
Antifungal Activities of Fluorinated Pyrazole Aldehydes on Phytopathogenic Fungi, and Their Effect on Entomopathogenic Nematodes, and Soil-Beneficial Bacteria

 , , , , ,
, , , , ,
Abstract
1. Introduction
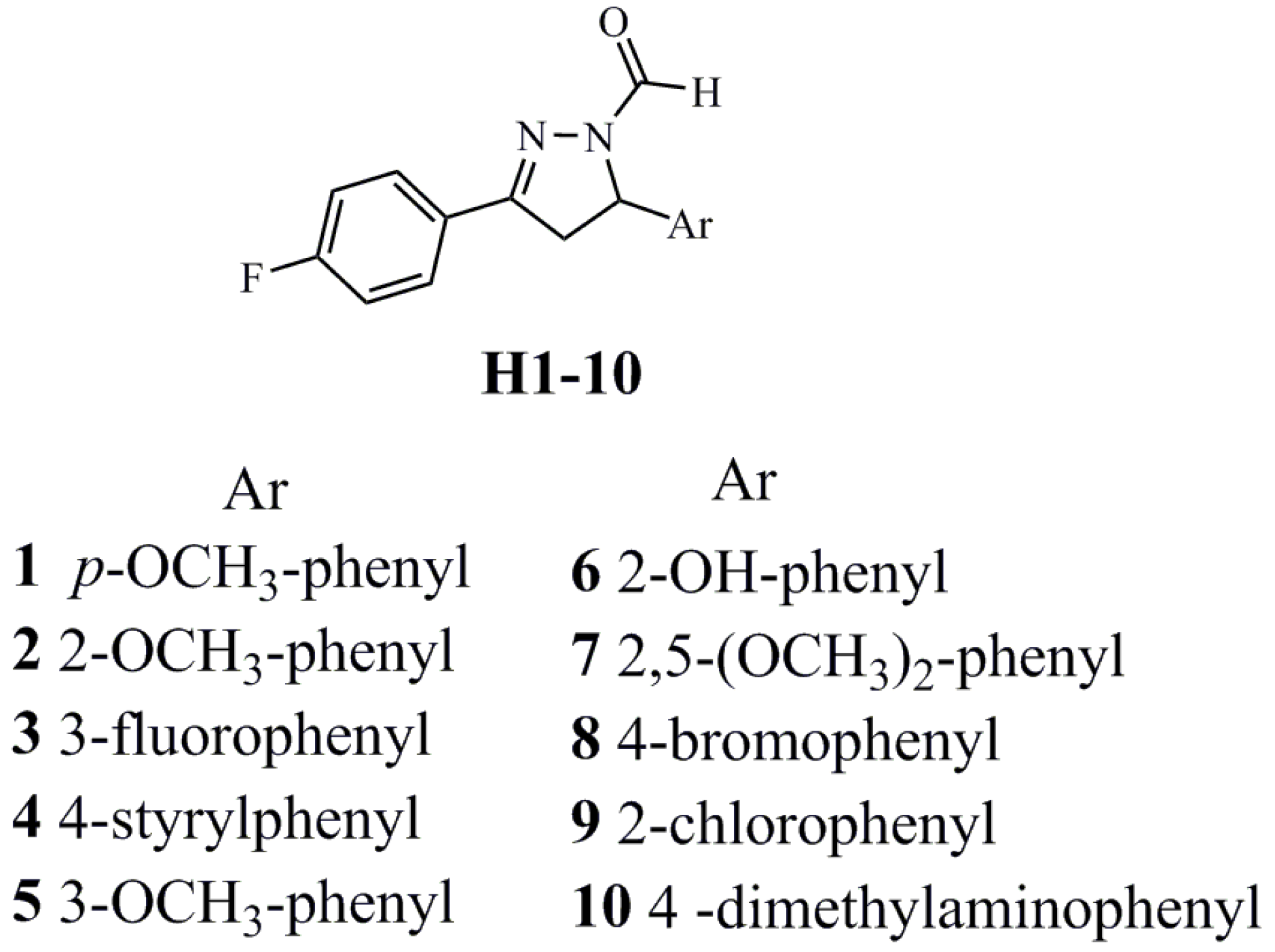
2. Results
2.1. Biological Activity Evaluation
2.1.1. Antifungal Activity
2.1.2. Antibacterial Activity
2.1.3. Nematicidal Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code of Molecule 1 | Antifungal Activity a,2 | Antibacterial Activity b | Nematicidal Activity c | Inhibition of AChE/% d,3 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Macrophomina phaseolina | Sclerotinia sclerotiorum | Fusarium oxysporum f. sp. lycopersici | Fusarium culmorum | Bacillus mycoides | Bradyrhizobium japonicum | Heterorhabditis bacteriophora | Steinernema feltiae | ||
| H1 | 23.81 ± 14.78 * | 17.74 ± 10.46 * | 9.42 ± 1.57 * | 23.37 ± 3.89 * | >512 | >512 | 0.00 ± 0.00 | 1.25 ± 0.00 | 44.20 ± 0.00 * |
| H2 | 10.42 ± 1.72 * | 14.36 ± 10.46 * | 18.05 ± 4.05 * | 38.62 ± 5.25 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 50.00 ± 5.18 * |
| H3 | 7.44 ± 2.98 | 5.91 ± 1.69 | 3.92 ± 4.44 | 28.46 ± 6.64 * | >512 | >512 | 0.00 ± 0.00 | 1.25 ± 0.00 | 37.00 ± 4.94 * |
| H4 | 8.18 ± 2.85 | 36.32 ± 4.25 * | 17.27 ± 6.47 * | 26.42 ± 5.25 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 60.00 ± 0.00 * |
| H5 | 16.37 ± 5.70 * | 39.70 ± 12.75 * | 1.57 ± 6.69 | 31.50 ± 6.94 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 74.40 ± 0.00 * |
| H6 | 11.16 ± 4.46 * | 28.72 ± 6.47 * | 22.76 ± 4.44 * | 28.46 ± 3.32 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 55.70 ± 0.02 * |
| H7 | 13.39 ± 7.87 * | 42.23 ± 1.95 * | 26.39 ± 1.57 * | 29.47 ± 3.89 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 63.30 ± 6.00 * |
| H8 | 20.83 ± 5.43 * | 10.98 ± 7.49 | 25.90 ± 9.24 * | 33.54 ± 3.89 * | >512 | >512 | 0.00 ± 0.00 | 2.50 ± 0.00 | 59.60 ± 0.00 * |
| H9 | 29.76 ± 9.41 * | 43.07 ± 10.46 * | 34.54 ± 10.37 * | 46.75 ± 4.07 * | >512 | >512 | 18.75 ± 0.00 | 0.25 ± 0.00 | 79.50 ± 0.00 * |
| H10 | 15.63 ± 7.83 * | 13.51 ± 18.09 | 20.67 ± 3.95 * | 25.41 ± 2.03 * | >512 | >512 | 0.00 ± 0.00 | 0.00 ± 0.00 | 72.60 ± 5.88 * |
| Control 4 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | / | / | / | / | / |
| Donepezil 5 | / | / | / | / | / | / | / | / | 99.89 ± 0.01 |
| LSD | 10.54 | 14.01 | 8.66 | 6.84 | / | / | / | / | 4.51 |
2.1.4. AChE Inhibition
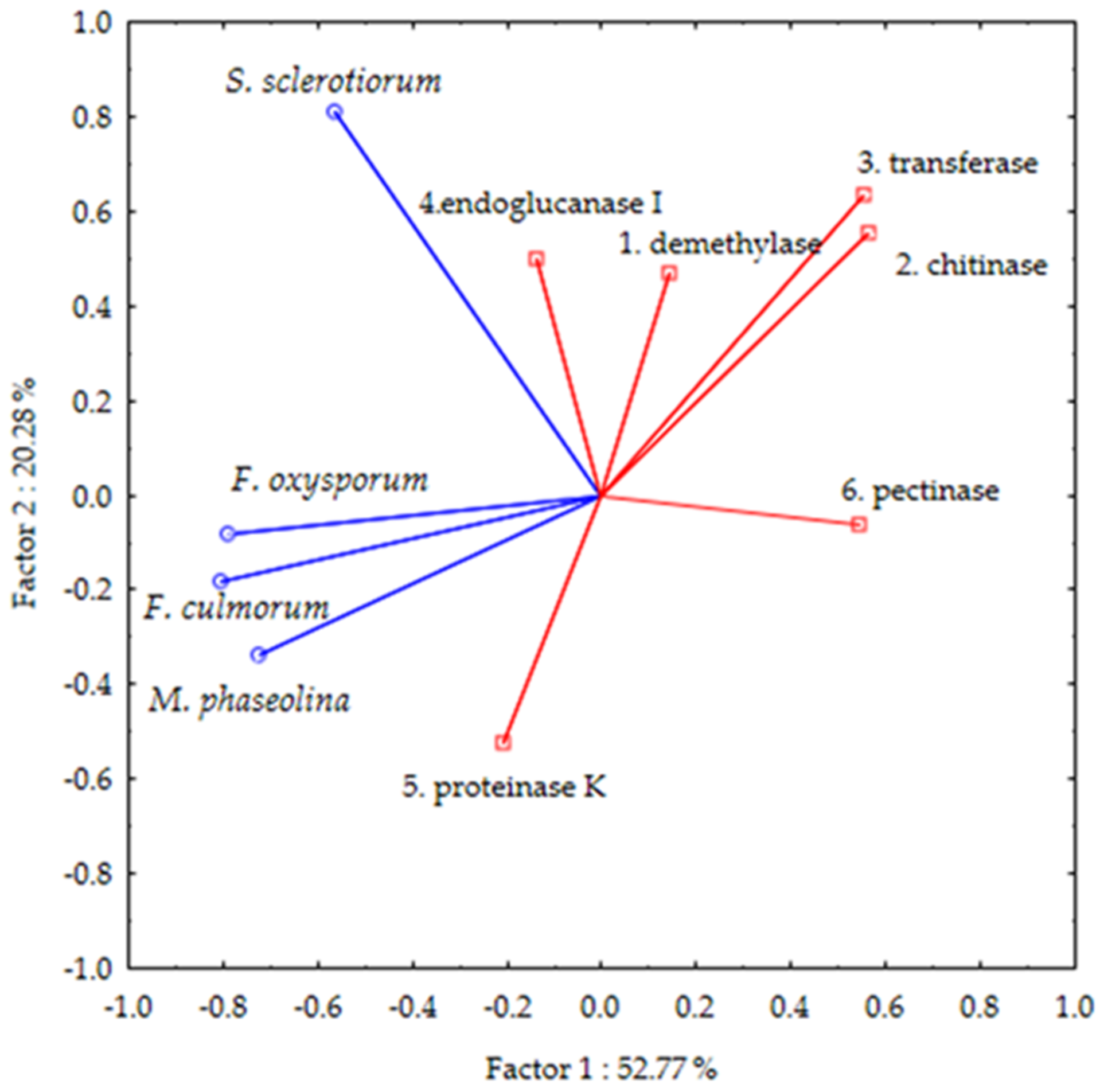
2.2. In-Silico Analyses
2.2.1. “Pesticide-Likeness” and Toxicity Parameters
| Code of Molecule | Oral Rat LD50 (mg/kg bw) a | Tetrahymena pyriformis pIGC50 48-h (mol/L) b | Fathead Minnow pLC50 96-h (mol/L) c | Mutagenicity Value (Result) d | Bioaccumulation Factor (logBAF/L kg−1) e |
|---|---|---|---|---|---|
| H1 | 977.34 (NN) | 4.69 (NN) | 5.78 | 0.70 (pos) | 1.37 |
| H2 | 977.34 (NN) | 4.69 (NN) | 5.82 | 0.67 (pos) | 1.39 |
| H3 | 937.90 (NN) | 4.58 (NN) | 5.37 | 0.57 (pos) | 1.51 |
| H4 | 464.23 (NN) | 5.10 (NN) | 8.24 | 0.37 (neg) | 1.30 |
| H5 | 977.34 (NN) | 4.69 (NN) | 5.80 | 0.69 (pos) | 1.40 |
| H6 | 931.38 (NN) | 4.58 (NN) | 5.26 | 0.64 (pos) | 1.27 |
| H7 | 1075.71 (NN) | 4.69 (NN) | 6.13 | 0.45 (neg) | 1.42 |
| H8 | 1137.40 (NN) | 4.58 (NN) | 5.89 | 0.63 (pos) | 1.55 |
| H9 | 991.78 (NN) | 4.58 (NN) | 5.90 | 0.63 (pos) | 1.63 |
| H10 | 1020.09 (NN) | 4.58 (NN) | 5.33 | 0.31 (neg) | 1.34 |
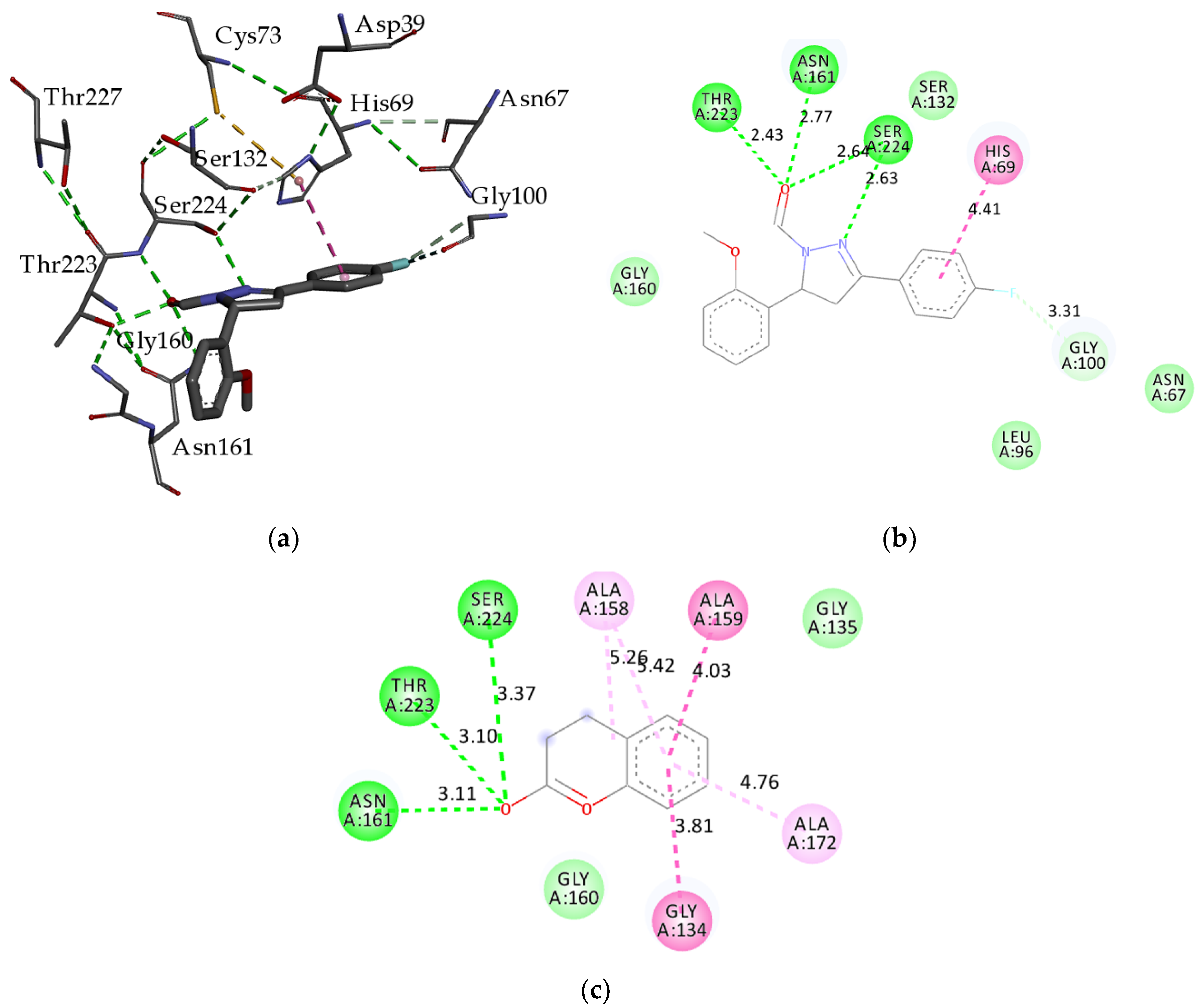
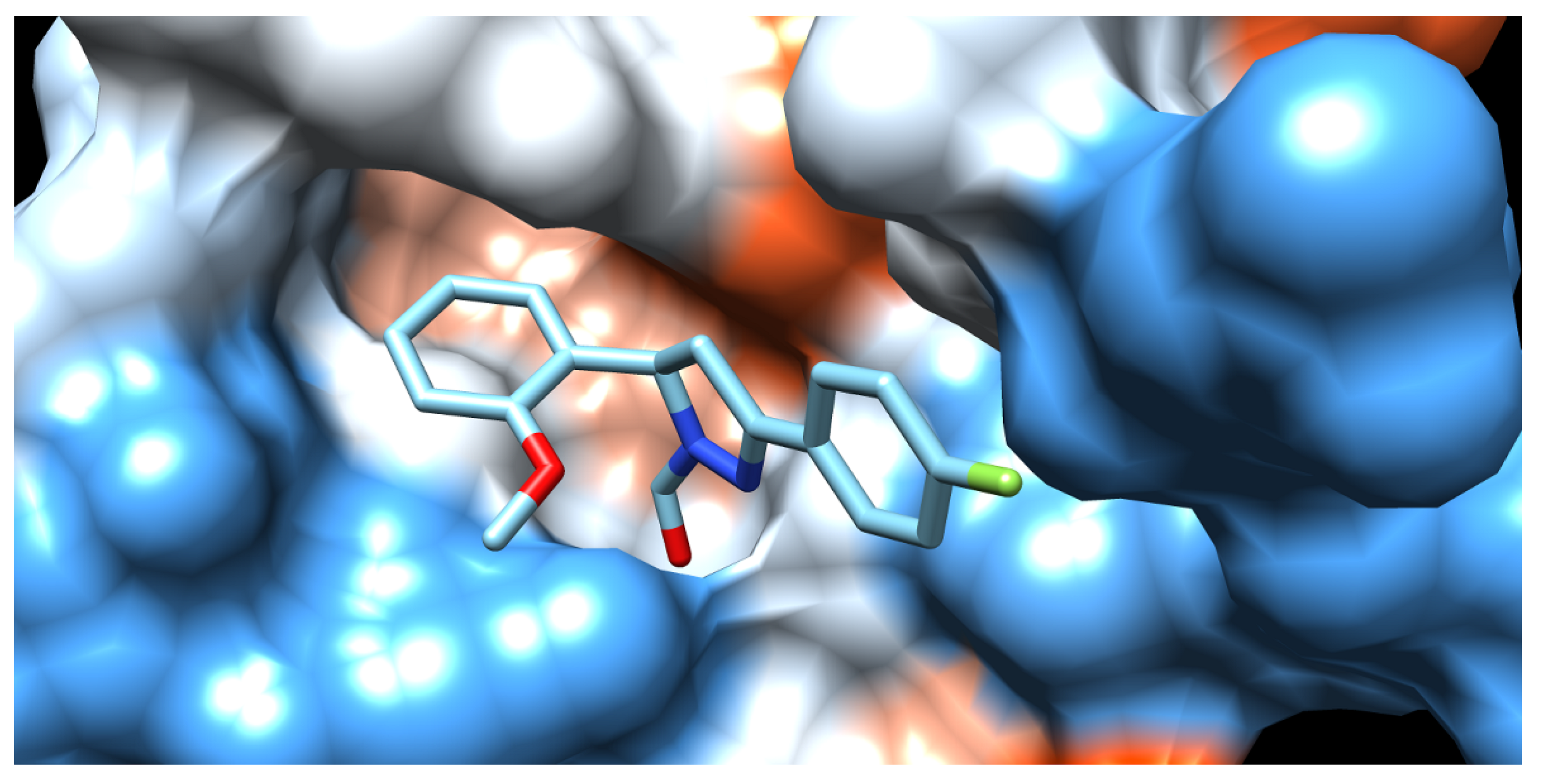
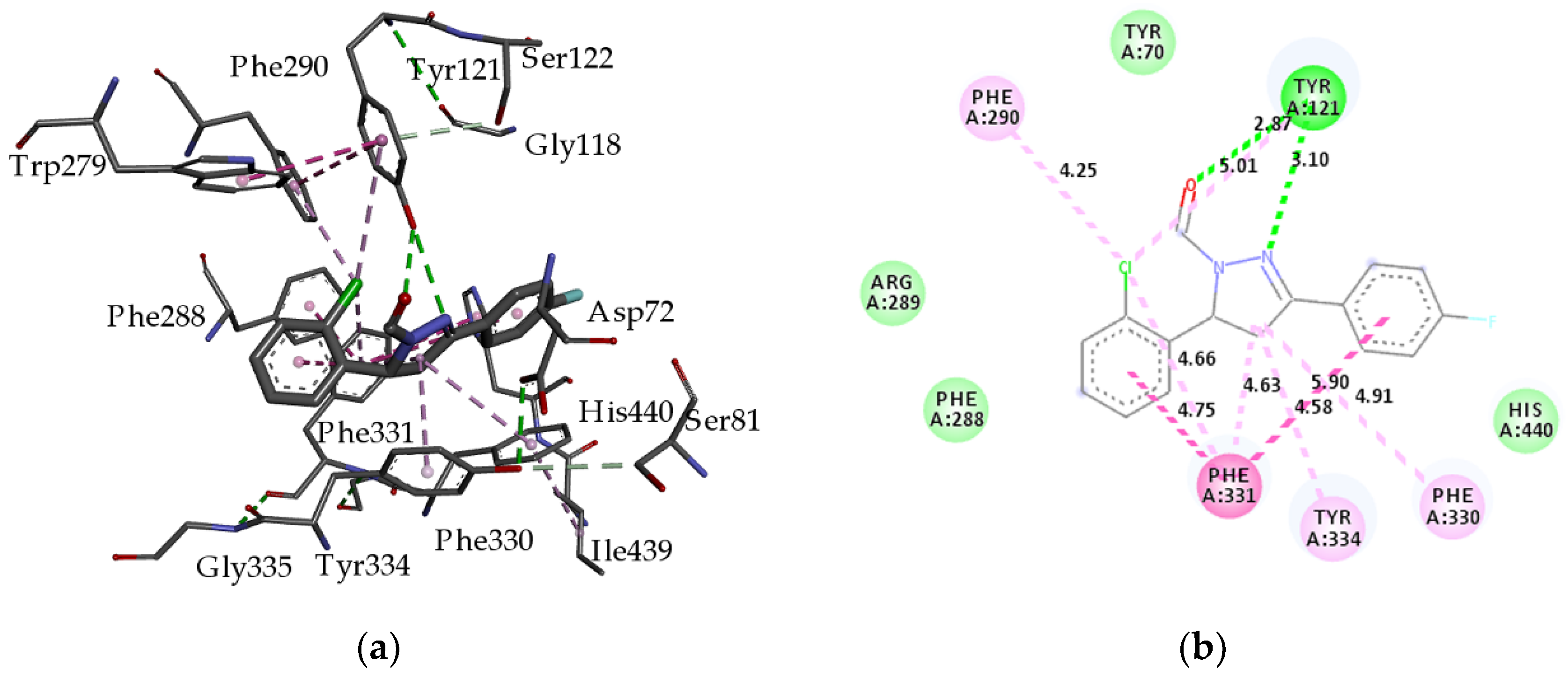
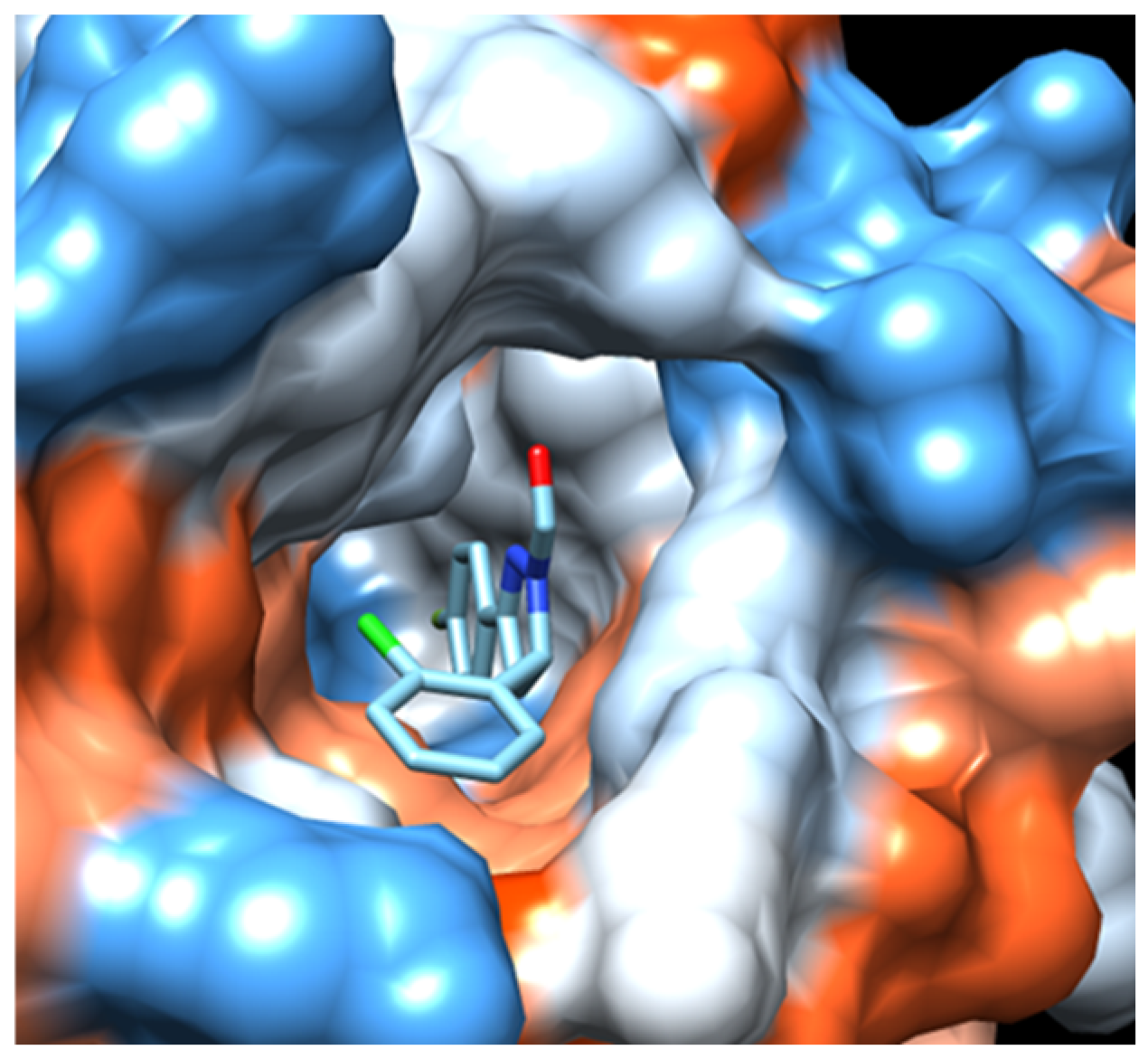
2.2.2. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Biological Assays
4.1.1. Antifungal Assays
4.1.2. Antibacterial Assays
4.1.3. Nematicidal Assays
4.1.4. Acetylcholinesterase Inhibition Assays
4.2. Computational Methods
4.2.1. Calculation of Pesticide-Likeness Molecular Descriptors
4.2.2. Calculation of Toxicity
4.2.3. Molecular Docking
4.2.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, K.; Evans, D.A.; El-Hiti, G.A. Role of modern chemistry in sustainable arable crop protection. Phil. Trans. R. Soc. B 2008, 363, 623–637. [Google Scholar] [CrossRef]
- European Parlament and of the Council, Directive 2009/128/EC, of 21 October 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32009L0128 (accessed on 4 November 2022).
- Umetsu, N.; Shirai, Y. Development of novel pesticides in the 21st century. J. Pestic. Sci. 2020, 45, 54–74. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Gao, Y.; He, L.; Zhu, J.; Cheng, J.; Li, B.; Liu, F.; Mu, W. The relationship between features enabling SDHI fungicide binding to the Sc-Sdh complex and its inhibitory activity against Sclerotinia sclerotiorum. Pest Manag. Sci. 2020, 76, 2799–2808. [Google Scholar] [CrossRef]
- Marquez, N.; Giachero, M.L.; Declerck, S.; Ducasse, D.A. Macrophomina phaseolina: General Characteristics of Pathogenicity and Methods of Control. Front. Plant Sci. 2021, 12, 634397. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- Srinivas, C.; Nirmala Devi, D.; Narasimha Murthy, K.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.P.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, H.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. The wheat pathogen Fusarium culmorum. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Mačkinaite, R. Fusarium culmorum (Wm. G. Sm.) Sacc. interaction with other root-associated fungi of cereals. Biologija 2001, 3, 30–33. [Google Scholar]
- Majic, I.; Sarajlic, A.; Lakatos, T.; Toth, T.; Raspudic, E.; Puskadija, Z.; Kanizai Saric, G.; Laznik, Z. Virulence of new strain of Heterorhabditis bacteriophora from Croatia against Lasioptera rubi. Plant Prot. Sci. 2019, 55, 131–141. [Google Scholar] [CrossRef]
- Bai, X.; Adams, B.J.; Ciche, T.A.; Clifton, S.; Gaugler, R.; Kim, K.S.; Grewal, P.S. A lover and a fighter: The genome sequence of an entomopathogenic nematode Heterorhabditis bacteriophora. PLoS ONE 2013, 8, e69618. [Google Scholar] [CrossRef]
- Mladenović, M.; Arsić, B.B.; Stanković, N.; Mihović, N.; Ragno, R.; Regan, A.; Milićević, J.S.; Trtić-Petrović, T.M.; Micić, R. The targeted pesticides as acetylcholinesterase inhibitors: Comprehensive cross-organism molecular modelling studies performed to anticipate the pharmacology of harmfulness to humans in vitro. Molecules 2018, 23, 2192. [Google Scholar] [CrossRef]
- Atwa, A.A.; Shamseldean, M.M.; Yonis, F.A. The effect of different pesticides on reproduction of entomopathogenic nematodes. Türk. Entomol. Derg. 2013, 37, 493–502. [Google Scholar]
- Rastija, V.; Brahmbhatt, H.; Molnar, M.; Lončarić, M.; Strelec, I.; Komar, M.; Pavić, V. Synthesis, tyrosinase inhibiting activity and molecular docking of fluorinated pyrazole aldehydes as phosphodiesterase inhibitors. Appl. Sci. 2019, 9, 1704. [Google Scholar] [CrossRef]
- Yi, Y.; de Jong, A.; Frenzel, E.; Kuipers, O.P. Comparative transcriptomics of Bacillus mycoides strains in response to potato-root exudates reveals different genetic adaptation of endophytic and soil isolates. Front. Microbiol. 2017, 8, 1487. [Google Scholar] [CrossRef]
- Shah, V.; Subramaniam, S. Bradyrhizobium japonicum USDA110: A representative model organism for studying the impact of pollutants on soil microbiota. Sci. Total Environ. 2018, 264, 963–967. [Google Scholar] [CrossRef]
- Alam, J.; Alam, O.; Alam, P.; Naim, M.J. A Review on pyrazole chemical entity and biological activity. Int. J. Pharm. Sci. Res. 2015, 6, 1433–1442. [Google Scholar]
- Sahu, S.K.; Banerjee, M.; Samantray, A.; Behera, C.; Azam, M.A. Synthesis, analgesic, anti-inflammatory and antimicrobial activities of some novel pyrazoline serivatives tropica. Trop. J. Pharm. Res. 2008, 7, 961–968. [Google Scholar] [CrossRef]
- Kumar, R.S.; Arfi, I.A.; Ahamed, A.; Idhayadhulla, A. Anti-inflammatory and antimicrobial activities of novel pyrazole anlogues. Saudi. J. Biol. Sci. 2016, 23, 614–620. [Google Scholar] [CrossRef]
- B’Bhatt, H.; Sharma, S. Synthesis and antimicrobial activity of pyrazole nucleus containing 2-thioxothiazolidin-4-one derivatives. Arab. J. Chem. 2017, 10, S1590–S1596. [Google Scholar] [CrossRef]
- Ouyang, G.; Cai, X.J.; Chen, Z.; Song, B.A.; Bhadury, P.S.; Yang, S.; Jin, L.H.; Xue, W.; Hu, D.Y.; Zeng, S. Synthesis and antiviral activities of pyrazole derivatives containing an oxime moiety. J. Agric. Food Chem. 2008, 56, 10160–10167. [Google Scholar] [CrossRef]
- El-Sabbagh, O.I.; Baraka, M.M.; Ibrahim, S.M.; Pannecouque, C.; Andrei, G.; Snoeck, R.; Balzarini, J.; Rashad, A.A. Synthesis and antiviral activity of new pyrazole and thiazole derivatives. Eur. J. Med. Chem. 2009, 44, 3746–3753. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Liu, X.H.; Wang, B.L.; Wang, S.H.; Li, Z.M. Synthesis and antifungal activities of new pyrazole derivatives via 1,3-dipolar cycloaddition reaction. Chem. Biol. Drug Des. 2010, 75, 489–493. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, Y. Synthesis and antifungal activity of the derivatives of novel pyrazole carboxamide and isoxazolol pyrazole carboxylate. Molecules 2015, 20, 4383–4394. [Google Scholar] [CrossRef]
- Yu, B.; Zhou, S.; Cao, L.; Hao, Z.; Yang, D.; Guo, X.; Zhang, N.; Bakulev, A.; Fan, Z. Design, synthesis, and evaluation of the antifungal activity of novel pyrazole–thiazole carboxamides as succinate dehydrogenase inhibitors. J. Agric. Food Chem. 2020, 68, 7093–7102. [Google Scholar] [CrossRef]
- Zhao, Y.; Gao, L.; Li, H.; Sun, P.; Meng, F.; Zhang, Y.; Xie, Y.; Sun, B.; Zhou, S.; Ma, Y.; et al. Synthesis, insecticidal activities, and structure-activity relationship of phenylpyrazole derivatives containing a fluoro-substituted benzene moiety. J. Agric. Food Chem. 2020, 68, 11282–11289. [Google Scholar] [CrossRef]
- Yang, S.; Peng, H.; Tang, J.; Fan, S.; Zhao, C.; Xu, H.; Yao, G. Discovery of novel N-pyridylpyrazole thiazole derivatives as insecticide leads. Agronomy 2022, 12, 2472. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, X.; Qu, L.; Xu, S.; Zhao, Y.; Xie, R.; Huang, M.; Zhang, Y. Design, synthesis, antibacterial and insecticidal activities of novel N-phenylpyrazole fraxinellone hybrid compounds. RSC Adv. 2017, 7, 11796–11802. [Google Scholar] [CrossRef]
- Dai, H.; Yao, W.; Fang, Y.; Sun, S.; Shi, Y.; Chen, J.; Jiang, G.; Shi, J. Design, synthesis and bioactivities of novel isoxazole-containing pyrazole oxime derivatives. Molecules 2017, 22, 2000. [Google Scholar] [CrossRef]
- Maienfisch, P.; Hall, R.G. The importance of fluorine in the life science industry. Chimia 2004, 58, 93–99. [Google Scholar] [CrossRef]
- Mykhailiuk, P.K. Fluorinated pyrazoles: From synthesis to applications. Chem. Rev. 2021, 121, 1670–1715. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Song, H.; Wang, Q. Fluorine-containing agrochemicals in the last decade and approaches for fluorine incorporation. Chin. Chem. Lett. 2022, 33, 626–642. [Google Scholar] [CrossRef]
- Fustero, S.; Román, R.; Sanz-Cervera, J.F.; Simón-Fuentes, A.; Bueno, J.; Villanova, S. Synthesis of new fluorinated Tebufenpyrad analogs with acaricidal activity through regioselective pyrazole formation. J. Org. Chem. 2008, 73, 8545–8552. [Google Scholar] [CrossRef] [PubMed]
- European Chemicals Agency. The Use of Alternatives to Testing on Animals for the REACH Regulation. ECHA-11-R-004.2-EN. 2011. Available online: https://echa.europa.eu/documents/10162/13639/alternatives_test_animals_2011_en.pdf/9b0f7e93-4d61-401d-ba2c-80b3b9faaf66 (accessed on 4 November 2022).
- Jitonnom, J. Computer-aided pesticide design: A short review. In Short Views on Insect Biochemistry and Molecular Biology; Section VIII: Insect, Bioinformatics; Chandrasekar, R., Tyagi, B.K., Gui, Z.Z., Reeck, G.R., Eds.; International Book Mission Academic Publisher: Tamilnadu, India, 2014; Volume 2, pp. 685–707. [Google Scholar]
- Meng, X.Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Tyndall, J.D.A.; Sabherwal, M.; Sagatova, A.A.; Keniya, M.V.; Negroni, J.; Wilson, R.K.; Woods, M.A.; Tietjen, K.; Monk, B.C. Structural and functional elucidation of yeast lanosterol 14-demethylase in complex with agrochemical antifungals. PLoS ONE 2016, 11, e0167485. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, X.; Li, B.; Ma, Z. Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F. graminearum collected from wheat in China. Phytopathology 2009, 99, 487–497. [Google Scholar] [CrossRef]
- Lockhart, D.E.A.; Schuettelkopf, A.; Blair, D.E.; van Aalten, D.M.F. Screening-based discovery of Aspergillus fumigatus plant-type chitinase inhibitors. FEBS Lett. 2014, 588, 3282–3290. [Google Scholar] [CrossRef]
- Wu, J.; Tao, Y.; Zhang, M.; Howard, M.H.; Gutteridge, S.; Ding, J. Crystal structures of Saccharomyces cerevisiae N-myristoyltransferase with bound myristoyl-CoA and inhibitors reveal the functional roles of the N-terminal region. J. Biol. Chem. 2007, 282, 22185–22194. [Google Scholar] [CrossRef]
- Guerrero-Perilla, C.; Bernal, F.A.; Coy-Barrera, E.D. Molecular docking study of naturally-occurring compounds as inhibitors of N-myristoyl transferase towards antifungal agents discovery. Rev. Colomb. Cienc. Quím. Farm. 2015, 44, 162–178. [Google Scholar] [CrossRef]
- Sulzenbacher, G.; Schülein, M.; Davies, G.J. Structure of the endoglucanase I from Fusarium oxysporum: Native, cellobiose, and 3,4-epoxybutyl β-D-cellobioside-inhibited forms, at 2.3 Å resolution. Biochemistry 1997, 36, 5902–5911. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Thangavelu, B.; Chun, S.C.; Sathiyabama, M. Proteases from phytopathogenic fungi and their importance in phytopathogenicity. J. Gen. Plant Pathol. 2016, 82, 233–239. [Google Scholar] [CrossRef]
- Olivieri, F.; Zanetti, E.; Oliva, C.R.; Covarrubias, A.A.; Casalongué, C.A. Characterization of an extracellular serine protease of Fusarium eumartii and its action on pathogenesis related proteins. Eur. J. Plant. Pathol. 2002, 108, 63–72. [Google Scholar] [CrossRef]
- Ahmad, M.S.; Akbar, Z.; Choudhary, M.I. Insight into the binding affinity of thiourea in the calcium binding pocket of proteinase K, through high resolution X-ray crystallography. Bioorg. Chem. 2020, 94, 103443. [Google Scholar] [CrossRef]
- Santen, Y.; Benen, J.A.E.; Schröter, K.H.; Kalk, K.H.; Armand, S.; Visser, J.; Dijkstra, B.W. 1.68-Å Crystal structure of endopolygalacturonase II from Aspergillus niger and identification of active site residues by site-directed mutagenesis. J. Biol. Chem. 1999, 274, 30474–30480. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Xu, L.; Peng, H.; Zhu, W.; Tanaka, K.; Cheng, J.; Sanguinet, K.A.; Vandemark, G.; Chen, W. A fungal extracellular effector inactivates plant polygalacturonase-inhibiting protein. Nat. Commun. 2022, 13, 2213. [Google Scholar] [CrossRef]
- Rastija, V.; Vrandečić, K.; Ćosić, J.; Majić, I.; Šarić, G.K.; Agić, D.; Karnaš, M.; Lončarić, M.; Molnar, M. Biological activities related to plant protection and environmental effects of coumarin derivatives: QSAR and molecular docking studies. Int. J. Mol. Sci. 2021, 22, 7283. [Google Scholar] [CrossRef]
- Molnar, M.; Pavić, V.; Šarkanj, B.; Čačić, M.; Vuković, D.; Klemkar, J. Mono- and bis-dipicolinic acid heterocyclic derivatives– thiosemicarbazides, triazoles, oxadiazoles and thiazolidinones as antifungal and antioxidant agents. Heterocycl. Comm. 2017, 23, 35–42. [Google Scholar] [CrossRef]
- Lončarić, M.; Strelec, I.; Pavić, V.; Šubarić, D.; Rastija, V.; Molnar, M. Lipoxygenase Inhibition Activity of Coumarin Derivatives—QSAR and Molecular Docking Study. Pharmaceuticals 2020, 13, 154. [Google Scholar] [CrossRef]
- Rastija, V.; Vrandečić, K.; Ćosić, J.; Šarić, G.K.; Majić, I.; Agić, D.; Šubarić, D.; Karnaš, M.; Bešlo, D.; Komar, M.; et al. Effects of coumarinyl Schiff bases against phytopathogenic fungi, the soil-beneficial bacteria and entomopathogenic nematodes: Deeper insight into the mechanism of action. Molecules 2022, 27, 2196. [Google Scholar] [CrossRef]
- Kang, J.S.; Kim, E.; Lee, S.H.; Park, I.K. Inhibition of acetylcholinesterases of the pinewood nematode, Bursaphelenchus xylophilus, by phytochemicals from plant essential oils. Pestic. Biochem. Physiol. 2013, 105, 50–56. [Google Scholar] [CrossRef]
- Hao, G.; Dong, Q.; Yang, G.A. Comparative study on the constitutive properties of marketed pesticides. Mol. Inform. 2011, 30, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Sripriya, N.; Ranjith, K.M.; Ashwin, K.N.; Bhuvaneswari, S.; Udaya, P.N.K. In silico evaluation of multispecies toxicity of natural compounds. Drug Chem. Toxicol. 2021, 44, 480–486. [Google Scholar] [CrossRef]
- Martin, T.M. User’s Guide for T.E.S.T., Version 5.1; Toxicity Estimation Software Tool: A Program to Estimate Toxicity from Molecular Structure; U.S. Environmental Protection Agency: Cincinati, OH, USA, 2020. Available online: https://www.epa.gov/sites/default/files/2016-05/documents/600r16058.pdf (accessed on 4 November 2022).
- Costanza, J.; Lynch, D.G.; Boethling, R.; Arnot, J.A. Use of the bioaccumulation factor to screen chemicals for bioaccumulation potential. Environ. Toxicol. Chem. 2012, 31, 2261–2268. [Google Scholar] [CrossRef] [PubMed]
- Bulgheroni, A.; Kinsner-Ovaskainen, A.; Hoffmann, S.; Hartung, T.; Prieto, P. Estimation of acute oral toxicity using the No Observed Adverse Effect Level (NOAEL) from the 28 day repeated dose toxicity studies in rats. Regul. Toxicol. Pharmacol. 2009, 53, 16–19. [Google Scholar] [CrossRef]
- Kryger, G.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with E2020 (Aricept®): Implications for the design of new anti-Alzheimer drugs. Structure 1999, 7, 297–307. [Google Scholar] [CrossRef]
- Müler, A.; Hinrichs, W.; Wolf, W.M.; Saenger, W. Crystal structure of calcium-free proteinase K at 1.5-Å resolution. J. Biol. Chem. 1994, 269, 23108–23111. [Google Scholar] [CrossRef]
- Bajda, M.; Więckowska, A.; Hebda, M.; Guzior, N.; Sotriffer, C.; Malawska, B. Structure-based search for new inhibitors of cholinesterases. Int. J. Mol. Sci. 2013, 14, 5608–5632. [Google Scholar] [CrossRef]
- Yang, E.; Xu, T.; Fan, J.; Zhang, Q.; Ding, M.; Huang, M.; Deng, L.; Lu, Y.; Guo, Y. Natural products-based pesticides: Design, synthesis and pesticidal activities of novel fraxinellone derivatives containing N-phenylpyrazole moiety. Ind. Crops Prod. 2018, 117, 50–57. [Google Scholar] [CrossRef]
- Qiao, L.; Zhai, Z.-W.; Cai, P.-P.; Tan, C.-X.; Weng, J.-Q.; Han, L.; Liu, X.-H.; Zhang, Y.-G. Synthesis, Crystal Structure, Antifungal Activity, and Docking Study of Difluoromethyl Pyrazole Derivatives. J. Heterocycl. Chem. 2019, 56, 2536–2541. [Google Scholar] [CrossRef]
- Wu, Z.; Hu, D.; Kuang, J.; Cai, H.; Wu, S.; Xue, W. Synthesis and antifungal activity of N-(substituted pyridinyl)-1-methyl(phenyl)-3-(trifluoromethyl)-1H-pyrazole-4-carboxamide derivatives. Molecules 2012, 17, 14205–14218. [Google Scholar] [CrossRef]
- Bondock, S.; Fadaly, W.; Metwally, M.A. Synthesis and antimicrobial activity of some new thiazole, thiophene and pyrazole derivatives containing benzothiazole moiety. Eur. J. Med. Chem. 2010, 45, 3692–3701. [Google Scholar] [CrossRef] [PubMed]
- Bajorath, J.; Saenger, W.; Pal, G.P. Autolysis and inhibition of proteinase K, a subtilisin-related serine proteinase isolated from the fungus Tritirachium album Limber. Biochim. Biophys. Acta 1988, 954, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Pekkarinen, A.I.; Jones, B.L.; Niku-Paavola, M.L. Purification and properties of an alkaline proteinase of Fusarium culmorum. Eur. J. Biochem. 2002, 269, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Pochet, L.; Frédérick, R.; Masereel, B. Coumarin and isocoumarin as serine protease inhibitors. Curr. Pharm. Des. 2004, 10, 3781–3796. [Google Scholar] [CrossRef]
- Lin, Y.; Wei, W.; Yuanyuan, P.; Fuping, L. Screening and identification of cellulase-producing strain of Fusarium oxysporum. Procedia Environ. Sci. 2012, 12, 1213–1219. [Google Scholar] [CrossRef]
- Ramos, A.M.; Gally, M.; Szapiro, G.; Itzcovich, T.; Carabajal, M.; Levin, L. In vitro growth and cell wall degrading enzyme production by Argentinean isolates of Macrophomina phaseolina, the causative agent of charcoal rot in corn. Rev. Argent. Microbiol. 2016, 48, 267–273. [Google Scholar] [CrossRef]
- Wang, H.; Jones, R.W. A unique endoglucanase-encoding gene cloned from the phytopathogenic fungus Macrophomina phaseolina. Appl. Environ. Microbiol. 1995, 61, 2004–2006. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Shen, Z.-H.; Xing, J.-H.; Yang, G.; Xu, T.-M.; Peng, W.-L.; Liu, X.-H. Synthesis and nematocidal activity of novel 1-(3-chloropyridin-2-yl)-3-(trifluoromethyl)-1H-pyrazole-4-carboxamide derivatives. Chem. Pap. 2017, 71, 921–928. [Google Scholar] [CrossRef]
- Liu, X.-H.; Zhao, W.; Shen, Z.-H.; Xing, J.-H.; Xu, T.-M.; Peng, W.-L. Synthesis, nematocidal activity and SAR study of novel difluoromethylpyrazole carboxamide derivatives containing flexible alkyl chain moieties. Eur. J. Med. Chem. 2017, 125, 881–889. [Google Scholar] [CrossRef]
- Radová, Š. Effect of selected pesticides on survival and virulence of two nematode species. Pol. J. Environ. Stud. 2011, 20, 181–185. Available online: http://www.pjoes.com/pdf-88544-22403?filename=Effects%20of%20Selected.pdf (accessed on 1 November 2022).
- De Boer, D.; Nguyen, N.; Mao, J.; Moore, J.; Sorin, E.J.A. Comprehensive review of cholinesterase modeling and simulation. Biomolecules 2021, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Kiametis, A.S.; Treptow, W. Donepezil inhibits acetylcholinesterase via multiple binding modes at room temperature. J. Chem. Inf. Modeling 2020, 60, 3463–3471. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Ali, M.T.; Shawan, M.M.A.K.; Sarwar, M.G.; Khan, M.A.K.; Halim, M.A. Halogen-directed drug design for Alzheimer’s disease: A combined density functional and molecular docking study. SpringerPlus 2016, 5, 1346. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, J.; Shen, H.; Cheng, J.; Li, Z.; Xu, X. Synthesis, nematicidal activity and docking study of novel chromone derivatives containing substituted pyrazole. Chin. Chem. Lett. 2019, 29, 911–914. [Google Scholar] [CrossRef]
- Gutiérret, C.; Campos-Herrera, R.; Jiménez, J. Comparative study of the effect of selected agrochemical products on Steinernema feltiae (Rhabditida: Steinernematidae). Biocontrol. Sci. Technol. 2008, 18, 101–108. [Google Scholar] [CrossRef]
- Worek, F.; Eyer, P.; Thiermann, H. Determination of acetylcholinesterase activity by the Ellman assay: A versatile tool for in vitro research on medical countermeasures against organophosphate poisoning. Drug Test. Anal. 2012, 4, 282–291. [Google Scholar] [CrossRef]
- Čolović, M.B.; Krstić, D.Z.; Lazarević-Pašti, T.D.; Bondžić, A.M.; Vasić, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Singh, K.D.; Labala, R.K.; Devi, T.B.; Singh, N.I.; Chanu, H.D.; Sougrakpam, S.; Nameirakpam, B.S.; Sahoo, D.; Rajashekar, Y. Biochemical efficacy, molecular docking and inhibitory effect of 2,3-dimethylmaleic anhydride on insect acetylcholinesterase. Sci. Rep. 2017, 7, 12483. [Google Scholar] [CrossRef]
- Bhurta, D.; Bharate, S.B. Styryl group, a friend or foe in medicinal chemistry. ChemMedChem 2022, 17, e202100706. [Google Scholar] [CrossRef]
- Siber, T.; Bušić, V.; Zobundžija, D.; Roca, S.; Vikić-Topić, D.; Vrandečić, K.; Gašo-Sokač, D. An improved method for the quaternization of nicotinamide and antifungal activities of its derivatives. Molecules 2019, 24, 1001. [Google Scholar] [CrossRef]
- Bušić, V.; Vrandečić, K.; Siber, T.; Roca, S.; Vikić Topić, D.; Gašo Sokač, D. A rapid microwave induced synthesis of isonicotinamide derivatives and their antifungal activity. Croat. Chem. Acta 2019, 92, 125–135. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hanckok, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Jagodič, A.; Majić, I.; Trdan, S.; Laznik, Ž. Are synthetic VOC, typically emitted by barley (Hordeum vulgare L.) roots, navigation signals for entomopathogenic nematodes (Steinernema and Heterorhabditis)? Russ. J. Nematol. 2020, 28, 29–39. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Schultz, T.W.; Sparfkin, C.L.; Aptula, A.O. Reactivity-based toxicity modelling of five-membered heterocyclic compounds: Application to Tetrahymena pyriformis. SAR QSAR Environ. Res. 2010, 21, 681–691. [Google Scholar] [CrossRef]
- Hansen, K.; Mika, S.; Schroeter, T.; Sutter, A.; ter Laak, A.; Steger-Hartmann, T.; Heinrich, N.; Müller, K.R. Benchmark Data set for in silico prediction of ames mutagenicity. J. Chem. Inf. Model. 2009, 49, 2077–2081. [Google Scholar] [CrossRef]
- Hocquet, A.; Langgård, M. An Evaluation of the MM+ Force Field. J. Mol. Model. 1998, 4, 94–112. [Google Scholar] [CrossRef]
- Stewart, J.J.P. Optimization of parameters for semiempirical methods I. Method. J. Comput. Chem. 1989, 10, 209–220. [Google Scholar] [CrossRef]
- Hsu, K.-C.; Chen, Y.-F.; Lin, S.-R.; Yang, J.-M. iGEMDOCK: A graphical environment of enhancing GEMDOCK using pharmacological interactions and post-screening analysis. BMC Bioinform. 2011, 12 (Suppl. S1), S33. [Google Scholar] [CrossRef]






| Code of Molecule | MW | MLOGP | HBA | HBD | RB | AB |
|---|---|---|---|---|---|---|
| H1 | 298.34 | 3.211 | 5 | 0 | 3 | 12 |
| H2 | 298.34 | 3.211 | 5 | 0 | 3 | 12 |
| H3 | 286.3 | 3.899 | 5 | 0 | 2 | 12 |
| H4 | 370.45 | 5.007 | 4 | 0 | 4 | 18 |
| H5 | 298.34 | 3.211 | 5 | 0 | 3 | 12 |
| H6 | 284.31 | 2.971 | 5 | 1 | 2 | 12 |
| H7 | 328.37 | 2.926 | 6 | 0 | 4 | 12 |
| H8 | 347.2 | 4.137 | 4 | 0 | 2 | 12 |
| H9 | 302.75 | 4.019 | 4 | 0 | 2 | 12 |
| H10 | 311.39 | 3.446 | 5 | 0 | 3 | 12 |
| Demethylase | Chitinase | Transferase | Endoglucanase I | Proteinase K | Pectinase | AChE | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Code of Mol. | Total E | Code of Mol. | Total E | Code of Mol. | Total E | Code of Mol. | Total E | Code of Mol. | Total E | Code of Mol. | Total E | Code of Mol. | Total E |
| H4 | −85.20 | 38f * | −120.51 | 3lp * | −87.65 | H4 | −106.52 | H2 | −97.70 | H10 | −76.66 | e20 * | −117.34 |
| H1 | −81.74 | H4 | −104.99 | H4 | −82.74 | H5 | −103.78 | H10 | −94.29 | H1 | −75.23 | H4 | −108.34 |
| 5lw * | −80.98 | H6 | −98.86 | H6 | −81.44 | in1 * | −101.69 | H9 | −92.93 | H4 | −74.98 | H1 | −100.08 |
| H7 | −79.88 | H5 | −97.41 | H5 | −77.38 | H6 | −101.60 | H5 | −87.31 | H6 | −73.74 | H10 | −99.86 |
| H2 | −78.71 | H1 | −96.77 | H1 | −76.96 | H2 | −101.03 | H8 | −86.93 | Nag * | −73.49 | H5 | −95.48 |
| H5 | −78.21 | H3 | −96.74 | H7 | −76.42 | H1 | −99.22 | H6 | −86.36 | H3 | −73.31 | H9 | −95.08 |
| H8 | −77.94 | H7 | −96.31 | H3 | −75.19 | H9 | −96.67 | H3 | −86.33 | H8 | −73.26 | H7 | −94.89 |
| H6 | −77.39 | H10 | −95.94 | H10 | −73.74 | H8 | −92.21 | H4 | −81.16 | H5 | −72.97 | H6 | −93.44 |
| H10 | −76.52 | H8 | −94.86 | H8 | −73.14 | H7 | −91.48 | H1 | −81.08 | H7 | −71.24 | H2 | −93.09 |
| H9 | −76.42 | H2 | −89.52 | H2 | −72.15 | H10 | −90.50 | H7 | −76.08 | H9 | −70.82 | H3 | −92.24 |
| H3 | −75.40 | H9 | −88.97 | H9 | −68.58 | H3 | −83.88 | Cou * | −72.31 | H2 | −70.26 | H8 | −90.52 |
| Residue | Energy (kcal mol−1) | Residue | Energy (kcal mol−1) |
|---|---|---|---|
| Hydrogen bond | Van der Waals interactions | ||
| S-Asn161 | −6.68 | M-Ala159 | −2.35 |
| M-Thr223 | −2.76 | M-Gly160 | −7.12 |
| S-Thr223 | −1.10 | M-Asn161 | −4.47 |
| M-Ser224 | −3.50 | S-Asn161 | −3.05 |
| S-Ser224 | −9.50 | S-Asn162 | −3.02 |
| Van der Waals interactions | S-Tyr169 | -3.46 | |
| S-Asn67 | −0.85 | M-Thr223 | −1.60 |
| S-Leu96 | −0.69 | M-Ser224 | −1.93 |
| M-Asn99 | −1.14 | S-Ser224 | −3.55 |
| M-Ser132 | −5.26 | Fluorine interactions | |
| M-Leu133 | −8.13 | M-Gly100 | −1.32 |
| M-Gly134 | −6.89 | ||
| M-Gly135 | −3.01 | π–π T−shaped interactions | |
| M-Ala158 | −2.11 | S-His69 | −8.06 |
| Residue | Energy (kcal mol−1) |
|---|---|
| hydrogen bond | |
| S-Tyr121 | −7.07 |
| S-Tyr121 | −6.07 |
| Van der Waals interactions | |
| S-Tyr70 | −1.31 |
| M-Trp84 | −0.02 |
| M-Gly117 | −0.45 |
| S-His440 | −3.87 |
| S-Trp279 | −1.09 |
| π–π stacked interactions | |
| S-Phe290 | −1.09 |
| π–π T-shaped interactions | |
| M-Phe331 | −5.43 |
| π-alkyl interactions | |
| S-Phe330 | −13.08 |
| M-Phe331 | −15.11 |
| S-Tyr334 | −10.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rastija, V.; Vrandečić, K.; Ćosić, J.; Kanižai Šarić, G.; Majić, I.; Agić, D.; Šubarić, D.; Karnaš, M.; Bešlo, D.; Brahmbhatt, H.; et al. Antifungal Activities of Fluorinated Pyrazole Aldehydes on Phytopathogenic Fungi, and Their Effect on Entomopathogenic Nematodes, and Soil-Beneficial Bacteria. Int. J. Mol. Sci. 2023, 24, 9335. https://doi.org/10.3390/ijms24119335
Rastija V, Vrandečić K, Ćosić J, Kanižai Šarić G, Majić I, Agić D, Šubarić D, Karnaš M, Bešlo D, Brahmbhatt H, et al. Antifungal Activities of Fluorinated Pyrazole Aldehydes on Phytopathogenic Fungi, and Their Effect on Entomopathogenic Nematodes, and Soil-Beneficial Bacteria. International Journal of Molecular Sciences. 2023; 24(11):9335. https://doi.org/10.3390/ijms24119335
Chicago/Turabian StyleRastija, Vesna, Karolina Vrandečić, Jasenka Ćosić, Gabriella Kanižai Šarić, Ivana Majić, Dejan Agić, Domagoj Šubarić, Maja Karnaš, Drago Bešlo, Harshad Brahmbhatt, and et al. 2023. "Antifungal Activities of Fluorinated Pyrazole Aldehydes on Phytopathogenic Fungi, and Their Effect on Entomopathogenic Nematodes, and Soil-Beneficial Bacteria" International Journal of Molecular Sciences 24, no. 11: 9335. https://doi.org/10.3390/ijms24119335
APA StyleRastija, V., Vrandečić, K., Ćosić, J., Kanižai Šarić, G., Majić, I., Agić, D., Šubarić, D., Karnaš, M., Bešlo, D., Brahmbhatt, H., & Komar, M. (2023). Antifungal Activities of Fluorinated Pyrazole Aldehydes on Phytopathogenic Fungi, and Their Effect on Entomopathogenic Nematodes, and Soil-Beneficial Bacteria. International Journal of Molecular Sciences, 24(11), 9335. https://doi.org/10.3390/ijms24119335