

Fangs in the Ghats: Preclinical Insights into the Medical Importance of Pit Vipers from the Western Ghats

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Venom Proteomics
2.2. Venom Biochemistry
2.3. Binding Efficiency and Cross-Reactivity of Commercial Antivenoms
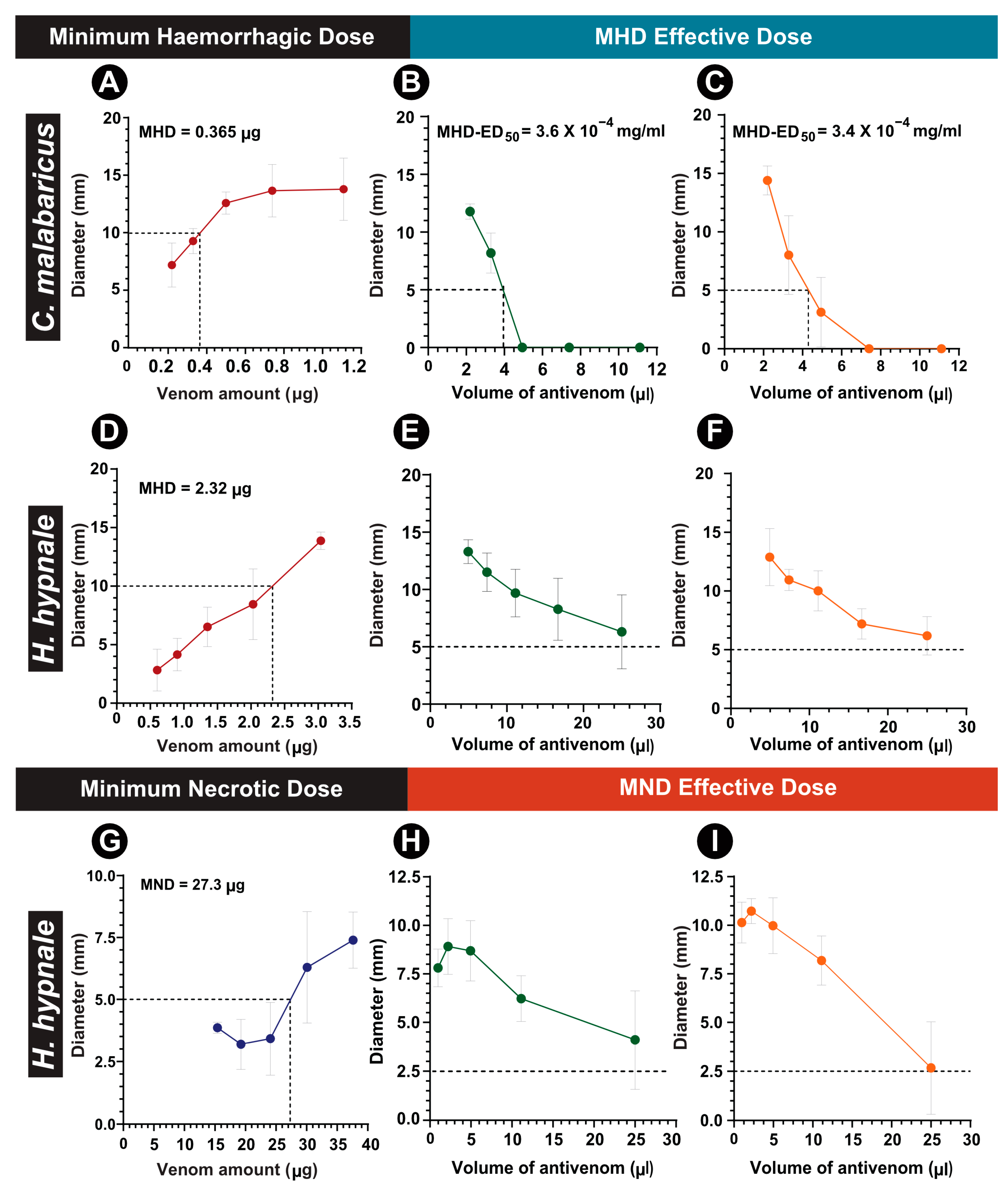
2.4. Preclinical Assessments
Venom Toxicity
2.5. Histopathological Evaluations
2.5.1. Renal Histology
2.5.2. Skeleton Muscle Histology
2.5.3. Neutralisation of Lethal and Morbid Effects of Pit Viper Venoms by Indian and Sri Lankan Polyvalent Antivenoms
3. Discussion
3.1. Venom Compositions of Pit Vipers from the Western Ghats
3.2. Clinically Important Venoms of Pit Vipers from the Western Ghats
3.3. Snakebite-Associated Acute Kidney Injury (SAKI)
3.4. Preclinical Failure of Commercial Polyvalent Antivenoms Highlights the Urgent Need for Region-Specific Antivenom Therapy
4. Materials and Methods
4.1. Venom and Antivenom Samples
4.2. Proteomic Analyses
4.2.1. Protein Estimation, One-Dimensional Gel Electrophoresis and In-Gel Digestion
4.2.2. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
4.3. Biochemical Characterization
4.3.1. Colorimetric Phospholipase A2 (PLA2) Assay
4.3.2. L-amino Acid Oxidase (LAAO) Assay
4.3.3. Snake Venom Protease Assay
4.3.4. Fibrinogenolytic Assay
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Preclinical Assessments
4.5.1. The Median Lethal Dose (LD50)
4.5.2. The Median Effective Dose (ED50)
4.5.3. The Minimum Haemorrhagic Dose (MHD)
4.5.4. The Minimum Necrotic Dose (MND)
4.6. Neutralisation of Venom-Induced Morbidities
4.7. Histopathological Evaluations
4.7.1. Renal Histology
4.7.2. Skeletal Muscle Histology
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suraweera, W.; Warrell, D.; Whitaker, R.; Menon, G.; Rodrigues, R.; Fu, S.H.; Begum, R.; Sati, P.; Piyasena, K.; Bhatia, M.; et al. Trends in snakebite deaths in India from 2000 to 2019 in a nationally representative mortality study. eLife 2020, 9, e54076. [Google Scholar] [CrossRef] [PubMed]
- Laxme, R.S.; Khochare, S.; de Souza, H.F.; Ahuja, B.; Suranse, V.; Martin, G.; Whitaker, R.; Sunagar, K. Beyond the ‘big four’: Venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl. Trop. Dis. 2019, 13, e0007899. [Google Scholar]
- Dutta, S.; Chanda, A.; Kalita, B.; Islam, T.; Patra, A.; Mukherjee, A.K. Proteomic analysis to unravel the complex venom proteome of eastern India Naja naja: Correlation of venom composition with its biochemical and pharmacological properties. J. Proteom. 2017, 156, 29–39. [Google Scholar] [CrossRef]
- Chanda, A.; Mukherjee, A.K. Mass spectrometric analysis to unravel the venom proteome composition of Indian snakes: Opening new avenues in clinical research. Expert Rev. Proteom. 2020, 17, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Swethakumar, B.; NaveenKumar, S.K.; Girish, K.S.; Kemparaju, K. The action of Echis carinatus and Naja naja venoms on human neutrophils; an emphasis on NETosis. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129561. [Google Scholar] [CrossRef] [PubMed]
- Kalita, B.; Mackessy, S.P.; Mukherjee, A.K. Proteomic analysis reveals geographic variation in venom composition of Russell’s Viper in the Indian subcontinent: Implications for clinical manifestations post-envenomation and antivenom treatment. Expert Rev. Proteom. 2018, 15, 837–849. [Google Scholar] [CrossRef]
- Kalita, B.; Patra, A.; Mukherjee, A.K. Unraveling the Proteome Composition and Immuno-profiling of Western India Russell’s Viper Venom for In-Depth Understanding of Its Pharmacological Properties, Clinical Manifestations, and Effective Antivenom Treatment. J. Proteome Res. 2017, 16, 583–598. [Google Scholar] [CrossRef]
- Rashmi, U.; Khochare, S.; Attarde, S.; Laxme, R.S.; Suranse, V.; Martin, G.; Sunagar, K. Remarkable intrapopulation venom variability in the monocellate cobra (Naja kaouthia) unveils neglected aspects of India’s snakebite problem. J. Proteom. 2021, 242, 104256. [Google Scholar] [CrossRef]
- Senji Laxme, R.; Khochare, S.; Attarde, S.; Suranse, V.; Iyer, A.; Casewell, N.R.; Whitaker, R.; Martin, G.; Sunagar, K. Biogeographic venom variation in Russell’s viper (Daboia russelii) and the preclinical inefficacy of antivenom therapy in snakebite hotspots. PLoS Negl. Trop. Dis. 2021, 15, e0009247. [Google Scholar] [CrossRef]
- Senji Laxme, R.R.; Attarde, S.; Khochare, S.; Suranse, V.; Martin, G.; Casewell, N.R.; Whitaker, R.; Sunagar, K. Biogeographical venom variation in the Indian spectacled cobra (Naja naja) underscores the pressing need for pan-India efficacious snakebite therapy. PLoS Negl. Trop. Dis. 2021, 15, e0009150. [Google Scholar] [CrossRef]
- Sunagar, K.; Khochare, S.; Senji Laxme, R.; Attarde, S.; Dam, P.; Suranse, V.; Khaire, A.; Martin, G.; Captain, A. A wolf in another wolf’s clothing: Post-genomic regulation dictates venom profiles of medically-important cryptic kraits in India. Toxins 2021, 13, 69. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, R.; Captain, A.; Ahmed, F. Snakes of India; Draco Books: Chengalpattu, India, 2004. [Google Scholar]
- Joseph, J.K.; Simpson, I.D.; Menon, N.C.; Jose, M.P.; Kulkarni, K.J.; Raghavendra, G.B.; Warrell, D.A. First authenticated cases of life-threatening envenoming by the hump-nosed pit viper (Hypnale hypnale) in India. Trans. R Soc. Trop. Med. Hyg. 2007, 101, 85–90. [Google Scholar] [CrossRef]
- Silva, A.; Gunawardena, P.; Weilgama, D.; Maduwage, K.; Gawarammana, I. Comparative in-vivo toxicity of venoms from South Asian hump-nosed pit vipers (Viperidae: Crotalinae: Hypnale). BMC Res. Notes 2012, 5, 471. [Google Scholar] [CrossRef]
- Ariaratnam, C.A.; Thuraisingam, V.; Kularatne, S.A.; Sheriff, M.H.; Theakston, R.D.; de Silva, A.; Warrell, D.A. Frequent and potentially fatal envenoming by hump-nosed pit vipers (Hypnale hypnale and H. nepa) in Sri Lanka: Lack of effective antivenom. Trans. R Soc. Trop. Med. Hyg. 2008, 102, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Ariaratnam, C.A.; Sheriff, M.H.; Arambepola, C.; Theakston, R.D.; Warrell, D.A. Syndromic approach to treatment of snake bite in Sri Lanka based on results of a prospective national hospital-based survey of patients envenomed by identified snakes. Am. J. Trop. Med. Hyg. 2009, 81, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Maduwage, K.; Scorgie, F.E.; Silva, A.; Shahmy, S.; Mohamed, F.; Abeysinghe, C.; Karunathilake, H.; Lincz, L.F.; Gnanathasan, C.A.; Isbister, G.K. Hump-nosed pit viper (Hypnale hypnale) envenoming causes mild coagulopathy with incomplete clotting factor consumption. Clin. Toxicol. 2013, 51, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Witharana, E.; Gnanathasan, A.; Dissanayake, A.S.; Wijesinghe, S.K.J.; Kadahetti, S.C.L.; Rajapaksha, R. Hump-nosed pit viper (Hypnale hypnale and Hypnale zara) envenoming in Deniyaya, Sri Lanka: Epidemiological and clinical features from a prospective observational study. Toxicon 2021, 189, 19–23. [Google Scholar] [CrossRef]
- Shivanthan, M.C.; Yudhishdran, J.; Navinan, R.; Rajapakse, S. Hump-nosed viper bite: An important but under-recognized cause of systemic envenoming. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 24. [Google Scholar] [CrossRef]
- Rathnayaka, R.; Ranathunga, P.; Kularatne, S.A.M. Kidney injury following envenoming by hump-nosed pit viper (Genus: Hypnale) in Sri Lanka: Proven and probable cases. Trans. R Soc. Trop. Med. Hyg. 2019, 113, 131–142. [Google Scholar] [CrossRef]
- Puthra, S.; Pirasath, S.; Hemal Sugathapala, A.G.; Gnanathasan, A. Thrombotic microangiopathy following hump-nosed viper ‘Hypnale’ envenomation. SAGE Open Med. Case Rep. 2020, 8, 2050313X20944308. [Google Scholar] [CrossRef]
- Ramachandran, S.; Ganaikabahu, B.; Pushparajan, K.; Wijesekera, J. Electroencephalographic abnormalities in patients with snake bites. Am. J. Trop. Med. Hyg. 1995, 52, 25–28. [Google Scholar] [CrossRef]
- Jeevagan, V.; Chang, T.; Gnanathasan, C.A. Acute ischemic stroke following Hump-nosed viper envenoming; first authenticated case. Thromb. J. 2012, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Thillainathan, S.; Priyangika, D.; Marasinghe, I.; Kanapathippillai, K.; Premawansa, G. Rare cardiac sequelae of a hump-nosed viper bite. BMC Res. Notes 2015, 8, 437. [Google Scholar] [CrossRef]
- Kumar, K.S.; Narayanan, S.; Udayabhaskaran, V.; Thulaseedharan, N.K. Clinical and epidemiologic profile and predictors of outcome of poisonous snake bites—An analysis of 1500 cases from a tertiary care center in Malabar, North Kerala, India. Int. J. Gen. Med. 2018, 11, 209–216. [Google Scholar] [CrossRef]
- Gowda, C.D.; Rajesh, R.; Nataraju, A.; Dhananjaya, B.L.; Raghupathi, A.R.; Gowda, T.V.; Sharath, B.K.; Vishwanath, B.S. Strong myotoxic activity of Trimeresurus malabaricus venom: Role of metalloproteases. Mol. Cell Biochem. 2006, 282, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Rathod, S.G.; Khan, A.A.; Geelani, S.A.; Roshan, R.; Bhat, J.R. Rare snake bite—A case report. Trop. Doct. 2022, 52, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Sirur, F.M.; Lath, V.; Chethana, A.S.; Mohan, G.; Shastry, S. A Tropical Kiss by a Malabar Pit Viper. Wilderness Env. Med. 2022, 33, 116–121. [Google Scholar] [CrossRef]
- Harikrishnan, M.; Rohan, K.V.; Ayyan, S.M. Trimeresurus Malabaricus Bites: Need of Species-specific Management for Snakebites. J. Emerg. Trauma Shock 2020, 13, 167. [Google Scholar] [PubMed]
- Tan, C.H.; Tan, N.H.; Sim, S.M.; Fung, S.Y.; Gnanathasan, C.A. Proteomic investigation of Sri Lankan hump-nosed pit viper (Hypnale hypnale) venom. Toxicon 2015, 93, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Maduwage, K.; Hodgson, W.C.; Konstantakopoulos, N.; O’Leary, M.A.; Gawarammana, I.; Isbister, G.K. The in vitro toxicity of venoms from South Asian hump-nosed pit vipers (Viperidae: Hypnale). J. Venom Res. 2011, 2, 17–23. [Google Scholar]
- Ali, S.A.; Baumann, K.; Jackson, T.N.; Wood, K.; Mason, S.; Undheim, E.A.; Nouwens, A.; Koludarov, I.; Hendrikx, I.; Jones, A.; et al. Proteomic comparison of Hypnale hypnale (hump-nosed pit-viper) and Calloselasma rhodostoma (Malayan pit-viper) venoms. J. Proteom. 2013, 91, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Vanuopadath, M.; Sajeev, N.; Murali, A.R.; Sudish, N.; Kangosseri, N.; Sebastian, I.R.; Jain, N.D.; Pal, A.; Raveendran, D.; Nair, B.G.; et al. Mass spectrometry-assisted venom profiling of Hypnale hypnale found in the Western Ghats of India incorporating de novo sequencing approaches. Int. J. Biol. Macromol. 2018, 118, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Vanuopadath, M.; Shaji, S.K.; Raveendran, D.; Nair, B.G.; Nair, S.S. Delineating the venom toxin arsenal of Malabar pit viper (Trimeresurus malabaricus) from the Western Ghats of India and evaluating its immunological cross-reactivity and in vitro cytotoxicity. Int. J. Biol. Macromol. 2020, 148, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.N.W.; Jouanne, H.; Vidal, N. Snake Venom in Context: Neglected Clades and Concepts. Front. Ecol. Evol. 2019, 7, 332. [Google Scholar] [CrossRef]
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharm. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- de Silva, A.; Wijekoon, A.S.; Jayasena, L.; Abeysekera, C.K.; Bao, C.X.; Hutton, R.A.; Warrell, D.A. Haemostatic dysfunction and acute renal failure following envenoming by Merrem’s hump-nosed viper (Hypnale hypnale) in Sri Lanka: First authenticated case. Trans. R Soc. Trop. Med. Hyg. 1994, 88, 209–212. [Google Scholar] [CrossRef]
- Mahale, R.; Mehta, A.; Javali, M.; Srinivasa, R. A case of bilateral occipital lobe infarcts following Indian tree viper bite. J. Stroke 2014, 16, 205–207. [Google Scholar] [CrossRef]
- Attarde, S.; Iyer, A.; Khochare, S.; Shaligram, U.; Vikharankar, M.; Sunagar, K. The Preclinical Evaluation of a Second-Generation Antivenom for Treating Snake Envenoming in India. Toxins 2022, 14, 168. [Google Scholar] [CrossRef]
- Tan, C.H.; Leong, P.K.; Fung, S.Y.; Sim, S.M.; Ponnudurai, G.; Ariaratnam, C.; Khomvilai, S.; Sitprija, V.; Tan, N.H. Cross neutralization of Hypnale hypnale (hump-nosed pit viper) venom by polyvalent and monovalent Malayan pit viper antivenoms in vitro and in a rodent model. Acta Trop. 2011, 117, 119–124. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, N.H.; Sim, S.M.; Fung, S.Y.; Gnanathasan, C.A. Immunological properties of Hypnale hypnale (hump-nosed pit viper) venom: Antibody production with diagnostic and therapeutic potentials. Acta Trop. 2012, 122, 267–275. [Google Scholar] [CrossRef]
- Vikrant, S.; Jaryal, A.; Parashar, A. Clinicopathological spectrum of snake bite-induced acute kidney injury from India. World J. Nephrol. 2017, 6, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Sinha, R.; Chaudhury, A.R.; Maduwage, K.; Abeyagunawardena, A.; Bose, N.; Pradhan, S.; Bresolin, N.L.; Garcia, B.A.; McCulloch, M. Snake bite associated with acute kidney injury. Pediatr. Nephrol. 2021, 36, 3829–3840. [Google Scholar] [CrossRef] [PubMed]
- Harshavardhan, L.; Lokesh, A.J.; Tejeshwari, H.L.; Halesha, B.R.; Metri, S.S. A study on the acute kidney injury in snake bite victims in a tertiary care centre. J. Clin. Diagn. Res. 2013, 7, 853–856. [Google Scholar]
- Bickler, P.E. Amplification of Snake Venom Toxicity by Endogenous Signaling Pathways. Toxins 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Sitprija, V. Snakebite nephropathy. Nephrology 2006, 11, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Iyer, A.; Sunagar, K. Evolution Bites—Timeworn Inefficacious Snakebite Therapy in the Era of Recombinant Vaccines. Indian Pediatr. 2021, 58, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Smith, B.J. SDS Polyacrylamide Gel Electrophoresis of Proteins. In Proteins; Methods in Molecular Biology™; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 1984; Volume 1, pp. 41–55. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Senji Laxme, R.R.; Khochare, S.; Attarde, S.; Kaur, N.; Jaikumar, P.; Shaikh, N.Y.; Aharoni, R.; Primor, N.; Hawlena, D.; Moran, Y.; et al. The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii. Toxins 2022, 14, 725. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Freitas-de-Sousa, L.A.; Nachtigall, P.G.; Portes-Junior, J.A.; Holding, M.L.; Nystrom, G.S.; Ellsworth, S.A.; Guimarães, N.C.; Tioyama, E.; Ortiz, F.; Silva, B.R. Size matters: An evaluation of the molecular basis of ontogenetic modifications in the composition of Bothrops jararacussu snake venom. Toxins 2020, 12, 791. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Lee, M.S.; Ziajko, M.; Dunstan, N.; Sumner, J.; Isbister, G.K. Activity of two key toxin groups in Australian elapid venoms show a strong correlation to phylogeny but not to diet. BMC Evol. Biol. 2020, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, M.; Takahashi, T. A spectrophotometric microplate assay for L-amino acid oxidase. Anal. Biochem. 2001, 298, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.; Miyoshi, S.; Shinoda, S. Purification and characterization of a protease produced by Vibrio mimicus. Infect. Immun. 1990, 58, 4159–4162. [Google Scholar] [CrossRef]
- Teng, C.-M.; Ouyang, C.; Lin, S.-C. Species difference in the fibrinogenolytic effects of α-and β-fibrinogenases from Trimeresurus mucrosquamatus snake venom. Toxicon 1985, 23, 777–782. [Google Scholar] [CrossRef]
- op den Brouw, B.; Ghezellou, P.; Casewell, N.R.; Ali, S.A.; Fathinia, B.; Fry, B.G.; Bos, M.H.; Ikonomopoulou, M.P. Pharmacological characterisation of Pseudocerastes and Eristicophis viper venoms reveal anticancer (Melanoma) properties and a potentially novel mode of fibrinogenolysis. Int. J. Mol. Sci. 2021, 22, 6896. [Google Scholar] [CrossRef]
- Casewell, N.R.; Cook, D.A.; Wagstaff, S.C.; Nasidi, A.; Durfa, N.; Wuster, W.; Harrison, R.A. Pre-clinical assays predict pan-African Echis viper efficacy for a species-specific antivenom. PLoS Negl. Trop. Dis. 2010, 4, e851. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: London, UK, 1971. [Google Scholar]
- Kondo, H.; Kondo, S.; Ikezawa, H.; Murata, R. Studies on the quantitative method for determination of hemorrhagic activity of Habu snake venom. Jpn. J. Med. Sci. Biol. 1960, 13, 43–52. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Solano, G.; Pla, D.; Herrera, M.; Segura, Á.; Vargas, M.; Villalta, M.; Sánchez, A.; Sanz, L.; Lomonte, B. Preclinical evaluation of the efficacy of antivenoms for snakebite envenoming: State-of-the-art and challenges ahead. Toxins 2017, 9, 163. [Google Scholar] [CrossRef]
- WHO. World Health Organisation Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; World Health Organisation: Geneva, Switzerland, 2018. [Google Scholar]
- Raij, L.; Azar, S.; Keane, W. Mesangial immune injury, hypertension, and progressive glomerular damage in Dahl rats. Kidney Int. 1984, 26, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Frohlich, E.D.; Ono, H.; Ono, Y.; Numabe, A.; Willis, G.W. Glomerular dynamics and morphology of aged spontaneously hypertensive rats. Effects of angiotensin-converting enzyme inhibition. Hypertension 1995, 25, 207–213. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. malabaricus | C. gramineus | H. hypnale | ||
|---|---|---|---|---|
| LD50 mg/kg | >6 | 4–6 | 1.32 | |
| MHD (μg/mouse) | 0.365 | NA | 2.32 | |
| MND (μg/mouse) | NA | NA | 22.3 | |
| ED50 (mL) | PSVPL-IN | NA | NA | — |
| PSVPL-SL | NA | NA | — | |
| MHD50 (mL) | PSVPL-IN | 3.6 × 10−4 | NA | — |
| PSVPL-SL | 3.4 × 10−4 | NA | — | |
| MND50 (mL) | PSVPL-IN | NA | NA | — |
| PSVPL-SL | NA | NA | — | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khochare, S.; Senji Laxme, R.R.; Jaikumar, P.; Kaur, N.; Attarde, S.; Martin, G.; Sunagar, K. Fangs in the Ghats: Preclinical Insights into the Medical Importance of Pit Vipers from the Western Ghats. Int. J. Mol. Sci. 2023, 24, 9516. https://doi.org/10.3390/ijms24119516
Khochare S, Senji Laxme RR, Jaikumar P, Kaur N, Attarde S, Martin G, Sunagar K. Fangs in the Ghats: Preclinical Insights into the Medical Importance of Pit Vipers from the Western Ghats. International Journal of Molecular Sciences. 2023; 24(11):9516. https://doi.org/10.3390/ijms24119516
Chicago/Turabian StyleKhochare, Suyog, R. R. Senji Laxme, Priyanka Jaikumar, Navneet Kaur, Saurabh Attarde, Gerard Martin, and Kartik Sunagar. 2023. "Fangs in the Ghats: Preclinical Insights into the Medical Importance of Pit Vipers from the Western Ghats" International Journal of Molecular Sciences 24, no. 11: 9516. https://doi.org/10.3390/ijms24119516
APA StyleKhochare, S., Senji Laxme, R. R., Jaikumar, P., Kaur, N., Attarde, S., Martin, G., & Sunagar, K. (2023). Fangs in the Ghats: Preclinical Insights into the Medical Importance of Pit Vipers from the Western Ghats. International Journal of Molecular Sciences, 24(11), 9516. https://doi.org/10.3390/ijms24119516