
Isolation and Characterization of scFv Antibody against Internal Ribosomal Entry Site of Enterovirus A71

 ,
,
Abstract
1. Introduction
2. Results
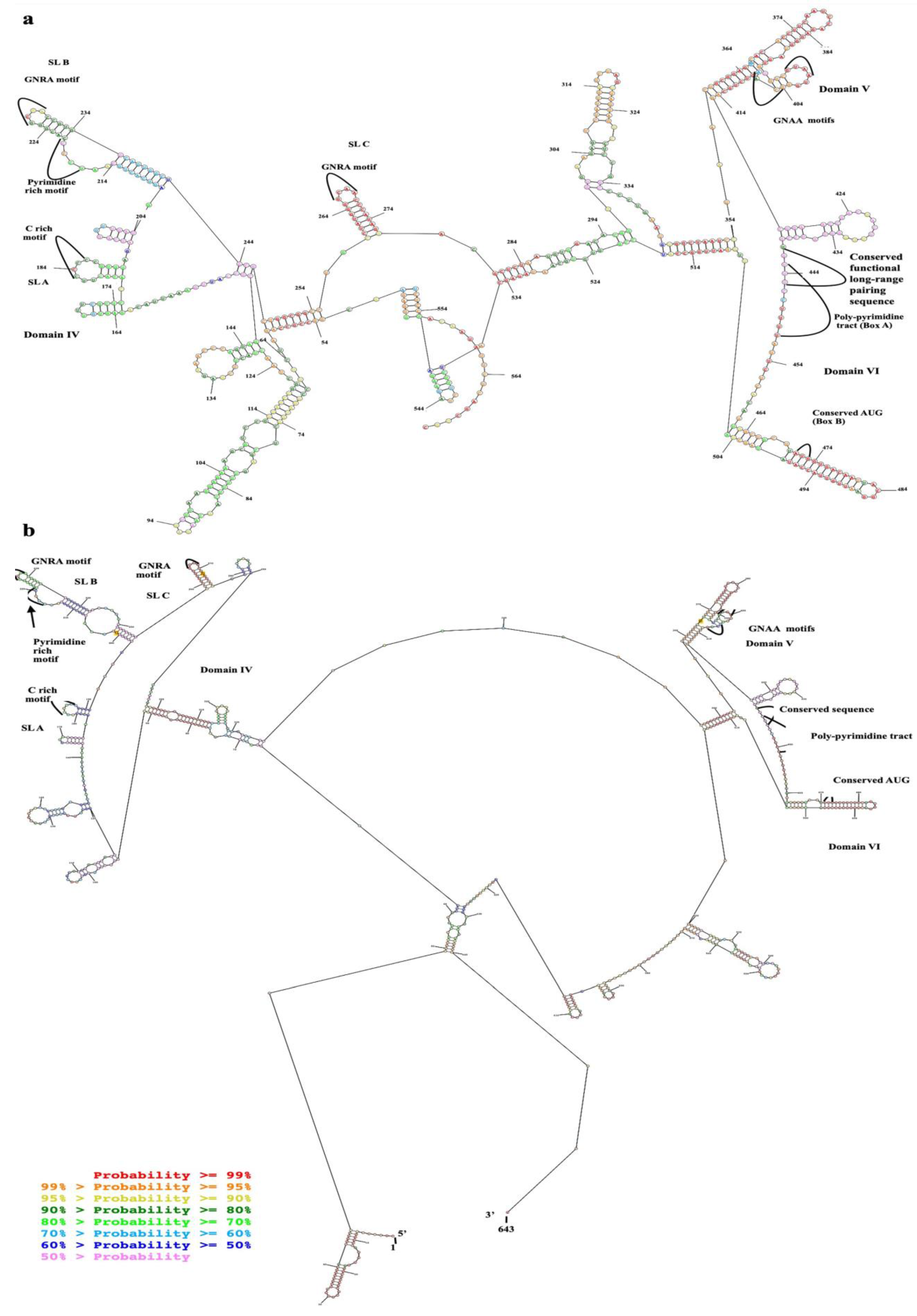
2.1. Domains IV, V, and VI Were Structurally Conserved Regions of EV-A71 IRES
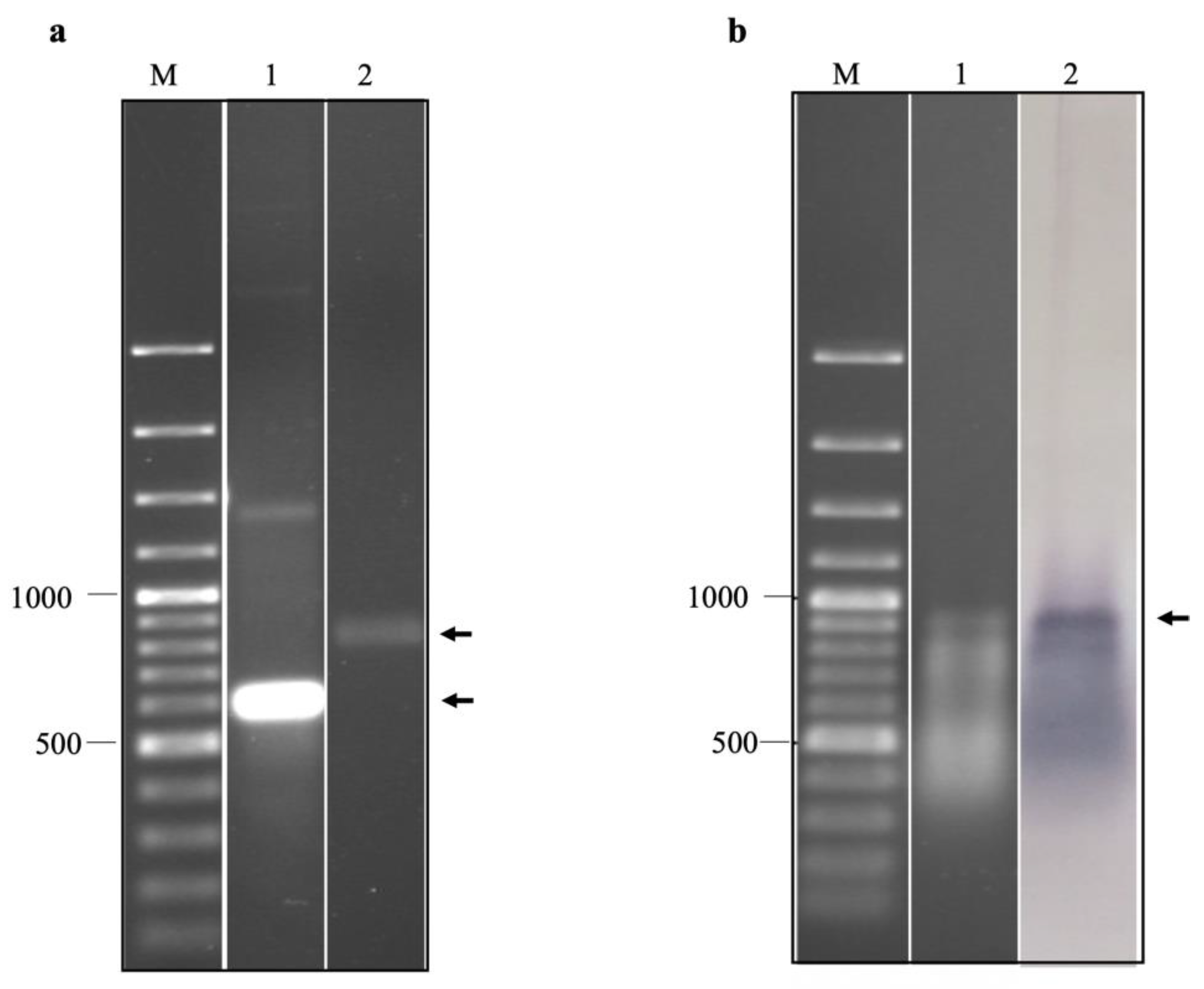
2.2. Production of EV-A71 IRES Domains IV, V, and VI RNA by In Vitro Transcription and Biotinylation
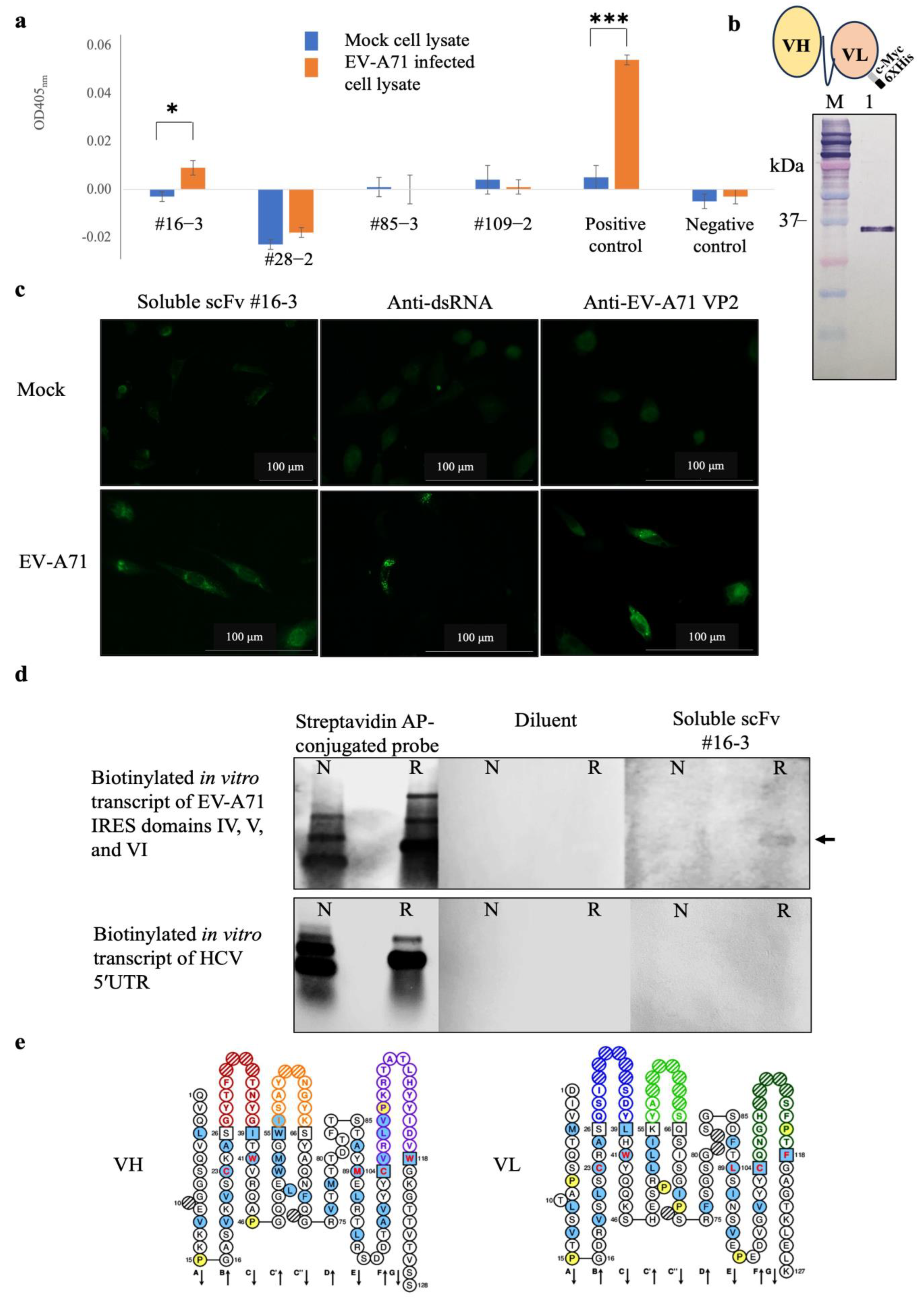
2.3. Characteristics of the Monoclonal scFv Antibody Specifically Binds EV-A71 IRES
2.4. Computational Prediction of Interaction between scFv and EV-A71 IRES
3. Discussion
4. Materials and Methods
4.1. Virus
4.2. Selection and Preparation of EV-A71 IRES Antigen
4.2.1. Identification of Conserved Regions of EV-A71 IRES
4.2.2. Preparation of the Selected Domains of EV-A71 IRES for Biopanning
4.3. Biopanning of Phage Display Single-Chain Variable Fragment Antibody (scFv)
4.3.1. Preparation of Antibody Phage Display Library
4.3.2. Immobilization of Antigens
4.3.3. Biopanning
4.4. Binding of the Selected scFv-Phages to EV-A71-Infected Cell Lysate
4.5. Production and Characterization of Soluble scFv Antibody
4.6. Antigenic Specificity of the Soluble scFv Antibody
4.7. Homology Modeling and Intermolecular Docking
4.8. Nucleotide Sequence
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishimaru, Y.; Nakano, S.; Yamaoka, K.; Takami, S. Outbreaks of hand, foot, and mouth disease by enterovirus 71. High incidence of complication disorders of central nervous system. Arch. Dis. Child. 1980, 55, 583–588. [Google Scholar] [CrossRef]
- Puenpa, J.; Wanlapakorn, N.; Vongpunsawad, S.; Poovorawan, Y. The History of Enterovirus A71 Outbreaks and Molecular Epidemiology in the Asia-Pacific Region. J. Biomed. Sci. 2019, 26, 75. [Google Scholar] [CrossRef] [PubMed]
- Kuo, R.-L.; Shih, S.-R. Strategies to develop antivirals against enterovirus 71. Virol. J. 2013, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Diarimalala, R.O.; Hu, M.; Wei, Y.; Hu, K. Recent advances of enterovirus 71 3Cpro targeting Inhibitors. Virol. J. 2020, 17, 173. [Google Scholar] [CrossRef]
- Ehrenfeld, E.; Domingo, E.; Roos, R.P.; American Society for, M. The Picornaviruses; ASM Press: Washington, DC, USA, 2010. [Google Scholar]
- Fitzgerald, K.D.; Semler, B.L. Bridging IRES elements in mRNAs to the eukaryotic translation apparatus. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2009, 1789, 518–528. [Google Scholar] [CrossRef]
- Sweeney, T.R.; Dhote, V.; Yu, Y.; Hellen, C.U. A distinct class of internal ribosomal entry site in members of the Kobuvirus and proposed Salivirus and Paraturdivirus genera of the Picornaviridae. J. Virol. 2012, 86, 1468–1486. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Miragall, O.; Lopez de Quinto, S.; Martinez-Salas, E. Relevance of RNA structure for the activity of picornavirus IRES elements. Virus Res. 2009, 139, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Salas, E.; Francisco-Velilla, R.; Fernandez-Chamorro, J.; Lozano, G.; Diaz-Toledano, R. Picornavirus IRES elements: RNA structure and host protein interactions. Virus Res. 2015, 206, 62–73. [Google Scholar] [CrossRef]
- Guan, L.; Hariharan, P. X-ray crystallography reveals molecular recognition mechanism for sugar binding in a melibiose transporter MelB. Commun. Biol. 2021, 4, 931. [Google Scholar] [CrossRef]
- Ostermeier, C.; Iwata, S.; Ludwig, B.; Michel, H. Fv fragment-mediated crystallization of the membrane protein bacterial cytochrome c oxidase. Nat. Struct. Biol. 1995, 2, 842. [Google Scholar] [CrossRef]
- Li, Y.; Yu, J.; Qi, X.; Yan, H. Monoclonal antibody against EV71 3Dpol inhibits the polymerase activity of RdRp and virus replication. BMC Immunol. 2019, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, K.J.; Ho, T.S.; Liu, C.C.; Lee, Y.R.; Lin, C.Y.; Shiuan, D.; Jiang, X.H. Monoclonal antibodies for diagnosis of enterovirus 71. Monoclon Antibodies Immunodiagn. Immunother. 2013, 32, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Pokkuluri, P.R.; Bouthillier, F.; Li, Y.; Kuderova, A.; Lee, J.; Cygler, M. Preparation, characterization and crystallization of an antibody Fab fragment that recognizes RNA. Crystal structures of native Fab and three Fab-mononucleotide complexes. J. Mol. Biol. 1994, 243, 283–297. [Google Scholar] [CrossRef]
- Lefranc, M.P.; Giudicelli, V.; Duroux, P.; Jabado-Michaloud, J.; Folch, G.; Aouinti, S.; Carillon, E.; Duvergey, H.; Houles, A.; Paysan-Lafosse, T.; et al. IMGT®, the international ImMunoGeneTics information system® 25 years on. Nucleic Acids Res. 2015, 43, D413–D422. [Google Scholar] [CrossRef] [PubMed]
- Souii, A.; Ben M’hadheb-Gharbi, M.; Gharbi, J. Role of RNA structure motifs in IRES-dependent translation initiation of the coxsackievirus B3: New insights for developing live-attenuated strains for vaccines and gene therapy. Mol. Biotechnol. 2013, 55, 179–202. [Google Scholar] [CrossRef]
- Koirala, D.; Lewicka, A.; Koldobskaya, Y.; Huang, H.; Piccirilli, J.A. Synthetic Antibody Binding to a Preorganized RNA Domain of Hepatitis C Virus Internal Ribosome Entry Site Inhibits Translation. ACS Chem. Biol. 2020, 15, 205–216. [Google Scholar] [CrossRef]
- Koide, S. Engineering of recombinant crystallization chaperones. Curr. Opin. Struct. Biol. 2009, 19, 449–457. [Google Scholar] [CrossRef]
- Murphy, K.; Travers, P.; Walport, M.; Janeway, C. Janeway’s Immunobiology; Garland Science: New York, NY, USA, 2012. [Google Scholar]
- Thanongsaksrikul, J.; Srimanote, P.; Tongtawe, P.; Glab-ampai, K.; Malik, A.A.; Supasorn, O.; Chiawwit, P.; Poovorawan, Y.; Chaicumpa, W. Identification and production of mouse scFv to specific epitope of enterovirus-71 virion protein-2 (VP2). Arch. Virol. 2018, 163, 1141–1152. [Google Scholar] [CrossRef]
- Thattanon, P.; Thanongsaksrikul, J.; Petvises, S.; Nathalang, O. Monoclonal antibody specific to the Di(a) blood group antigen generated by phage display technology. Blood Transfus 2020, 18, 366–373. [Google Scholar] [CrossRef]
- Jung, E.; Lee, J.; Hong, H.J.; Park, I.; Lee, Y. RNA recognition by a human antibody against brain cytoplasmic 200 RNA. RNA 2014, 20, 805–814. [Google Scholar] [CrossRef]
- Das, R.; Kudaravalli, M.; Jonikas, M.; Laederach, A.; Fong, R.; Schwans, J.P.; Baker, D.; Piccirilli, J.A.; Altman, R.B.; Herschlag, D. Structural inference of native and partially folded RNA by high-throughput contact mapping. Proc. Natl. Acad. Sci. USA 2008, 105, 4144–4149. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, N.G.; Gamarnik, A.V. Dynamic RNA structures in the dengue virus genome. RNA Biol. 2011, 8, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Tuplin, A. Diverse roles and interactions of RNA structures during the replication of positive-stranded RNA viruses of humans and animals. J. Gen. Virol. 2015, 96, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Salas, E. The impact of RNA structure on picornavirus IRES activity. Trends Microbiol. 2008, 16, 230–237. [Google Scholar] [CrossRef]
- Weber, F.; Wagner, V.; Rasmussen, S.B.; Hartmann, R.; Paludan, S.R. Double-stranded RNA is produced by positive-strand RNA viruses and DNA viruses but not in detectable amounts by negative-strand RNA viruses. J. Virol. 2006, 80, 5059–5064. [Google Scholar] [CrossRef]
- Son, K.-N.; Liang, Z.; Lipton, H.L. Double-Stranded RNA Is Detected by Immunofluorescence Analysis in RNA and DNA Virus Infections, Including Those by Negative-Stranded RNA Viruses. J. Virol. 2015, 89, 9383–9392. [Google Scholar] [CrossRef]
- Shao, Y.; Huang, H.; Qin, D.; Li, N.S.; Koide, A.; Staley, J.P.; Koide, S.; Kossiakoff, A.A.; Piccirilli, J.A. Specific Recognition of a Single-Stranded RNA Sequence by a Synthetic Antibody Fragment. J. Mol. Biol. 2016, 428, 4100–4114. [Google Scholar] [CrossRef]
- Ye, J.D.; Tereshko, V.; Frederiksen, J.K.; Koide, A.; Fellouse, F.A.; Sidhu, S.S.; Koide, S.; Kossiakoff, A.A.; Piccirilli, J.A. Synthetic antibodies for specific recognition and crystallization of structured RNA. Proc. Natl. Acad. Sci. USA 2008, 105, 82–87. [Google Scholar] [CrossRef]
- Koldobskaya, Y.; Duguid, E.M.; Shechner, D.M.; Suslov, N.B.; Ye, J.; Sidhu, S.S.; Bartel, D.P.; Koide, S.; Kossiakoff, A.A.; Piccirilli, J.A. A portable RNA sequence whose recognition by a synthetic antibody facilitates structural determination. Nat. Struct. Mol. Biol. 2011, 18, 100–106. [Google Scholar] [CrossRef]
- Sherman, E.M.; Holmes, S.; Ye, J.D. Specific RNA-binding antibodies with a four-amino-acid code. J. Mol. Biol. 2014, 426, 2145–2157. [Google Scholar] [CrossRef]
- Koirala, D.; Shao, Y.; Koldobskaya, Y.; Fuller, J.R.; Watkins, A.M.; Shelke, S.A.; Pilipenko, E.V.; Das, R.; Rice, P.A.; Piccirilli, J.A. A conserved RNA structural motif for organizing topology within picornaviral internal ribosome entry sites. Nat. Commun. 2019, 10, 3629. [Google Scholar] [CrossRef] [PubMed]
- Bellaousov, S.; Reuter, J.S.; Seetin, M.G.; Mathews, D.H. RNAstructure: Web servers for RNA secondary structure prediction and analysis. Nucleic Acids Res. 2013, 41, W471–W474. [Google Scholar] [CrossRef] [PubMed]
- Giudicelli, V.; Duroux, P.; Kossida, S.; Lefranc, M.-P. IG and TR single chain fragment variable (scFv) sequence analysis: A new advanced functionality of IMGT/V-QUEST and IMGT/HighV-QUEST. BMC Immunol. 2017, 18, 35. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK server for integrated protein–protein docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, D.; Zhou, P.; Li, B.; Huang, S.Y. HDOCK: A web server for protein-protein and protein-DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res. 2017, 45, W365–W373. [Google Scholar] [CrossRef]
- Yan, Y.; Wen, Z.; Wang, X.; Huang, S. Addressing recent docking challenges: A hybrid strategy to integrate template based and free proteié protein docking. Proteins Struct. Funct. Bioinform. 2017, 85, 497–512. [Google Scholar] [CrossRef]
- Huang, S.-Y.; Zou, X. A knowledge-based scoring function for protein-RNA interactions derived from a statistical mechanics-based iterative method. Nucleic Acids Res. 2014, 42, e55. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Zou, X. An iterative knowledge-based scoring function for protein-protein recognition. Proteins 2008, 72, 557–579. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motifs | Domains IV to VI with Flanking Regions of Cloning Plasmid (528 nts) | Whole IRES (Domains II to VI) (643 nts) |
|---|---|---|
| Minimal free energy (kcal/mol) | −192.6 | −227.1 |
| C rich motif (CCCC) of domain IV | in hairpin loop | in hairpin loop |
| Pyrimidine rich bulge (UCCCC) of domain IV | in lateral bulge | in lateral bulge |
| 1st GNRA motif (GUCA) of domain IV | in hairpin loop | in hairpin loop |
| 2nd GNRA motif (GGGA) of domain IV | in hairpin loop | in hairpin loop |
| 1st GNAA motif (GUAA) of domain V | in hairpin loop | in hairpin loop |
| 2nd GNAA motif (GCAA) of domain V | at the base of the hairpin | at the base of the hairpin |
| UGUUU in connecting region of domains V and VI | in multi-branch loop | in multi-branch loop |
| UUUCCUUUU (Box A) of domain VI | in multi-branch loop | in multi-branch loop |
| AUG (Box B) of domain VI | in stem/helix | in stem/helix |
| ScFv #16-3 | EV-A71 IRES | Interactive Bond | ||
|---|---|---|---|---|
| Residue | Domain | Nucleotide | Domain | |
| Q1 | VH-FR1 | G277 | V | Hydrogen Bond |
| V2 | VH-FR1 | U278 | V | Hydrogen Bond |
| N62 | VH-CDR2 | C107 | IV | Hydrogen Bond |
| G63 | VH-CDR2 | C66 | IV | Hydrogen Bond |
| G63 | VH-CDR2 | C67 | IV | Hydrogen Bond |
| Y64 | VH-CDR2 | C66 | IV | π-Hydrogen Bond |
| Y64 | VH-CDR2 | G105 | IV | Hydrogen Bond |
| Y64 | VH-CDR2 | C107 | IV | π-π interaction |
| R95 | VH-FR3 | A95 | IV | Salt Bridge, π-Alkyl |
| R106 | VH-CDR3 | G277 | V | Electrostatic |
| K110 | VH-CDR3 | C147 | IV | Electrostatic, Hydrogen Bond |
| R111 | VH-CDR3 | C56 | IV | Electrostatic, Hydrogen Bond |
| R111 | VH-CDR3 | A57 | IV | Electrostatic |
| R111 | VH-CDR3 | G145 | IV | Electrostatic |
| R111 | VH-CDR3 | G146 | IV | Electrostatic, π-Alkyl |
| T112 | VH-CDR3 | G108 | IV | Hydrogen Bond |
| W118 | VH-FR4 | U278 | V | Hydrogen Bond |
| S36 | VL-CDR1 | U54 | IV | Hydrogen Bond |
| R51 | VL-FR2 | G277 | V | Hydrogen Bond |
| S65 | VL-CDR2 | G1 | IV | Hydrogen Bond |
| S69 | VL-FR3 | C276 | V | Hydrogen Bond |
| R75 | VL-FR3 | G284 | V | Electrostatic |
| E97 | VL-FR3 | G283 | V | Hydrogen Bond |
| H108 | VL-CDR3 | A148 | IV | π-Anion |
| Genotype | Strain | GenBank® Accession No. | Country |
|---|---|---|---|
| A | BrCr, prototype | U22521 | USA |
| B1 | Nagoya | AB482183.1 | Japan |
| B2 | MS/7423/87 | U22522.1 | USA |
| B3 | SK-EV006 | AB550334.1 | Malaysia |
| B4 | 5666/sin/002209 | AF352027.1 | Singapore |
| B5 | THA-EV71-019 | JF738001.1 | Thailand |
| C1 | THA-EV71-002 | JF738000.1 | Thailand |
| C2 | Hau1 | KM077140.1 | Taiwan |
| C2 | 4643-TW98 | JN544418.1 | Taiwan |
| C3 | 03-KOR-00 | DQ341356.1 | Republic of Korea |
| C4 | SH12-036 | KC570452.1 | China |
| C5 | 2007-07364 | EU527983.1 | Taiwan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hlaing, S.T.; Srimanote, P.; Tongtawe, P.; Khantisitthiporn, O.; Glab-ampai, K.; Chulanetra, M.; Thanongsaksrikul, J. Isolation and Characterization of scFv Antibody against Internal Ribosomal Entry Site of Enterovirus A71. Int. J. Mol. Sci. 2023, 24, 9865. https://doi.org/10.3390/ijms24129865
Hlaing ST, Srimanote P, Tongtawe P, Khantisitthiporn O, Glab-ampai K, Chulanetra M, Thanongsaksrikul J. Isolation and Characterization of scFv Antibody against Internal Ribosomal Entry Site of Enterovirus A71. International Journal of Molecular Sciences. 2023; 24(12):9865. https://doi.org/10.3390/ijms24129865
Chicago/Turabian StyleHlaing, Su Thandar, Potjanee Srimanote, Pongsri Tongtawe, Onruedee Khantisitthiporn, Kittirat Glab-ampai, Monrat Chulanetra, and Jeeraphong Thanongsaksrikul. 2023. "Isolation and Characterization of scFv Antibody against Internal Ribosomal Entry Site of Enterovirus A71" International Journal of Molecular Sciences 24, no. 12: 9865. https://doi.org/10.3390/ijms24129865
APA StyleHlaing, S. T., Srimanote, P., Tongtawe, P., Khantisitthiporn, O., Glab-ampai, K., Chulanetra, M., & Thanongsaksrikul, J. (2023). Isolation and Characterization of scFv Antibody against Internal Ribosomal Entry Site of Enterovirus A71. International Journal of Molecular Sciences, 24(12), 9865. https://doi.org/10.3390/ijms24129865