
The Epigenetics of Migraine
 , , , ,
, , , , 
Abstract
1. Introduction
2. Methodology
2.1. Literature and Search Strategy
2.2. Inclusion and Exclusion Criteria
- (1)
- The study contained original data.
- (2)
- The study was in vitro or in vivo.
- (3)
- The study subjects were human or animal.
- (4)
- The study was written in English.
- (5)
- The study described the interplay between the migraine pathogenesis or potential targets for obtaining effective therapeutic responses and epigenetics.
2.3. Selected Studies
3. Principal Mechanisms of DNA Methylation and Histone Modifications
3.1. DNA Methylation and Demethylation
3.2. Histone Acetylation and Deacetylation
3.2.1. The Principle of Histone Acetylation
3.2.2. Histone Acetylation and Cancer
3.2.3. Histone Acetylation and Immune Response
3.2.4. Histone Acetylation and Embryonic Development
3.2.5. Histone Deacetylase Inhibitors as Therapeutic Agents in Neurologic Disorders
3.3. Histone Methylation and Demethylation
4. Environment, Epigenetics, and Migraine
4.1. Stress
4.2. Diet
4.3. Toxins
5. Aberrant DNA Methylation Patterns in Migraine
6. Migraine Chronification and DNA Methylation
7. Changed Histone Acetylation Patterns in Migraine
8. Epigenetics of Specific Pathways in Migraine
8.1. CGRP, RAMP1, and Migraine
8.1.1. CALCA Gene Epigenetics in Migraine
8.1.2. RAMP1 Epigenetics in Migraine
8.2. The Endocannabinoid System and Migraine
8.2.1. The Endocannabinoid System—A Short Overview
8.2.2. General Aspects of the Epigenetic Regulation of the Endocannabinoid System
8.2.3. Epigenetics of the Endocannabinoid System in Migraine
8.2.4. The Endocannabinoid System Influences Epigenetics
8.3. The Epigenetic Regulation of Other Pathways in Migraine
9. Epigenetics as a Therapeutic Target in Migraine
10. General Aspects of microRNAs and circRNAs in Migraine
11. Principal Mechanisms of Action of microRNAs and circRNAs
11.1. microRNAs—Key Functions and Mechanisms of Action
11.2. circRNAs: Key Functions and Mechanisms of Action
12. Changed Expression Patterns of microRNAs and circRNAs in Migraine
12.1. The Role of microRNA in the Pathophysiology of Migraine
12.1.1. The Role of microRNA in the Pathophysiology of Migraine in Animal Models
12.1.2. The Role of microRNA in the Pathophysiology of Migraine in Humans
12.2. microRNA and Treatment of Migraine
12.3. microRNAs and Clinical Characteristics of Migraine
12.4. The Role of circRNAs in Migraine Pathophysiology
13. Discussion
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AEA | anandamide |
| CSD | cortical spreading depression |
| CGRP | calcitonin gene-related peptide |
| CGIs | CpG islands |
| DNMTs | DNA methyl transferases |
| EM | episodic migraine |
| eCB | endogenous ligands |
| FAAH | fatty acid amide hydrolase |
| GR | glucocorticoid receptor |
| HAT | histone acetyltransferase |
| HDACs | histone deacetylases |
| IDHs | isocitrate dehydrogenase inhibitors |
| JNKs | c-Jun N-terminal kinases |
| MOH | medication overuse headache |
| MTHFR | methylenetetrahydrofolate reductase |
| MZ | monozygotic |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| PTSD | post-traumatic stress disorder |
| RAMP1 | receptor activity-modifying protein 1 |
| TET | ten-eleven translocation |
| THC | Δ9-tetrahydrocannabinol |
| TG | trigeminal ganglion |
References
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 328 Diseases and Injuries for 195 Countries, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M. Headache Classification Committee of the International Headache Society (IHS) the International Classification of Headache Disorders, 3rd Edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef]
- Cvetković, V.V. Is the management of migraine and tension headache in Croatia satisfactory? Acta Med. Croat. 2013, 67, 415–424. [Google Scholar]
- Tzankova, V.; Becker, W.J.; Chan, T.L.H. Diagnosis and Acute Management of Migraine. CMAJ 2023, 195, E153–E158. [Google Scholar] [CrossRef]
- Zobdeh, F.; Kraiem, A.; Attwood, M.M.; Chubarev, V.N.; Tarasov, V.V.; Schiöth, H.B.; Mwinyi, J. Pharmacological Treatment of Migraine: Drug Classes, Mechanisms of Action, Clinical Trials and New Treatments. Br. J. Pharmacol. 2021, 178, 4588–4607. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, T.; Viana, M.; Tassorelli, C. Current Prophylactic Medications for Migraine and Their Potential Mechanisms of Action. Neurotherapeutics 2018, 15, 313–323. [Google Scholar] [CrossRef]
- Burstein, R.; Noseda, R.; Borsook, D. Migraine: Multiple Processes, Complex Pathophysiology. J. Neurosci. 2015, 35, 6619–6629. [Google Scholar] [CrossRef]
- Karsan, N.; Prabhakar, P.; Goadsby, P.J. Characterising the Premonitory Stage of Migraine in Children: A Clinic-Based Study of 100 Patients in a Specialist Headache Service. J. Headache Pain 2016, 17, 94. [Google Scholar] [CrossRef]
- Giffin, N.J.; Ruggiero, L.; Lipton, R.B.; Silberstein, S.D.; Tvedskov, J.F.; Olesen, J.; Altman, J.; Goadsby, P.J.; Macrae, A. Premonitory Symptoms in Migraine: An Electronic Diary Study. Neurology 2003, 60, 935–940. [Google Scholar] [CrossRef]
- Russell, M.; Rasmussen, B.; Fenger, K.; Olesen, J. Migraine without Aura and Migraine with Aura Are Distinct Clinical Entities: A Study of Four Hundred and Eighty-Four Male and Female Migraineurs from the General Population. Cephalalgia 1996, 16, 239–245. [Google Scholar] [CrossRef]
- Pietrobon, D.; Moskowitz, M.A. Pathophysiology of Migraine. Annu. Rev. Physiol. 2013, 75, 365–391. [Google Scholar] [CrossRef] [PubMed]
- Leo, A.A.P.; Morison, R.S. Propagation of spreading cortical depression. J. Neurophysiol. 1945, 8, 33–45. [Google Scholar] [CrossRef]
- Harriott, A.M.; Takizawa, T.; Chung, D.Y.; Chen, S.-P. Spreading Depression as a Preclinical Model of Migraine. J. Headache Pain 2019, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Quintela, E.; Castillo, J.; Muñoz, P.; Pascual, J. Premonitory and Resolution Symptoms in Migraine: A Prospective Study in 100 Unselected Patients. Cephalalgia 2006, 26, 1051–1060. [Google Scholar] [CrossRef]
- Russell, M.B.; Hilden, J.; Sørensen, S.A.; Olesen, J. Familial Occurrence of Migraine without Aura and Migraine with Aura. Neurology 1993, 43, 1369. [Google Scholar] [CrossRef]
- Polderman, T.J.C.; Benyamin, B.; de Leeuw, C.A.; Sullivan, P.F.; van Bochoven, A.; Visscher, P.M.; Posthuma, D. Meta-Analysis of the Heritability of Human Traits Based on Fifty Years of Twin Studies. Nat. Genet. 2015, 47, 702–709. [Google Scholar] [CrossRef]
- Pisanu, C.; Preisig, M.; Castelao, E.; Glaus, J.; Pistis, G.; Squassina, A.; Del Zompo, M.; Merikangas, K.R.; Waeber, G.; Vollenweider, P.; et al. A Genetic Risk Score Is Differentially Associated with Migraine with and without Aura. Hum. Genet. 2017, 136, 999–1008. [Google Scholar] [CrossRef]
- Sutherland, H.G.; Albury, C.L.; Griffiths, L.R. Advances in Genetics of Migraine. J. Headache Pain 2019, 20, 72. [Google Scholar] [CrossRef]
- Russell, M.B.; Ducros, A. Sporadic and Familial Hemiplegic Migraine: Pathophysiological Mechanisms, Clinical Characteristics, Diagnosis, and Management. Lancet Neurol. 2011, 10, 457–470. [Google Scholar] [CrossRef]
- Silveira-Moriyama, L.; Kovac, S.; Kurian, M.A.; Houlden, H.; Lees, A.J.; Walker, M.C.; Roze, E.; Paciorkowski, A.R.; Mink, J.W.; Warner, T.T. Phenotypes, Genotypes, and the Management of Paroxysmal Movement Disorders. Dev. Med. Child Neurol. 2018, 60, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Lafrenière, R.G.; Cader, M.Z.; Poulin, J.-F.; Andres-Enguix, I.; Simoneau, M.; Gupta, N.; Boisvert, K.; Lafrenière, F.; McLaughlan, S.; Dubé, M.-P.; et al. A Dominant-Negative Mutation in the TRESK Potassium Channel Is Linked to Familial Migraine with Aura. Nat. Med. 2010, 16, 1157–1160. [Google Scholar] [CrossRef] [PubMed]
- Fila, M.; Sobczuk, A.; Pawlowska, E.; Blasiak, J. Epigenetic Connection of the Calcitonin Gene-Related Peptide and Its Potential in Migraine. Int. J. Mol. Sci. 2022, 23, 6151. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.; Dias, A.; Sousa, A.; Lopes, A.M.; Martins, S.; Pinto, N.; Lemos, C.; Alves-Ferreira, M. A High Methylation Level of a Novel −284 Bp CpG Island in the RAMP1 Gene Promoter Is Potentially Associated with Migraine in Women. Brain Sci. 2022, 12, 526. [Google Scholar] [CrossRef]
- Park, K.-Y.; Fletcher, J.R.; Raddant, A.C.; Russo, A.F. Epigenetic Regulation of the Calcitonin Gene–Related Peptide Gene in Trigeminal Glia. Cephalalgia 2011, 31, 614–624. [Google Scholar] [CrossRef]
- Greco, R.; De Icco, R.; Demartini, C.; Zanaboni, A.M.; Tumelero, E.; Sances, G.; Allena, M.; Tassorelli, C. Plasma Levels of CGRP and Expression of Specific MicroRNAs in Blood Cells of Episodic and Chronic Migraine Subjects: Towards the Identification of a Panel of Peripheral Biomarkers of Migraine? J. Headache Pain 2020, 21, 122. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of Epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Chuang, J.C.; Jones, P.A. Epigenetics and MicroRNAs. Pediatr. Res. 2007, 61 Pt 2, 24R–29R. [Google Scholar] [CrossRef]
- Burggren, W.W. Dynamics of Epigenetic Phenomena: Intergenerational and Intragenerational Phenotype ‘Washout’. J. Exp. Biol. 2015, 218, 80–87. [Google Scholar] [CrossRef]
- Kane, M.F.; Loda, M.; Gaida, G.M.; Lipman, J.; Mishra, R.; Goldman, H.; Jessup, J.M.; Kolodner, R. Methylation of the HMLH1 Promoter Correlates with Lack of Expression of HMLH1 in Sporadic Colon Tumors and Mismatch Repair-Defective Human Tumor Cell Lines. Cancer Res. 1997, 57, 808–811. [Google Scholar]
- Oostra, B.A.; Jacky, P.B.; Brown, W.T.; Rousseau, F. Guidelines for the Diagnosis of Fragile X Syndrome. National Fragile X Foundation. J. Med. Genet. 1993, 30, 410–413. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in Epigenetics Link Genetics to the Environment and Disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Eising, E.; A Datson, N.; van den Maagdenberg, A.M.; Ferrari, M.D. Epigenetic Mechanisms in Migraine: A Promising Avenue? BMC Med. 2013, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.H.; Duroux, M.; Gazerani, P. Serum MicroRNA Signatures in Migraineurs During Attacks and in Pain-Free Periods. Mol. Neurobiol. 2016, 53, 1494–1500. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. MiRBase: Tools for MicroRNA Genomics. Nucleic Acids Res. 2007, 36, D154–D158. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. The Biological Functions of MiRNAs: Lessons from in Vivo Studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef]
- Gazerani, P. Current Evidence on Potential Uses of MicroRNA Biomarkers for Migraine: From Diagnosis to Treatment. Mol. Diagn. Ther. 2019, 23, 681–694. [Google Scholar] [CrossRef]
- Gallelli, L.; Cione, E.; Caroleo, M.C.; Carotenuto, M.; Lagana, P.; Siniscalchi, A.; Guidetti, V. MicroRNAs to Monitor Pain-Migraine and Drug Treatment. MIRNA 2017, 6, 152–156. [Google Scholar] [CrossRef]
- Labruijere, S.; Stolk, L.; Verbiest, M.; de Vries, R.; Garrelds, I.M.; Eilers, P.H.C.; Danser, A.H.J.; Uitterlinden, A.G.; MaassenVanDenBrink, A. Methylation of Migraine-Related Genes in Different Tissues of the Rat. PLoS ONE 2014, 9, e87616. [Google Scholar] [CrossRef]
- Bainomugisa, C.K.; Sutherland, H.G.; Parker, R.; Mcrae, A.F.; Haupt, L.M.; Griffiths, L.R.; Heath, A.; Nelson, E.C.; Wright, M.J.; Hickie, I.B.; et al. Using Monozygotic Twins to Dissect Common Genes in Posttraumatic Stress Disorder and Migraine. Front. Neurosci. 2021, 15, 678350. [Google Scholar] [CrossRef] [PubMed]
- Gerring, Z.F.; McRae, A.F.; Montgomery, G.W.; Nyholt, D.R. Genome-Wide DNA Methylation Profiling in Whole Blood Reveals Epigenetic Signatures Associated with Migraine. BMC Genom. 2018, 19, 69. [Google Scholar] [CrossRef]
- Winsvold, B.S.; Palta, P.; Eising, E.; Page, C.M.; The International Headache Genetics Consortium; van den Maagdenberg, A.M.; Palotie, A.; Zwart, J.-A. Epigenetic DNA Methylation Changes Associated with Headache Chronification: A Retrospective Case-Control Study. Cephalalgia 2018, 38, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Pérez Pereda, S.; Toriello Suárez, M.; González Quintanilla, V.; Oterino, A. Methylation Analysis of NPTX2 and SH2D5 Genes in Chronic Migraine: A Case–Control Study. Cephalalgia Rep. 2020, 3, 251581632092359. [Google Scholar] [CrossRef]
- Terlizzi, R.; Bacalini, M.G.; Pirazzini, C.; Giannini, G.; Pierangeli, G.; Garagnani, P.; Franceschi, C.; Cevoli, S.; Cortelli, P. Epigenetic DNA Methylation Changes in Episodic and Chronic Migraine. Neurol. Sci. 2018, 39 (Suppl. S1), 67–68. [Google Scholar] [CrossRef]
- Kwiatkowska, K.M.; Bacalini, M.G.; Sala, C.; Kaziyama, H.; de Andrade, D.C.; Terlizzi, R.; Giannini, G.; Cevoli, S.; Pierangeli, G.; Cortelli, P.; et al. Analysis of Epigenetic Age Predictors in Pain-Related Conditions. Front. Public Health 2020, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Bertels, Z.; Singh, H.; Dripps, I.; Siegersma, K.; Tipton, A.F.; Witkowski, W.D.; Sheets, Z.; Shah, P.; Conway, C.; Mangutov, E.; et al. Neuronal Complexity Is Attenuated in Preclinical Models of Migraine and Restored by HDAC6 Inhibition. eLife 2021, 10, e63076. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, X.; Nauta, H.J.; Lin, Q.; Li, J.; Fang, L. JNK1 Regulates Histone Acetylation in Trigeminal Neurons Following Chemical Stimulation. Biochem. Biophys. Res. Commun. 2008, 376, 781–786. [Google Scholar] [CrossRef]
- Rubino, E.; Boschi, S.; Giorgio, E.; Pozzi, E.; Marcinnò, A.; Gallo, E.; Roveta, F.; Grassini, A.; Brusco, A.; Rainero, I. Analysis of the DNA Methylation Pattern of the Promoter Region of Calcitonin Gene-Related Peptide 1 Gene in Patients with Episodic Migraine: An Exploratory Case-Control Study. Neurobiol. Pain 2022, 11, 100089. [Google Scholar] [CrossRef]
- Wan, D.; Hou, L.; Zhang, X.; Han, X.; Chen, M.; Tang, W.; Liu, R.; Dong, Z.; Yu, S. DNA Methylation of RAMP1 Gene in Migraine: An Exploratory Analysis. J. Headache Pain 2015, 16, 90. [Google Scholar] [CrossRef]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Tumelero, E.; Icco, R.D.; Sances, G.; Allena, M.; Tassorelli, C. Peripheral Changes of Endocannabinoid System Components in Episodic and Chronic Migraine Patients: A Pilot Study. Cephalalgia 2021, 41, 185–196. [Google Scholar] [CrossRef]
- Wen, Q.; Wang, Y.; Pan, Q.; Tian, R.; Zhang, D.; Qin, G.; Zhou, J.; Chen, L. MicroRNA-155-5p Promotes Neuroinflammation and Central Sensitization via Inhibiting SIRT1 in a Nitroglycerin-Induced Chronic Migraine Mouse Model. J. Neuroinflamm. 2021, 18, 287. [Google Scholar] [CrossRef] [PubMed]
- Aczél, T.; Benczik, B.; Ágg, B.; Körtési, T.; Urbán, P.; Bauer, W.; Gyenesei, A.; Tuka, B.; Tajti, J.; Ferdinandy, P.; et al. Disease- and Headache-Specific MicroRNA Signatures and Their Predicted MRNA Targets in Peripheral Blood Mononuclear Cells in Migraineurs: Role of Inflammatory Signalling and Oxidative Stress. J. Headache Pain 2022, 23, 113. [Google Scholar] [CrossRef]
- Baksa, D.; Gonda, X.; Eszlari, N.; Petschner, P.; Acs, V.; Kalmar, L.; Deakin, J.F.W.; Bagdy, G.; Juhasz, G. Financial Stress Interacts with CLOCK Gene to Affect Migraine. Front. Behav. Neurosci. 2020, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chang, Y.; Chou, C.; Juan, C.; Lee, H.; Chen, L.; Wu, P.; Wang, Y.; Fuh, J.; Lirng, J.; et al. Circulating microRNAs Associated with Reversible Cerebral Vasoconstriction Syndrome. Ann. Neurol. 2021, 89, 459–473. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Chen, S.-P.; Liao, Y.-C.; Fuh, J.-L.; Wang, Y.-F.; Wang, S.-J. Elevated Circulating Endothelial-Specific MicroRNAs in Migraine Patients: A Pilot Study. Cephalalgia 2018, 38, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- De Icco, R.; Fiamingo, G.; Greco, R.; Bottiroli, S.; Demartini, C.; Zanaboni, A.M.; Allena, M.; Guaschino, E.; Martinelli, D.; Putortì, A.; et al. Neurophysiological and Biomolecular Effects of Erenumab in Chronic Migraine: An Open Label Study. Cephalalgia 2020, 40, 1336–1345. [Google Scholar] [CrossRef]
- Gallelli, L.; Cione, E.; Peltrone, F.; Siviglia, S.; Verano, A.; Chirchiglia, D.; Zampogna, S.; Guidetti, V.; Sammartino, L.; Montana, A.; et al. Hsa-MiR-34a-5p and Hsa-MiR-375 as Biomarkers for Monitoring the Effects of Drug Treatment for Migraine Pain in Children and Adolescents: A Pilot Study. JCM 2019, 8, 928. [Google Scholar] [CrossRef]
- Tafuri, E.; Santovito, D.; de Nardis, V.; Marcantonio, P.; Paganelli, C.; Affaitati, G.; Bucci, M.; Mezzetti, A.; Giamberardino, M.A.; Cipollone, F. MicroRNA Profiling in Migraine without Aura: Pilot Study. Ann. Med. 2015, 47, 468–473. [Google Scholar] [CrossRef]
- Zhai, Y.; Zhu, Y.-Y. MiR-30a Relieves Migraine by Degrading CALCA. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2022–2028. [Google Scholar] [CrossRef]
- Lin, J.; Shi, S.; Chen, Q.; Pan, Y. Differential Expression and Bioinformatic Analysis of the CircRNA Expression in Migraine Patients. BioMed Res. Int. 2020, 2020, 4710780. [Google Scholar] [CrossRef]
- Roos-Araujo, D.; Stuart, S.; Lea, R.A.; Haupt, L.M.; Griffiths, L.R. Epigenetics and Migraine; Complex Mitochondrial Interactions Contributing to Disease Susceptibility. Gene 2014, 543, 1–7. [Google Scholar] [CrossRef]
- Fila, M.; Chojnacki, C.; Chojnacki, J.; Blasiak, J. Is an “Epigenetic Diet” for Migraines Justified? The Case of Folate and DNA Methylation. Nutrients 2019, 11, 2763. [Google Scholar] [CrossRef]
- Miller, J.L.; Grant, P.A. The Role of DNA Methylation and Histone Modifications in Transcriptional Regulation in Humans. In Epigenetics: Development and Disease; Kundu, T.K., Ed.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2013; Volume 61, pp. 289–317. [Google Scholar] [CrossRef]
- Lyko, F. The DNA Methyltransferase Family: A Versatile Toolkit for Epigenetic Regulation. Nat. Rev. Genet. 2018, 19, 81–92. [Google Scholar] [CrossRef]
- Younesian, S.; Yousefi, A.-M.; Momeny, M.; Ghaffari, S.H.; Bashash, D. The DNA Methylation in Neurological Diseases. Cells 2022, 11, 3439. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Veland, N.; Lu, Y.; Hardikar, S.; Gaddis, S.; Zeng, Y.; Liu, B.; Estecio, M.R.; Takata, Y.; Lin, K.; Tomida, M.W.; et al. DNMT3L Facilitates DNA Methylation Partly by Maintaining DNMT3A Stability in Mouse Embryonic Stem Cells. Nucleic Acids Res. 2019, 47, 152–167. [Google Scholar] [CrossRef]
- Bronner, C.; Alhosin, M.; Hamiche, A.; Mousli, M. Coordinated Dialogue between UHRF1 and DNMT1 to Ensure Faithful Inheritance of Methylated DNA Patterns. Genes 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. ChemBioChem 2011, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Saravanaraman, P.; Selvam, M.; Ashok, C.; Srijyothi, L.; Baluchamy, S. De Novo Methyltransferases: Potential Players in Diseases and New Directions for Targeted Therapy. Biochimie 2020, 176, 85–102. [Google Scholar] [CrossRef]
- Illingworth, R.S.; Bird, A.P. CpG Islands—‘A Rough Guide’. FEBS Lett. 2009, 583, 1713–1720. [Google Scholar] [CrossRef]
- Amante, S.M.; Montibus, B.; Cowley, M.; Barkas, N.; Setiadi, J.; Saadeh, H.; Giemza, J.; Contreras-Castillo, S.; Fleischanderl, K.; Schulz, R.; et al. Transcription of Intragenic CpG Islands Influences Spatiotemporal Host Gene Pre-MRNA Processing. Nucleic Acids Res. 2020, 48, 8349–8359. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.A.; Montibus, B.; Oakey, R.J. Intragenic CpG Islands and Their Impact on Gene Regulation. Front. Cell Dev. Biol. 2022, 10, 832348. [Google Scholar] [CrossRef] [PubMed]
- Sarda, S.; Hannenhalli, S. Orphan CpG Islands as Alternative Promoters. Transcription 2018, 9, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, Y. TET-Mediated Active DNA Demethylation: Mechanism, Function and Beyond. Nat. Rev. Genet. 2017, 18, 517–534. [Google Scholar] [CrossRef]
- Ito, S.; D’Alessio, A.C.; Taranova, O.V.; Hong, K.; Sowers, L.C.; Zhang, Y. Role of Tet Proteins in 5mC to 5hmC Conversion, ES-Cell Self-Renewal and Inner Cell Mass Specification. Nature 2010, 466, 1129–1133. [Google Scholar] [CrossRef]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [PubMed]
- Maiti, A.; Drohat, A.C. Thymine DNA Glycosylase Can Rapidly Excise 5-Formylcytosine and 5-Carboxylcytosine. J. Biol. Chem. 2011, 286, 35334–35338. [Google Scholar] [CrossRef]
- Ross, S.E.; Bogdanovic, O. TET Enzymes, DNA Demethylation and Pluripotency. Biochem. Soc. Trans. 2019, 47, 875–885. [Google Scholar] [CrossRef]
- Khare, T.; Pai, S.; Koncevicius, K.; Pal, M.; Kriukiene, E.; Liutkeviciute, Z.; Irimia, M.; Jia, P.; Ptak, C.; Xia, M.; et al. 5-HmC in the Brain Is Abundant in Synaptic Genes and Shows Differences at the Exon-Intron Boundary. Nat. Struct. Mol. Biol. 2012, 19, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, A.; Freeman, W.M.; Jackson, J.; Wren, J.D.; Porter, H.; Richardson, A. The Role of DNA Methylation in Epigenetics of Aging. Pharmacol. Ther. 2019, 195, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Skvortsova, K.; Stirzaker, C.; Taberlay, P. The DNA Methylation Landscape in Cancer. Essays Biochem. 2019, 63, 797–811. [Google Scholar] [CrossRef]
- Locke, W.J.; Guanzon, D.; Ma, C.; Liew, Y.J.; Duesing, K.R.; Fung, K.Y.C.; Ross, J.P. DNA Methylation Cancer Biomarkers: Translation to the Clinic. Front. Genet. 2019, 10, 1150. [Google Scholar] [CrossRef]
- Pries, L.-K.; Gülöksüz, S.; Kenis, G. DNA Methylation in Schizophrenia. In Neuroepigenomics in Aging and Disease; Delgado-Morales, R., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 978, pp. 211–236. [Google Scholar] [CrossRef]
- Pishva, E.; Rutten, B.P.F.; Van Den Hove, D. DNA Methylation in Major Depressive Disorder. In Neuroepigenomics in Aging and Disease; Delgado-Morales, R., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 978, pp. 185–196. [Google Scholar] [CrossRef]
- Mastroeni, D.; Grover, A.; Delvaux, E.; Whiteside, C.; Coleman, P.D.; Rogers, J. Epigenetic Changes in Alzheimer’s Disease: Decrements in DNA Methylation. Neurobiol. Aging 2010, 31, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Wüllner, U.; Kaut, O.; de Boni, L.; Piston, D.; Schmitt, I. DNA Methylation in Parkinson’s Disease. J. Neurochem. 2016, 139, 108–120. [Google Scholar] [CrossRef]
- Goel, D.; Un Nisa, K.; Reza, M.I.; Rahman, Z.; Aamer, S. Aberrant DNA Methylation Pattern May Enhance Susceptibility to Migraine: A Novel Perspective. CNSNDDT 2019, 18, 504–515. [Google Scholar] [CrossRef]
- Daskalaki, M.G.; Tsatsanis, C.; Kampranis, S.C. Histone Methylation and Acetylation in Macrophages as a Mechanism for Regulation of Inflammatory Responses. J. Cell Physiol. 2018, 233, 6495–6507. [Google Scholar] [CrossRef] [PubMed]
- Anamika, K.; Krebs, A.R.; Thompson, J.; Poch, O.; Devys, D.; Tora, L. Lessons from Genome-Wide Studies: An Integrated Definition of the Coactivator Function of Histone Acetyl Transferases. Epigenetics Chromatin 2010, 3, 18. [Google Scholar] [CrossRef]
- de Ruijter, A.J.M.; van Gennip, A.H.; Caron, H.N.; Kemp, S.; van Kuilenburg, A.B.P. Histone Deacetylases (HDACs): Characterization of the Classical HDAC Family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [CrossRef]
- Wang, Z.; Zang, C.; Cui, K.; Schones, D.E.; Barski, A.; Peng, W.; Zhao, K. Genome-Wide Mapping of HATs and HDACs Reveals Distinct Functions in Active and Inactive Genes. Cell 2009, 138, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Dokmanovic, M.; Clarke, C.; Marks, P.A. Histone Deacetylase Inhibitors: Overview and Perspectives. Mol. Cancer Res. 2007, 5, 981–989. [Google Scholar] [CrossRef]
- Poziello, A.; Nebbioso, A.; Stunnenberg, H.G.; Martens, J.H.A.; Carafa, V.; Altucci, L. Recent Insights into Histone Acetyltransferase-1: Biological Function and Involvement in Pathogenesis. Epigenetics 2021, 16, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tian, Y.; Zhu, W.-G. The Roles of Histone Deacetylases and Their Inhibitors in Cancer Therapy. Front. Cell Dev. Biol. 2020, 8, 576946. [Google Scholar] [CrossRef] [PubMed]
- Kulthinee, S.; Yano, N.; Zhuang, S.; Wang, L.; Zhao, T.C. Critical Functions of Histone Deacetylases (HDACs) in Modulating Inflammation Associated with Cardiovascular Diseases. Pathophysiology 2022, 29, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Atlasi, Y.; Stunnenberg, H.G. The Interplay of Epigenetic Marks during Stem Cell Differentiation and Development. Nat. Rev. Genet. 2017, 18, 643–658. [Google Scholar] [CrossRef]
- Broide, R.S.; Redwine, J.M.; Aftahi, N.; Young, W.; Bloom, F.E.; Winrow, C.J. Distribution of Histone Deacetylases 1–11 in the Rat Brain. J. Mol. Neurosci. 2007, 31, 47–58. [Google Scholar] [CrossRef]
- Shivakumar, M.; Subbanna, S.; Joshi, V.; Basavarajappa, B.S. Postnatal Ethanol Exposure Activates HDAC-Mediated Histone Deacetylation, Impairs Synaptic Plasticity Gene Expression and Behavior in Mice. Int. J. Neuropsychopharmacol. 2020, 23, 324–338. [Google Scholar] [CrossRef]
- Bardai, F.H.; Price, V.; Zaayman, M.; Wang, L.; D’Mello, S.R. Histone Deacetylase-1 (HDAC1) Is a Molecular Switch between Neuronal Survival and Death. J. Biol. Chem. 2012, 287, 35444–35453. [Google Scholar] [CrossRef]
- Kumar, V.; Kundu, S.; Singh, A.; Singh, S. Understanding the Role of Histone Deacetylase and Their Inhibitors InNeurodegenerative Disorders: Current Targets and Future Perspective. CN 2022, 20, 158–178. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.L.; Davis, C.A.; Potthoff, M.J.; Haberland, M.; Fielitz, J.; Qi, X.; Hill, J.A.; Richardson, J.A.; Olson, E.N. Histone Deacetylases 1 and 2 Redundantly Regulate Cardiac Morphogenesis, Growth, and Contractility. Genes Dev. 2007, 21, 1790–1802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Cai, Y.; Liu, D.; Li, M.; Sha, Y.; Zhang, W.; Wang, K.; Gong, J.; Tang, N.; Huang, A.; et al. Pharmacological or Transcriptional Inhibition of Both HDAC1 and 2 Leads to Cell Cycle Blockage and Apoptosis via P21 Waf1/Cip1 and P19 INK4d Upregulation in Hepatocellular Carcinoma. Cell Prolif. 2018, 51, e12447. [Google Scholar] [CrossRef]
- Datta, M.; Staszewski, O.; Raschi, E.; Frosch, M.; Hagemeyer, N.; Tay, T.L.; Blank, T.; Kreutzfeldt, M.; Merkler, D.; Ziegler-Waldkirch, S.; et al. Histone Deacetylases 1 and 2 Regulate Microglia Function during Development, Homeostasis, and Neurodegeneration in a Context-Dependent Manner. Immunity 2018, 48, 514–529.e6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, M.; Bushong, E.; Phan, S.; Uytiepo, M.; Beutter, E.; Boemer, D.; Tsui, K.; Ellisman, M.; Maximov, A. Class IIa HDACs Regulate Learning and Memory through Dynamic Experience-Dependent Repression of Transcription. Nat. Commun. 2019, 10, 3469. [Google Scholar] [CrossRef]
- Sando, R.; Gounko, N.; Pieraut, S.; Liao, L.; Yates, J.; Maximov, A. HDAC4 Governs a Transcriptional Program Essential for Synaptic Plasticity and Memory. Cell 2012, 151, 821–834. [Google Scholar] [CrossRef]
- Daśko, M.; de Pascual-Teresa, B.; Ortín, I.; Ramos, A. HDAC Inhibitors: Innovative Strategies for Their Design and Applications. Molecules 2022, 27, 715. [Google Scholar] [CrossRef]
- Biersack, B.; Nitzsche, B.; Höpfner, M. HDAC Inhibitors with Potential to Overcome Drug Resistance in Castration-Resistant Prostate Cancer. CDR 2022, 5, 64. [Google Scholar] [CrossRef]
- Ziemka-Nalecz, M.; Jaworska, J.; Sypecka, J.; Zalewska, T. Histone Deacetylase Inhibitors: A Therapeutic Key in Neurological Disorders? J. Neuropathol. Exp. Neurol. 2018, 77, 855–870. [Google Scholar] [CrossRef]
- Urru, M.; Buonvicino, D.; Pistolesi, A.; Paccosi, S.; Chiarugi, A. Histone Deacetylase Inhibitors Counteract CGRP Signaling and Pronociceptive Sensitization in a Rat Model of Medication Overuse Headache. J. Pain 2022, 23, 1874–1884. [Google Scholar] [CrossRef]
- Gong, F.; Miller, K.M. Histone Methylation and the DNA Damage Response. Mutat. Res. Rev. Mutat. Res. 2019, 780, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S.; Subbanna, S. Histone Methylation Regulation in Neurodegenerative Disorders. Int. J. Mol. Sci. 2021, 22, 4654. [Google Scholar] [CrossRef] [PubMed]
- Alavian-Ghavanini, A.; Rüegg, J. Understanding Epigenetic Effects of Endocrine Disrupting Chemicals: From Mechanisms to Novel Test Methods. Basic Clin. Pharmacol. Toxicol. 2018, 122, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lei, L.; Laplante, D.P.; King, S. Prenatal Maternal Stress and Epigenetics: Review of the Human Research. Curr. Mol. Biol. Rep. 2016, 2, 16–25. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Gold, P.W. The Concepts of Stress and Stress System Disorders. Overview of Physical and Behavioral Homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Shimba, A.; Ikuta, K. Control of Immunity by Glucocorticoids in Health and Disease. Semin. Immunopathol. 2020, 42, 669–680. [Google Scholar] [CrossRef]
- Gassen, N.C.; Chrousos, G.P.; Binder, E.B.; Zannas, A.S. Life Stress, Glucocorticoid Signaling, and the Aging Epigenome: Implications for Aging-Related Diseases. Neurosci. Biobehav. Rev. 2017, 74, 356–365. [Google Scholar] [CrossRef]
- Mourtzi, N.; Sertedaki, A.; Charmandari, E. Glucocorticoid Signaling and Epigenetic Alterations in Stress-Related Disorders. Int. J. Mol. Sci. 2021, 22, 5964. [Google Scholar] [CrossRef]
- Gazerani, P. Migraine and Diet. Nutrients 2020, 12, 1658. [Google Scholar] [CrossRef]
- Migliore, L.; Coppedè, F. Gene–Environment Interactions in Alzheimer Disease: The Emerging Role of Epigenetics. Nat. Rev. Neurol. 2022, 18, 643–660. [Google Scholar] [CrossRef]
- Guo, J.; Riley, K.W.; Durham, T.; Margolis, A.E.; Wang, S.; Perera, F.; Herbstman, J.B. Association Studies of Environmental Exposures, DNA Methylation and Children’s Cognitive, Behavioral, and Mental Health Problems. Front. Genet. 2022, 13, 871820. [Google Scholar] [CrossRef]
- Cediel Ulloa, A.; Gliga, A.; Love, T.M.; Pineda, D.; Mruzek, D.W.; Watson, G.E.; Davidson, P.W.; Shamlaye, C.F.; Strain, J.J.; Myers, G.J.; et al. Prenatal Methylmercury Exposure and DNA Methylation in Seven-Year-Old Children in the Seychelles Child Development Study. Environ. Int. 2021, 147, 106321. [Google Scholar] [CrossRef]
- Smeester, L.; Fry, R.C. Long-Term Health Effects and Underlying Biological Mechanisms of Developmental Exposure to Arsenic. Curr. Environ. Health Rep. 2018, 5, 134–144. [Google Scholar] [CrossRef]
- Elkin, E.R.; Higgins, C.; Aung, M.T.; Bakulski, K.M. Metals Exposures and DNA Methylation: Current Evidence and Future Directions. Curr. Environ. Health Rep. 2022, 9, 673–696. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, L.; Nasuti, C.; Fedeli, D.; Galeazzi, R.; Laudadio, E.; Massaccesi, L.; López-Rodas, G.; Gabbianelli, R. Early Impairment of Epigenetic Pattern in Neurodegeneration: Additional Mechanisms behind Pyrethroid Toxicity. Exp. Gerontol. 2019, 124, 110629. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA Hypermethylation in Disease: Mechanisms and Clinical Relevance. Epigenetics 2019, 14, 1141–1163. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Liu, Y. DNA Methylation in Human Diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Weisenberger, D.J.; Liang, G.; Lenz, H.-J. DNA Methylation Aberrancies Delineate Clinically Distinct Subsets of Colorectal Cancer and Provide Novel Targets for Epigenetic Therapies. Oncogene 2018, 37, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Joosten, S.C.; Feng, Z.; de Ruijter, T.C.; Draht, M.X.; Melotte, V.; Smits, K.M.; Veeck, J.; Herman, J.G.; Van Neste, L.; et al. Analysis of DNA Methylation in Cancer: Location Revisited. Nat. Rev. Clin. Oncol. 2018, 15, 459–466. [Google Scholar] [CrossRef]
- Howell, K.J.; Kraiczy, J.; Nayak, K.M.; Gasparetto, M.; Ross, A.; Lee, C.; Mak, T.N.; Koo, B.-K.; Kumar, N.; Lawley, T.; et al. DNA Methylation and Transcription Patterns in Intestinal Epithelial Cells from Pediatric Patients with Inflammatory Bowel Diseases Differentiate Disease Subtypes and Associate with Outcome. Gastroenterology 2018, 154, 585–598. [Google Scholar] [CrossRef]
- Shirvani-Farsani, Z.; Maloum, Z.; Bagheri-Hosseinabadi, Z.; Vilor-Tejedor, N.; Sadeghi, I. DNA Methylation Signature as a Biomarker of Major Neuropsychiatric Disorders. J. Psychiatr. Res. 2021, 141, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Scher, I.A.; Stewart, F.W.; Ricci, A.J.; Lipton, B.R. Factors Associated with the Onset and Remission of Chronic Daily Headache in a Population-Based Study. Pain 2003, 106, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Mungoven, T.J.; Henderson, L.A.; Meylakh, N. Chronic Migraine Pathophysiology and Treatment: A Review of Current Perspectives. Front. Pain Res. 2021, 2, 705276. [Google Scholar] [CrossRef] [PubMed]
- Diener, H.-C.; Dodick, D.W.; Goadsby, P.J.; Lipton, R.B.; Olesen, J.; Silberstein, S.D. Chronic Migraine—Classification, Characteristics and Treatment. Nat. Rev. Neurol. 2012, 8, 162–171. [Google Scholar] [CrossRef]
- Natoli, J.; Manack, A.; Dean, B.; Butler, Q.; Turkel, C.; Stovner, L.; Lipton, R. Global Prevalence of Chronic Migraine: A Systematic Review. Cephalalgia 2010, 30, 599–609. [Google Scholar] [CrossRef]
- Guo, J.U.; Ma, D.K.; Mo, H.; Ball, M.P.; Jang, M.-H.; Bonaguidi, M.A.; Balazer, J.A.; Eaves, H.L.; Xie, B.; Ford, E.; et al. Neuronal Activity Modifies the DNA Methylation Landscape in the Adult Brain. Nat. Neurosci. 2011, 14, 1345–1351. [Google Scholar] [CrossRef]
- Horvath, S.; Raj, K. DNA Methylation-Based Biomarkers and the Epigenetic Clock Theory of Ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef]
- Horvath, S.; Ritz, B.R. Increased Epigenetic Age and Granulocyte Counts in the Blood of Parkinson’s Disease Patients. Aging 2015, 7, 1130–1142. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Bennett, D.A.; Horvath, S. Epigenetic Age of the Pre-Frontal Cortex Is Associated with Neuritic Plaques, Amyloid Load, and Alzheimer’s Disease Related Cognitive Functioning. Aging 2015, 7, 1198–1211. [Google Scholar] [CrossRef]
- Cruz-Almeida, Y.; Sinha, P.; Rani, A.; Huo, Z.; Fillingim, R.B.; Foster, T. Epigenetic Aging Is Associated with Clinical and Experimental Pain in Community-Dwelling Older Adults. Mol. Pain 2019, 15, 174480691987181. [Google Scholar] [CrossRef]
- Krishnan, H.R.; Sakharkar, A.J.; Teppen, T.L.; Berkel, T.D.M.; Pandey, S.C. The Epigenetic Landscape of Alcoholism. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 115, pp. 75–116. [Google Scholar] [CrossRef]
- Descalzi, G.; Ikegami, D.; Ushijima, T.; Nestler, E.J.; Zachariou, V.; Narita, M. Epigenetic Mechanisms of Chronic Pain. Trends Neurosci. 2015, 38, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Fernández, A.; Cabrero, J.R.; Serrador, J.M.; Sánchez-Madrid, F. HDAC6: A Key Regulator of Cytoskeleton, Cell Migration and Cell–Cell Interactions. Trends Cell Biol. 2008, 18, 291–297. [Google Scholar] [CrossRef] [PubMed]
- d’Ydewalle, C.; Krishnan, J.; Chiheb, D.M.; Van Damme, P.; Irobi, J.; Kozikowski, A.P.; Berghe, P.V.; Timmerman, V.; Robberecht, W.; Van Den Bosch, L. HDAC6 Inhibitors Reverse Axonal Loss in a Mouse Model of Mutant HSPB1–Induced Charcot-Marie-Tooth Disease. Nat. Med. 2011, 17, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Zeke, A.; Misheva, M.; Reményi, A.; Bogoyevitch, M.A. JNK Signaling: Regulation and Functions Based on Complex Protein-Protein Partnerships. Microbiol. Mol. Biol. Rev. 2016, 80, 793–835. [Google Scholar] [CrossRef] [PubMed]
- Ryder, K.; Lau, L.F.; Nathans, D. A Gene Activated by Growth Factors Is Related to the Oncogene V-Jun. Proc. Natl. Acad. Sci. USA 1988, 85, 1487–1491. [Google Scholar] [CrossRef] [PubMed]
- Herdegen, T.; Leah, J.D. Inducible and Constitutive Transcription Factors in the Mammalian Nervous System: Control of Gene Expression by Jun, Fos and Krox, and CREB/ATF Proteins. Brain Res. Rev. 1998, 28, 370–490. [Google Scholar] [CrossRef] [PubMed]
- Mielke, K.; Herdegen, T. JNK and P38 Stresskinases—Degenerative Effectors of Signal-Transduction-Cascades in the Nervous System. Prog. Neurobiol. 2000, 61, 45–60. [Google Scholar] [CrossRef]
- Wang, J.; Yun, F.; Sui, J.; Liang, W.; Shen, D.; Zhang, Q. HAT- and HDAC-Targeted Protein Acetylation in the Occurrence and Treatment of Epilepsy. Biomedicines 2022, 11, 88. [Google Scholar] [CrossRef]
- Edvinsson, L. Role of CGRP in Migraine. In Calcitonin Gene-Related Peptide (CGRP) Mechanisms; Brain, S.D., Geppetti, P., Eds.; Handbook of Experimental Pharmacology; Springer International Publishing: Cham, Switzerland, 2019; Volume 255, pp. 121–130. [Google Scholar] [CrossRef]
- Durham, P.L. Inhibition of Calcitonin Gene-Related Peptide Function: A Promising Strategy for Treating Migraine. Headache J. Head Face Pain 2008, 48, 1269–1275. [Google Scholar] [CrossRef]
- Rosenfeld, M.G.; Amara, S.G.; Evans, R.M. Alternative RNA Processing: Determining Neuronal Phenotype. Science 1984, 225, 1315–1320. [Google Scholar] [CrossRef]
- Tendl, K.A.; Schulz, S.M.; Mechtler, T.P.; Bohn, A.; Metz, T.; Greber-Platzer, S.; Kasper, D.C.; Herkner, K.R.; Item, C.B. DNA Methylation Pattern of CALCA in Preterm Neonates with Bacterial Sepsis as a Putative Epigenetic Biomarker. Epigenetics 2013, 8, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R. Understanding Migraine: Potential Role of Neurogenic Inflammation. Ann. Indian Acad. Neurol. 2016, 19, 175. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Warfvinge, K. Recognizing the Role of CGRP and CGRP Receptors in Migraine and Its Treatment. Cephalalgia 2019, 39, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L. Novel Migraine Therapy with Calcitonin Gene-Regulated Peptide Receptor Antagonists. Expert Opin. Ther. Targets 2007, 11, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.F. CGRP as a Neuropeptide in Migraine: Lessons from Mice: CGRP as a Neuropeptide in Migraine. Br. J. Clin. Pharmacol. 2015, 80, 403–414. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Edvinsson, L.; Ekman, R. Vasoactive Peptide Release in the Extracerebral Circulation of Humans during Migraine Headache. Ann. Neurol. 1990, 28, 183–187. [Google Scholar] [CrossRef]
- Cernuda-Morollon, E.; Larrosa, D.; Ramon, C.; Vega, J.; Martinez-Camblor, P.; Pascual, J. Interictal Increase of CGRP Levels in Peripheral Blood as a Biomarker for Chronic Migraine. Neurology 2013, 81, 1191–1196. [Google Scholar] [CrossRef]
- Edvinsson, L.; Grell, A.-S.; Warfvinge, K. Expression of the CGRP Family of Neuropeptides and Their Receptors in the Trigeminal Ganglion. J. Mol. Neurosci. 2020, 70, 930–944. [Google Scholar] [CrossRef]
- Barwell, J.; Wootten, D.; Simms, J.; Hay, D.L.; Poyner, D.R. RAMPs and CGRP Receptors. In RAMPs; Spielman, W.S., Parameswaran, N., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2012; Volume 744, pp. 13–24. [Google Scholar] [CrossRef]
- Gingell, J.J.; Hendrikse, E.R.; Hay, D.L. New Insights into the Regulation of CGRP-Family Receptors. Trends Pharmacol. Sci. 2019, 40, 71–83. [Google Scholar] [CrossRef]
- Marquez de Prado, B.; Hammond, D.L.; Russo, A.F. Genetic Enhancement of Calcitonin Gene-Related Peptide-Induced Central Sensitization to Mechanical Stimuli in Mice. J. Pain 2009, 10, 992–1000. [Google Scholar] [CrossRef]
- Recober, A.; Kaiser, E.A.; Kuburas, A.; Russo, A.F. Induction of Multiple Photophobic Behaviors in a Transgenic Mouse Sensitized to CGRP. Neuropharmacology 2010, 58, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Meccariello, R.; Santoro, A.; D’Angelo, S.; Morrone, R.; Fasano, S.; Viggiano, A.; Pierantoni, R. The Epigenetics of the Endocannabinoid System. Int. J. Mol. Sci. 2020, 21, 1113. [Google Scholar] [CrossRef] [PubMed]
- Lomazzo, E.; König, F.; Abassi, L.; Jelinek, R.; Lutz, B. Chronic Stress Leads to Epigenetic Dysregulation in the Neuropeptide-Y and Cannabinoid CB1 Receptor Genes in the Mouse Cingulate Cortex. Neuropharmacology 2017, 113, 301–313. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Wiley, J.W. The Role of the Endocannabinoid System in the Brain–Gut Axis. Gastroenterology 2016, 151, 252–266. [Google Scholar] [CrossRef]
- Gomes, T.M.; Dias Da Silva, D.; Carmo, H.; Carvalho, F.; Silva, J.P. Epigenetics and the Endocannabinoid System Signaling: An Intricate Interplay Modulating Neurodevelopment. Pharmacol. Res. 2020, 162, 105237. [Google Scholar] [CrossRef]
- D’Addario, C.; Di Francesco, A.; Arosio, B.; Gussago, C.; Dell’Osso, B.; Bari, M.; Galimberti, D.; Scarpini, E.; Altamura, A.C.; Mari, D.; et al. Epigenetic Regulation of Fatty Acid Amide Hydrolase in Alzheimer Disease. PLoS ONE 2012, 7, e39186. [Google Scholar] [CrossRef]
- Akerman, S.; Holland, P.R.; Lasalandra, M.P.; Goadsby, P.J. Endocannabinoids in the Brainstem Modulate Dural Trigeminovascular Nociceptive Traffic via CB1 and “Triptan” Receptors: Implications in Migraine. J. Neurosci. 2013, 33, 14869–14877. [Google Scholar] [CrossRef]
- Baron, E.P. Comprehensive Review of Medicinal Marijuana, Cannabinoids, and Therapeutic Implications in Medicine and Headache: What a Long Strange Trip It’s Been…. Headache J. Head Face Pain 2015, 55, 885–916. [Google Scholar] [CrossRef]
- Tassorelli, C.; Greco, R.; Silberstein, S.D. The Endocannabinoid System in Migraine: From Bench to Pharmacy and Back. Curr. Opin. Neurol. 2019, 32, 405–412. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the Expanded Endocannabinoid System in Neurological Disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Börner, C.; Martella, E.; Höllt, V.; Kraus, J. Regulation of Opioid and Cannabinoid Receptor Genes in Human Neuroblastoma and T Cells by the Epigenetic Modifiers Trichostatin A and 5-Aza-2′-Deoxycytidine. Neuroimmunomodulation 2012, 19, 180–186. [Google Scholar] [CrossRef] [PubMed]
- D’Addario, C.; Di Francesco, A.; Pucci, M.; Finazzi Agrò, A.; Maccarrone, M. Epigenetic Mechanisms and Endocannabinoid Signalling. FEBS J. 2013, 280, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Upham, B.L.; Rummel, A.M.; Carbone, J.M.; Trosko, J.E.; Ouyang, Y.; Crawford, R.B.; Kaminski, N.E. Cannabinoids Inhibit Gap Junctional Intercellular Communication and Activate ERK in a Rat Liver Epithelial Cell Line. Int. J. Cancer 2003, 104, 12–18. [Google Scholar] [CrossRef]
- Nagre, N.N.; Subbanna, S.; Shivakumar, M.; Psychoyos, D.; Basavarajappa, B.S. CB1-Receptor Knockout Neonatal Mice Are Protected against Ethanol-Induced Impairments of DNMT1, DNMT3A, and DNA Methylation. J. Neurochem. 2015, 132, 429–442. [Google Scholar] [CrossRef]
- Tomasiewicz, H.C.; Jacobs, M.M.; Wilkinson, M.B.; Wilson, S.P.; Nestler, E.J.; Hurd, Y.L. Proenkephalin Mediates the Enduring Effects of Adolescent Cannabis Exposure Associated with Adult Opiate Vulnerability. Biol. Psychiatry 2012, 72, 803–810. [Google Scholar] [CrossRef]
- Prini, P.; Rusconi, F.; Zamberletti, E.; Gabaglio, M.; Penna, F.; Fasano, M.; Battaglioli, E.; Parolaro, D.; Rubino, T. Adolescent THC Exposure in Female Rats Leads to Cognitive Deficits through a Mechanism Involving Chromatin Modifications in the Prefrontal Cortex. Jpn 2018, 43, 87–101. [Google Scholar] [CrossRef]
- Gerra, M.C.; Jayanthi, S.; Manfredini, M.; Walther, D.; Schroeder, J.; Phillips, K.A.; Cadet, J.L.; Donnini, C. Gene Variants and Educational Attainment in Cannabis Use: Mediating Role of DNA Methylation. Transl. Psychiatry 2018, 8, 23. [Google Scholar] [CrossRef]
- Schürks, M.; Rist, P.M.; Kurth, T. MTHFR 677C>T and ACE D/I Polymorphisms in Migraine: A Systematic Review and Meta-Analysis. Headache J. Head Face Pain 2010, 50, 588–599. [Google Scholar] [CrossRef]
- Hautakangas, H.; Winsvold, B.S.; Ruotsalainen, S.E.; Bjornsdottir, G.; Harder, A.V.E.; Kogelman, L.J.A.; Thomas, L.F.; Noordam, R.; Benner, C.; Gormley, P.; et al. Genome-Wide Analysis of 102,084 Migraine Cases Identifies 123 Risk Loci and Subtype-Specific Risk Alleles. Nat. Genet. 2022, 54, 152–160. [Google Scholar] [CrossRef]
- Hillyar, C.; Rallis, K.S.; Varghese, J. Advances in Epigenetic Cancer Therapeutics. Cureus 2020, 12, e11725. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA Profiling: Approaches and Considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Charles, A. The Pathophysiology of Migraine: Implications for Clinical Management. Lancet Neurol. 2018, 17, 174–182. [Google Scholar] [CrossRef]
- Ebahimzadeh, K.; Gholipour, M.; Samadian, M.; Taheri, M.; Ghafouri-Fard, S. A Comprehensive Review on the Role of Genetic Factors in the Pathogenesis of Migraine. J. Mol. Neurosci. 2021, 71, 1987–2006. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Rani, V.; Sengar, R.S. Biogenesis and Mechanisms of MicroRNA-mediated Gene Regulation. Biotechnol. Bioeng. 2022, 119, 685–692. [Google Scholar] [CrossRef]
- Chhabra, R. MiRNA and Methylation: A Multifaceted Liaison. ChemBioChem 2015, 16, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Piletič, K.; Kunej, T. MicroRNA Epigenetic Signatures in Human Disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef]
- Iwakawa, H.; Tomari, Y. Life of RISC: Formation, Action, and Degradation of RNA-Induced Silencing Complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef]
- Misir, S.; Wu, N.; Yang, B.B. Specific Expression and Functions of Circular RNAs. Cell Death Differ. 2022, 29, 481–491. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-Protein Interactions: Functions, Mechanisms, and Identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-Y.; Cai, Z.-R.; Liu, J.; Wang, D.-S.; Ju, H.-Q.; Xu, R.-H. Circular RNA: Metabolism, Functions and Interactions with Proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.C. Circular RNAs Act as MiRNA Sponges. In Circular RNAs; Xiao, J., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2018; Volume 1087, pp. 67–79. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Das, A.; Sinha, T.; Shyamal, S.; Panda, A.C. Emerging Role of Circular RNA–Protein Interactions. ncRNA 2021, 7, 48. [Google Scholar] [CrossRef]
- Shi, Y.; Jia, X.; Xu, J. The New Function of CircRNA: Translation. Clin. Transl. Oncol. 2020, 22, 2162–2169. [Google Scholar] [CrossRef]
- Schuch, J.B.; Genro, J.P.; Bastos, C.R.; Ghisleni, G.; Tovo-Rodrigues, L. The Role of CLOCK Gene in Psychiatric Disorders: Evidence from Human and Animal Research. Am. J. Med. Genet. 2018, 177, 181–198. [Google Scholar] [CrossRef]
- Dupras, C.; Saulnier, K.M.; Joly, Y. Epigenetics, Ethics, Law and Society: A Multidisciplinary Review of Descriptive, Instrumental, Dialectical and Reflexive Analyses. Soc. Stud. Sci. 2019, 49, 785–810. [Google Scholar] [CrossRef]
- Cerdeña, J.P. Epigenetic Citizenship and Political Claims-Making: The Ethics of Molecularizing Structural Racism. BioSocieties 2022. [Google Scholar] [CrossRef]
- Santaló, J.; Berdasco, M. Ethical Implications of Epigenetics in the Era of Personalized Medicine. Clin. Epigenet. 2022, 14, 44. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Study | Title | First Author | Sample | Ethnicity | Genes/Molecules Assessed | Findings |
|---|---|---|---|---|---|---|
| [24] | A high methylation level of a novel −284 bp CpG island in the RAMP1 gene promoter is potentially associated with migraine in women | Carvalho Estefânia | 104 females (54 migraineurs, 50 controls) | Portuguese | RAMP1 gene promotor methylation | 5 differently methylated CpG dinucleotides (−346, −334, −284, −276, and −234 1) were assessed; −284 CpG unit found to be significantly methylated in migraineurs |
| [25] | Epigenetic regulation of the calcitonin gene-related peptide gene in trigeminal glia | Ki-Youb Park | Rat and human model cell lines; primary cultures of rat TG 11 glia | - | DNA methylation and histone acetylation in the promotor region of the CALCA gene | CT and CGRP mRNAs were assessed; CpG island methylation and histone H3 acetylation at the 18 bp cell-specific enhancer correlated with CALCA gene expression |
| [41] | Methylation of migraine-related genes in different tissues of the rat | Labruijere, S. | Rat tissues (leukocytes, thoracic aorta, dura mater, TG, caudal nuclei); 395 healthy women | - | Comparison of DNA methylation of migraine-specific genes in different migraine-related tissues: Calca, Ramp1, Crcp, Calcrl, Usf2, Esr1, Gper, Nos3, Mthfr Comparison of DNA methylation of migraine-specific genes between rats and humans | Methylation of the Crcp, Calcrl, Esr1, and Nos3 genes is tissue-specific; methylation in leukocytes does not correlate with that in other tissues |
| [42] | Using monozygotic twins to dissect common genes in posttraumatic stress disorder and migraine | Charlotte K Bainomugisa | Total: 42 monozygote twins PTSD: 12 participants (10 males, 2 females) Migraineurs: 30 participants (14 males, 16 females) | Caucasian | Genome-wide DNA methylation levels assessment | Differently methylated genes ADCYAP1, AIM2, CRHR1, DBH, DOCK2, FKBP5, HTR3A, OXTR, RORA, WWC1, and TSNARE1 |
| [43] | Genome-wide DNA methylation profiling in whole blood reveals epigenetic signatures associated with migraine | Gerring Zachary F. | 67 migraineurs and 67 age- and sex = matched controls | Northern Europeans | Epigenome-wide analysis of differently methylated regions | No single methylation probe reached genome-wide significance |
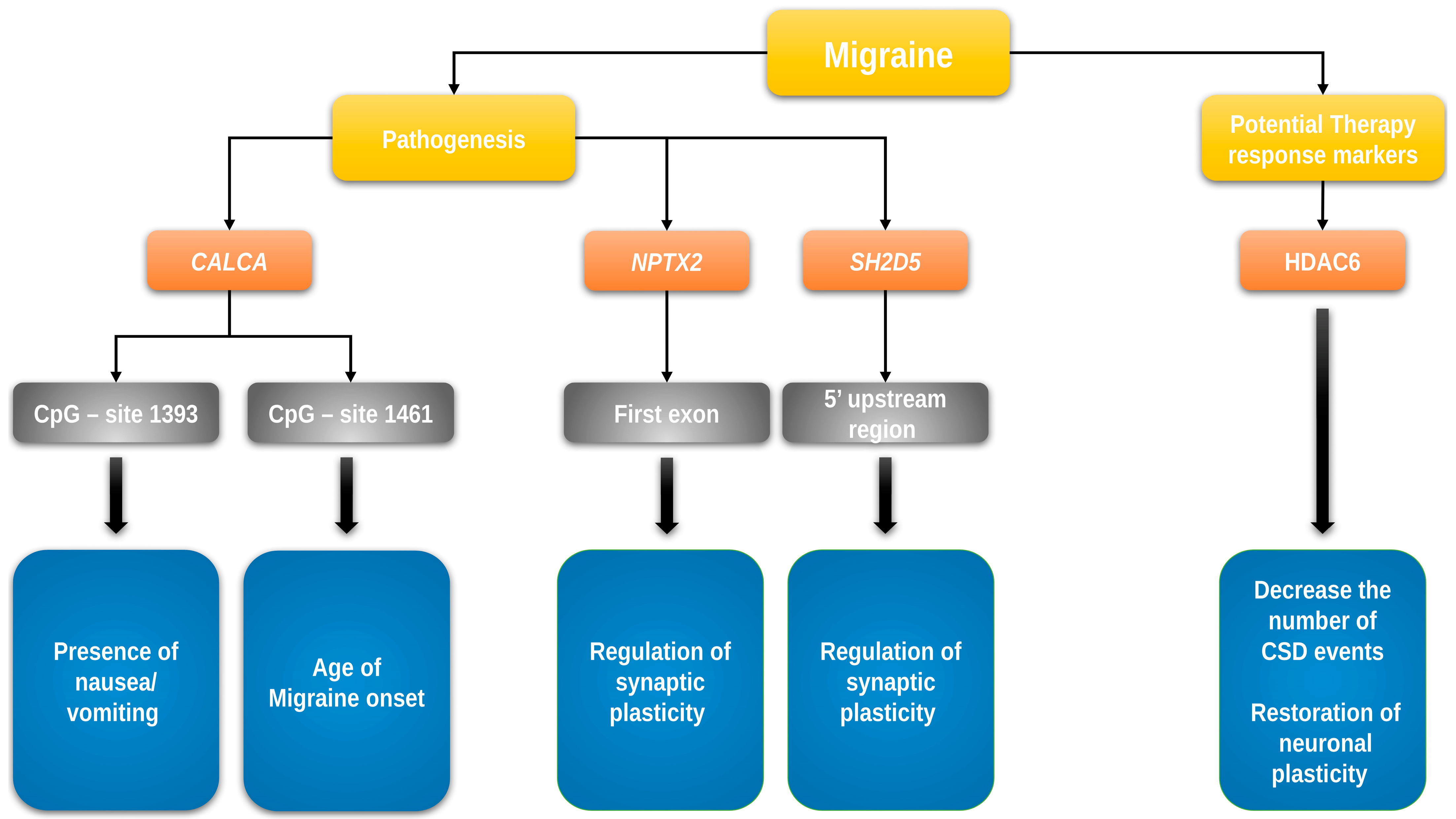
| [44] | Epigenetic DNA methylation changes associated with headache chronification: A retrospective case–control study | Winsvold, B.S. | 36 female headache patients 35 controls with episodic headache | Norwegian | Assessment of DNA methylation at 485,000 CpG sites | Possible association of 2 CpG sites in SH2D5 and NPTX2 genes with chronification of episodic headache |
| [45] | Methylation analysis of NPTX2 and SH2D5 genes in chronic migraine: A case–control study | Sara Perez Pereda | 109 CM patients; 98 EM patients; 98 controls | N/A | Assessment of methylation of two CpG sites related to NPTX2 7 and SH2D5 8 | No significant differences in methylation levels between CM, EM, and HC 9 in the first exon of the NPTX2 gene or the 50 upstream region of the SH2D5 gene |
| [46] | Epigenetic DNA methylation changes in episodic and chronic migraine | Terlizzi, R. | 18 MOH; 20 EM patients; 11 HC | N/A | Genome-wide DNA methylation levels association with headache chronification | Hypermethylation of chr10:76993892 island in COMT gene in MOH cases compared to HCs; hypomethylation of GpG site at chr12:110433797-110434205*Island in MOH cases compared to HCs (GIT2 gene), at chr19:44645494-44646069*N_Shore in MOH cases compared to HCs (ZNF234 gene), and at chr16:11348541-11350803*Island in MOH cases compared to HCs (SOCS1 gene) |
| [47] | Analysis of epigenetic age predictors in pain-related conditions | K. M. Kwiatkowska | 22 MOH 10 patients; 18 EM patients; 13 HC | Italian | Association between epigenetic age and chronic pain, by investigating first- and second-generation epigenetic clocks and DNA methylation surrogates of plasma proteins, blood cell counts, and telomere length in headache conditions | No significant difference in epigenetic age acceleration, DNA methylation surrogates comprised in GrimAg, and estimates of telomere length and blood cell counts between MOH cases and HCs or between EM cases and HCs. |
| [48] | Neuronal complexity is attenuated in preclinical models of migraine and restored by HDAC6 inhibition | Bertels Z. | Mouse models of migraine | - | HDAC6 inhibition and its effect in mouse models of migraine | HDAC6 inhibition restored neuronal plasticity and decreased the number of cortical spreading depression events |
| [49] | JNK1 regulates histone acetylation in trigeminal neurons following chemical stimulation | Wu, J. | TG neuron culture | - | The role of JNK/c-Jun cascade in the regulation of acetylation of H3 following chemical stimulation in TG neurons | Mustard oil stimulation activated the JNK/c-Jun pathway significantly by enhancing phospho-JNK1, phospho-c-Jun expression, and c-Jun activity, which were correlated with elevated acetylated H3 histone in TG neurons |
| [50] | Analysis of the DNA methylation pattern of the promoter region of calcitonin gene-related peptide 1 gene in patients with EM: An exploratory case–control study | Elisa Rubino | 22 EM patients; 20 controls | N/A | Evaluation of DNA methylation of CALCA gene in patients with EM | No differences in methylation of the 30 CpG sites at the distal region of CALCA were found in migraineurs compared with controls; no overall difference was found in the methylation level among these six detected CpG sites at the CALCA proximal promoter between migraineurs and controls; however, the DNA methylation profile in two CpG sites at the proximal promoter region of CALCA was lower in migraineurs when compared to HCs |
| [51] | DNA methylation of RAMP1 gene in migraine: An exploratory analysis | Dongjun Wan | 26 migraineurs, 25 matched controls | Chinese | DNA methylation levels at RAMP1 promoter region | No significant differences in 13 detected CpG sites or units at the RAMP1 promoter region in migraineurs compared with HC |
| [52] | Peripheral changes of endocannabinoid system components in episodic and chronic migraine patients: A pilot study | Rosaria Greco | 25 EM 2 patients (24 females) 26 CM 3-MO 4 (22 females) 24 Controls (18 females) | N/A 5 | DNA methylation changes in genes involved in ES 6 components | Methylation of CNR1, CNR2, DAGLA, FAAH, MGLL, NAPEPLD, and RPL19P12 was assessed; DNA methylation analysis did not show any significant differences between patients and controls with regard to the detected CpG sites at the promoter region levels of all the evaluated genes |
| Study | Title | First Author | Sample | Ethnicity | Genes/Molecules Assessed | Molecules That Are Expressed Differently |
|---|---|---|---|---|---|---|
| [26] | Plasma levels of CGRP and expression of specific microRNAs in blood cells of episodic and chronic migraine subjects: Toward the identification of a panel of peripheral biomarkers of migraine? | Rosaria Greco | 27 EM patients, 28 patients with CM-MO 6 | N/A | Evaluation of plasma levels of CGRP 7 and the expression of miR34a-5p and miR-382-5p in peripheral blood mononuclear cells | CGRP, miR-382-5p, and miR-34a-5p levels were significantly higher in CM-MO subjects when compared to EM patients |
| [34] | Serum MicroRNA signatures in migraineurs during attacks and in pain-free periods | Andersen, H. H. | 2 cohorts, 28 migraineurs, 20 HC 3 | N/A | Serum microRNA profiles of migraineurs during attacks and pain-free periods compared with healthy controls | miR-34a-5p miR-29c-5p, miR-1231, miR-328-3p, miR-382-5p*, miR-1207-5p, miR-1301-3p, miR-375, miR-26b-3p*, miR-4505, miR-424-5p, miR-320b, miR-320e, miR-629-3p, miR-1193, miR-142-5p, miR-188-5p, miR-1539, miR-373-3p, mar-1909-5p, miR-378e, miR-15a-3p, miR-324-3p, miR-34c-3p, miR-532-5p, miR-1183, miR-877-3p, miR-124-3p, miR-3120-3p, miR-1237-3p, miR-335-3p, miR-374c-5p |
| [53] | MicroRNA-155-5p promotes neuroinflammation and central sensitization via inhibiting SIRT1 in a nitroglycerin-induced chronic migraine mouse model | Wen Q. | Nitroglycerin-induced CM mouse model | - | miR-155-5p expression, SIRT1 protein levels; | Increased expression of miR-155-5p and decreased levels of SIRT1 in CM mouse model |
| [54] | Disease- and headache-specific microRNA signatures and their predicted mRNA targets in peripheral blood mononuclear cells in migraineurs: Role of inflammatory signaling and oxidative stress | Timea Aczél | 28 participants (16 with migraine, 12 healthy participants) | N/A 1 | miRNA of peripheral blood mononuclear cells | miRNAs: hsa-miR-5189-3p (2.59) 2, hsa-miR-96-5p (−2.4), hsa-miR-3613-5p (2.55), hsa-miR-99a-3p (2.37), hsa-miR-542-3p (2.4), hsa-miR-6803-3p (2.19), hsa-miR-6731-3p (−2.14), hsa-miR-577 (−2.17), hsa-miR-95-3p (−2.06), hsa-miR-556-3p (−2.18), hsa-miR-412-5p (−2.36), hsa-miR-5701 (−2.24), hsa-miR-3064-5p (2.1), hsa-miR-196a-5p (−2.55), hsa-miR-5189-5p (1.93), hsa-let-7i-3p (−1.82), hsa-miR-1277-5p (2.07), hsa-miR-29b-3p (−1.85), hsa-miR-4676-3p (1.87), hsa-miR-548j-3p (1.91), hsa-miR-1260b (1.78), hsa-miR-326 (1.62), hsa-miR-3174 (1.79), hsa-miR-210-3p (1.77), hsa-miR-32-5p (−1.65), hsa-miR-342-3p (−1.6), hsa-miR-3607-3p (−1.59), hsa-miR-142-5p (−1.54), hsa-miR-192-5p (−1.56), hsa-miR-155-5p (−1.43), and hsa-let-7 g-5p (−1.43) |
| [55] | Financial stress interacts with CLOCK gene to affect migraine | Baksa, D. | 2157 participants; 1503 females | N/A | Effect of rs10462028 of CLOCK gene on migraine | No direct effect of rs10462028 SNP on migraine; change in miRNA bindings in the 3′UTR of CLOCK gene in rs1801260 (G/A): miR-365b-3p G↓, miR-365a-3p G↓, and miR-664a-5p G↓, as well as in rs10462028 (A/G): miR-409-5p A↓ |
| [56] | Circulating microRNAs associated with reversible cerebral vasoconstriction syndrome | Chen S. | 30 patients with EM 4 during the ictal stage; 30 during the interictal stage; 30 age- and sex-matched HC | Taiwanese | Level of five miRNAs in 30 EM patients during the ictal stage and 30 EM patients during the interictal stage compared with 30 controls | MiR-130a-3p, miR-130b-3p, let-7a-5p, let-7b-5p, and let-7f-5p were investigated; the abundance of let-7a-5p, let-7b-5p, and let-7f-5p was significantly higher in ictal migraine patients compared to that of HC and interictal migraine patients |
| [57] | Elevated circulating endothelial-specific microRNAs in migraine patients: A pilot study | Cheng C. Y. | 30 migraineurs (20 females), 30 age- and sex-matched HC | Taiwanese | miR-155, miR-126, miR-21, and let-7g levels comparison in migraine patients with those in HC level of ICAM-1 (a marker of endothelial dysfunction) | miR-155, miR-126, and let-7g levels were 2–7-fold higher in the interictal migraine patients than in HCs; miR-155, miR-126, and let-7g were positively associated with the level of ICAM-1 in migraine patients |
| [58] | Neurophysiological and biomolecular effects of erenumab in chronic migraine: An open-label study | De Icco Roberto | 40 CM 5 patients | N/A | Evaluation of effects of erenumab treatment on the expression levels of miR34a-5p and miR-382-5p. | MiR-382-5p and miR-34a-5p levels were significantly lower after erenumab administration in the overall study population. After erenumab treatment, no significant differences between 30% responder and 30% nonresponder groups were found |
| [59] | Hsa-miR-34a-5p and hsa-miR-375 as biomarkers for monitoring the effects of drug treatment for migraine pain in children and adolescents: A pilot study | Galleli Luca | 24 migraine patients (50% females) without aura in 2 equal groups: treated; untreated 12 age- and sex-matched controls to the untreated group | N/A | Difference in saliva or in blood expression of hsa-miR-34a-5p and hsa-miR-375 between treated group and untreated group. | All enrolled migraineurs constitutively expressed both hsa-miR-34a-5p and hsa-miR-375, without difference with respect to age or gender; decrease of about 50% for hsa-miR-34a-5p and hsa-miR-375 in treated patients compared to untreated patients without difference with respect to age or gender |
| [60] | MicroRNA profiling in migraine without aura: A pilot study | E. Tafuri | 15 females suffering from migraine without aura; 13 HC | N/A | Validation of the following miRNAs by quantitative real-time polymerase chain reaction: miR-22, miR-26a, miR-26b, miR-27b, miR-29b, let-7b, miR-181a, miR-221, miR-30b, miR-30e | miR-27b was significantly upregulated; miR-181a, let-7b, and miR-22 were significantly downregulated |
| [61] | MiR-30a relieves migraine by degrading CALCA | Y. Zhai | N/A | Chinese | Relationship between miR-30a and CALCA in migraine | Expression levels of miR-30a in the peripheral blood of migraine patients were significantly reduced compared with HC |
| [62] | Differential expression and bioinformatic analysis of the circRNA expression in migraine patients | Jinghan Lin | 4 migraine patients, 3 controls | Chinese | Microarray analysis of circRNA of the plasma of migraine patients and healthy controls | 2039 circRNAs were detected in patient samples; 794 upregulated, 1245 downregulated relative to controls (fold change ≥ 1:5, p < 0:01); the top 10 upregulated circRNAs were hsa_circRNA_103670, hsa_circRNA_101833, hsa_circRNA_103809, hsa_circRNA_104855, hsa_circRNA_104761, hsa_circRNA_102610, hsa_circRNA_103444, hsa_circRNA_100257, hsa_circRNA_103149, and hsa_circRNA_100983; the top 10 downregulated circRNAs were hsa_circRNA_000367, hsa_circRNA_100236, hsa_circRNA_100790, hsa_circRNA_100789, hsa_circRNA_102413, hsa_circRNA_103689, hsa_circRNA_101784, hsa_circRNA_104950, hsa_circRNA_103846, and hsa_circRNA_101698 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zobdeh, F.; Eremenko, I.I.; Akan, M.A.; Tarasov, V.V.; Chubarev, V.N.; Schiöth, H.B.; Mwinyi, J. The Epigenetics of Migraine. Int. J. Mol. Sci. 2023, 24, 9127. https://doi.org/10.3390/ijms24119127
Zobdeh F, Eremenko II, Akan MA, Tarasov VV, Chubarev VN, Schiöth HB, Mwinyi J. The Epigenetics of Migraine. International Journal of Molecular Sciences. 2023; 24(11):9127. https://doi.org/10.3390/ijms24119127
Chicago/Turabian StyleZobdeh, Farzin, Ivan I. Eremenko, Mikail A. Akan, Vadim V. Tarasov, Vladimir N. Chubarev, Helgi B. Schiöth, and Jessica Mwinyi. 2023. "The Epigenetics of Migraine" International Journal of Molecular Sciences 24, no. 11: 9127. https://doi.org/10.3390/ijms24119127
APA StyleZobdeh, F., Eremenko, I. I., Akan, M. A., Tarasov, V. V., Chubarev, V. N., Schiöth, H. B., & Mwinyi, J. (2023). The Epigenetics of Migraine. International Journal of Molecular Sciences, 24(11), 9127. https://doi.org/10.3390/ijms24119127