
Exploring the Potential Role of Circulating microRNAs as Biomarkers for Predicting Clinical Response to Neoadjuvant Therapy in Breast Cancer
 ,
,  , and
, and
Abstract
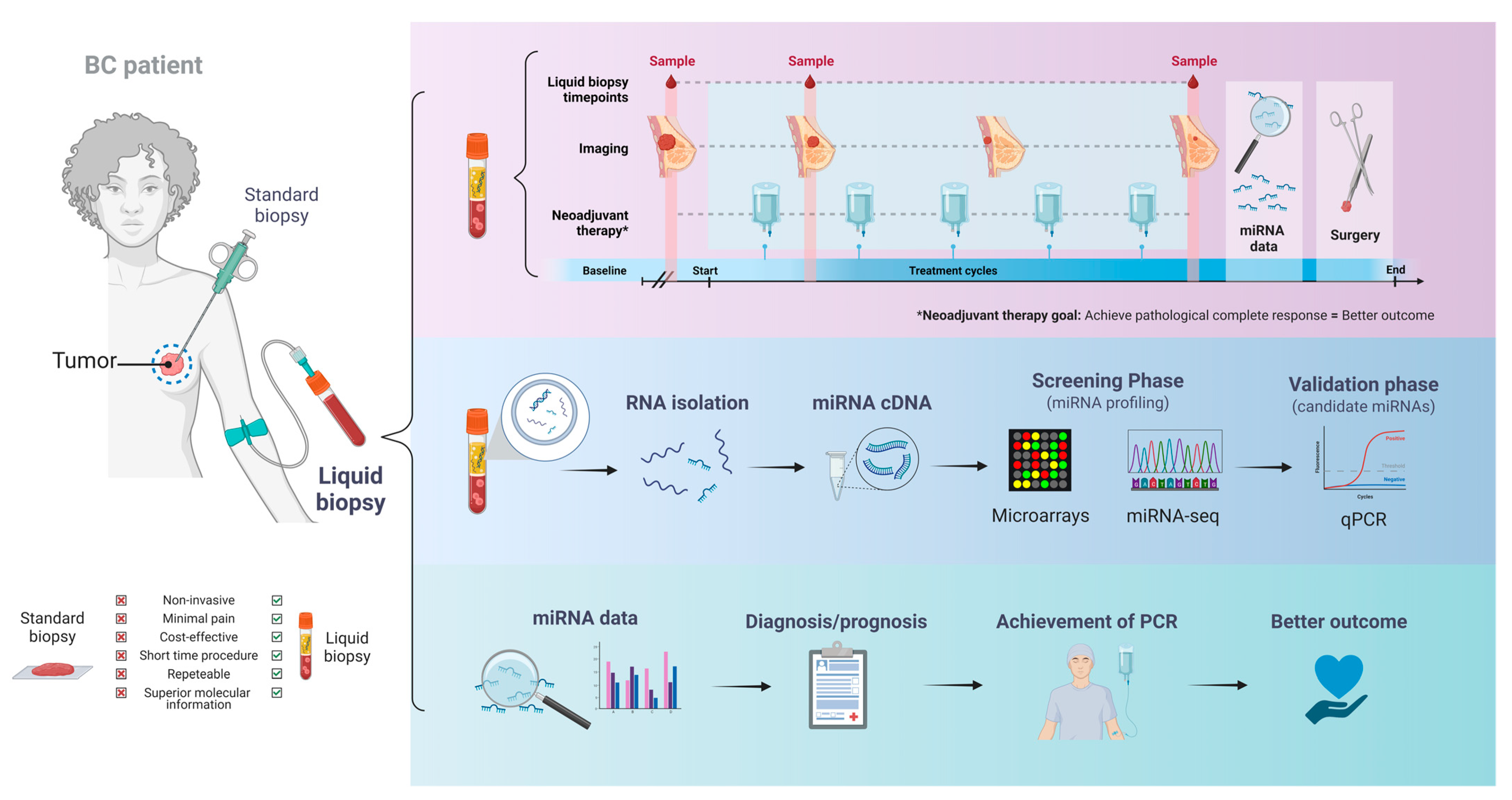
1. Introduction
2. Circulating miRNAs as Indicators of Clinical Response to NAT in BC
3. Discussion
4. Conclusions
5. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Breast cancer | BC |
| Neoadjuvant therapy | NAT |
| microRNAs | miRNAs |
| Pathologic complete response | pCR |
| Triple-negative BC | TNBC |
| 3′-untranslated region | 3′-UTR |
| High-density lipoproteins | HDL |
| Trichorhinophalangeal syndrome type 1 protein | TRPS1 |
| Epithelial-to-mesenchymal transition | EMT |
| Multidrug resistance protein 1 | MRP-1 |
| Estrogen receptor 1 | ESR1 |
| Circulating tumor cells | CTCs |
| Progesterone receptor | PGR |
| Estrogen receptor | ER |
| Event-free survival | EFS |
| BC stem cell | BCSC |
| neoadjuvant polychemotherapy | NPCT |
| Mitofusin-2 | MFN2 |
| Adriamycin | ADR |
| Clinical complete response | cCR |
| Protein inhibitor of activated signal transducer and activator of transcription 4 | PIAS4 |
| Miller–Payne | MP |
| Programmed cell death 4 | PDCD4 |
| Invasive ductal carcinoma | IDC |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Pashayan, N.; Antoniou, A.C.; Ivanus, U.; Esserman, L.J.; Easton, D.F.; French, D.; Sroczynski, G.; Hall, P.; Cuzick, J.; Evans, D.G.; et al. Personalized Early Detection and Prevention of Breast Cancer: ENVISION Consensus Statement. Nat. Rev. Clin. Oncol. 2020, 17, 687–705. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment. Am. Fam. Physician 2021, 104, 171–178. [Google Scholar] [CrossRef]
- Selli, C.; Sims, A.H. Neoadjuvant Therapy for Breast Cancer as a Model for Translational Research. Breast Cancer Basic Clin. Res. 2019, 13, 1178223419829072. [Google Scholar] [CrossRef]
- Tilsed, C.M.; Fisher, S.A.; Nowak, A.K.; Lake, R.A.; Lesterhuis, W.J. Cancer Chemotherapy: Insights into Cellular and Tumor Microenvironmental Mechanisms of Action. Front. Oncol. 2022, 12, 960317. [Google Scholar] [CrossRef] [PubMed]
- Masoud, V.; Pagès, G. Targeted Therapies in Breast Cancer: New Challenges to Fight against Resistance. World J. Clin. Oncol. 2017, 8, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.J.; Cheung, K.-L. Endocrine Therapy for Breast Cancer: A Model of Hormonal Manipulation. Oncol. Ther. 2018, 6, 141–156. [Google Scholar] [CrossRef]
- Asaoka, M.; Gandhi, S.; Ishikawa, T.; Takabe, K. Neoadjuvant Chemotherapy for Breast Cancer: Past, Present, and Future. Breast Cancer Basic Clin. Res. 2020, 14, 1178223420980377. [Google Scholar] [CrossRef]
- Korde, L.A.; Somerfield, M.R.; Carey, L.A.; Crews, J.R.; Denduluri, N.; Shelley Hwang, E.; Khan, S.A.; Loibl, S.; Morris, E.A.; Perez, A.; et al. Neoadjuvant Chemotherapy, Endocrine Therapy, and Targeted Therapy for Breast Cancer: ASCO Guideline. J. Clin. Oncol. 2021, 39, 1485–1505. [Google Scholar] [CrossRef]
- Rizzo, A.; Cusmai, A.; Massafra, R.; Bove, S.; Comes, M.; Fanizzi, A.; Rinaldi, L.; Acquafredda, S.; Gadaleta-Caldarola, G.; Oreste, D.; et al. Pathological Complete Response to Neoadjuvant Chemoimmunotherapy for Early Triple-Negative Breast Cancer: An Updated Meta-Analysis. Cells 2022, 11, 1857. [Google Scholar] [CrossRef]
- Spring, L.M.; Fell, G.; Arfe, A.; Sharma, C.; Greenup, R.; Reynolds, K.L.; Smith, B.L.; Alexander, B.; Moy, B.; Isakoff, S.J.; et al. Pathologic Complete Response after Neoadjuvant Chemotherapy and Impact on Breast Cancer Recurrence and Survival: A Comprehensive Meta-Analysis. Clin. Cancer Res. 2020, 26, 2838–2848. [Google Scholar] [CrossRef]
- Wang-Lopez, Q.; Chalabi, N.; Abrial, C.; Radosevic-Robin, N.; Durando, X.; Mouret-Reynier, M.-A.; Benmammar, K.-E.; Kullab, S.; Bahadoor, M.; Chollet, P.; et al. Can Pathologic Complete Response (PCR) Be Used as a Surrogate Marker of Survival after Neoadjuvant Therapy for Breast Cancer? Crit. Rev. Oncol. Hematol. 2015, 95, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Torrisi, R.; Marrazzo, E.; Agostinetto, E.; De Sanctis, R.; Losurdo, A.; Masci, G.; Tinterri, C.; Santoro, A. Neoadjuvant Chemotherapy in Hormone Receptor-Positive/HER2-Negative Early Breast Cancer: When, Why and What? Crit. Rev. Oncol. Hematol. 2021, 160, 103280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, H.; Li, C.; Xiang, Q.; Xu, L.; Liu, Q.; Pang, X.; Zhang, W.; Zhang, H.; Zhang, S.; et al. Circulating MicroRNAs as Indicators in the Prediction of Neoadjuvant Chemotherapy Response in Luminal B Breast Cancer. Thorac. Cancer 2021, 12, 3396–3406. [Google Scholar] [CrossRef]
- Asselain, B.; Barlow, W.; Bartlett, J.; Bergh, J.; Bergsten-Nordström, E.; Bliss, J.; Boccardo, F.; Boddington, C.; Bogaerts, J.; Bonadonna, G.; et al. Long-Term Outcomes for Neoadjuvant versus Adjuvant Chemotherapy in Early Breast Cancer: Meta-Analysis of Individual Patient Data from Ten Randomised Trials. Lancet Oncol. 2018, 19, 27–39. [Google Scholar] [CrossRef]
- Derouane, F.; van Marcke, C.; Berlière, M.; Gerday, A.; Fellah, L.; Leconte, I.; Van Bockstal, M.R.; Galant, C.; Corbet, C.; Duhoux, F.P. Predictive Biomarkers of Response to Neoadjuvant Chemotherapy in Breast Cancer: Current and Future Perspectives for Precision Medicine. Cancers 2022, 14, 3876. [Google Scholar] [CrossRef]
- de Freitas, A.J.A.; Causin, R.L.; Varuzza, M.B.; Hidalgo Filho, C.M.T.; da Silva, V.D.; Souza, C.d.P.; Marques, M.M.C. Molecular Biomarkers Predict Pathological Complete Response of Neoadjuvant Chemotherapy in Breast Cancer Patients: Review. Cancers 2021, 13, 5477. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Kajiwara, Y.; Zhu, Y.; Iha, S. Biomarkers of Neoadjuvant/Adjuvant Chemotherapy for Breast Cancer. Chin. Clin. Oncol. 2020, 9, 27. [Google Scholar] [CrossRef]
- Bownes, R.; Turnbull, A.; Cameron, D.; Sims, A.; Oikonomidou, O. On-Treatment Biomarkers Can Improve Prediction of Response to Neoadjuvant Chemotherapy in Breast Cancer. Cancer Res. 2019, 79, P3-11-13. [Google Scholar] [CrossRef]
- Asaoka, M.; Narui, K.; Suganuma, N.; Chishima, T.; Yamada, A.; Sugae, S.; Kawai, S.; Uenaka, N.; Teraoka, S.; Miyahara, K.; et al. Clinical and Pathological Predictors of Recurrence in Breast Cancer Patients Achieving Pathological Complete Response to Neoadjuvant Chemotherapy. Eur. J. Surg. Oncol. 2019, 45, 2289–2294. [Google Scholar] [CrossRef]
- Wu, H.J.; Chu, P.Y. Current and Developing Liquid Biopsy Techniques for Breast Cancer. Cancers 2022, 14, 2052. [Google Scholar] [CrossRef]
- Cirmena, G.; Dameri, M.; Ravera, F.; Fregatti, P.; Ballestrero, A.; Zoppoli, G. Assessment of Circulating Nucleic Acids in Cancer: From Current Status to Future Perspectives and Potential Clinical Applications. Cancers 2021, 13, 3460. [Google Scholar] [CrossRef] [PubMed]
- Rahat, B.; Ali, T.; Sapehia, D.; Mahajan, A.; Kaur, J. Circulating Cell-Free Nucleic Acids as Epigenetic Biomarkers in Precision Medicine. Front. Genet. 2020, 11, 844. [Google Scholar] [CrossRef] [PubMed]
- Tierno, D.; Grassi, G.; Zanconati, F.; Bortul, M.; Scaggiante, B. An Overview of Circulating Cell-Free Nucleic Acids in Diagnosis and Prognosis of Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2023, 24, 1799. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, B.; Tasso, R.; Piccioli, P.; Ciferri, M.C.; Quarto, R.; Del Mastro, L. Circulating MiRNAs in Breast Cancer Diagnosis and Prognosis. Cancers 2022, 14, 2317. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of MicroRNA Biogenesis and Its Crosstalk with Other Cellular Pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Benavides-Aguilar, J.A.; Morales-Rodríguez, J.I.; Ambriz-González, H.; Ruiz-Manriquez, L.M.; Banerjee, A.; Pathak, S.; Duttaroy, A.K.; Paul, S. The Regulatory Role of MicroRNAs in Common Eye Diseases: A Brief Review. Front. Genet. 2023, 14, 1152110. [Google Scholar] [CrossRef]
- Brümmer, A.; Hausser, J. MicroRNA Binding Sites in the Coding Region of MRNAs: Extending the Repertoire of Post-Transcriptional Gene Regulation. BioEssays 2014, 36, 617–626. [Google Scholar] [CrossRef]
- Dexheimer, P.J.; Cochella, L. MicroRNAs: From Mechanism to Organism. Front. Cell Dev. Biol. 2020, 8, 409. [Google Scholar] [CrossRef]
- Shu, J.; Silva, B.V.R.e.; Gao, T.; Xu, Z.; Cui, J. Dynamic and Modularized MicroRNA Regulation and Its Implication in Human Cancers. Sci. Rep. 2017, 7, 13356. [Google Scholar] [CrossRef]
- Ledesma-Pacheco, S.J.; Uriostegui-Pena, A.G.; Rodriguez-Jacinto, E.; Gomez-Hernandez, E.; Estrada-Meza, C.; Banerjee, A.; Pathak, S.; Ruiz-Manriquez, L.M.; Duttaroy, A.K.; Paul, S. Regulatory Mechanisms of MicroRNAs in Endocrine Disorders and Their Therapeutic Potential. Front. Genet. 2023, 14, 1137017. [Google Scholar] [CrossRef]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. Mirna-Based Biomarkers, Therapies, and Resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Bravo-Vázquez, L.A.; Frías-Reid, N.; Ramos-Delgado, A.G.; Osorio-Pérez, S.M.; Zlotnik-Chávez, H.R.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Duttaroy, A.K.; Paul, S. MicroRNAs and Long Non-Coding RNAs in Pancreatic Cancer: From Epigenetics to Potential Clinical Applications. Transl. Oncol. 2023, 27, 101579. [Google Scholar] [CrossRef]
- Ruiz-Manriquez, L.M.; Carrasco-Morales, O.; Sanchez, Z.E.A.; Osorio-Perez, S.M.; Estrada-Meza, C.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Duttaroy, A.K.; Paul, S. MicroRNA-Mediated Regulation of Key Signaling Pathways in Hepatocellular Carcinoma: A Mechanistic Insight. Front. Genet. 2022, 13, 910733. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.O.; Reiisi, S.; Shareef, S. MiRNAs, Oxidative Stress, and Cancer: A Comprehensive and Updated Review. J. Cell. Physiol. 2020, 235, 8812–8825. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Deb, M.; Kar, S.; Pradhan, N.; Parbin, S.; Kirtana, R.; Singh, S.P.; Suma, S.; Niharika; Roy, A.; et al. Dissecting MiRNA Facilitated Physiology and Function in Human Breast Cancer for Therapeutic Intervention. Semin. Cancer Biol. 2021, 72, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Lokhotiya, K.; Kumarasamy, C.; Sunil, K.; Suresh, D.; Shetty, S.; Muthukaliannan, G.K.; Baxi, S.; Mani, R.R.; Sivanandy, P.; et al. Mapping Research on MiRNAs in Cancer: A Global Data Analysis and Bibliometric Profiling Analysis. Pathophysiology 2022, 29, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. MiRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef]
- Hamam, R.; Hamam, D.; Alsaleh, K.A.; Kassem, M.; Zaher, W.; Alfayez, M.; Aldahmash, A.; Alajez, N.M. Circulating MicroRNAs in Breast Cancer: Novel Diagnostic and Prognostic Biomarkers. Cell Death Dis. 2017, 8, e3045. [Google Scholar] [CrossRef]
- El-Daly, S.M.; Gouhar, S.A.; Elmageed, Z.Y.A. Circulating MicroRNAs as Reliable Tumor Biomarkers: Opportunities and Challenges Facing Clinical Application. J. Pharmacol. Exp. Ther. 2023, 384, 35–51. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA Biogenesis Pathways in Cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef]
- Filipów, S.; Łaczmański, Ł. Blood Circulating MiRNAs as Cancer Biomarkers for Diagnosis and Surgical Treatment Response. Front. Neurosci. 2019, 13, 169. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating MicroRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin. Epigenet. 2018, 10, 59. [Google Scholar] [CrossRef]
- Tan, S.; Xia, L.; Yi, P.; Han, Y.; Tang, L.; Pan, Q.; Tian, Y.; Rao, S.; Oyang, L.; Liang, J.; et al. Exosomal MiRNAs in Tumor Microenvironment. J. Exp. Clin. Cancer Res. 2020, 39, 67. [Google Scholar] [CrossRef]
- Xu, R.; Rai, A.; Chen, M.; Suwakulsiri, W.; Greening, D.W.; Simpson, R.J. Extracellular Vesicles in Cancer—Implications for Future Improvements in Cancer Care. Nat. Rev. Clin. Oncol. 2018, 15, 617–638. [Google Scholar] [CrossRef]
- Li, I.; Nabet, B.Y. Exosomes in the Tumor Microenvironment as Mediators of Cancer Therapy Resistance. Mol. Cancer 2019, 18, 32. [Google Scholar] [CrossRef]
- Patel, H.; Nilendu, P.; Jahagirdar, D.; Pal, J.K.; Sharma, N.K. Modulating Secreted Components of Tumor Microenvironment: A Masterstroke in Tumor Therapeutics. Cancer Biol. Ther. 2018, 19, 3–12. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. MicroRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, M.; Fan, Y.; Ma, F.; Xu, N.; Xu, B. Dynamics of Circulating MicroRNAs as a Novel Indicator of Clinical Response to Neoadjuvant Chemotherapy in Breast Cancer. Cancer Med. 2018, 7, 4420–4433. [Google Scholar] [CrossRef]
- Stevic, I.; Müller, V.; Weber, K.; Fasching, P.A.; Karn, T.; Marmé, F.; Schem, C.; Stickeler, E.; Denkert, C.; Van Mackelenbergh, M.; et al. Specific MicroRNA Signatures in Exosomes of Triple-Negative and HER2-Positive Breast Cancer Patients Undergoing Neoadjuvant Therapy within the GeparSixto Trial. BMC Med. 2018, 16, 179. [Google Scholar] [CrossRef] [PubMed]
- Akkiprik, M.; Koca, S.; Uğurlu, M.; Ekren, R.; Eyüboğlu, I.P.; Alan, Ö.; Erzik, C.; Amuran, G.G.; Telli, T.A.; Güllüoğlu, M.B.; et al. Response Assessment with Molecular Characterization of Circulating Tumor Cells and Plasma MicroRNA Profiling in Patients with Locally Advanced Breast Cancer during Neoadjuvant Chemotherapy. Clin. Breast Cancer 2020, 20, 332–343.e3. [Google Scholar] [CrossRef]
- Di Cosimo, S.; Appierto, V.; Pizzamiglio, S.; Silvestri, M.; Baselga, J.; Piccart, M.; Huober, J.; Izquierdo, M.; de la Pena, L.; Hilbers, F.S.; et al. Early Modulation of Circulating MicroRNAs Levels in HER2-Positive Breast Cancer Patients Treated with Trastuzumab-Based Neoadjuvant Therapy. Int. J. Mol. Sci. 2020, 21, 1386. [Google Scholar] [CrossRef]
- Di Cosimo, S.; Appierto, V.; Pizzamiglio, S.; Tiberio, P.; Iorio, M.V.; Hilbers, F.; De Azambuja, E.; De La Peña, L.; Izquierdo, M.; Huober, J.; et al. Plasma MiRNA Levels for Predicting Therapeutic Response to Neoadjuvant Treatment in HER2-Positive Breast Cancer: Results from the NeoALTTO Trial. Clin. Cancer Res. 2019, 25, 3887–3895. [Google Scholar] [CrossRef]
- Baldasici, O.; Balacescu, L.; Cruceriu, D.; Roman, A.; Lisencu, C.; Fetica, B.; Visan, S.; Cismaru, A.; Jurj, A.; Barbu-Tudoran, L.; et al. Circulating Small EVs MiRNAs as Predictors of Pathological Response to Neo-Adjuvant Therapy in Breast Cancer Patients. Int. J. Mol. Sci. 2022, 23, 2625. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Su, F.; Lv, X.; Zhang, W.; Shang, X.; Zhang, Y.; Zhang, J. Serum MicroRNA-21 Predicted Treatment Outcome and Survival in HER2-Positive Breast Cancer Patients Receiving Neoadjuvant Chemotherapy Combined with Trastuzumab. Cancer Chemother. Pharmacol. 2019, 84, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Sukhija, S.; Purohit, P.; Pareek, P.; Garg, P.K.; Vishnoi, J.R.; Elhence, P.A.; Varthya, S.B.; Sharma, P.; Ambwani, S.; Charan, J. Circulating MiRNA-21 Levels in Breast Cancer Patients Before and After Chemotherapy and Its Association with Clinical Improvement. Indian J. Clin. Biochem. 2023. [Google Scholar] [CrossRef]
- McGuire, A.; Casey, M.C.; Waldron, R.M.; Heneghan, H.; Kalinina, O.; Holian, E.; McDermott, A.; Lowery, A.J.; Newell, J.; Dwyer, R.M.; et al. Prospective Assessment of Systemic MicroRNAs as Markers of Response to Neoadjuvant Chemotherapy in Breast Cancer. Cancers 2020, 12, 1820. [Google Scholar] [CrossRef]
- Ibrahim, A.M.; Said, M.M.; Hilal, A.M.; Medhat, A.M.; Abd Elsalam, I.M. Candidate Circulating MicroRNAs as Potential Diagnostic and Predictive Biomarkers for the Monitoring of Locally Advanced Breast Cancer Patients. Tumour. Biol. 2020, 42, 1010428320963811. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, A.; De Miguel-Pérez, D.; Ortega, F.G.; García-Puche, J.L.; Robles-Fernández, I.; Exposito, J.; Martorell-Marugan, J.; Carmona-Sáez, P.; Garrido-Navas, M.D.C.; Rolfo, C.; et al. Exosomal MiRNA Profile as Complementary Tool in the Diagnostic and Prediction of Treatment Response in Localized Breast Cancer under Neoadjuvant Chemotherapy. Breast Cancer Res. 2019, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.K.; Byrum, S.D.; Gies, A.J.; Haynie, C.; Smith, H.; Reyna, N.S.; Makhoul, I. Circulating Exosomal MicroRNAs as Predictive Biomarkers of Neoadjuvant Chemotherapy Response in Breast Cancer. Curr. Oncol. 2022, 29, 613–630. [Google Scholar] [CrossRef]
- Chekhun, V.F.; Borikun, T.V.; Bazas, V.; Andriiv, A.V.; Klyusov, O.; Yalovenko, T.; Lukianova, N.Y. Association of Circulating MiR-21, -205, and -182 with Response of Luminal Breast Cancers to Neoadjuvant FAC and AC Treatment. Exp. Oncol. 2020, 42, 162–166. [Google Scholar] [CrossRef]
- Ritter, A.; Hirschfeld, M.; Berner, K.; Rücker, G.; Jäger, M.; Weiss, D.; Medl, M.; Nöthling, C.; Gassner, S.; Asberger, J.; et al. Circulating Non-Coding RNA-Biomarker Potential in Neoadjuvant Chemotherapy of Triple Negative Breast Cancer? Int. J. Oncol. 2020, 56, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.G.; Casey, M.C.; McGuire, A.; Waldron, R.M.; Paganga, M.; Holian, E.; Newell, J.; Heneghan, H.M.; McDermott, A.M.; Keane, M.M.; et al. Evaluating the Role of Circulating MicroRNAs to Aid Therapeutic Decision Making for Neoadjuvant Chemotherapy in Breast Cancer: A Prospective, Multicenter Clinical Trial. Ann. Surg. 2022, 276, 905. [Google Scholar] [CrossRef]
- Liang, Y.K.; Lin, H.Y.; Dou, X.W.; Chen, M.; Wei, X.L.; Zhang, Y.Q.; Wu, Y.; Chen, C.F.; Bai, J.W.; Xiao, Y.S.; et al. MiR-221/222 Promote Epithelial-Mesenchymal Transition by Targeting Notch3 in Breast Cancer Cell Lines. NPJ Breast Cancer 2018, 4, 20. [Google Scholar] [CrossRef]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.-J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. TRPS1 Targeting by MiR-221/222 Promotes the Epithelial-to-Mesenchymal Transition in Breast Cancer. Sci. Signal 2011, 4, ra41. [Google Scholar] [CrossRef] [PubMed]
- Emran, T.B.; Shahriar, A.; Mahmud, A.R.; Rahman, T.; Abir, M.H.; Siddiquee, M.F.R.; Ahmed, H.; Rahman, N.; Nainu, F.; Wahyudin, E.; et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front. Oncol. 2022, 12, 891652. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, H.; Liu, X.; Evans, B.R.; Medina, D.J.; Liu, C.-G.; Yang, J.-M. Role of MicroRNA MiR-27a and MiR-451 in the Regulation of MDR1/P-Glycoprotein Expression in Human Cancer Cells. Biochem. Pharmacol. 2008, 76, 582–588. [Google Scholar] [CrossRef]
- Luengo-Gil, G.; Gonzalez-Billalabeitia, E.; Perez-Henarejos, S.A.; Navarro Manzano, E.; Chaves-Benito, A.; Garcia-Martinez, E.; Garcia-Garre, E.; Vicente, V.; De La Peña, F.A. Angiogenic Role of MiR-20a in Breast Cancer. PLoS ONE 2018, 13, e0194638. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Chadalapaka, G.; Lee, S.-O.; Yamada, D.; Sastre-Garau, X.; Defossez, P.-A.; Park, Y.-Y.; Lee, J.-S.; Safe, S. Identification of Oncogenic MicroRNA-17-92/ZBTB4/Specificity Protein Axis in Breast Cancer. Oncogene 2012, 31, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Imani, S.; Wu, R.C.; Fu, J. MicroRNA-34 Family in Breast Cancer: From Research to Therapeutic Potential. J. Cancer 2018, 9, 3765–3775. [Google Scholar] [CrossRef]
- von Minckwitz, G.; Loibl, S.; Untch, M.; Eidtmann, H.; Rezai, M.; Fasching, P.A.; Tesch, H.; Eggemann, H.; Schrader, I.; Kittel, K.; et al. Survival after Neoadjuvant Chemotherapy with or without Bevacizumab or Everolimus for HER2-Negative Primary Breast Cancer (GBG 44-GeparQuinto). Ann. Oncol. 2014, 25, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- von Minckwitz, G.; Schneeweiss, A.; Loibl, S.; Salat, C.; Denkert, C.; Rezai, M.; Blohmer, J.U.; Jackisch, C.; Paepke, S.; Gerber, B.; et al. Neoadjuvant Carboplatin in Patients with Triple-Negative and HER2-Positive Early Breast Cancer (GeparSixto; GBG 66): A Randomised Phase 2 Trial. Lancet Oncol. 2014, 15, 747–756. [Google Scholar] [CrossRef] [PubMed]
- He, X.-H.; Zhu, W.; Yuan, P.; Jiang, S.; Li, D.; Zhang, H.-W.; Liu, M.-F. MiR-155 Downregulates ErbB2 and Suppresses ErbB2-Induced Malignant Transformation of Breast Epithelial Cells. Oncogene 2016, 35, 6015–6025. [Google Scholar] [CrossRef]
- Lettlova, S.; Brynychova, V.; Blecha, J.; Vrana, D.; Vondrusova, M.; Soucek, P.; Truksa, J. MiR-301a-3p Suppresses Estrogen Signaling by Directly Inhibiting ESR1 in ERα Positive Breast Cancer. Cell. Physiol. Biochem. 2018, 46, 2601–2615. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhang, J.; Zhong, L.; Wang, L.; Liu, Y.; Wang, Y.; Peng, L.; Guo, B. Upregulated MicroRNA-301a in Breast Cancer Promotes Tumor Metastasis by Targeting PTEN and Activating Wnt/β-Catenin Signaling. Gene 2014, 535, 191–197. [Google Scholar] [CrossRef]
- Zhang, G.J.; Xiao, H.X.; Tian, H.P.; Liu, Z.L.; Xia, S.; Zhou, T. Upregulation of MicroRNA-155 Promotes the Migration and Invasion of Colorectal Cancer Cells through the Regulation of Claudin-1 Expression. Int. J. Mol. Med. 2013, 31, 1375–1380. [Google Scholar] [CrossRef]
- Ali, S.A.; Abdulrahman, Z.F.A.; Faraidun, H.N. Circulatory MiRNA-155, MiRNA-21 Target PTEN Expression and Activity as a Factor in Breast Cancer Development. Cell. Mol. Biol. 2020, 66, 44–50. [Google Scholar] [CrossRef]
- Jabbari, N.; Akbariazar, E.; Feqhhi, M.; Rahbarghazi, R.; Rezaie, J. Breast Cancer-Derived Exosomes: Tumor Progression and Therapeutic Agents. J. Cell. Physiol. 2020, 235, 6345–6356. [Google Scholar] [CrossRef]
- Qattan, A. Novel Mirna Targets and Therapies in the Triple-Negative Breast Cancer Microenvironment: An Emerging Hope for a Challenging Disease. Int. J. Mol. Sci. 2020, 21, 8905. [Google Scholar] [CrossRef]
- Zhao, F.; Zhong, M.; Pei, W.; Tian, B.; Cai, Y. MiR-376c-3p Modulates the Properties of Breast Cancer Stem Cells by Targeting RAB2A. Exp. Ther. Med. 2020, 20, 68. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; He, S. Hsa_circ_0025202 Suppresses Cell Tumorigenesis and Tamoxifen Resistance via MiR-197-3p/HIPK3 Axis in Breast Cancer. World J. Surg. Oncol. 2021, 19, 39. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Yang, R.; Zhao, S.; Chen, Y.; Hong, S.; Wang, K.; Wang, T.; Cheng, J.; Zhang, T.; Chen, D. Decreased MiR-320 Expression Is Associated with Breast Cancer Progression, Cell Migration, and Invasiveness via Targeting Aquaporin 1. Acta Biochim. Biophys. Sin. 2018, 50, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; Li, J.; Geng, H.; Zhou, B.; Zhang, B.; Chen, H. MiR-320/ELF3 Axis Inhibits the Progression of Breast Cancer via the PI3K/AKT Pathway. Oncol. Lett. 2020, 19, 3239–3248. [Google Scholar] [CrossRef] [PubMed]
- Fuso, P.; Di Salvatore, M.; Santonocito, C.; Guarino, D.; Autilio, C.; Mulè, A.; Arciuolo, D.; Rinninella, A.; Mignone, F.; Ramundo, M.; et al. Let-7a-5p, Mir-100-5p, Mir-101-3p, and Mir-199a-3p Hyperexpression as Potential Predictive Biomarkers in Early Breast Cancer Patients. J. Pers. Med. 2021, 11, 816. [Google Scholar] [CrossRef]
- An, C.; Hu, Z.; Li, Y.; Zhao, P.; Liu, R.; Zhang, Q.; Zhu, P.; Li, Y.; Wang, Y. LINC00662 Enhances Cell Progression and Stemness in Breast Cancer by MiR-144-3p/SOX2 Axis. Cancer Cell Int. 2022, 22, 184. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, X.; Tan, W.; Gao, J.; Pan, L.; Ye, X.; Chen, L.; Zheng, W. MiR-145-5p Suppresses Breast Cancer Progression by Inhibiting SOX2. J. Surg. Res. 2019, 236, 278–287. [Google Scholar] [CrossRef]
- Assiri, A.A.; Mourad, N.; Shao, M.; Kiel, P.; Liu, W.; Skaar, T.C.; Overholser, B.R. MicroRNA 362-3p Reduces HERG-Related Current and Inhibits Breast Cancer Cells Proliferation. Cancer Genom. Proteom. 2019, 16, 433–442. [Google Scholar] [CrossRef]
- Son, D.; Kim, Y.; Lim, S.; Kang, H.-G.; Kim, D.-H.; Park, J.W.; Cheong, W.; Kong, H.K.; Han, W.; Park, W.-Y.; et al. MiR-374a-5p Promotes Tumor Progression by Targeting ARRB1 in Triple Negative Breast Cancer. Cancer Lett. 2019, 454, 224–233. [Google Scholar] [CrossRef]
- Zha, Z.; Jia, F.; Hu, P.; Mai, E.; Lei, T. MicroRNA-574-3p Inhibits the Malignant Behavior of Liver Cancer Cells by Targeting ADAM28. Oncol. Lett. 2020, 20, 3015–3023. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, J.; Lu, Y.; Bo, S.; Li, L.; Wang, L.; Zhang, Q.; Mao, J. MiR-140-5p Inhibits the Proliferation and Enhances the Efficacy of Doxorubicin to Breast Cancer Stem Cells by Targeting Wnt1. Cancer Gene Ther. 2019, 26, 74–82. [Google Scholar] [CrossRef]
- Deng, S.; Wang, M.; Wang, C.; Zeng, Y.; Qin, X.; Tan, Y.; Liang, B.; Cao, Y. P53 Downregulates PD-L1 Expression via MiR-34a to Inhibit the Growth of Triple-Negative Breast Cancer Cells: A Potential Clinical Immunotherapeutic Target. Mol. Biol. Rep. 2023, 50, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Luo, X.; Ma, L.; Wang, Y.; Zhang, F. Identifying of MiR-98-5p/IGF1 Axis Contributes Breast Cancer Progression Using Comprehensive Bioinformatic Analyses Methods and Experiments Validation. Life Sci. 2020, 261, 118435. [Google Scholar] [CrossRef]
- Ruiz-Manriquez, L.M.; Estrada-Meza, C.; Benavides-Aguilar, J.A.; Ledesma-Pacheco, S.J.; Torres-Copado, A.; Serrano-Cano, F.I.; Bandyopadhyay, A.; Pathak, S.; Chakraborty, S.; Srivastava, A.; et al. Phytochemicals Mediated Modulation of MicroRNAs and Long Non-Coding RNAs in Cancer Prevention and Therapy. Phyther. Res. 2022, 36, 705–729. [Google Scholar] [CrossRef] [PubMed]
- Park, H.A.; Brown, S.R.; Kim, Y. Cellular Mechanisms of Circulating Tumor Cells during Breast Cancer Metastasis. Int. J. Mol. Sci. 2020, 21, 5040. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalaf, H.H.; Aboussekhra, A. P16INK4A Induces Senescence and Inhibits EMT through MicroRNA-141/MicroRNA-146b-5p-Dependent Repression of AUF1. Mol. Carcinog. 2017, 56, 985–999. [Google Scholar] [CrossRef]
- Chen, J.; Shin, V.Y.; Siu, M.T.; Ho, J.C.W.; Cheuk, I.; Kwong, A. MiR-199a-5p Confers Tumor-Suppressive Role in Triple-Negative Breast Cancer. BMC Cancer 2016, 16, 887. [Google Scholar] [CrossRef]
- Amirfallah, A.; Knutsdottir, H.; Arason, A.; Hilmarsdottir, B.; Johannsson, O.T.; Agnarsson, B.A.; Barkardottir, R.B.; Reynisdottir, I. Hsa-MiR-21-3p Associates with Breast Cancer Patient Survival and Targets Genes in Tumor Suppressive Pathways. PLoS ONE 2021, 16, e0260327. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, D.; Ren, H.; Shi, Y.; Gao, Y. MiR-145-Targeted HBXIP Modulates Human Breast Cancer Cell Proliferation. Thorac. Cancer 2019, 10, 71–77. [Google Scholar] [CrossRef]
- Wang, J.; Yan, Y.; Zhang, Z.; Li, Y. Role of MiR-10b-5p in the Prognosis of Breast Cancer. PeerJ 2019, 2019, e7728. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Mu, T.; Zhao, L.; Li, Y.; Zhu, D.; Pan, Y. MiR-181a-5p Facilitates Proliferation, Invasion, and Glycolysis of Breast Cancer through NDRG2-Mediated Activation of PTEN/AKT Pathway. Bioengineered 2022, 13, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. MicroRNA-21 Promotes Breast Cancer Proliferation and Metastasis by Targeting LZTFL1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef]
- Yu, J.; Shen, W.; Gao, B.; Zhao, H.; Xu, J.; Gong, B. MicroRNA-182 Targets FOXF2 to Promote the Development of Triple-Negative Breast Cancer. Neoplasma 2017, 64, 209–215. [Google Scholar] [CrossRef]
- Huang, R.; Li, J.; Pan, F.; Zhang, B.; Yao, Y. The Activation of GPER Inhibits Cells Proliferation, Invasion and EMT of Triple-Negative Breast Cancer via CD151/MiR-199a-3p Bio-Axis. Am. J. Transl. Res. 2020, 12, 32–44. [Google Scholar] [PubMed]
- Shen, Y.; Xu, Y.; Huang, L.; Chi, Y.; Meng, L. MiR-205 Suppressed the Malignant Behaviors of Breast Cancer Cells by Targeting CLDN11 via Modulation of the Epithelial-to-Mesenchymal Transition. Aging 2021, 13, 13073–13085. [Google Scholar] [CrossRef]
- Fu, H.; Fu, L.; Xie, C.; Zuo, W.S.; Liu, Y.S.; Zheng, M.Z.; Yu, J.M. CancermiR-375 Inhibits Cancer Stem Cell Phenotype and Tamoxifen Resistance by Degrading HOXB3 in Human ER-Positive Breast Cancer. Oncol. Rep. 2017, 37, 1093–1099. [Google Scholar] [CrossRef]
- Purohit, P.K.; Edwards, R.; Tokatlidis, K.; Saini, N. MiR-195 Regulates Mitochondrial Function by Targeting Mitofusin-2 in Breast Cancer Cells. RNA Biol. 2019, 16, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Y.; Jasper, J.; Lykken, E.; Alexander, P.B.; Markowitz, G.J.; McDonnell, D.P.; Li, Q.J.; Wang, X.F. MiR-148a Functions to Suppress Metastasis and Serves as a Prognostic Indicator in Triple-Negative Breast Cancer. Oncotarget 2016, 7, 20381–20394. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, L.-Z.; Yin, Y.; He, J.; Li, Q.; Qian, X.; You, Y.; Lu, Z.; Peiper, S.C.; Shu, Y.; et al. Regulatory Circuit of PKM2/NF-ΚB/MiR-148a/152-Modulated Tumor Angiogenesis and Cancer Progression. Oncogene 2015, 34, 5482–5493. [Google Scholar] [CrossRef] [PubMed]
- Furrer, D.; Paquet, C.; Jacob, S.; Diorio, C. The Human Epidermal Growth Factor Receptor 2 (HER2) as a Prognostic and Predictive Biomarker: Molecular Insights into HER2 Activation and Diagnostic Implications. In Cancer Prognosis; Lemamy, G.-J., Ed.; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar]
- Zhang, S.; Wang, Y.; Wang, Y.; Peng, J.; Yuan, C.; Zhou, L.; Xu, S.; Lin, Y.; Du, Y.; Yang, F.; et al. Serum MiR-222-3p as a Double-Edged Sword in Predicting Efficacy and Trastuzumab-Induced Cardiotoxicity for HER2-Positive Breast Cancer Patients Receiving Neoadjuvant Target Therapy. Front. Oncol. 2020, 10, 631. [Google Scholar] [CrossRef]
- Shen, H.; Wang, D.; Li, L.; Yang, S.; Chen, X.; Zhou, S.; Zhong, S.; Zhao, J.; Tang, J. MiR-222 Promotes Drug-Resistance of Breast Cancer Cells to Adriamycin via Modulation of PTEN/Akt/FOXO1 Pathway. Gene 2017, 596, 110–118. [Google Scholar] [CrossRef]
- Zhong, W.; Chen, S.; Qin, Y.; Zhang, H.; Wang, H.; Meng, J.; Huai, L.; Zhang, Q.; Yin, T.; Lei, Y.; et al. Doxycycline Inhibits Breast Cancer EMT and Metastasis through PAR-1/NF-ΚB/MiR-17/E-Cadherin Pathway. Oncotarget 2017, 8, 104855–104866. [Google Scholar] [CrossRef]
- Zhao, L.; Zhao, Y.; He, Y.; Mao, Y. MiR-19b Promotes Breast Cancer Metastasis through Targeting MYLIP and Its Related Cell Adhesion Molecules. Oncotarget 2017, 8, 64330–64343. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wang, Y.; Wu, Y.; Zhang, X.; Zhang, X.; Liu, J.; Wang, T.; Fan, J.; Sun, J.; Yang, A.; et al. EZH2-Mediated Epigenetic Silencing of MiR-29/MiR-30 Targets LOXL4 and Contributes to Tumorigenesis, Metastasis, and Immune Microenvironment Remodeling in Breast Cancer. Theranostics 2020, 10, 8494–8512. [Google Scholar] [CrossRef]
- Xing, Z.; Wang, X.; Liu, J.; Liu, G.; Zhang, M.; Feng, K.; Wang, X. Effect of MiR-210 on the Chemosensitivity of Breast Cancer by Regulating JAK-STAT Signaling Pathway. Biomed Res. Int. 2021, 2021, 7703159. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Yang, J.; Han, Y.; Lu, Q.; Cao, J.; Syed, L. High Expression of MiR-210 Predicts Poor Survival in Patients with Breast Cancer: A Meta-Analysis. Gene 2012, 507, 135–138. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Jung, E.J.; Kim, J.-M.; Son, Y.; Lee, H.S.; Kwag, S.-J.; Park, J.-H.; Cho, J.-K.; Kim, H.-G.; Park, T.; et al. MiR-221 and MiR-222 Regulate Cell Cycle Progression and Affect Chemosensitivity in Breast Cancer by Targeting ANXA3. Exp. Ther. Med. 2023, 25, 11826. [Google Scholar] [CrossRef]
- Moorthy, R.K.; Srinivasan, C.; Kannan, M.; Arockiam, A.J.V. Deregulation of MiR-375 Inhibits HOXA5 and Promotes Migration, Invasion, and Cell Proliferation in Breast Cancer. Appl. Biochem. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Cui, F.; Zhou, Q.; Xiao, K.; Ma, S. The MicroRNA Hsa-Let-7g Promotes Proliferation and Inhibits Apoptosis in Lung Cancer by Targeting HOXB1. Yonsei Med. J. 2020, 61, 210–217. [Google Scholar] [CrossRef]
- Najjary, S.; Mohammadzadeh, R.; Mokhtarzadeh, A.; Mohammadi, A.; Kojabad, A.B.; Baradaran, B. Role of MiR-21 as an Authentic Oncogene in Mediating Drug Resistance in Breast Cancer. Gene 2020, 738, 144453. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, X.; Li, T.; Ren, Q.; Li, L.; Sun, X.; Zhang, B.; Wang, X.; Han, H.; He, Y.; et al. MicroRNA-221 Promotes Breast Cancer Resistance to Adriamycin via Modulation of PTEN/Akt/MTOR Signaling. Cancer Med. 2020, 9, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Si, C.; Yu, Q.; Yao, Y. Effect of MiR-146a-5p on Proliferation and Metastasis of Triple-Negative Breast Cancer via Regulation of SOX5. Exp. Ther. Med. 2018, 15, 4515–4521. [Google Scholar] [CrossRef] [PubMed]
- Tormo, E.; Adam-Artigues, A.; Ballester, S.; Pineda, B.; Zazo, S.; González-Alonso, P.; Albanell, J.; Rovira, A.; Rojo, F.; Lluch, A.; et al. The Role of MiR-26a and MiR-30b in HER2+ Breast Cancer Trastuzumab Resistance and Regulation of the CCNE2 Gene. Sci. Rep. 2017, 7, srep41309. [Google Scholar] [CrossRef]
- Wu, T.; Song, H.; Xie, D.; Hua, K.; Hu, J.; Deng, Y.; Ji, C.; Fang, L. Mir-30b-5p Promotes Proliferation, Migration, and Invasion of Breast Cancer Cells via Targeting ASPP2. Biomed Res. Int. 2020, 2020, 7907269. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ye, P.; Ye, Y.; Lu, S.; Han, B. Circular RNA Hsa_circRNA_002178 Silencing Retards Breast Cancer Progression via MicroRNA-328-3p-Mediated Inhibition of COL1A1. J. Cell. Mol. Med. 2020, 24, 2189–2201. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, B.; Li, J.K.; Li, S.Y.; Niu, R.L.; Fu, N.Q.; Zheng, J.J.; Liu, G.; Wang, Z.L. Collagen Fiber Features and COL1A1: Are They Associated with Elastic Parameters in Breast Lesions, and Can COL1A1 Predict Axillary Lymph Node Metastasis? BMC Cancer 2022, 22, 1004. [Google Scholar] [CrossRef]
- Dai, T.; Zhao, X.; Li, Y.; Yu, L.; Li, Y.; Zhou, X.; Gong, Q. Mir-423 Promotes Breast Cancer Invasion by Activating Nf-Κb Signaling. Onco. Targets. Ther. 2020, 13, 5467–5478. [Google Scholar] [CrossRef]
- Umeh-Garcia, M.; Simion, C.; Ho, P.Y.; Batra, N.; Berg, A.L.; Carraway, K.L.; Yu, A.; Sweeney, C. A Novel Bioengineered MiR-127 Prodrug Suppresses the Growth and Metastatic Potential of Triple-Negative Breast Cancer Cells. Cancer Res. 2020, 80, 418–429. [Google Scholar] [CrossRef]
- Wen, X.; Dai, Y.; Wu, S.; Li, J. MiR-127-3p Inhibits Breast Cancer Cell Behaviors via Targeting Benzodiazepine Receptor-Associated Protein 1 (BZRAP1). J. Biomater. Tissue Eng. 2022, 12, 2038–2043. [Google Scholar] [CrossRef]
- Song, W.; Wu, S.; Wu, Q.; Zhou, L.; Yu, L.; Zhu, B.; Gong, X. The MicroRNA-141-3p/ CDK8 Pathway Regulates the Chemosensitivity of Breast Cancer Cells to Trastuzumab. J. Cell. Biochem. 2019, 120, 14095–14106. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Duan, Z.; Zhang, C.; Wang, W.; He, H.; Liu, Y.; Wu, P.; Wang, S.; Song, M.; Chen, H.; et al. Mouse 4T1 Breast Cancer Cell–Derived Exosomes Induce Proinflammatory Cytokine Production in Macrophages via MiR-183. J. Immunol. 2020, 205, 2916–2925. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Z.; Fang, M.; Qiao, Y. MicroRNA Let-7a Inhibits Proliferation of Breast Cancer Cell by Downregulating USP32 Expression. Transl. Cancer Res. 2019, 8, 1763–1771. [Google Scholar] [CrossRef]
- Jordan-Alejandre, E.; Campos-Parra, A.D.; Castro-López, D.L.; Silva-Cázares, M.B. Potential MiRNA Use as a Biomarker: From Breast Cancer Diagnosis to Metastasis. Cells 2023, 12, 525. [Google Scholar] [CrossRef]
- Grimaldi, A.M.; Salvatore, M.; Incoronato, M. MiRNA-Based Therapeutics in Breast Cancer: A Systematic Review. Front. Oncol. 2021, 11, 668464. [Google Scholar] [CrossRef]
- Saiyed, A.N.; Vasavada, A.R.; Johar, S.R.K. Recent Trends in MiRNA Therapeutics and the Application of Plant MiRNA for Prevention and Treatment of Human Diseases. Futur. J. Pharm. Sci. 2022, 8, 24. [Google Scholar] [CrossRef]
- Bayraktar, E.; Bayraktar, R.; Oztatlici, H.; Lopez-Berestein, G.; Amero, P.; Rodriguez-Aguayo, C. Targeting MiRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update. Non-Coding RNA 2023, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G. MiRNAs: The Road from Bench to Bedside. Genes 2023, 14, 314. [Google Scholar] [CrossRef]
- Diener, C.; Keller, A.; Meese, E. Emerging Concepts of MiRNA Therapeutics: From Cells to Clinic. Trends Genet. 2022, 38, 613–626. [Google Scholar] [CrossRef]
- Samad, A.F.A.; Kamaroddin, M.F. Innovative Approaches in Transforming MicroRNAs into Therapeutic Tools. WIREs RNA 2023, 14, e1768. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Associated miRNAs | Target mRNA | Altered Biological Mechanism | Source | Reference |
|---|---|---|---|---|
| miR-222 ↑ | TRPS1 Notch3 | EMT | Plasma | [52] |
| miR-20a ↑ | ZBTB4 | Cell growth and invasion | ||
| miR-451 ↓ | MRP-1 | Anthracycline resistance | ||
| miR-34a ↓ | Bcl-2 SIRT1 | Apoptosis | ||
| miR-155 ↓ | ErbB2 HDAC2 | Cell survival, growth, and chemosensitivity | Plasma | [53] |
| miR-301 ↓ | ESR1 | Cell growth and invasion | ||
| miR-146b-5p ↓ | ZEB1 | EMT | Plasma | [54] |
| miR-199a-5p ↑ | CDH1, ZEB1, TWIST | |||
| miR-148a-3p ↑ | PKM2 | Malignant phenotype maintenance | Plasma | [55] |
| miR-374a-5p ↑ | ARRB1 | Cell survival, proliferation, and migration | [55,56] | |
| miR-21-5p ↓ | PTEN PDCD4 | Chemoresistance | Plasma | [57] |
| miR-221-3p ↓ | PTEN | |||
| miR-146a-5p ↓ | SOX5 | EMT | ||
| miR-26a-5p ↓ | CCNE2 | Chemoresistance | ||
| miR-21 ↓ | PTEN | Apoptosis and cell proliferation | Serum | [58] |
| miR-21 ↑ | LZTFL1 | Cell proliferation, invasion, and migration | Whole blood | [59] |
| miR-195 ↓ | MFN2 | Mitochondrial metabolism | [60] | |
| miR-210 ↑ | PIAS4 | Apoptosis, anthracycline resistance | Serum | [14] |
| miR-222 ↓ | ANXA3 | Cell proliferation, invasion, and migration | ||
| miR-375 ↑ | HOXA5 | Apoptosis, cell proliferation, invasion, and migration | ||
| let-7g ↓ | FOXC2 | Cell migration | ||
| miR-21-3p ↑ | MAT2 STARD13 ZNF132 | Cell proliferation, invasion, and migration | Serum | [61] |
| miR-145-5p ↑ | SOX2 | Cell proliferation | ||
| miR-145 ↑ | HBXIP | |||
| miR-10b-5 ↑ | BIRC5 E2F2 KIF2C FOXM1 MCM5 | |||
| miR-181a ↑ | SOCS3 PIAS3 ATM | Cell proliferation, invasion, and migration | Whole blood | [62] |
| miR-181a-5p ↑ | NDRG2 | |||
| miR-105 ↑ | ZO-1 | |||
| miR-221 ↑ | ERα | |||
| miR-222 ↑ | ERα | |||
| miR-155 ↑ | PTEN | |||
| miR-30b-5p ↑ | ASPP2 | Apoptosis and EMT | Plasma | [63] |
| miR-328-3p ↑ | COL1A1 | Inflammation | ||
| miR-423-5p ↑ | TNIP2 | Cell invasion | ||
| miR-127-3p ↑ | CERK NANOS1 FOXO6 SOX11 SOX12 FASN SUSD2 BZRAP1 | Cell proliferation, invasion, and migration | ||
| miR-141-3p ↑ | CDK8 | Chemoresistance | ||
| miR-34a-5p ↑ | B7-H1 | Apoptosis, cell proliferation, invasion, and migration | ||
| miR-183-5p ↑ | PPP2CA | Cell proliferation, invasion, and migration | ||
| miR-182-5p ↑ | CMTM7 | Cell proliferation, invasion, and migration | ||
| miR-376c-3p ↓ | RAB2A | Malignant phenotype maintenance Cell stemness | Plasma | [56] |
| miR-197-3p ↓ | HIPK3 | Apoptosis, cell proliferation, invasion, and migration | ||
| miR-874-3p ↑ | SOX2 | Tumor growth | ||
| miR-320c ↑ | AQP1 | Cell proliferation | ||
| miR-100-5p ↑ | PlK1 | Cell proliferation, invasion, and migration | ||
| miR-144-3p ↑ | SOX2 | Tumor growth Cell stemness | ||
| miR-362-3p ↑ | hERG | Apoptosis, cell proliferation, invasion, and migration | ||
| miR-374a-5p ↑ | ARRB1 | Tumor progression | ||
| miR-574-3p ↑ | ADAM28 | Tumor progression | ||
| miR-140-5p ↑ | Wnt1 | Cell proliferation, invasion, and migration | ||
| miR-145-5p ↑ | SOX2 | Tumor growth Cell stemness | ||
| miR-328-3p ↓ | Ki-67 | Cell proliferation | ||
| miR-34a-5p ↓ | PD-L1 P53 | Apoptosis, cell proliferation, invasion, and migration | ||
| miR-98-5p ↑ | IGF1 | Cell proliferation, invasion, and migration | ||
| miR-182 ↑ | FOXF2 | Cell proliferation and migration | Serum | [64] |
| miR-375 ↓ | HOXB3 | Malignant phenotype maintenance Cell stemness Chemoresistance | ||
| miR-205 ↑ | CLDN11 | Apoptosis and cell proliferation | ||
| miR-21 ↓ | LZTFL1 | EMT | ||
| miR-199a ↓ | CD151 | Cell proliferation, invasion, and migration Angiogenesis | ||
| miR-155 ↑ | SOCS1 MMP16 | Cell proliferation and migration | ||
| miR-17 ↓ | E-cadherin | EMT | Serum | [65] |
| miR-19b ↓ | MYLIP E-cadherin ICAM-1 Inregrin β1 | Cell adhesion | ||
| miR-30 ↓ | LOXL4 | Cell proliferation, invasion, and migration | ||
| let-7a ↑ | USP32 | Cell proliferation | Whole blood | [66] |
| miR-21 ↑ | LZTFL1 | Cell proliferation, invasion, and migration | ||
| miR-145 ↓ | SOX2 | Tumor growth Cell stemness |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Manriquez, L.M.; Villarreal-Garza, C.; Benavides-Aguilar, J.A.; Torres-Copado, A.; Isidoro-Sánchez, J.; Estrada-Meza, C.; Arvizu-Espinosa, M.G.; Paul, S.; Cuevas-Diaz Duran, R. Exploring the Potential Role of Circulating microRNAs as Biomarkers for Predicting Clinical Response to Neoadjuvant Therapy in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 9984. https://doi.org/10.3390/ijms24129984
Ruiz-Manriquez LM, Villarreal-Garza C, Benavides-Aguilar JA, Torres-Copado A, Isidoro-Sánchez J, Estrada-Meza C, Arvizu-Espinosa MG, Paul S, Cuevas-Diaz Duran R. Exploring the Potential Role of Circulating microRNAs as Biomarkers for Predicting Clinical Response to Neoadjuvant Therapy in Breast Cancer. International Journal of Molecular Sciences. 2023; 24(12):9984. https://doi.org/10.3390/ijms24129984
Chicago/Turabian StyleRuiz-Manriquez, Luis M., Cynthia Villarreal-Garza, Javier A. Benavides-Aguilar, Andrea Torres-Copado, José Isidoro-Sánchez, Carolina Estrada-Meza, María Goretti Arvizu-Espinosa, Sujay Paul, and Raquel Cuevas-Diaz Duran. 2023. "Exploring the Potential Role of Circulating microRNAs as Biomarkers for Predicting Clinical Response to Neoadjuvant Therapy in Breast Cancer" International Journal of Molecular Sciences 24, no. 12: 9984. https://doi.org/10.3390/ijms24129984
APA StyleRuiz-Manriquez, L. M., Villarreal-Garza, C., Benavides-Aguilar, J. A., Torres-Copado, A., Isidoro-Sánchez, J., Estrada-Meza, C., Arvizu-Espinosa, M. G., Paul, S., & Cuevas-Diaz Duran, R. (2023). Exploring the Potential Role of Circulating microRNAs as Biomarkers for Predicting Clinical Response to Neoadjuvant Therapy in Breast Cancer. International Journal of Molecular Sciences, 24(12), 9984. https://doi.org/10.3390/ijms24129984