

Possible Molecular Mechanisms Underlying the Decrease in the Antibacterial Activity of Protamine-like Proteins after Exposure of Mytilus galloprovincialis to Chromium and Mercury
 ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
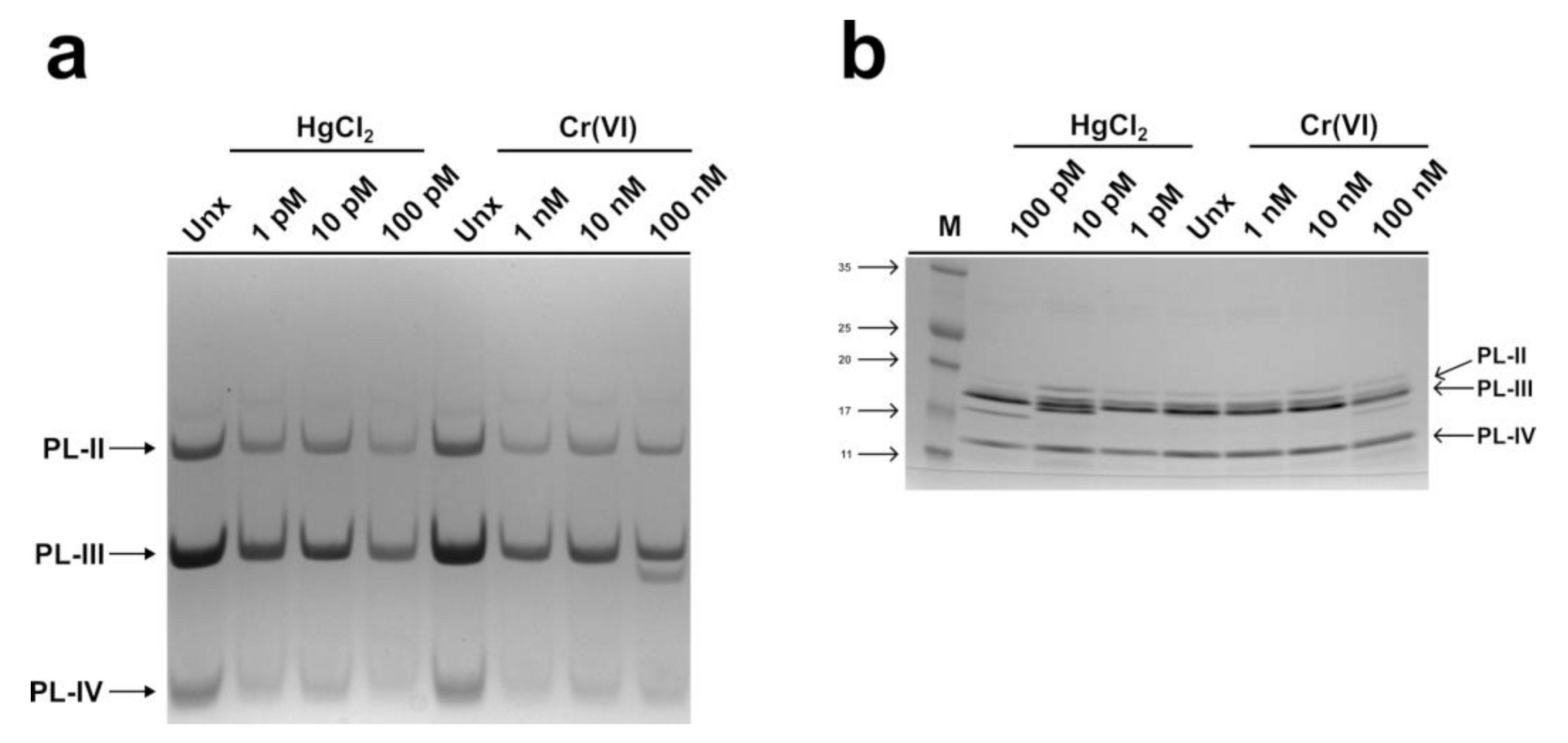
2.1. Electrophoretic Analysis of the PLs of M. galloprovincialis
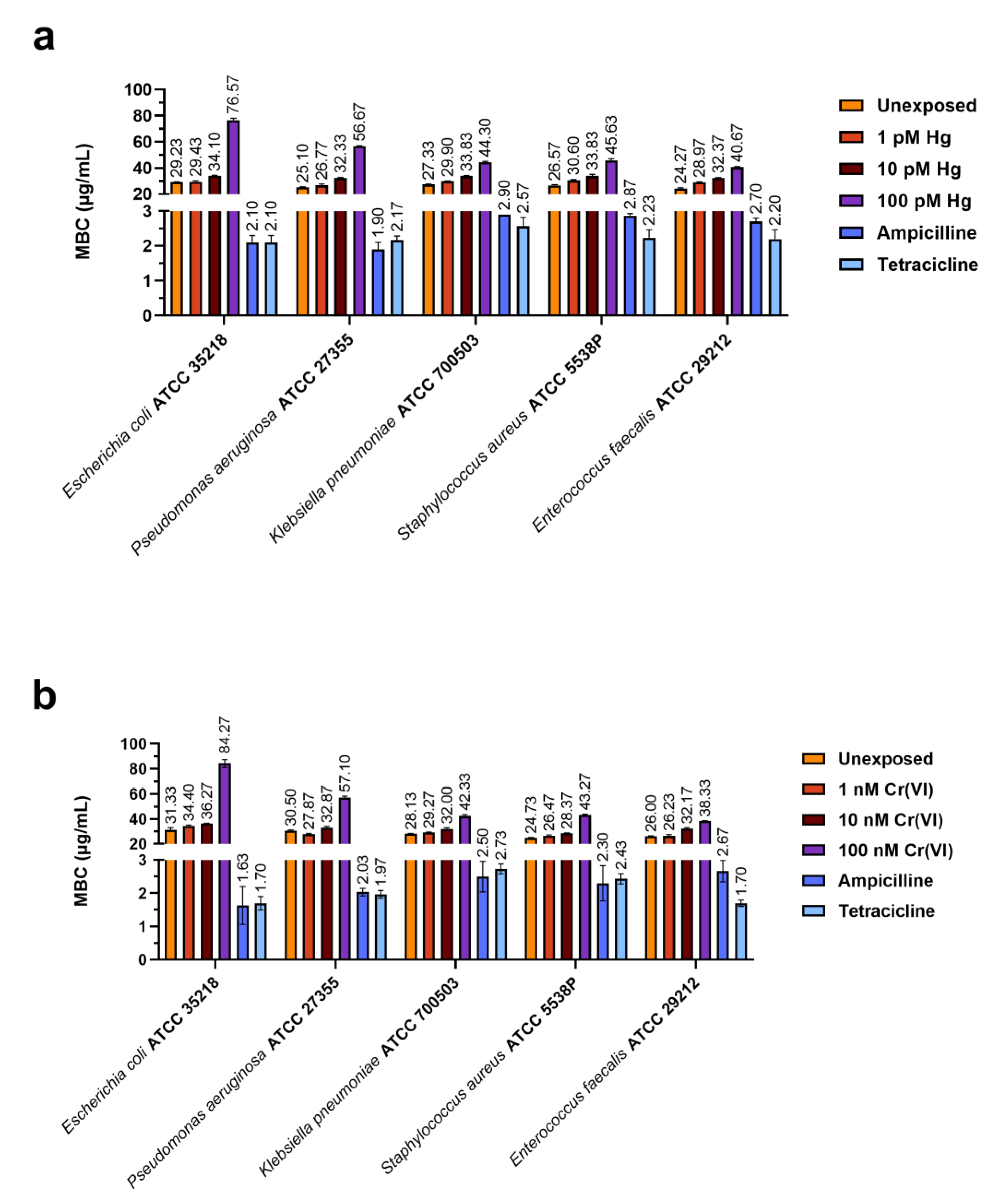
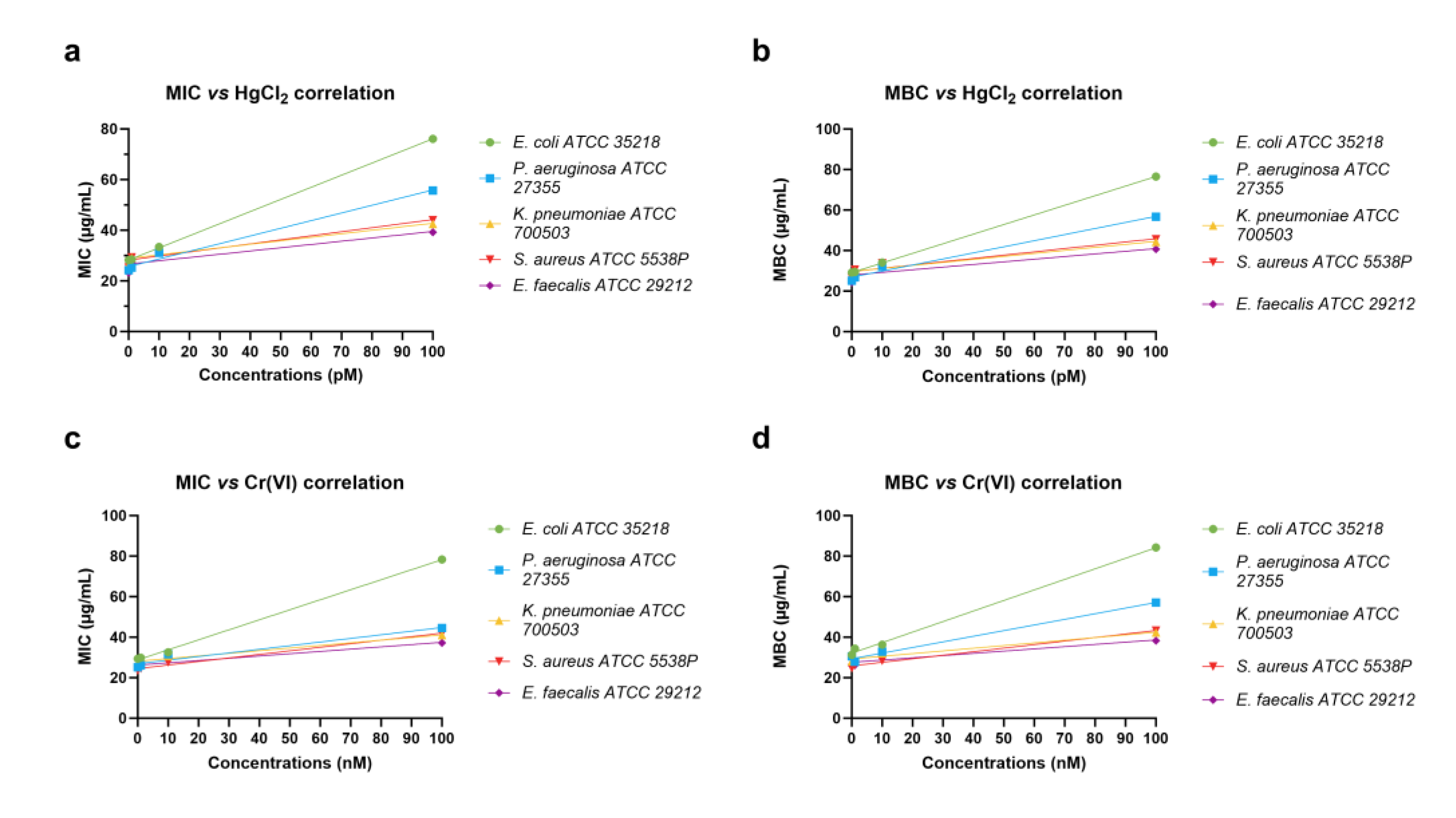
2.2. Determination of MIC and MBC of the PLs of M. galloprovincialis
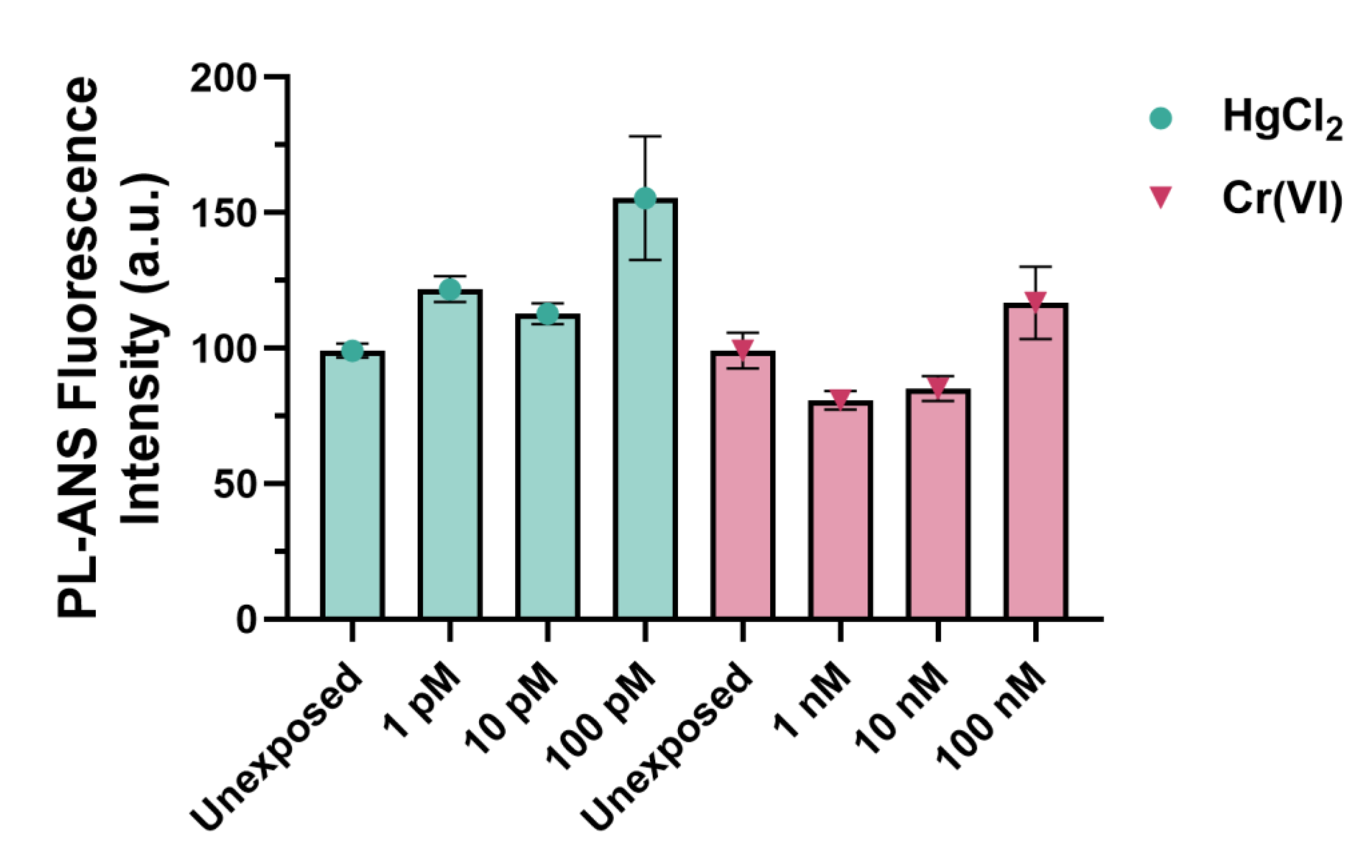
2.3. Fluorescence Analyses of PLs
2.4. Thermal Stability of PLs
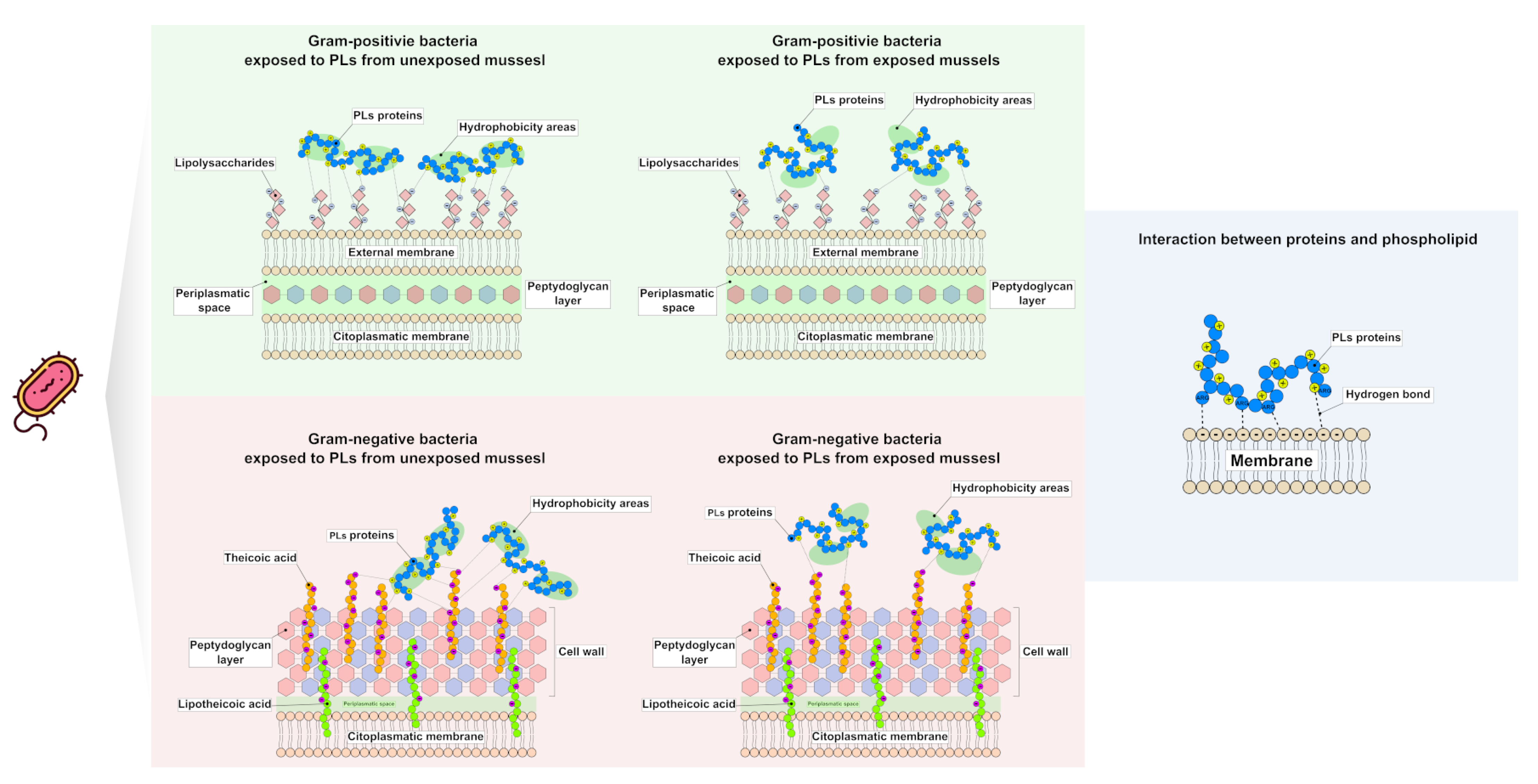
3. Discussion
4. Materials and Methods
4.1. The Ethics Statement
4.2. Materials and Animal Collection
4.3. Treatment of Mussels and Spermatozoa Collection
4.4. Extraction and Purification of M. galloprovincialis PLs
4.5. Electrophoretic Analyses on Polyacryilamide Gels of PLs from Unexposed and Exposed M. galloprovincialis
4.6. Thermal Stability of PLs
4.7. Antibacterial Activity of M. galloprovincialis PLs
4.8. Fluorescence Measurements of M. galloprovincialis PLs
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- AL-Eitan, L.; Sendyani, S.; Alnemri, M. Applications of the One Health Concept: Current Status in the Middle East. J. Biosaf. Biosecur. 2023, 5, 21–31. [Google Scholar] [CrossRef]
- Rossi, F.; Péguilhan, R.; Turgeon, N.; Veillette, M.; Baray, J.-L.; Deguillaume, L.; Amato, P.; Duchaine, C. Quantification of Antibiotic Resistance Genes (ARGs) in Clouds at a Mountain Site (Puy de Dôme, Central France). Sci. Total Environ. 2023, 865, 161264. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Gao, M.; Lv, L.; Wang, J.; Cai, Z.; Bai, Y.; Gao, X.; Gao, G.; Pu, W.; Jiao, Y.; et al. Persistence and Molecular Epidemiology of BlaNDM-Positive Gram-Negative Bacteria in Three Broiler Farms: A Longitudinal Study (2015–2021). J. Hazard. Mater. 2023, 446, 130725. [Google Scholar] [CrossRef]
- Ramesan, C.K.V.; Vinod, N.V.; Skariyachan, S. 14-Antimicrobial Peptide Resistance and Scope of Computational Biology in Antimicrobial Peptide Research. In Antimicrobial Peptides; Ajesh, K., Sreejith, K., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 261–295. ISBN 978-0-323-85682-9. [Google Scholar]
- Pane, K.; Verrillo, M.; Avitabile, A.; Pizzo, E.; Varcamonti, M.; Zanfardino, A.; Di Maro, A.; Rega, C.; Amoresano, A.; Izzo, V.; et al. Chemical Cleavage of an Asp-Cys Sequence Allows Efficient Production of Recombinant Peptides with an N-Terminal Cysteine Residue. Bioconjug. Chem. 2018, 29, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, M.V.; Lekshmi, S.; Castillo-Godina, R.; Campos-Muzquiz, L.G.; Rodriguez-Herrera, R.; Biju, S.; Krishnan, R.; Swapna, T.S. Bioresources and Diversity. In Conservation and Sustainable Utilization of Bioresources; Sukumaran, S.T., Keerthi, T.R., Eds.; Sustainable Development and Biodiversity; Springer Nature: Singapore, 2023; pp. 1–37. ISBN 978-981-19584-1-0. [Google Scholar]
- Osorio-Reyes, J.G.; Valenzuela-Amaro, H.M.; Pizaña-Aranda, J.J.P.; Ramírez-Gamboa, D.; Meléndez-Sánchez, E.R.; López-Arellanes, M.E.; Castañeda-Antonio, M.D.; Coronado-Apodaca, K.G.; Gomes Araújo, R.; Sosa-Hernández, J.E.; et al. Microalgae-Based Biotechnology as Alternative Biofertilizers for Soil Enhancement and Carbon Footprint Reduction: Advantages and Implications. Mar. Drugs 2023, 21, 93. [Google Scholar] [CrossRef]
- Gram, L.; Melchiorsen, J.; Bruhn, J.B. Antibacterial Activity of Marine Culturable Bacteria Collected from a Global Sampling of Ocean Surface Waters and Surface Swabs of Marine Organisms. Mar. Biotechnol. 2010, 12, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lu, Y.; Sun, B.; Zhang, M.; Wang, C.; Xiu, C.; Johnson, A.C.; Wang, P. Ecological and Human Health Risks of Antibiotics in Marine Species through Mass Transfer from Sea to Land in a Coastal Area: A Case Study in Qinzhou Bay, the South China Sea. Environ. Pollut. 2023, 316, 120502. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, C.; Frenzilli, G.; Lasagna, C.; Perrone, E.; Roggieri, P. Genotoxicity Biomarkers in Mytilus galloprovincialis: Wild versus Caged Mussels. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2004, 552, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.; Garcia, A.R.; Pereira, B.P.; Fonseca, M.; Mestre, N.C.; Fonseca, T.G.; Ilharco, L.M.; Bebianno, M.J. Microplastics Effects in Scrobicularia Plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef]
- Provenza, F.; Rampih, D.; Pignattelli, S.; Pastorino, P.; Barceló, D.; Prearo, M.; Specchiulli, A.; Renzi, M. Mussel Watch Program for Microplastics in the Mediterranean Sea: Identification of Biomarkers of Exposure Using Mytilus galloprovincialis. Ecol. Indic. 2022, 142, 109212. [Google Scholar] [CrossRef]
- Piscopo, M.; Trifuoggi, M.; Notariale, R.; Labar, S.; Troisi, J.; Giarra, A.; Rabbito, D.; Puoti, R.; de Benedictis, D.; Brundo, M.V.; et al. Protamine-like Proteins’ Analysis as an Emerging Biotechnique for Cadmium Impact Assessment on Male Mollusk Mytilus galloprovincialis (Lamarck 1819). Acta Biochim. Pol. 2018, 65, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Notariale, R.; Basile, A.; Montana, E.; Romano, N.C.; Cacciapuoti, M.G.; Aliberti, F.; Gesuele, R.; Ruberto, F.D.; Sorbo, S.; Tenore, G.C.; et al. Protamine-like Proteins Have Bactericidal Activity. The First Evidence in Mytilus galloprovincialis. Acta Biochim. Pol. 2018, 65, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Notariale, R.; Carusone, N.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. New Insights into Alterations in PL Proteins Affecting Their Binding to DNA after Exposure of Mytilus galloprovincialis to Mercury—A Possible Risk to Sperm Chromatin Structure? Int. J. Mol. Sci. 2021, 22, 5893. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef]
- Carbone, G.; Lettieri, G.; Marinaro, C.; Costabile, M.; Notariale, R.; Bianchi, A.R.; De Maio, A.; Piscopo, M. A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus galloprovincialis: An In Vitro Study. Biomolecules 2023, 13, 520. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef]
- Moriello, C.; Costabile, M.; Spinelli, M.; Amoresano, A.; Palumbo, G.; Febbraio, F.; Piscopo, M. Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis? Biomolecules 2022, 12, 700. [Google Scholar] [CrossRef]
- Witkowska, D.; Słowik, J.; Chilicka, K. Heavy Metals and Human Health: Possible Exposure Pathways and the Competition for Protein Binding Sites. Molecules 2021, 26, 6060. [Google Scholar] [CrossRef]
- Mizrahi, L.; Achituv, Y. Effect of Heavy Metals Ions on Enzyme Activity in the Mediterranean Mussel, Donax Trunculus. Bull. Environ. Contam. Toxicol. 1989, 42, 854–859. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Moore, B.S. Lessons from the Past and Charting the Future of Marine Natural Products Drug Discovery and Chemical Biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.-Y.; Li, H.-J.; Li, Q.-Y.; Wu, Y.-C. Application of Marine Natural Products in Drug Research. Bioorg. Med. Chem. 2021, 35, 116058. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy Metal Pollution in the Environment and Their Toxicological Effects on Humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Khan, A.H.; Sharholy, M.; Alam, P.; Al-Mansour, A.I.; Ahmad, K.; Kamal, M.A.; Alam, S.; Pervez, M.N.; Naddeo, V. Evaluation of Cost Benefit Analysis of Municipal Solid Waste Management Systems. J. King Saud Univ.—Sci. 2022, 34, 101997. [Google Scholar] [CrossRef]
- Piscopo, M.; Notariale, R.; Rabbito, D.; Ausió, J.; Olanrewaju, O.S.; Guerriero, G. Mytilus galloprovincialis (Lamarck, 1819) Spermatozoa: Hsp70 Expression and Protamine-like Protein Property Studies. Environ. Sci. Pollut. Res. 2018, 25, 12957–12966. [Google Scholar] [CrossRef] [PubMed]
- Anedda, E.; Farrell, M.L.; Morris, D.; Burgess, C.M. Evaluating the Impact of Heavy Metals on Antimicrobial Resistance in the Primary Food Production Environment: A Scoping Review. Environ. Pollut. 2023, 320, 121035. [Google Scholar] [CrossRef]
- Maphuhla, N.G.; Lewu, F.B.; Oyedeji, O.O. Enzyme Activities in Reduction of Heavy Metal Pollution from Alice Landfill Site in Eastern Cape, South Africa. Int. J. Environ. Res. Public Health 2022, 19, 12054. [Google Scholar] [CrossRef]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and Arginine-Rich Antimicrobial Peptides: Structures and Mechanisms of Action. Biochim. Biophys. Acta (BBA)—Biomembr. 2006, 1758, 1184–1202. [Google Scholar] [CrossRef]
- Shai, Y.; Oren, Z. From “Carpet” Mechanism to de-Novo Designed Diastereomeric Cell-Selective Antimicrobial Peptides. Peptides 2001, 22, 1629–1641. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.; Lee, S.; Kim, H.; Sim, J.-Y.; Pak, B.; Kim, K.; Il Kim, J. Matching Amino Acids Membrane Preference Profile to Improve Activity of Antimicrobial Peptides. Commun. Biol. 2022, 5, 1199. [Google Scholar] [CrossRef]
- Bacalum, M.; Janosi, L.; Zorila, F.; Tepes, A.-M.; Ionescu, C.; Bogdan, E.; Hadade, N.; Craciun, L.; Grosu, I.; Turcu, I.; et al. Modulating Short Tryptophan- and Arginine-Rich Peptides Activity by Substitution with Histidine. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2017, 1861, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Ding, Y.; Yao, K.; Gao, W.; Wang, Y. Antimicrobial Resistance Analysis of Clinical Escherichia Coli Isolates in Neonatal Ward. Front. Pediatr. 2021, 9, 670470. [Google Scholar] [CrossRef]
- Pérez, L.; Pinazo, A.; Morán, M.C.; Pons, R. Aggregation Behavior, Antibacterial Activity and Biocompatibility of Catanionic Assemblies Based on Amino Acid-Derived Surfactants. Int. J. Mol. Sci. 2020, 21, 8912. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Maniti, O.; Piao, H.-R.; Ayala-Sanmartin, J. Basic Cell Penetrating Peptides Induce Plasma Membrane Positive Curvature, Lipid Domain Separation and Protein Redistribution. Int. J. Biochem. Cell Biol. 2014, 50, 73–81. [Google Scholar] [CrossRef]
- Hawe, A.; Sutter, M.; Jiskoot, W. Extrinsic Fluorescent Dyes as Tools for Protein Characterization. Pharm. Res. 2008, 25, 1487–1499. [Google Scholar] [CrossRef]
- Bismuto, E.; Irace, G.; Sirangelo, I.; Gratton, E. Pressure-Induced Perturbation of ANS-Apomyoglobin Complex: Frequency Domain Fluorescence Studies on Native and Acidic Compact States. Protein. Sci. 1996, 5, 121–126. [Google Scholar] [CrossRef]
- Singh, K.; Hussain, I.; Mishra, V.; Akhtar, M.S. New Insight on 8-Anilino-1-Naphthalene Sulfonic Acid Interaction with TgFNR for Hydrophobic Exposure Analysis. Int. J. Biol. Macromol. 2019, 122, 636–643. [Google Scholar] [CrossRef]
- Gorinstein, S.; Jung, S.-T.; Moncheva, S.; Arancibia-Avila, P.; Park, Y.-S.; Kang, S.-G.; Goshev, I.; Trakhtenberg, S.; Namiesnik, J. Partial Characterization of Proteins from Mussel Mytilus galloprovincialis as a Biomarker of Contamination. Arch. Environ. Contam. Toxicol. 2005, 49, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of Peptide Hydrophobicity in the Mechanism of Action of Alpha-Helical Antimicrobial Peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Vassalli, Q.A.; Caccavale, F.; Avagnano, S.; Murolo, A.; Guerriero, G.; Fucci, L.; Ausió, J.; Piscopo, M. New Insights into Protamine-Like Component Organization in Mytilus galloprovincialis’ Sperm Chromatin. DNA Cell Biol. 2015, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Notariale, R.; Tortora, F.; Lettieri, G.; Palumbo, G.; Manna, C. Novel Insights into Mercury Effects on Hemoglobin and Membrane Proteins in Human Erythrocytes. Molecules 2020, 25, 3278. [Google Scholar] [CrossRef]
- Uschewa, A.; Patriotis, C.; Avramova, Z. An H1-like Protein from the Sperm Chromatin of Mytilus galloprovincialis. Cell Biol. Int. Rep. 1985, 9, 253–263. [Google Scholar] [CrossRef]
- Ausió, J.; Subirana, J.A. Nuclear Proteins and the Organization of Chromatin in Spermatozoa of Mythus Edulis. Exp. Cell Res. 1982, 141, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ausio, J.; McPARLAND, R. Sequence and Characterization of the Sperm-Specific Protein Φ3 from Mytilus Californianus. Eur. J. Biochem. 1989, 182, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, C.; Carlos, S.; Ausio, J. Microheterogeneity and Interspecific Variability of the Nuclear Sperm Proteins from Mytilus. FEBS Lett. 1991, 282, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Rocchini, C.; Rice, P.; Ausio, J. Complete Sequence and Characterization of the Major Sperm Nuclear Basic Protein from Mytilus Trossulus. FEBS Lett. 1995, 363, 37–40. [Google Scholar] [CrossRef]
- Eirín-López, J.M.; Lewis, J.D.; Howe, L.A.; Ausió, J. Common Phylogenetic Origin of Protamine-like (PL) Proteins and Histone H1: Evidence from Bivalve PL Genes. Mol. Biol. Evol. 2006, 23, 1304–1317. [Google Scholar] [CrossRef]
- Ausió, J.; Subirana, J.A. A High Molecular Weight Nuclear Basic Protein from the Bivalve Mollusc Spisula Solidissima. J. Biol. Chem. 1982, 257, 2802–2805. [Google Scholar] [CrossRef]
- Sheikh, C.; Houssain, M.S.; Easmin, M.S.; Islam, M.S.; Houssain, M.A.; Rashid, M. New Coordination Complexes of Chromium as Cytotoxic and Antimicrobial Agents. Pak. J. Biol. Sci. 2004, 7, 335–339. [Google Scholar] [CrossRef]
- Verrillo, M.; Koellensperger, G.; Puehringer, M.; Cozzolino, V.; Spaccini, R.; Rampler, E. Evaluation of Sustainable Recycled Products to Increase the Production of Nutraceutical and Antibacterial Molecules in Basil Plants by a Combined Metabolomic Approach. Plants 2023, 12, 513. [Google Scholar] [CrossRef]
- Lettieri, G.; Maione, M.; Ranauda, M.A.; Mele, E.; Piscopo, M. Molecular Effects on Spermatozoa of Mytilus galloprovincialis Exposed to Hyposaline Conditions. Mol. Reprod. Dev. 2019, 86, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Verrillo, M.; Salzano, M.; Savy, D.; Di Meo, V.; Valentini, M.; Cozzolino, V.; Piccolo, A. Antibacterial and Antioxidant Properties of Humic Substances from Composted Agricultural Biomasses. Chem. Biol. Technol. Agric. 2022, 9, 28. [Google Scholar] [CrossRef]
- Verrillo, M.; Cozzolino, V.; Spaccini, R.; Piccolo, A. Humic Substances from Green Compost Increase Bioactivity and Antibacterial Properties of Essential Oils in Basil Leaves. Chem. Biol. Technol. Agric. 2021, 8, 28. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E. coli ATCC 35218 | P. aeruginosa ATCC 27355 | K. pneumoniae ATCC 700503 | S. aureus ATCC 5538P | E. faecalis ATCC 29212 | |
|---|---|---|---|---|---|
| r | 0.9999 | 0.9934 | 0.9476 | 0.9617 | 0.9116 |
| 95% confidence interval | 0.9971 to 1.000 | 0.7140 to 0.9999 | −0.1508 to 0.9989 | 0.007551 to 0.9992 | −0.3993 to 0.9982 |
| R-squared | 0.9999 | 0.9868 | 0.8980 | 0.9248 | 0.8311 |
| p (two-tailed) | <0.0001 | 0.0066 | 0.0524 | 0.0383 | 0.0884 |
| p value summary | **** | ** | ns | * | ns |
| Significant? (alpha = 0.05) | Yes | Yes | No | Yes | No |
| E. coli ATCC 35218 | P. aeruginosa ATCC 27355 | K. pneumoniae ATCC 700503 | S. aureus ATCC 5538P | E. faecalis ATCC 29212 | |
|---|---|---|---|---|---|
| r | 1.000 | 0.9927 | 0.9614 | 0.9577 | 0.9120 |
| 95% confidence interval | 0.9987 to 1.000 | 0.6866 to 0.9999 | 0.003564 to 0.9992 | −0.04278 to 0.9991 | −0.3975 to 0.9982 |
| R-squared | 0.9999 | 0.9854 | 0.9242 | 0.9172 | 0.8317 |
| p (two-tailed) | <0.0001 | 0.0073 | 0.0386 | 0.0423 | 0.0880 |
| p value summary | **** | ** | * | * | ns |
| Significant? (alpha = 0.05) | Yes | Yes | Yes | Yes | No |
| E. coli ATCC 35218 | P. aeruginosa ATCC 27355 | K. pneumoniae ATCC 700503 | S. aureus ATCC 5538P | E. faecalis ATCC 29212 | |
|---|---|---|---|---|---|
| r | 0.9993 | 0.9735 | 0.9888 | 0.9934 | 0.9364 |
| 95% confidence interval | 0.9669 to 1.000 | 0.1931 to 0.9995 | 0.5578 to 0.9998 | 0.7149 to 0.9999 | −0.2466 to 0.9987 |
| R-squared | 0.9987 | 0.9477 | 0.9777 | 0.9869 | 0.8769 |
| p (two-tailed) | 0.0007 | 0.0265 | 0.0112 | 0.0066 | 0.0636 |
| p value summary | *** | * | * | ** | ns |
| Significant? (alpha = 0.05) | Yes | Yes | Yes | Yes | No |
| E. coli ATCC 35218 | P. aeruginosa ATCC 27355 | K. pneumoniae ATCC 700503 | S. aureus ATCC 5538P | E. faecalis ATCC 29212 | |
|---|---|---|---|---|---|
| r | 0.9987 | 0.9954 | 0.9867 | 0.9953 | 0.9142 |
| 95% confidence interval | 0.9379 to 1.000 | 0.7937 to 0.9999 | 0.4959 to 0.9997 | 0.7874 to 0.9999 | −0.3863 to 0.9982 |
| R-squared | 0.9975 | 0.9909 | 0.9736 | 0.9906 | 0.8358 |
| p (two-tailed) | 0.0013 | 0.0046 | 0.0133 | 0.0047 | 0.0858 |
| p value summary | ** | ** | * | ** | ns |
| Significant? (alpha = 0.05) | Yes | Yes | Yes | Yes | No |
| Bacteria Strains | Gram Stain |
|---|---|
| Escherichia coli ATCC 35218 | Gram− |
| Pseudomonas aeruginosa ATCC 27355 | Gram− |
| Klebsiella pneumoniae ATCC 700503 | Gram− |
| Staphylococcus aureus ATCC 5538P | Gram+ |
| Enterococcus faecalis ATCC 29212 | Gram+ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marinaro, C.; Lettieri, G.; Verrillo, M.; Morelli, M.; Carraturo, F.; Guida, M.; Piscopo, M. Possible Molecular Mechanisms Underlying the Decrease in the Antibacterial Activity of Protamine-like Proteins after Exposure of Mytilus galloprovincialis to Chromium and Mercury. Int. J. Mol. Sci. 2023, 24, 9345. https://doi.org/10.3390/ijms24119345
Marinaro C, Lettieri G, Verrillo M, Morelli M, Carraturo F, Guida M, Piscopo M. Possible Molecular Mechanisms Underlying the Decrease in the Antibacterial Activity of Protamine-like Proteins after Exposure of Mytilus galloprovincialis to Chromium and Mercury. International Journal of Molecular Sciences. 2023; 24(11):9345. https://doi.org/10.3390/ijms24119345
Chicago/Turabian StyleMarinaro, Carmela, Gennaro Lettieri, Mariavittoria Verrillo, Michela Morelli, Federica Carraturo, Marco Guida, and Marina Piscopo. 2023. "Possible Molecular Mechanisms Underlying the Decrease in the Antibacterial Activity of Protamine-like Proteins after Exposure of Mytilus galloprovincialis to Chromium and Mercury" International Journal of Molecular Sciences 24, no. 11: 9345. https://doi.org/10.3390/ijms24119345
APA StyleMarinaro, C., Lettieri, G., Verrillo, M., Morelli, M., Carraturo, F., Guida, M., & Piscopo, M. (2023). Possible Molecular Mechanisms Underlying the Decrease in the Antibacterial Activity of Protamine-like Proteins after Exposure of Mytilus galloprovincialis to Chromium and Mercury. International Journal of Molecular Sciences, 24(11), 9345. https://doi.org/10.3390/ijms24119345