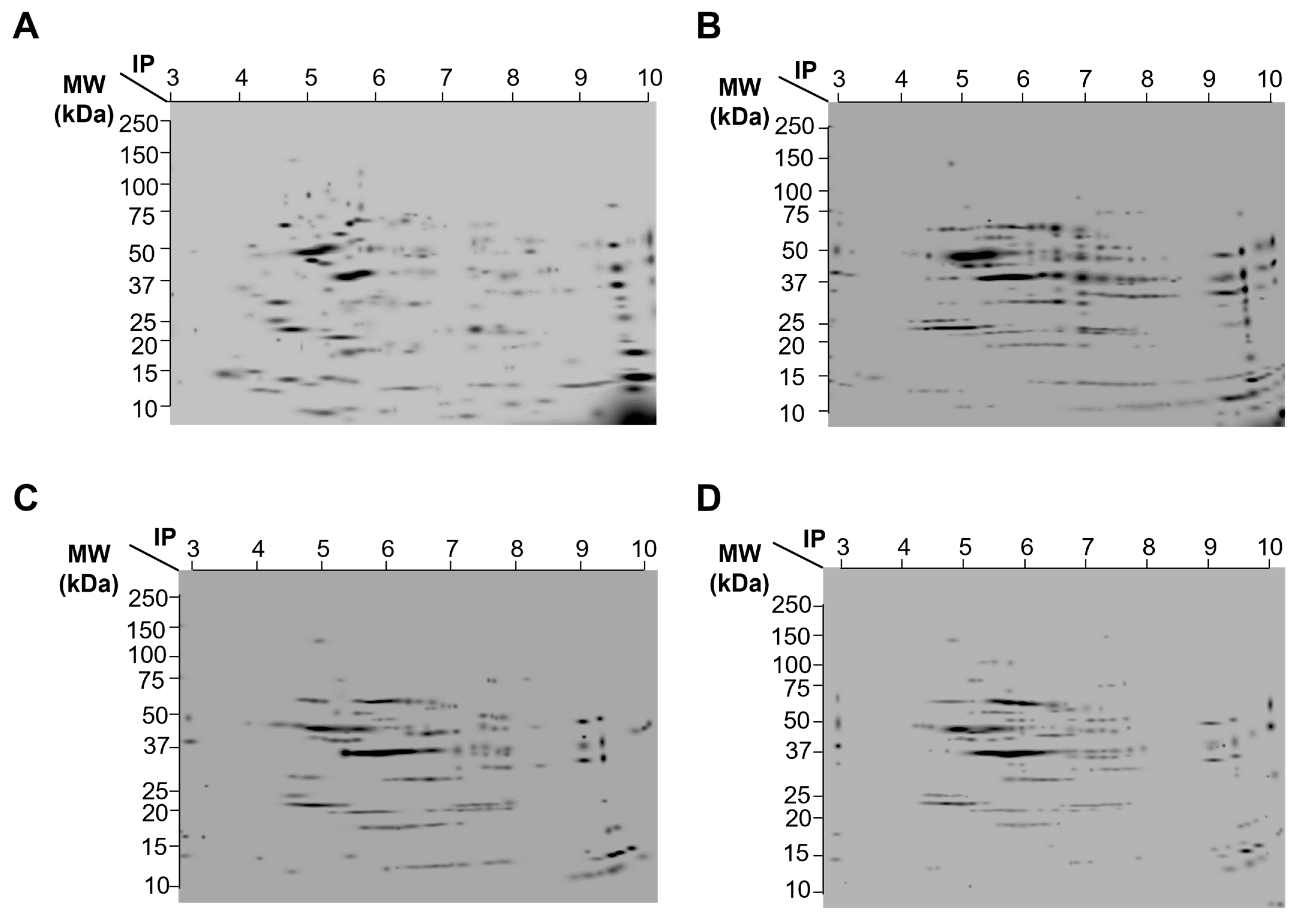
2.5. Protein Expression Patterns from Striatum from Control, Simvastatin, MPP+, and Simvastatin-MPP+-Treated Rats by 2D-SDS-Page
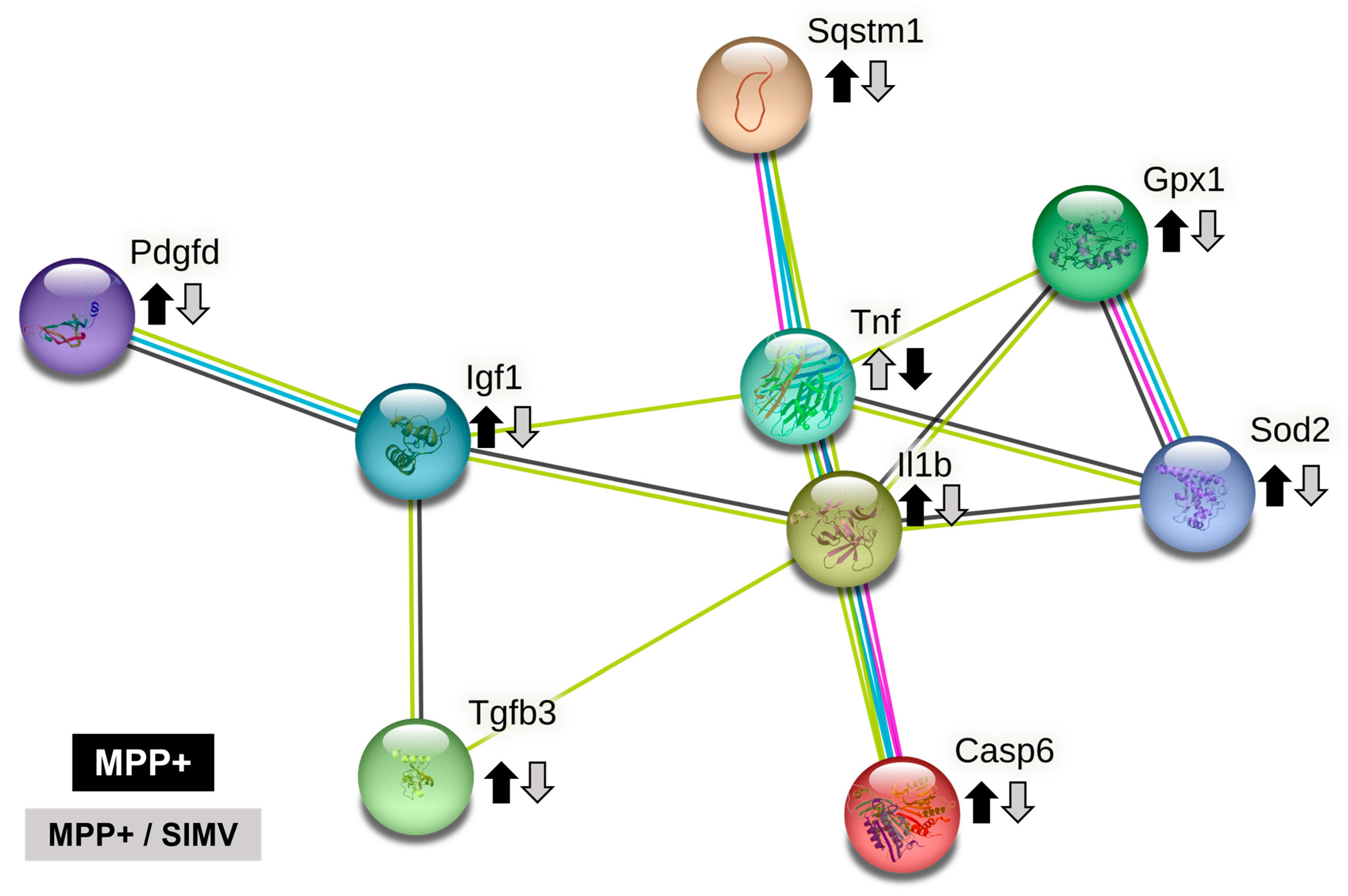
2.6. Bioinformatic Analysis
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Treatments
4.3. Striatal Dopamine Levels Measurement
4.4. Fluorescent Lipidic Products Assay
4.5. Circling Behavior
4.6. Immunofluorescence for Tyrosine Hydroxylase in the Striatum of the Rat
4.7. Samples Protein Extraction
4.8. Two-Dimensional-SDS-Page (2D-SDS-Page)
4.9. Bioinformatic Analysis
4.10. String Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tolleson, C.M.; Fang, J.Y. Advances in the mechanisms of Parkinson’s disease. Discov. Med. 2013, 15, 61–66. [Google Scholar]
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. 2014, 6, 65–74. [Google Scholar] [CrossRef]
- Dorsey, E.R.; Constantinescu, R.; Thompson, J.P.; Biglan, K.M.; Holloway, R.G.; Kieburtz, K.; Marshall, F.J.; Ravina, B.M.; Schifitto, G.; Siderowf, A.; et al. Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology 2007, 68, 384–386. [Google Scholar] [CrossRef]
- Hare, D.J.; Adlard, P.A.; Doble, P.A.; Finkelstein, D.I. Metallobiology of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine neurotoxicity. Metallomics 2013, 5, 91. [Google Scholar] [CrossRef]
- Bournival, J.; Plouffe, M.; Renaud, J.; Provencher, C.; Martinoli, M.-G. Quercetin and Sesamin Protect Dopaminergic Cells from MPP + -Induced Neuroinflammation in a Microglial (N9)-Neuronal (PC12) Coculture System. Oxidative Med. Cell. Longev. 2012, 2012, 921941. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Hwang, I.; Park, S.; Hong, S.; Hwang, B.; Cho, Y.; Son, J.; Yu, J.-W. MPTP-driven NLRP3 inflammasome activation in microglia plays a central role in dopaminergic neurodegeneration. Cell Death Differ. 2019, 26, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Z.; Cao, B.-B.; Qiu, Y.-H.; Peng, Y.-P. TGF-β1 Neuroprotection via Inhibition of Microglial Activation in a Rat Model of Parkinson’s Disease. J. Neuroimmune Pharmacol. 2017, 12, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed]
- Vitlic, A.; Lord, J.M.; Phillips, A.C. Stress ageing and their influence on functional, cellular and molecular aspects of the immune system. AGE 2014, 36, 9631. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, T.R.; Tobert, J.A. Simvastatin: A review. Expert Opin. Pharmacother. 2004, 5, 2583–2596. [Google Scholar] [CrossRef]
- Prasad, E.M.; Hung, S.-Y. Behavioral Tests in Neurotoxin-Induced Animal Models of Parkinson’s Disease. Antioxidants 2020, 9, 1007. [Google Scholar] [CrossRef]
- Selley, M.L. Simvastatin prevents 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced striatal dopamine depletion and protein tyrosine nitration in mice. Brain Res. 2005, 1037, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Vidal, Y.; Montes, S.; Tristan-López, L.; Anaya-Ramos, L.; Teiber, J.; Ríos, C.; Baron-Flores, V.; Monroy-Noyola, A. The neuroprotective effect of lovastatin on MPP + -induced neurotoxicity is not mediated by PON2. Neurotoxicology 2015, 48, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, K.; Ullah, Z.; Al Masoudi, A.S.; Ahmad, I. Simultaneous administration of fluoxetine and simvastatin ameliorates lipid profile, improves brain level of neurotransmitters, and increases bioavailability of simvastatin. J. Exp. Pharmacol. 2017, 11, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Garg, A.; Jin, B.; Keshavarz, S.S.; Bieberdorf, F.A.; Chodakewitz, J.; Wagner, J.A. Assessment of a pharmacokinetic and pharmacodynamic interaction between simvastatin and anacetrapib, a potent cholesteryl ester transfer protein (CETP) inhibitor, in healthy subjects. Br. J. Clin. Pharmacol. 2009, 67, 520–526. [Google Scholar] [CrossRef]
- Santiago, M.; Hernández-Romero, M.C.; Machado, A.; Cano, J. Zocor Forte® (simvastatin) has a neuroprotective effect against LPS striatal dopaminergic terminals injury, whereas against MPP+ does not. Eur. J. Pharmacol. 2009, 609, 58–64. [Google Scholar] [CrossRef]
- Aguirre-Vidal, Y.; Morales-Montor, J.; de León, C.T.G.; Ostoa-Saloma, P.; Díaz-Zaragoza, M.; Montes, S.; Arteaga-Silva, M.; Monroy-Noyola, A. Protection induced by estradiol benzoate in the MPP+ rat model of Parkinson’s disease is associated with the regulation of the inflammatory cytokine profile in the nigro striatum. J. Neuroimmunol. 2020, 349, 577426. [Google Scholar] [CrossRef]
- Brzozowski, M.J.; Jenner, P.; Rose, S. Inhibition of i-NOS but not n-NOS protects rat primary cell cultures against MPP(+)-induced neuronal toxicity. J. Neural Transm. 2015, 122, 779–788. [Google Scholar] [CrossRef]
- Hernández-Romero, M.D.C.; Argüelles, S.; Villarán, R.F.; de Pablos, R.M.; Delgado-Cortés, M.J.; Santiago, M.; Herrera, A.J.; Cano, J.; Machado, A. Simvastatin prevents the inflammatory process and the dopaminergic degeneration induced by the intranigral injection of lipopolysaccharide. J. Neurochem. 2008, 105, 445–459. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef]
- Yan, J.; Qiao, L.; Wu, J.; Fan, H.; Sun, J.; Zhang, Y. Simvastatin Protects Dopaminergic Neurons Against MPP+-Induced Oxidative Stress and Regulates the Endogenous Anti-Oxidant System Through ERK. Cell. Physiol. Biochem. 2018, 51, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Sun, J.; Huang, L.; Fu, Q.; Du, G. Simvastatin prevents neuroinflammation by inhibiting N-methyl-D-aspartic acid receptor 1 in 6-hydroxydopamine-treated PC12 cells. J. Neurosci. Re.s 2014, 92, 634–640. [Google Scholar] [CrossRef] [PubMed]
- ZWang, L.; Yuan, L.; Li, W.; Li, J.-Y. Ferroptosis in Parkinson’s disease: Glia–neuron crosstalk. Trends Mol. Med. 2022, 28, 258–269. [Google Scholar] [CrossRef]
- Vasefi, M.; Beazely, M.A. Neuroprotective effects of direct activation and transactivation of PDGFβ receptors. Vessel. Plus 2020, 4, 24. [Google Scholar] [CrossRef]
- Zou, B.; Liu, J.; Klionsky, D.J.; Tang, D.; Kang, R. Extracellular SQSTM1 as an inflammatory mediator. Autophagy 2020, 16, 2313–2315. [Google Scholar] [CrossRef]
- Cui, X.; Tang, J.; Hartanto, Y.; Zhang, J.; Bi, J.; Dai, S.; Qiao, S.Z.; Cheng, K.; Zhang, H. NIPAM-based Microgel Microenvironment Regulates the Therapeutic Function of Cardiac Stromal Cells. ACS Appl. Mater. Interfaces 2018, 10, 37783–37796. [Google Scholar] [CrossRef]
- Rubio-Osornio, M.; Orozco-Ibarra, M.; Díaz-Ruiz, A.; Brambila, E.; Boll, M.-C.; Monroy-Noyola, A.; Guevara, J.; Montes, S.; Ríos, C. Copper sulfate pretreatment prevents mitochondrial electron transport chain damage and apoptosis against MPP+ -induced neurotoxicity. Chem. Biol. Interact. 2017, 271, 1–8. [Google Scholar] [CrossRef]
- Rubio-Osornio, M.; Montes, S.; Pérez-Severiano, F.; Aguilera, P.; Floriano-Sánchez, E.; Monroy-Noyola, A.; Rubio, C.; Ríos, C. Copper reduces striatal protein nitration and tyrosine hydroxylase inactivation induced by MPP+ in rats. Neurochem. Int. 2009, 54, 447–451. [Google Scholar] [CrossRef]
- Rubio-Osornio, M.; Montes, S.; Heras-Romero, Y.; Guevara, J.; Rubio, C.; Aguilera, P.; Rivera-Mancia, S.; Floriano-Sánchez, E.; Monroy-Noyola, A.; Ríos, C. Induction of ferroxidase enzymatic activity by copper reduces MPP+-evoked neurotoxicity in rats. Neurosci. Res. 2013, 75, 250–255. [Google Scholar] [CrossRef]
- Triggs, W.J.; Willmore, L.J. In vivo lipid peroxidation in rat brain following intracortical Fe2+ injection. J. Neurochem. 1984, 42, 976–980. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Spots | Number of Spots That Diminishes Their Expression | Number of Spots That Increases Their Expression |
|---|---|---|---|
| Control | 224 | - | - |
| Simvastatin | 239 | 174 | 52 |
| MPP+ | 177 | 194 | 32 |
| MPP+/Simvastatin | 130 | 211 | 13 |
| SPOT | Protein Name | UniProtK B Accession Number | Theoretical Mr/pI | Experimental Mr/pI | Possible Function | Relative Expression | |||
|---|---|---|---|---|---|---|---|---|---|
| ST1 | ST2 | ST3 | ST4 | ||||||
| 3501 | Carcinoembryonic antigen-related cell adhesion molecule 1 (Ceacam1) | P16573 | 50.76/5.31 | 50.27/5.31 | Cell adhesion, Positive regulation of activation-induced cell death of T cells. | + | ++++ | ++ | ++++ |
| 5103 | TGF-β2 | Q07257 | 12.69/6.88 | 12.09/6.91 | Cytokine | + | - | - | - |
| 7403 | TGF-β1 (Tgfb1) | P17246 | 44.33/8.59 | 40.94/8.52 | Inflammatory response, Positive regulation of activation-induced cell death of T cells, Negative regulation of macrophage cytokine production. | ++ | + | - | - |
| 4603 | TGF-β3 (Tgfb3) | Q07258 | 47.12/6.10 | 48.11/6.06 | Activation of MAPK activity. | ++ | ++ | + | - |
| 6401 | Proteinase-activated receptor 1 | P26824 | 43.31/7.39 | 41.24/7.41 | Inflammatory response. | + | - | - | - |
| 4402 | Heme oxygenase 1 | P06762 | 33.00/6.08 | 33.57/6.09 | Oxidoreductase, Apoptosis. | + | - | - | - |
| 3302 | IL-1α | P16598 | 30.86/5.59 | 30.32/5.53 | Inflammatory response. | + | ++ | - | - |
| 8301 | IL-1β | Q63264 | 30.64/8.37 | 31.80/8.34 | Inflammatory response. | - | ++ | + | - |
| 3104 | Interleukin-23 subunit alpha (IL-23α) | Q91Z84 | 19.60/5.63 | 17.92/5.62 | Immunit, Inflammatory response. | + | - | - | - |
| 3004 | Protein S100-A8 | P50115 | 10.10/5.69 | 9.46/5.64 | Inflammatory response. | ++ | + | - | - |
| 1302 | Amyloid-beta A4 protein | P08592 | 30.03/4.15 | 26.92/4.13 | Apoptosis, Cell adhesion, Endocytosis, Notch signaling pathway. | + | - | - | - |
| 6206 | Allograft inflammatory factor 1 (Aif1) | P55009 | 16.69/7.83 | 18.75/7.87 | Cytoskeleton. | ++ | + | - | - |
| 5302 | B- and T-lymphocyte attenuator | Q6PNM1 | 31.11/6.77 | 30.35/6.77 | Adaptive immunity. | + | - | - | - |
| 3003 | TYRO protein tyrosine kinase-binding protein | Q6 × 9T7 | 9.54/5.69 | 8.29/5.63 | Immunity. | + | - | - | - |
| 6203 | IL-6r | P22273 | 24.36/7.77 | 22.37/7.66 | Response to lipopolysaccharide. | + | ++ | - | - |
| 5504 | Corticotropin-releasing factor receptor 2 | P47866 | 47.69/6.85 | 47.18/6.86 | G-protein coupled receptor. | + | + | ++++ | ++ |
| 2203 | Leucine repeat adapter protein 25 | Q566R4 | 18.71/4.95 | 18.97/4.95 | Negative regulation of transforming growth factor beta receptor signaling pathway. | + | ++ | - | - |
| 6503 | Serine protease HTRA1 | Q9QZK5 | 48.97/7.55 | 49.83/7.56 | Negative regulation of transforming growth factor beta receptor signaling pathway. | + | - | - | - |
| 9103 | Insulin-like growth factor I (Igf1) | P08025 | 17.83/9.77 | 17.91/9.75 | Growth factor. | ++++ | +++ | ++ | - |
| 2404 | cAMP-dependent protein kinase type I-alpha regulatory subunit | P09456 | 43.09/5.28 | 43.18/5.27 | cAMP, cAMP-binding, Nucleotide-binding. | + | ++ | ++ | - |
| 8003 | Caspase-8 | Q9JHX4 | 10.83/9.13 | 11.21/9.13 | Apoptosis. | + | - | - | - |
| 3204 | Myc box-dependent-interacting protein 1 | O08839 | 25.23/5.72 | 24.85/5.73 | Positive regulation of astrocyte. | + | - | - | - |
| 2604 | Myelin-associated glycoprotein | P07722 | 67.17/4.94 | 68.81/4.93 | Axon regeneration. | + | - | - | - |
| 4207 | Matrilysin | P50280 | 18.93/6.22 | 19.35/6.21 | Metalloprotease. | + | - | - | - |
| 8102 | WAP four-disulfide core domain protein 2 | Q8CHN3 | 12.39/8.89 | 12.50/8.88 | Serine protease inhibitor. | + | - | - | - |
| 7002 | Protein WFDC9 | Q6IE41 | 6.39/8.19 | 6.78/8.20 | Unknown. | + | - | - | - |
| 6201 | Vascular endothelial growth factor B | O35485 | 19.56/7.36 | 18.92/7.41 | Angiogenesis. | + | - | - | - |
| 2501 | Calreticulin | P18418 | 46.34/4.34 | 46.96/4.34 | Chaperone, Metal-binding, Zinc. | + | - | - | - |
| 6101 | Macrophage migration inhibitory factor | P30904 | 12.34/7.28 | 12.62/7.38 | Inflammatory response. | + | - | - | - |
| 2702 | Cadherin-17 | P55281 | 89.56/4.72 | 87.22/4.72 | Cell adhesion. | + | - | - | - |
| 3704 | Protein artemis | Q5XIX3 | 78.18/5.63 | 79.45/5.62 | DNA repair, Immunity. | + | - | - | - |
| 6205 | Interferon alpha-1 (Ifna1) | P05011 | 19.42/7.83 | 22.54/7.84 | Cytokine pro inflamamtory. | +++ | ++ | + | ++ |
| 8104 | Platelet-derived growth factor D (Pdgf-d) | Q9EQT1 | 13.96/9.39 | 13.22/9.35 | Developmental protein, Growth factor, Mitogen. | ++ | - | + | - |
| 8005 | Tumor necrosis factor ligand superfamily member 6 (Faslg) | P36940 | 8.52/9.24 | 8.88/9.24 | Proinflammatory. | ++ | + | - | - |
| 2001 | Protein S100-B | P04631 | 10.61/4.53 | 11.74/4.56 | Positive regulation of myelination. | + | - | - | - |
| 2705 | Ubiquitin carboxyl-terminal hydrolase 10 | Q3KR59 | 87.18/5.05 | 87.64/5.05 | DNA repair. | + | - | - | - |
| 2503 | Sequestosome-1 (Sqstm1) | O08623 | 47.55/5.05 | 48.01/5.07 | Apoptosis, Autophagy, Differentiation, Immunity. | +++ | +++++ | ++ | + |
| 3201 | Cdc42 effector protein 2 | Q5PQP4 | 22.91/5.31 | 23.14/5.30 | Rho protein signal transduction. | + | - | - | - |
| 5204 | Metalloproteinase inhibitor 4 | P81556 | 22.55/6.88 | 22.76/6.87 | Notch signaling pathway, central nervous system development. | + | - | - | - |
| 5201 | Caspase-6 | O35397 | 18.06/6.48 | 18.10/6.49 | Apoptosis. | ++ | + | + | - |
| 5606 | Synaptic functional regulator FMR1 | Q80WE1 | 66.78/6.77 | 68.63/6.78 | Translation regulation. | + | - | - | - |
| 2607 | Interleukin-2 receptor subunit beta | P26896 | 57.71/5.20 | 56.75/5.18 | Cytokine pro inflamamtory. | + | - | - | - |
| 6601 | Ectonucleoside triphosphate diphosphohydrolase 1 | P97687 | 57.40/7.49 | 63.42/7.45 | Regulate purinergic neurotransmission. | ++ | + | - | - |
| 2410 | Tumor necrosis factor (TNF-α) | P16599 | 25.81/5.14 | 24.27/5.19 | Cytokine pro inflammatory. | + | +++ | ++ | ++++ |
| SPOT | Protein Name | UniProtKB Accession Number | Theoretical Mr/pI | Experimental Mr/pI | Possible Function | Relative Expression | |||
|---|---|---|---|---|---|---|---|---|---|
| ST1 | ST2 | ST3 | ST4 | ||||||
| 3306 | Superoxide Dismutase 2 (SOD2) | P07895 | 15.91/5.89 | 19.55/5.88 | Vasodilation. | - | ++ | +++ | - |
| 5705 | Catalase (CAT) | P04762 | 59.63/7.15 | 58.07/7.20 | Positive regulation of NF-kappaB transcription factor activity. | - | + | - | - |
| 7203 | Glutathione Peroxidase 1 (GPX1) | P04041 | 22.35/7.70 | 21.51/7.70 | Angiogenesis. | ++ | +++ | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio-Osornio, M.; León, C.T.G.-D.; Montes, S.; Rubio, C.; Ríos, C.; Monroy, A.; Morales-Montor, J. Repurposing Simvastatin in Parkinson’s Disease Model: Protection Is throughout Modulation of the Neuro-Inflammatory Response in the Substantia nigra. Int. J. Mol. Sci. 2023, 24, 10414. https://doi.org/10.3390/ijms241310414
Rubio-Osornio M, León CTG-D, Montes S, Rubio C, Ríos C, Monroy A, Morales-Montor J. Repurposing Simvastatin in Parkinson’s Disease Model: Protection Is throughout Modulation of the Neuro-Inflammatory Response in the Substantia nigra. International Journal of Molecular Sciences. 2023; 24(13):10414. https://doi.org/10.3390/ijms241310414
Chicago/Turabian StyleRubio-Osornio, Moisés, Carmen T. Goméz-De León, Sergio Montes, Carmen Rubio, Camilo Ríos, Antonio Monroy, and Jorge Morales-Montor. 2023. "Repurposing Simvastatin in Parkinson’s Disease Model: Protection Is throughout Modulation of the Neuro-Inflammatory Response in the Substantia nigra" International Journal of Molecular Sciences 24, no. 13: 10414. https://doi.org/10.3390/ijms241310414
APA StyleRubio-Osornio, M., León, C. T. G.-D., Montes, S., Rubio, C., Ríos, C., Monroy, A., & Morales-Montor, J. (2023). Repurposing Simvastatin in Parkinson’s Disease Model: Protection Is throughout Modulation of the Neuro-Inflammatory Response in the Substantia nigra. International Journal of Molecular Sciences, 24(13), 10414. https://doi.org/10.3390/ijms241310414