
Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Projection Neurons and Interneurons of Basal Ganglia and Motor Thalamus in the Adult Human and Macaque Brains
 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. General Pattern of MCT8 and OATP1C1 Immunostaining in the Adult Human and Macaque Basal Ganglia Nuclei
2.2. MCT8 and OATP1C1 in Blood Vessels and Barriers in Macaque and Human Basal Ganglia
2.3. MCT8 and OATP1C1 Are Expressed in Input Nuclei of the Human and Macaque Basal Ganglia
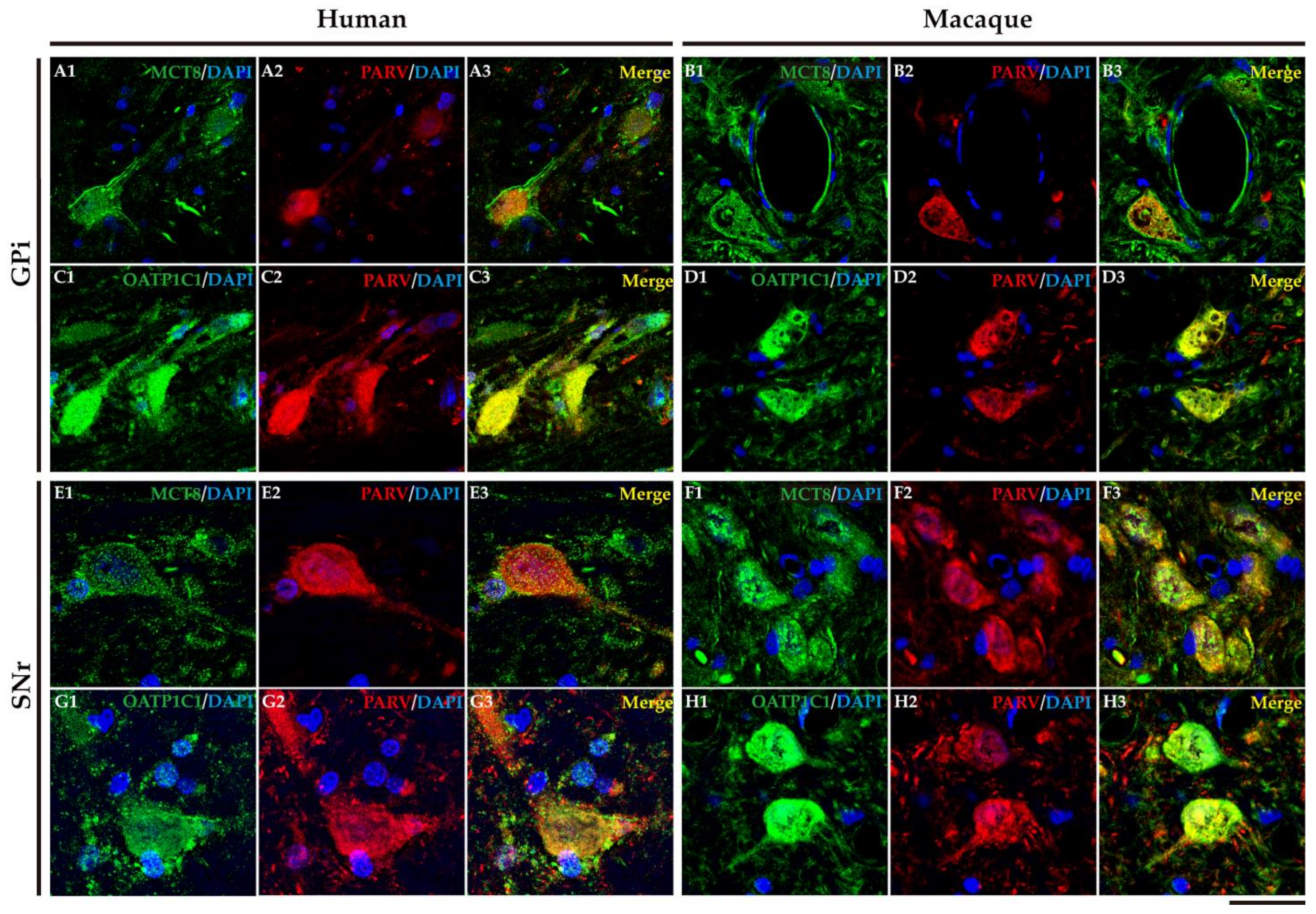
2.4. MCT8 and OATP1C1 Are Expressed in Projection Neurons of Output Nuclei of the Human and Macaque Basal Ganglia
2.5. MCT8 and OAdTP1C1 Are Expressed in Projection Neurons of the Human and Macaque Intrinsic Nuclei
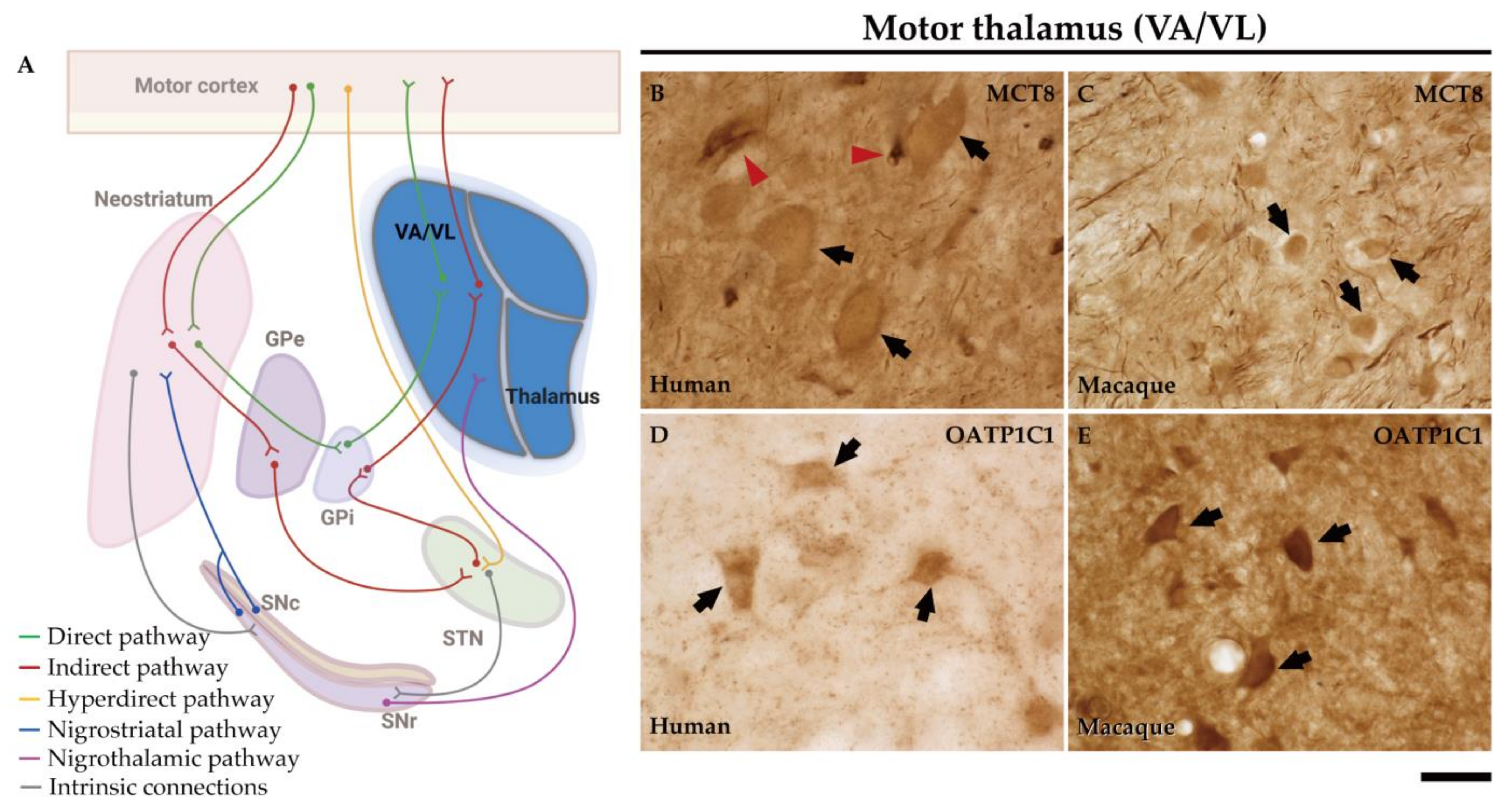
2.6. MCT8 and OATP1C1 Are Expressed in Thalamocortical Neurons of the Human and Macaque Motor Thalamus
2.7. MCT8 and OATP1C1 Are Expressed in the Cholinergic Neurons of the Human and Macaque Nucleus Basalis of Meynert (Ventral Pallidum)
3. Discussion
3.1. Study Limitations
3.2. Expression Pattern of TH Transporters in the Human and Nonhuman Primate Basal Ganglia and Thalamus
3.3. TH Transporters in the Nucleus Basalis of Meynert
3.4. Model of TH Transport in Human and Nonhuman Primate Neostriatum Microcircuitry
4. Materials and Methods
4.1. Human Samples and Tissue Preparation
4.2. Monkey Samples and Tissue Preparation
4.3. Histological Analysis
4.4. Immunohistochemistry (IHC)
4.5. Double/Triple Immunofluorescence (IF)
4.6. Image Acquisition and Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernal, J.; Guadaño-Ferraz, A.; Morte, B. Perspectives in the Study of Thyroid Hormone Action on Brain Development and Function. Thyroid 2003, 13, 1005–1012. [Google Scholar] [CrossRef]
- Tedeschi, L.; Vassalle, C.; Iervasi, G.; Sabatino, L. Main Factors Involved in Thyroid Hormone Action. Molecules 2021, 26, 7337. [Google Scholar] [CrossRef]
- Giammanco, M.; Di Liegro, C.M.; Schiera, G.; Di Liegro, I. Genomic and Non-Genomic Mechanisms of Action of Thyroid Hormones and Their Catabolite 3,5-Diiodo-L-Thyronine in Mammals. Int. J. Mol. Sci. 2020, 21, 4140. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.C.; Kim, B.W. Deiodinases: Implications of the local control of thyroid hormone action. J. Clin. Investig. 2006, 116, 2571–2579. [Google Scholar] [CrossRef] [PubMed]
- Guadaño-Ferraz, A.; Obregon, M.J.; St Germain, D.L.; Bernal, J. The type 2 iodothyronine deiodinase is expressed primarily in glial cells in the neonatal rat brain. Proc. Natl. Acad. Sci. USA 1997, 94, 10391–10396. [Google Scholar] [CrossRef]
- Friesema, E.C.; Ganguly, S.; Abdalla, A.; Manning Fox, J.E.; Halestrap, A.P.; Visser, T.J. Identification of Monocarboxylate Transporter 8 as a Specific Thyroid Hormone Transporter. J. Biol. Chem. 2003, 278, 40128–40135. [Google Scholar] [CrossRef]
- Pizzagalli, F.; Hagenbuch, B.; Stieger, B.; Klenk, U.; Folkers, G.; Meier, P.J. Identification of a Novel Human Organic Anion Transporting Polypeptide as a High Affinity Thyroxine Transporter. Mol. Endocrinol. 2002, 16, 2283–2296. [Google Scholar] [CrossRef]
- Bernal, J.; Guadaño-Ferraz, A.; Morte, B. Thyroid hormone transporters—Functions and clinical implications. Nat. Rev. Endocrinol. 2015, 11, 406–417. [Google Scholar] [CrossRef]
- Friesema, E.C.; Grueters, A.; Biebermann, H.; Krude, H.; von Moers, A.; Reeser, M.; Barrett, T.G.; E Mancilla, E.E.; Svensson, J.; Kester, M.H.; et al. Association between mutations in a thyroid hormone transporter and severe X-linked psychomotor retardation. Lancet 2004, 364, 1435–1437. [Google Scholar] [CrossRef]
- Dumitrescu, A.M.; Liao, X.-H.; Best, T.B.; Brockmann, K.; Refetoff, S. A Novel Syndrome Combining Thyroid and Neurological Abnormalities Is Associated with Mutations in a Monocarboxylate Transporter Gene. Am. J. Hum. Genet. 2004, 74, 168–175. [Google Scholar] [CrossRef]
- Tonduti, D.; Vanderver, A.; Berardinelli, A.; Schmidt, J.L.; Collins, C.D.; Novara, F.; Genni, A.D.; Mita, A.; Triulzi, F.; Brunstrom-Hernandez, J.E.; et al. MCT8 Deficiency: Extrapyramidal symptoms and delayed myelination as prominent features. J. Child Neurol. 2013, 28, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Masnada, S.; Sarret, C.; Antonello, C.E.; Fadilah, A.; Krude, H.; Mura, E.; Mordekar, S.; Nicita, F.; Olivotto, S.; Orcesi, S.; et al. Movement disorders in MCT8 deficiency/Allan-Herndon-Dudley Syndrome. Mol. Genet. Metab. 2022, 135, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Remerand, G.; Boespflug-Tanguy, O.; Tonduti, D.; Touraine, R.; Rodriguez, D.; Curie, A.; Perreton, N.; Des Portes, V.; Sarret, C.; Group, R.A.S. Expanding the phenotypic spectrum of Allan–Herndon–Dudley syndrome in patients with SLC 16A2 mutations. Dev. Med. Child Neurol. 2019, 61, 1439–1447. [Google Scholar] [CrossRef]
- Ono, E.; Ariga, M.; Oshima, S.; Hayakawa, M.; Imai, M.; Ochiai, Y.; Mochizuki, H.; Namba, N.; Ozono, K.; Miyata, I. Three novel mutations of the MCT8 (SLC16A2) gene: Individual and temporal variations of endocrinological and radiological features. Clin. Pediatr. Endocrinol. 2016, 25, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Groeneweg, S.; van Geest, F.S.; Abacı, A.; Alcantud, A.; Ambegaonkar, G.P.; Armour, C.M.; Bakhtiani, P.; Barca, D.; Bertini, E.S.; van Beynum, I.M.; et al. Disease characteristics of MCT8 deficiency: An international, retrospective, multicentre cohort study. Lancet Diabetes Endocrinol. 2020, 8, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Zhao, P.; Chunquan, C.; Chen, J.; Chen, S.; Sheng, Q. Identification of a novel SLC16A2 mutation that causes Allan-Herndon-Dudley syndrome. Tianjin Med. J. 2020, 48, 132–136. [Google Scholar] [CrossRef]
- Kubota, M.; Yakuwa, A.; Terashima, H.; Hoshino, H. A nationwide survey of monocarboxylate transporter 8 deficiency in Japan: Its incidence, clinical course, MRI and laboratory findings. Brain Dev. 2022, 44, 699–705. [Google Scholar] [CrossRef]
- Chen, X.; Liu, L.; Zeng, C. A novel variant in SLC16A2 associated with typical Allan-Herndon-Dudley syndrome: A case report. BMC Pediatr. 2022, 22, 180. [Google Scholar] [CrossRef]
- Strømme, P.; Groeneweg, S.; Lima de Souza, E.C.; Zevenbergen, C.; Torgersbråten, A.; Holmgren, A.; Gurcan, E.; Meima, M.E.; Peeters, R.P.; Visser, W.E.; et al. Mutated Thyroid Hormone Transporter OATP1C1 Associates with Severe Brain Hypometabolism and Juvenile Neurodegeneration. Thyroid 2018, 28, 1406–1415. [Google Scholar] [CrossRef]
- López-Espíndola, D.; Morales-Bastos, C.; Grijota-Martinez, C.; Liao, X.-H.; Lev, D.; Sugo, E.; Verge, C.F.; Refetoff, S.; Bernal, J.; Guadaño-Ferraz, A. Mutations of the Thyroid Hormone Transporter MCT8 Cause Prenatal Brain Damage and Persistent Hypomyelination. J. Clin. Endocrinol. Metab. 2014, 99, E2799–E2804. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, T.; Montero-Pedrazuela, A.; Guadaño-Ferraz, A.; Rausell, E. Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Pyramidal Neurons and Interneurons in the Adult Motor Cortex of Human and Macaque Brain. Int. J. Mol. Sci. 2023, 24, 3207. [Google Scholar] [CrossRef] [PubMed]
- Haines, D.E.; Mihailoff, G.A. Fundamental Neuroscience for Basic and Clinical Applications, 5th ed.; Elsevier: Philadelphia, PA, USA, 2018; 528p. [Google Scholar]
- Lanciego, J.L.; Luquin, N.; Obeso, J.A. Functional Neuroanatomy of the Basal Ganglia. Cold Spring Harb. Perspect. Med. 2012, 2, a009621. [Google Scholar] [CrossRef]
- Albin, R.L.; Young, A.B.; Penney, J.B. The functional anatomy of disorders of the basal ganglia. Trends Neurosci. 1995, 18, 63–64. [Google Scholar] [CrossRef]
- Brazis, P.W.; Masdeu, J.C.; Biller, J. Localization in Clinical Neurology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- DeLong, M.R. Primate models of movement disorders of basal ganglia origin. Trends Neurosci. 1990, 13, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Gerfen, C.R.; Engber, T.M.; Mahan, L.C.; Susel, Z.; Chase, T.N.; Monsma, F.J., Jr.; Sibley, D.R. D1 and D2 Dopamine Receptor-regulated Gene Expression of Striatonigral and Striatopallidal Neurons. Science 1990, 250, 1429–1432. [Google Scholar] [CrossRef]
- Benarroch, E.E. Intrinsic circuits of the striatum: Complexity and clinical correlations. Neurology 2016, 86, 1531–1542. [Google Scholar] [CrossRef]
- Prensa, L.; Gimenez-Amaya, J.M.; Parent, A. Chemical heterogeneity of the striosomal compartment in the human striatum. J. Comp. Neurol. 1999, 413, 603–618. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Wilson, C.J.; Augood, S.J.; Emson, P.C. Striatal interneurones: Chemical, physiological and morphological characterization. Trends Neurosci. 1995, 18, 527–535. [Google Scholar] [CrossRef]
- Bernácer, J.; Prensa, L.; Giménez-Amaya, J.M. Morphological features, distribution and compartmental organization of the nicotinamide adenine dinucleotide phosphate reduced-diaphorase interneurons in the human striatum. J. Comp. Neurol. 2005, 489, 311–327. [Google Scholar] [CrossRef]
- Kita, H. Globus pallidus external segment. In Gaba and the Basal Ganglia—From Molecules to Systems; Progress in Brain Research; Elsevier: Philadelphia, PA, USA, 2007; pp. 111–133. [Google Scholar]
- Nambu, A. Globus pallidus internal segment. In Gaba and the Basal Ganglia—From Molecules to Systems; Progress in Brain Research; Elsevier: Philadelphia, PA, USA, 2007; pp. 135–150. [Google Scholar]
- Gonzalez-Hernandez, T.; Rodriguez, M. Compartmental organization and chemical profile of dopaminergic and GABAergic neurons in the substantia nigra of the rat. J. Comp. Neurol. 2000, 421, 107–135. [Google Scholar] [CrossRef]
- Levesque, J.C.; Parent, A. GABAergic interneurons in human subthalamic nucleus. Mov. Disord. 2005, 20, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G. The Thalamus; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Rausell, E.; Bae, C.S.; Vinuela, A.; Huntley, G.W.; Jones, E.G. Calbindin and parvalbumin cells in monkey VPL thalamic nucleus: Distribution, laminar cortical projections, and relations to spinothalamic terminations. J. Neurosci. 1992, 12, 4088–4111. [Google Scholar] [CrossRef] [PubMed]
- Rausell, E.; Jones, E.G. Chemically distinct compartments of the thalamic VPM nucleus in monkeys relay principal and spinal trigeminal pathways to different layers of the somatosensory cortex. J. Neurosci. 1991, 11, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Heuer, H. Expression Pattern of Thyroid Hormone Transporters in the Postnatal Mouse Brain. Front. Endocrinol. 2014, 5, 92. [Google Scholar] [CrossRef]
- Heuer, H.; Maier, M.K.; Iden, S.; Mittag, J.; Friesema, E.C.; Visser, T.J.; Bauer, K. The Monocarboxylate Transporter 8 Linked to Human Psychomotor Retardation Is Highly Expressed in Thyroid Hormone-Sensitive Neuron Populations. Endocrinology 2005, 146, 1701–1706. [Google Scholar] [CrossRef]
- Consortium, G. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Visser, W.E.; Swagemakers, S.M.; Őzgűr, Z.; Schot, R.; Verheijen, F.W.; van Ijcken, W.F.; van der Spek, P.J.; Visser, T.J. Transcriptional profiling of fibroblasts from patients with mutations in MCT8 and comparative analysis with the human brain transcriptome. Hum. Mol. Genet. 2010, 19, 4189–4200. [Google Scholar] [CrossRef]
- Grijota-Martínez, C.; Bárez-López, S.; Gómez-Andrés, D.; Guadaño-Ferraz, A. MCT8 Deficiency: The Road to Therapies for a Rare Disease. Front. Neurosci. 2020, 14, 380. [Google Scholar] [CrossRef]
- Mai, J.K.; Majtanik, M.; Paxinos, G. Atlas of the Human Brain; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Winters, W.D.; Kado, R.; Adey, W.R. A Stereotaxic Brain atlas for Macaca nemestrina; University of California Press: Oakland, CA, USA, 1969. [Google Scholar]
- Ito, K.; Uchida, Y.; Ohtsuki, S.; Aizawa, S.; Kawakami, H.; Katsukura, Y.; Kamiie, J.; Terasaki, T. Quantitative Membrane Protein Expression at the Blood–Brain Barrier of Adult and Younger Cynomolgus Monkeys. J. Pharm. Sci. 2011, 100, 3939–3950. [Google Scholar] [CrossRef]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the Thyroid Hormone Transporters Monocarboxylate Transporter-8 (SLC16A2) and Organic Ion Transporter-14 (SLCO1C1) at the Blood-Brain Barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef]
- Alkemade, A.; Friesema, E.C.; Kalsbeek, A.; Swaab, D.F.; Visser, T.J.; Fliers, E. Expression of Thyroid Hormone Transporters in the Human Hypothalamus. J. Clin. Endocrinol. Metab. 2011, 96, E967–E971. [Google Scholar] [CrossRef]
- Riba, M.; Augé, E.; Campo-Sabariz, J.; Moral-Anter, D.; Molina-Porcel, L.; Ximelis, T.; Ferrer, R.; Martín-Venegas, R.; Pelegrí, C.; Vilaplana, J. Corpora amylacea act as containers that remove waste products from the brain. Proc. Natl. Acad. Sci. USA 2019, 116, 26038–26048. [Google Scholar] [CrossRef] [PubMed]
- Augé, E.; Bechmann, I.; Llor, N.; Vilaplana, J.; Krueger, M.; Pelegrí, C. Corpora amylacea in human hippocampal brain tissue are intracellular bodies that exhibit a homogeneous distribution of neo-epitopes. Sci. Rep. 2019, 9, 2063. [Google Scholar] [CrossRef] [PubMed]
- Augé, E.; Cabezón, I.; Pelegrí, C.; Vilaplana, J. New perspectives on corpora amylacea in the human brain. Sci. Rep. 2017, 7, 41807. [Google Scholar] [CrossRef]
- Jones, E.G. Synchrony in the Interconnected Circuitry of the Thalamus and Cerebral Cortex. Ann. N. Y. Acad. Sci. 2009, 1157, 10–23. [Google Scholar] [CrossRef]
- Jones, E.G. The thalamic matrix and thalamocortical synchrony. Trends Neurosci. 2001, 24, 595–601. [Google Scholar] [CrossRef]
- Liu, A.K.L.; Chang, R.C.-C.; Pearce, R.K.B.; Gentleman, S.M. Nucleus basalis of Meynert revisited: Anatomy, history and differential involvement in Alzheimer’s and Parkinson’s disease. Acta Neuropathol. 2015, 129, 527–540. [Google Scholar] [CrossRef]
- Boelen, A.; Kwakkel, J.; Fliers, E. Beyond Low Plasma T3: Local Thyroid Hormone Metabolism during Inflammation and Infection. Endocr. Rev. 2011, 32, 670–693. [Google Scholar] [CrossRef]
- López-Espíndola, D.; García-Aldea, A.; Gomez de la Riva, I.; Rodríguez-García, A.M.; Salvatore, D.; Visser, T.J.; Bernal, J.; Guadaño-Ferraz, A. Thyroid hormone availability in the human fetal brain: Novel entry pathways and role of radial glia. Brain Struct. Funct. 2019, 224, 2103–2119. [Google Scholar] [CrossRef]
- Wilpert, N.-M.; Krueger, M.; Opitz, R.; Sebinger, D.; Paisdzior, S.; Mages, B.; Schulz, A.; Spranger, J.; Wirth, E.K.; Stachelscheid, H.; et al. Spatiotemporal Changes of Cerebral Monocarboxylate Transporter 8 Expression. Thyroid 2020, 30, 1366–1383. [Google Scholar] [CrossRef]
- Alkemade, A.; Friesema, E.C.; Unmehopa, U.A.; Fabriek, B.O.; Kuiper, G.G.; Leonard, J.L.; Wiersinga, W.M.; Swaab, D.F.; Visser, T.J.; Fliers, E. Neuroanatomical Pathways for Thyroid Hormone Feedback in the Human Hypothalamus. J. Clin. Endocrinol. Metab. 2005, 90, 4322–4334. [Google Scholar] [CrossRef]
- Friesema, E.C.; Visser, T.J.; Borgers, A.J.; Kalsbeek, A.; Swaab, D.F.; Fliers, E.; Alkemade, A. Thyroid hormone transporters and deiodinases in the developing human hypothalamus. Eur. J. Endocrinol. 2012, 167, 379–386. [Google Scholar] [CrossRef]
- Hennemann, G.; Docter, R.; Friesema, E.C.; de Jong, M.; Krenning, E.P.; Visser, T.J. Plasma Membrane Transport of Thyroid Hormones and Its Role in Thyroid Hormone Metabolism and Bioavailability. Endocr. Rev. 2001, 22, 451–476. [Google Scholar] [CrossRef]
- Visser, W.E.; Friesema, E.C.; Jansen, J.; Visser, T.J. Thyroid hormone transport in and out of cells. Trends Endocrinol. Metab. 2008, 19, 50–56. [Google Scholar] [CrossRef]
- Morreale de Escobar, G.; Obregon, M.J.; Escobar del Rey, F. Role of thyroid hormone during early brain development. Eur. J. Endocrinol. 2004, 151, U25–U37. [Google Scholar] [CrossRef]
- Lu, E.J.; Brown, W.J. An electron microscopic study of the developing caudate nucleus in euthyroid and hypothyroid states. Anat. Embryol. 1977, 150, 335–364. [Google Scholar] [CrossRef]
- Morte, B.; Gil-Ibañez, P.; Heuer, H.; Bernal, J. Brain Gene Expression in Systemic Hypothyroidism and Mouse Models of MCT8 Deficiency: The Mct8-Oatp1c1-Dio2 Triad. Thyroid 2021, 31, 985–993. [Google Scholar] [CrossRef]
- Vargiu, P.; Morte, B.; Manzano, J.; Perez, J.; de Abajo, R.; Gregor Sutcliffe, J.; Bernal, J. Thyroid hormone regulation of rhes, a novel Ras homolog gene expressed in the striatum. Brain Res. Mol. Brain Res. 2001, 94, 1–8. [Google Scholar] [CrossRef]
- Manzano, J.; Morte, B.; Scanlan, T.S.; Bernal, J. Differential Effects of Triiodothyronine and the Thyroid Hormone Receptor β-Specific Agonist GC-1 on Thyroid Hormone Target Genes in the Brain. Endocrinology 2003, 144, 5480–5487. [Google Scholar] [CrossRef]
- Vallortigara, J.; Chassande, O.; Higueret, P.; Enderlin, V. Thyroid Hormone Receptor Alpha Plays an Essential Role in the Normalisation of Adult-Onset Hypothyroidism-Related Hypoexpression of Synaptic Plasticity Target Genes in Striatum. J. Neuroendocr. 2009, 21, 49–56. [Google Scholar] [CrossRef]
- Dussault, J.H.; Ruel, J. Thyroid Hormones and Brain Development. Annu. Rev. Physiol. 1987, 49, 321–334. [Google Scholar] [CrossRef]
- Oh, J.D.; Butcher, L.L.; Woolf, N.J. Thyroid hormone modulates the development of cholinergic terminal fields in the rat forebrain: Relation to nerve growth factor receptor. Brain Res. Dev. Brain Res. 1991, 59, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Hayashi, M.; Hunt, A. Role of Thyroid Hormone and Nerve Growth Factor in the Development of Choline Acetyltransferase and Other Cell-Specific Marker Enzymes in the Basal Forebrain of the Rat. J. Neurochem. 1988, 50, 803–811. [Google Scholar] [CrossRef]
- Patel, A.J.; Hayashi, M.; Hunt, A. Selective persistent reduction in choline acetyltransferase activity in basal forebrain of the rat after thyroid deficiency during early life. Brain Res. 1987, 422, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Butcher, L.L. Developing cholinergic basal forebrain neurons are sensitive to thyroid hormone. J. Neurosci. 1989, 9, 3347–3358. [Google Scholar] [CrossRef]
- Calzà, L.; Giardino, L.; Aloe, L. Thyroid Hormone Regulates NGF Content and p75LNGFRExpression in the Basal Forebrain of Adult Rats. Exp. Neurol. 1997, 143, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.K.; Ray, A.K. Involvement of L-Triiodothyronine in Acetylcholine Metabolism in Adult Rat Cerebrocortical Synaptosomes. Horm. Metab. Res. 2001, 33, 270–275. [Google Scholar] [CrossRef]
- Moskovkin, G.N.; Kálmán, M.; Kardos, J.; Yarigin, K.N.; Hajós, F. Effect of Triiodothryonine on the Muscarinic Receptors and Acetylcholinesterase Activity of Developing Rat Brain. Int. J. Neurosci. 1989, 44, 83–89. [Google Scholar] [CrossRef]
- Lebel, J.M.; Dussault, J.H.; Puymirat, J. Overexpression of the beta 1 thyroid receptor induces differentiation in neuro-2a cells. Proc. Natl. Acad. Sci. USA 1994, 91, 2644–2648. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Bower, J.M. Acetylcholine and memory. Trends Neurosci. 1993, 16, 218–222. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P. Cognitive functions of cortical acetylcholine: Toward a unifying hypothesis. Brain Res. Rev. 1997, 23, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol. 2006, 16, 710–715. [Google Scholar] [CrossRef]
- Quirin-Stricker, C.; Nappey, V.; Simoni, P.; Toussaint, J.L.; Schmitt, M. Trans-activation by thyroid hormone receptors of the 5′ flanking region of the human ChAT gene. Mol. Brain Res. 1994, 23, 253–265. [Google Scholar] [CrossRef]
- Juarez de Ku, L.M.; Sharma-Stokkermans, M.; Meserve, L.A. Thyroxine normalizes polychlorinated biphenyl (PCB) dose-related depression of choline acetyltransferase (ChAT) activity in hippocampus and basal forebrain of 15-day-old rats. Toxicology 1994, 94, 19–30. [Google Scholar] [CrossRef]
- Sawin, S.; Brodish, P.; Carter, C.S.; Stanton, M.E.; Lau, C. Development of Cholinergic Neurons in Rat Brain Regions: Dose-Dependent Effects of Propylthiouracil-Induced Hypothyroidism. Neurotoxicol. Teratol. 1998, 20, 627–635. [Google Scholar] [CrossRef]
- Provost, T.L.; Juarez de Ku, L.M.; Zender, C.; Meserve, L.A. Dose- and age-dependent alterations in choline acetyltransferase (ChAT) activity, learning and memory, and thyroid hormones in 15- and 30-day old rats exposed to 1.25 or 12.5 ppm polychlorinated biphenyl (PCB) beginning at conception. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1999, 23, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.J. The generation of natural firing patterns in neostriatal neurons. Prog. Brain Res. 1993, 99, 277–297. [Google Scholar] [CrossRef]
- Berke, J.D.; Okatan, M.; Skurski, J.; Eichenbaum, H.B. Oscillatory Entrainment of Striatal Neurons in Freely Moving Rats. Neuron 2004, 43, 883–896. [Google Scholar] [CrossRef]
- Adler, A.; Katabi, S.; Finkes, I.; Israel, Z.; Prut, Y.; Bergman, H. Temporal Convergence of Dynamic Cell Assemblies in the Striato-Pallidal Network. J. Neurosci. 2012, 32, 2473–2484. [Google Scholar] [CrossRef]
- Purves, D.; Augustine, G.J.; Fitzpatrick, D.; Hall, W.C.; LaManita, A.-S.; White, L.E. Neuroscience, 5th ed.; Sinauer Associates: Sunderland, MA, USA, 2012; p. 759. [Google Scholar]
- Iglesias, J.E.; Insausti, R.; Lerma-Usabiaga, G.; Bocchetta, M.; Van Leemput, K.; Greve, D.N.; van der Kouwe, A.; Fischl, B.; Caballero-Gaudes, C.; Paz-Alonso, P.M. A probabilistic atlas of the human thalamic nuclei combining ex vivo MRI and histology. Neuroimage 2018, 183, 314–326. [Google Scholar] [CrossRef]
- Uroz, V.; Prensa, L.; Giménez-Amaya, J.M. Chemical anatomy of the human paraventricular thalamic nucleus. Synapse 2004, 51, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Guadaño-Ferraz, A.; Viñuela, A.; Oeding, G.; Bernal, J.; Rausell, E. RC3/neurogranin is expressed in pyramidal neurons of motor and somatosensory cortex in normal and denervated monkeys. J. Comp. Neurol. 2005, 493, 554–570. [Google Scholar] [CrossRef] [PubMed]
- Emmers, R.; Akert, K. A Stereotaxic Atlas of the Brain of the Squirrel Monkey (Saimiri sciureus); The University of Wisconsin Press: Madison, WI, USA, 1963. [Google Scholar]
- Smyth, L.C.D.; Rustenhoven, J.; Scotter, E.L.; Schweder, P.; Faull, R.L.M.; Park, T.I.H.; Dragunow, M. Markers for human brain pericytes and smooth muscle cells. J. Chem. Neuroanat. 2018, 92, 48–60. [Google Scholar] [CrossRef]
- Kadota, E.; Tanji, K.; Nishida, S.; Takahashi, M.; Maeda, M.; Hiruma, S.; Enomoto, Y.; Hashimoto, S.; Akai, F. Lectin (UEA-1) reaction of capillary endothelium with reference to permeability in autopsied cases of cerebral infarction. Histol. Histopathol. 1986, 1, 219–226. [Google Scholar] [PubMed]
- Rausell, E.; Avendaño, C. Thalamocortical neurons projecting to superficial and to deep layers in parietal, frontal and prefrontal regions in the cat. Brain Res. 1985, 347, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gerfen, C.R.; Baimbridge, K.G.; Miller, J.J. The neostriatal mosaic: Compartmental distribution of calcium-binding protein and parvalbumin in the basal ganglia of the rat and monkey. Proc. Natl. Acad. Sci. USA 1985, 82, 8780–8784. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Age (y) | Postmortem Interval (h) | Brain Weight (g) | Cause of Death |
|---|---|---|---|---|
| Male | 29 | 4 | 1500 | Lung tumor |
| Male | 32 | 3 | 1420 | Hemorrhagic gastroenteritis |
| Male | 54 | 12 | 1350 | Aortic aneurysm |
| Male | 59 | <24 | 1020 | Pneumonia |
| Male | 86 | <24 | Not available | Not available |
| Male | 97 | 9 | 1238 | Septic shock |
| Female | 98 | 6 | 1168 | Not available |
| Species | Age (y) | Sex |
|---|---|---|
| M. fascicularis | 3 | Female |
| Saimiri sciureus | 3 | Female |
| M. fascicularis | 5 | Male |
| M. fascicularis | 5 | Male |
| M. fascicularis | 5 | Female |
| M. fascicularis | 5 | Male |
| M. fascicularis | 5 | Male |
| M. fascicularis | 5 | Female |
| M. fascicularis | 5 | Male |
| M. mulatta | 7 | Male |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Wang, Y.; Montero-Pedrazuela, A.; Prensa, L.; Guadaño-Ferraz, A.; Rausell, E. Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Projection Neurons and Interneurons of Basal Ganglia and Motor Thalamus in the Adult Human and Macaque Brains. Int. J. Mol. Sci. 2023, 24, 9643. https://doi.org/10.3390/ijms24119643
Wang T, Wang Y, Montero-Pedrazuela A, Prensa L, Guadaño-Ferraz A, Rausell E. Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Projection Neurons and Interneurons of Basal Ganglia and Motor Thalamus in the Adult Human and Macaque Brains. International Journal of Molecular Sciences. 2023; 24(11):9643. https://doi.org/10.3390/ijms24119643
Chicago/Turabian StyleWang, Ting, Yu Wang, Ana Montero-Pedrazuela, Lucía Prensa, Ana Guadaño-Ferraz, and Estrella Rausell. 2023. "Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Projection Neurons and Interneurons of Basal Ganglia and Motor Thalamus in the Adult Human and Macaque Brains" International Journal of Molecular Sciences 24, no. 11: 9643. https://doi.org/10.3390/ijms24119643
APA StyleWang, T., Wang, Y., Montero-Pedrazuela, A., Prensa, L., Guadaño-Ferraz, A., & Rausell, E. (2023). Thyroid Hormone Transporters MCT8 and OATP1C1 Are Expressed in Projection Neurons and Interneurons of Basal Ganglia and Motor Thalamus in the Adult Human and Macaque Brains. International Journal of Molecular Sciences, 24(11), 9643. https://doi.org/10.3390/ijms24119643