
NGS Data Repurposing Allows Detection of tRNA Fragments as Gastric Cancer Biomarkers in Patient-Derived Extracellular Vesicles

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
2.1. tRFs Are Highly Expressed in GC Tumors
2.2. tRF Expression in GC Cell Lines and Derived EVs
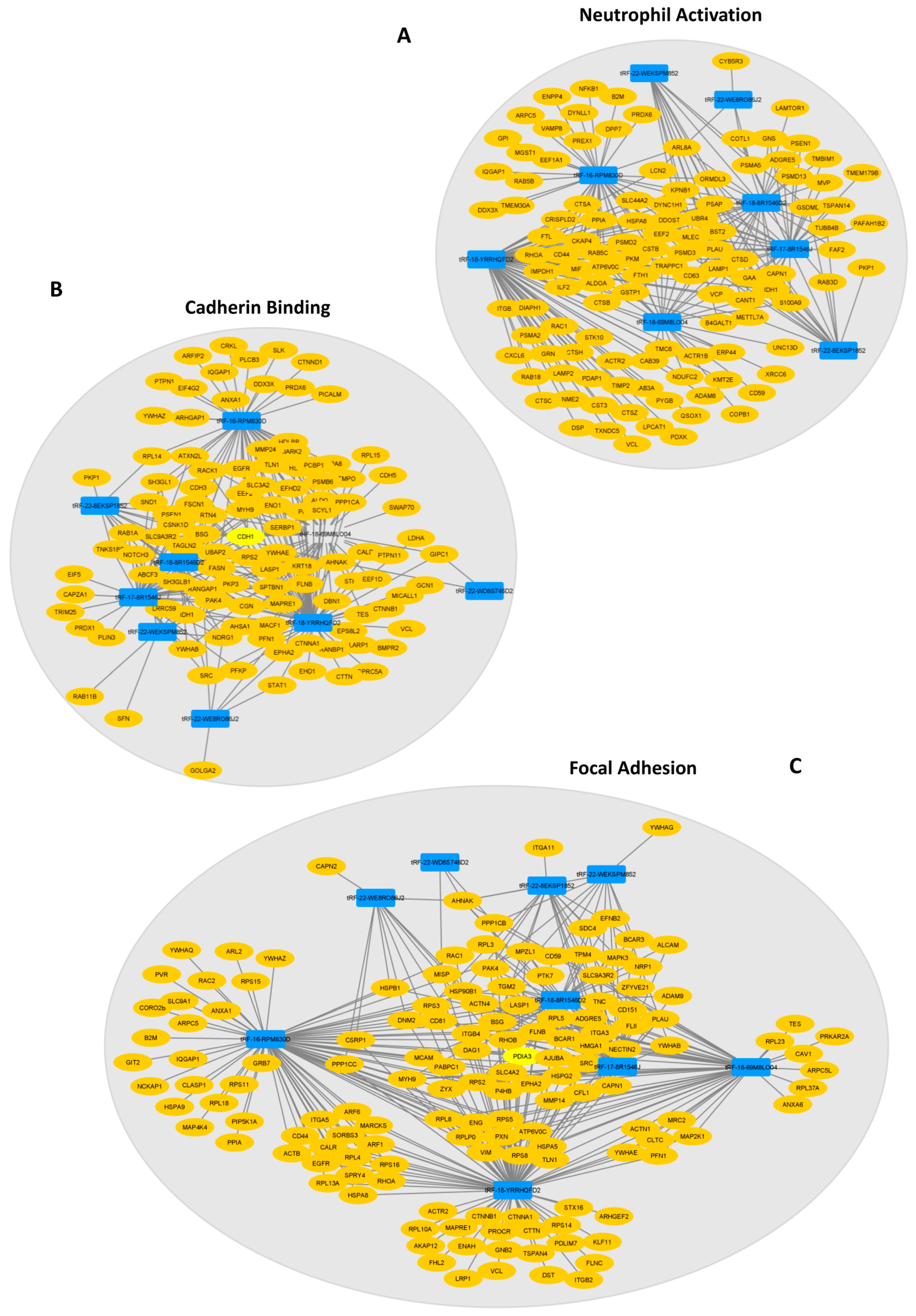
2.3. DE-tRFs Are Predicted to Modulate Immune Response and Cell Adhesion
2.4. Nine DE-tRFs Are Also Present in Patient-Derived EVs
3. Discussion
4. Materials and Methods
4.1. tRF Sequencing Data Collection and Pre-Processing
4.1.1. TCGA
4.1.2. S. Rocha et al.
4.1.3. GC Patients
Blood Sample Collection from Gastric Cancer (GC) Patients
EV Isolation and Characterization from Plasma of GC Patients
RNA Extraction from Human Plasma EVs (GC)
Small RNA Library Preparation and Sequencing from Human Plasma EV-sRNA (GC)
Pre-Processing of Human Plasma EV-sRNA Sequencing Data (GC)
4.2. tRF Expression Estimation
4.2.1. TCGA
4.2.2. GC Study
4.2.3. GC Patient EVs
4.3. Target Prediction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zang, L. The Effectiveness of Gastrectomy with Chemoradiotherapy Among Stage IV Gastric Adenocarcinoma: A Population-Based Analysis. Front. Oncol. 2020, 10, 630. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Sagaert, X.; Topal, B.; Haustermans, K.; Prenen, H. Gastric cancer. Lancet 2016, 388, 2654–2664. [Google Scholar] [CrossRef] [PubMed]
- Pasechnikov, V.; Chukov, S.; Fedorov, E.; Kikuste, I.; Leja, M. Gastric cancer: Prevention, screening and early diagnosis. World J. Gastroenterol. 2014, 20, 13842–13862. [Google Scholar] [CrossRef]
- Necula, L.; Matei, L.; Dragu, D.; Neagu, A.I.; Mambet, C.; Nedeianu, S.; Bleotu, C.; Diaconu, C.C.; Chivu-Economescu, M. Recent advances in gastric cancer early diagnosis. World J. Gastroenterol. 2019, 25, 2029–2044. [Google Scholar] [CrossRef]
- Wu, J.; Li, G.; Wang, Z.; Yao, Y.; Chen, R.; Pu, X.; Wang, J. Circulating MicroRNA-21 Is a Potential Diagnostic Biomarker in Gastric Cancer. Dis. Markers 2015, 2015, 435656. [Google Scholar] [CrossRef]
- Huang, Z.; Zhu, D.; Wu, L.; He, M.; Zhou, X.; Zhang, L.; Zhang, H.; Wang, W.; Zhu, J.; Cheng, W.; et al. Six Serum-Based miRNAs as Potential Diagnostic Biomarkers for Gastric Cancer. Cancer Epidemiol. Biomarkers Prev. 2017, 26, 188–196. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef]
- Rocha, S.; Carvalho, J.; Oliveira, P.; Voglstaetter, M.; Schvartz, D.; Thomsen, A.R.; Walter, N.; Khanduri, R.; Sanchez, J.-C.; Keller, A.; et al. 3D Cellular Architecture Affects MicroRNA and Protein Cargo of Extracellular Vesicles. Adv. Sci. 2019, 6, 1800948. [Google Scholar] [CrossRef]
- Millan, C.; Prause, L.; Vallmajo-Martin, Q.; Hensky, N.; Eberli, D. Extracellular Vesicles from 3D Engineered Microtissues Harbor Disease-Related Cargo Absent in EVs from 2D Cultures. Adv. Healthc. Mater. 2021, 11, 2002067. [Google Scholar] [CrossRef]
- Chiou, N.-T.; Kageyama, R.; Ansel, K.M. Selective Export into Extracellular Vesicles and Function of tRNA Fragments during T Cell Activation. Cell Rep. 2018, 25, 3356–3370.e4. [Google Scholar] [CrossRef] [PubMed]
- Shurtleff, M.J.; Yao, J.; Qin, Y.; Nottingham, R.M.; Temoche-Diaz, M.M.; Schekman, R.; Lambowitz, A.M. Broad role for YBX1 in defining the small noncoding RNA composition of exosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E8987–E8995. [Google Scholar] [CrossRef]
- Baglio, S.R.; Rooijers, K.; Koppers-Lalic, D.; Verweij, F.J.; Pérez Lanzón, M.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.M.; Baldini, N.; et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 2015, 6, 127. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zeringer, E.; Barta, T.; Schageman, J.; Cheng, A.; Vlassov, A. Analysis of the RNA content of the exosomes derived from blood serum and urine and its potential as biomarkers. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130502. [Google Scholar] [CrossRef]
- Santos, M.; Pereira, P.M.; Varanda, A.S.; Carvalho, J.; Azevedo, M.; Mateus, D.D.; Mendes, N.; Oliveira, P.; Trindade, F.; Pinto, M.T.; et al. Codon misreading tRNAs promote tumor growth in mice. RNA Biol. 2018, 15, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, H.; Nguyen, H.C.B.; Zhang, S.; Dill, B.D.; Molina, H.; Tavazoie, S.F. Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression. Cell 2016, 165, 1416–1427. [Google Scholar] [CrossRef]
- Krishnan, P.; Sunita, G.; Wang, B.; Heyns, M.; Li, D.; Mackey, J.R.; Kovalchuk, O.; Damaraju, S. Genome-wide profiling of transfer RNAs and their role as novel prognostic markers for breast cancer. Sci. Rep. 2016, 6, 32843. [Google Scholar] [CrossRef]
- Santos, M.; Fidalgo, A.; Varanda, A.S.; Soares, A.R.; Almeida, G.M.; Martins, D.; Mendes, N.; Oliveira, C.; Santos, M.A.S. Upregulation of tRNA-Ser-AGA-2-1 Promotes Malignant Behavior in Normal Bronchial Cells. Front. Mol. Biosci. 2022, 9, 809985. [Google Scholar] [CrossRef]
- Santos, M.; Fidalgo, A.; Varanda, A.S.; Oliveira, C.; Santos, M.A.S. tRNA Deregulation and Its Consequences in Cancer. Trends Mol. Med. 2019, 25, 853–865. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, H.; He, J.; Gao, W. Mechanisms of tRNA-derived fragments and tRNA halves in cancer treatment resistance. Biomark. Res. 2020, 8, 52. [Google Scholar] [CrossRef]
- Thompson, D.M.; Lu, C.; Green, P.J.; Parker, R. tRNA cleavage is a conserved response to oxidative stress in eukaryotes. RNA 2008, 14, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Ivanov, P.; Hu, G.-F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA-Related Fragments (tRFs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef]
- Hogg, M.C.; Raoof, R.; El Naggar, H.; Monsefi, N.; Delanty, N.; O’Brien, D.F.; Bauer, S.; Rosenow, F.; Henshall, D.C.; Prehn, J.H.M. Elevation of plasma tRNA fragments precedes seizures in human epilepsy. J. Clin. Investig. 2019, 129, 2946–2951. [Google Scholar] [CrossRef] [PubMed]
- Jacovetti, C.; Bayazit, M.B.; Regazzi, R. Emerging Classes of Small Non-Coding RNAs with Potential Implications in Diabetes and Associated Metabolic Disorders. Front. Endocrinol. 2021, 12, 503. [Google Scholar] [CrossRef]
- Yu, M.; Lu, B.; Zhang, J.; Ding, J.; Liu, P.; Lu, Y. tRNA-derived RNA fragments in cancer: Current status and future perspectives. J. Hematol. Oncol. 2020, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, J.; Gong, Y.; Wu, Q.; Tan, S.; Sun, D.; Xu, X.; Zuo, Y.; Zhao, Y.; Wei, Y.-Q.; et al. Exosomal tRNA-derived small RNA as a promising biomarker for cancer diagnosis. Mol. Cancer 2019, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Koi, Y.; Tsutani, Y.; Nishiyama, Y.; Ueda, D.; Ibuki, Y.; Sasada, S.; Akita, T.; Masumoto, N.; Kadoya, T.; Yamamoto, Y.; et al. Predicting the presence of breast cancer using circulating small RNAs, including those in the extracellular vesicles. Cancer Sci. 2020, 111, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Fan, X.; He, X.; Chen, S.; Huang, W.; Gao, J.; Huang, Y.; Wang, H. Comprehensively Identifying the Key tRNA-Derived Fragments and Investigating Their Function in Gastric Cancer Processes. OncoTargets Ther. 2020, 13, 10931–10943. [Google Scholar] [CrossRef]
- Xu, W.; Zhou, B.; Wang, J.; Tang, L.; Hu, Q.; Wang, J.; Chen, H.; Zheng, J.; Yan, F.; Chen, H. tRNA-Derived Fragment tRF-Glu-TTC-027 Regulates the Progression of Gastric Carcinoma via MAPK Signaling Pathway. Front. Oncol. 2021, 11, 3310. [Google Scholar] [CrossRef]
- Tong, L.; Zhang, W.; Qu, B.; Zhang, F.; Wu, Z.; Shi, J.; Chen, X.; Song, Y.; Wang, Z. The tRNA-Derived Fragment-3017A Promotes Metastasis by Inhibiting NELL2 in Human Gastric Cancer. Front. Oncol. 2021, 10, 3468. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Ma, S.; Liang, B.; Ju, S. Serum hsa_tsr016141 as a Kind of tRNA-Derived Fragments Is a Novel Biomarker in Gastric Cancer. Front. Oncol. 2021, 11, 1639. [Google Scholar] [CrossRef] [PubMed]
- Rasnic, R.; Brandes, N.; Zuk, O.; Linial, M. Substantial batch effects in TCGA exome sequences undermine pan-cancer analysis of germline variants. BMC Cancer 2019, 19, 783. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Martí, E. Toward an Understanding of Extracellular tRNA Biology. Front. Mol. Biosci. 2021, 8, 662620. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T.; Wada, R.; Kure, S.; Ishino, K.; Kudo, M.; Ohashi, R.; Fujita, I.; Uchida, E.; Yoshida, H.; Naito, Z. Expression of protein disulfide isomerase A3 and its clinicopathological association in gastric cancer. Oncol. Rep. 2019, 41, 2265–2272. [Google Scholar] [CrossRef]
- Sun, X.; Yang, J.; Yu, M.; Yao, D.; Zhou, L.; Li, X.; Qiu, Q.; Lin, W.; Lu, B.; Chen, E.; et al. Global identification and characterization of tRNA-derived RNA fragment landscapes across human cancers. NAR Cancer 2020, 2, zcaa031. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Chen, Y.P.; Lin, J.-Y. Increased expression of angiogenin in gastric carcinoma in correlation with tumor angiogenesis and proliferation. World J. Gastroenterol. 2006, 12, 5135–5139. [Google Scholar] [CrossRef]
- Lengyel, C.G.; Hussain, S.; Trapani, D.; El Bairi, K.; Altuna, S.C.; Seeber, A.; Odhiambo, A.; Habeeb, B.S.; Seid, F. The Emerging Role of Liquid Biopsy in Gastric Cancer. J. Clin. Med. 2021, 10, 2108. [Google Scholar] [CrossRef]
- Kusuma, G.D.; Li, A.; Zhu, D.; McDonald, H.; Inocencio, I.M.; Chambers, D.C.; Sinclair, K.; Fang, H.; Greening, D.W.; Frith, J.E.; et al. Effect of 2D and 3D Culture Microenvironments on Mesenchymal Stem Cell-Derived Extracellular Vesicles Potencies. Front. Cell Dev. Biol. 2022, 10, 819726. [Google Scholar] [CrossRef]
- Thippabhotla, S.; Zhong, C.; He, M. 3D cell culture stimulates the secretion of in vivo like extracellular vesicles. Sci. Rep. 2019, 9, 13012. [Google Scholar] [CrossRef]
- Veziroglu, E.M.; Mias, G.I. Characterizing Extracellular Vesicles and Their Diverse RNA Contents. Front. Genet. 2020, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Jean-Toussaint, R.; Sacan, A.; Ajit, S.K. Differential RNA packaging into small extracellular vesicles by neurons and astrocytes. Cell Commun. Signal. 2021, 19, 75. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514. [Google Scholar] [CrossRef] [PubMed]
- Haga, H.; Yan, I.K.; Takahashi, K.; Wood, J.; Zubair, A.; Patel, T. Tumour cell–derived extracellular vesicles interact with mesenchymal stem cells to modulate the microenvironment and enhance cholangiocarcinoma growth. J. Extracell. Vesicles 2015, 4, 24900. [Google Scholar] [CrossRef]
- Zhang, D.X.; Vu, L.T.; Ismail, N.N.; Le, M.T.N.; Grimson, A. Landscape of extracellular vesicles in the tumour microenvironment: Interactions with stromal cells and with non-cell components, and impacts on metabolic reprogramming, horizontal transfer of neoplastic traits, and the emergence of therapeutic resistance. Semin. Cancer Biol. 2021, 74, 24–44. [Google Scholar] [CrossRef]
- Yamada, N.; Kuranaga, Y.; Kumazaki, M.; Shinohara, H.; Taniguchi, K.; Akao, Y. Colorectal cancer cell-derived extracellular vesicles induce phenotypic alteration of T cells into tumor-growth supporting cells with transforming growth factor-β1-mediated suppression. Oncotarget 2016, 7, 27033–27043. [Google Scholar] [CrossRef]
- Fu, H.; Ma, Y.; Yang, M.; Zhang, C.; Huang, H.; Xia, Y.; Lu, L.; Jin, W.; Cui, D. Persisting and Increasing Neutrophil Infiltration Associates with Gastric Carcinogenesis and E-cadherin Downregulation. Sci. Rep. 2016, 6, 29762. [Google Scholar] [CrossRef]
- Mao, D.; Xu, R.; Chen, H.; Chen, X.; Li, D.; Song, S.; He, Y.; Wei, Z.; Zhang, C. Cross-Talk of Focal Adhesion-Related Gene Defines Prognosis and the Immune Microenvironment in Gastric Cancer. Front. Cell Dev. Biol. 2021, 9, 716461. [Google Scholar] [CrossRef]
- Lee, Y.; Finch-Edmondson, M.; Cognart, H.; Zhu, B.; Song, H.; Low, B.C.; Sudol, M. Common and Unique Transcription Signatures of YAP and TAZ in Gastric Cancer Cells. Cancers 2020, 12, 3667. [Google Scholar] [CrossRef]
- Andreuzzi, E.; Capuano, A.; Poletto, E.; Pivetta, E.; Fejza, A.; Favero, A.; Doliana, R.; Cannizzaro, R.; Spessotto, P.; Mongiat, M. Role of Extracellular Matrix in Gastrointestinal Cancer-Associated Angiogenesis. Int. J. Mol. Sci. 2020, 21, 3686. [Google Scholar] [CrossRef]
- Oliveira, C.; Pinheiro, H.; Figueiredo, J.; Seruca, R.; Carneiro, F. Familial gastric cancer: Genetic susceptibility, pathology, and implications for management. Lancet Oncol. 2015, 16, e60–e70. [Google Scholar] [CrossRef] [PubMed]
- Tischler, G.; Leonard, S. biobambam: Tools for read pair collation based algorithms on BAM files. Source Code Biol. Med. 2014, 9, 13. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Andrews, S.; Krueger, F.; Seconds-Pichon, A.; Biggins, F.; Wingett, S.; Fast, Q.C. Babraham Bioinformatics [Internet]. Volume 1, B.I. 2015. p. 1. A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/%0Ahttp://www.bioinformatics.bbsrc.ac.uk/projects/fastqc (accessed on 20 November 2020).
- Loher, P.; Telonis, A.G.; Rigoutsos, I. MINTmap: Fast and exhaustive profiling of nuclear and mitochondrial tRNA fragments from short RNA-seq data. Sci. Rep. 2017, 7, 41184. [Google Scholar] [CrossRef]
- Meseguer, S.; Navarro-González, C.; Panadero, J.; Villarroya, M.; Boutoual, R.; Sánchez-Alcázar, J.A.; Armengod, M.-E. The MELAS mutation m.3243A>G alters the expression of mitochondrial tRNA fragments. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1433–1449. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. gplots: Various R Programming Tools for Plotting Data. R Package Version 3.0.1.1. 2019. Available online: http://CRAN.R-project.org/package=gplots (accessed on 7 April 2021).
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, H.; Cui, Q.; Zhou, Y. tRFTar: Prediction of tRF-target gene interactions via systemic re-analysis of Argonaute CLIP-seq datasets. Methods 2021, 187, 57–67. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Inter J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | tRF_Type | Sequence | Exclusivity | Anticodon | logFC | logCPM | LR | Pvalue | FDR |
|---|---|---|---|---|---|---|---|---|---|
| tRF-23-YJE76INB0J | i-tRF | TTAGCACTCTGGACTCTGAATCC | UNIQUE | GlnCTG | −3.47878163 | 10.6598912 | 118.981005 | 1.06 × 10−27 | 1.04 × 10−24 |
| tRF-22-91PJB7MNK | i-tRF | TGGCCGCAGCAACCTCGGTTCG | UNIQUE | HisGTG | −4.01790481 | 9.10726781 | 118.346426 | 1.46 × 10−27 | 1.04 × 10−24 |
| tRF-22-WB8US5652 | 3′-tRF | TCGAATCCGAGTCACGGCACCA | UNIQUE | HisGTG | −2.19175055 | 8.94123967 | 116.35393 | 3.98 × 10−27 | 1.90 × 10−24 |
| tRF-21-91PJB7MND | i-tRF | TGGCCGCAGCAACCTCGGTTC | UNIQUE | HisGTG | −3.54575799 | 7.36208523 | 100.300282 | 1.31 × 10−23 | 4.69 × 10−21 |
| tRF-22-VF4YO9XEJ | i-tRF | TAGCACTCTGGACTCTGAATCC | UNIQUE | GlnCTG | −3.21016875 | 10.1858067 | 99.1946855 | 2.29 × 10−23 | 6.55 × 10−21 |
| tRF-24-SWRYVMMVHX | i-tRF | GTCGTGGTTGTAGTCCGTGCGAGA | MT | GluTTC | 6.01156036 | 10.0232838 | 93.2423782 | 4.63 × 10−22 | 1.10 × 10−19 |
| tRF-21-WB8US565D | 3′-tRF | TCGAATCCGAGTCACGGCACC | UNIQUE | HisGTG | −2.15755385 | 7.62862441 | 87.2918519 | 9.36 × 10−21 | 1.92 × 10−18 |
| tRF-23-91PJB7MNDL | i-tRF | TGGCCGCAGCAACCTCGGTTCGA | UNIQUE | HisGTG | −4.27131139 | 7.79586266 | 82.6891628 | 9.60 × 10−20 | 1.72 × 10−17 |
| tRF-23-VF4YO9XED2 | i-tRF | TAGCACTCTGGACTCTGAATCCA | UNIQUE | GlnCTG | −2.77409913 | 9.43587704 | 77.8958834 | 1.09 × 10−18 | 1.73 × 10−16 |
| tRF-21-EXEY0VWUD | 3′-tRF | ACTTAACTTGACCGCTCTGAC | MT | ValTAC | 3.47697974 | 12.9374387 | 76.9701494 | 1.74 × 10−18 | 2.49 × 10−16 |
| tRF-24-8DYDZDL9JR | 3′-tRF | TCAACTTAACTTGACCGCTCTGAC | MT | ValTAC | 3.34575605 | 9.93002784 | 74.5263922 | 5.98 × 10−18 | 7.79 × 10−16 |
| tRF-22-8B8SOUPR2 | 3′-tRF | TCAAATCCCGGACGAGCCCCCA | AMBIGUOUS | ProAGG | −1.86693881 | 9.09960168 | 73.4251504 | 1.05 × 10−17 | 1.25 × 10−15 |
| tRF-20-NONU3IND | 3′-tRF | CTTAACTTGACCGCTCTGAC | MT | ValTAC | 3.10311975 | 10.6738238 | 73.1309279 | 1.21 × 10−17 | 1.34 × 10−15 |
| tRF-18-INVDRID1 | i-tRF | ATGTTTAGACGGGCTCAC | MT | PheGAA | −2.7987848 | 8.31865382 | 72.1257612 | 2.02 × 10−17 | 2.07 × 10−15 |
| tRF-23-ZVELXKKSDZ | i-tRF | TTTGCACGTATGAGGCCCCGGGT | UNIQUE | AlaTGC | −2.02023747 | 6.92621223 | 70.3849902 | 4.88 × 10−17 | 4.66 × 10−15 |
| tRF_ID | tRF_Type | tRF_Sequence | Exclusive | Anticodon | Tumor vs. NAT (TCGA) |
|---|---|---|---|---|---|
| tRF-16-RPM830D | 5′-tRF | GGTAGCGTGGCCGAGC | AMBIGUOUS | LeuAAG | Downregulated |
| tRF-17-8R1546J | 3′-tRF | TCCCCAGTACCTCCACC | UNIQUE | AlaAGC | Upregulated |
| tRF-18-69M8LO04 | 5′-tRF | GGCTCCGTGGCGCAATGG | UNIQUE | ArgTCT | Upregulated |
| tRF-18-8R1546D2 | 3′-tRF | TCCCCAGTACCTCCACCA | UNIQUE | AlaAGC | Upregulated |
| tRF-18-YRRHQFD2 | 3′-tRF | TTCCCGGGCGGCGCACCA | UNIQUE | GlyCCC | Downregulated |
| tRF-22-8EKSP1852 | 3′-tRF | TCAATCCCCGGCACCTCCACCA | UNIQUE | AlaAGC | Downregulated |
| tRF-22-WD8S746D2 | 3′-tRF | TCGACTCCCGGTGTGGGAACCA | UNIQUE | GluTTC | Downregulated |
| tRF-22-WE8RO86J2 | 3′-tRF | TCGATTCCCCGACGGGGAGCCA | UNIQUE | AspGTC | Downregulated |
| tRF-22-WEKSPM852 | 3′-tRF | TCGATCCCCGGCATCTCCACCA | AMBIGUOUS | AlaTGC | Downregulated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maqueda, J.J.; Santos, M.; Ferreira, M.; Marinho, S.; Rocha, S.; Rocha, M.; Saraiva, N.; Bonito, N.; Carvalho, J.; Oliveira, C. NGS Data Repurposing Allows Detection of tRNA Fragments as Gastric Cancer Biomarkers in Patient-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 8961. https://doi.org/10.3390/ijms24108961
Maqueda JJ, Santos M, Ferreira M, Marinho S, Rocha S, Rocha M, Saraiva N, Bonito N, Carvalho J, Oliveira C. NGS Data Repurposing Allows Detection of tRNA Fragments as Gastric Cancer Biomarkers in Patient-Derived Extracellular Vesicles. International Journal of Molecular Sciences. 2023; 24(10):8961. https://doi.org/10.3390/ijms24108961
Chicago/Turabian StyleMaqueda, Joaquín J., Mafalda Santos, Marta Ferreira, Sérgio Marinho, Sara Rocha, Mafalda Rocha, Nadine Saraiva, Nuno Bonito, Joana Carvalho, and Carla Oliveira. 2023. "NGS Data Repurposing Allows Detection of tRNA Fragments as Gastric Cancer Biomarkers in Patient-Derived Extracellular Vesicles" International Journal of Molecular Sciences 24, no. 10: 8961. https://doi.org/10.3390/ijms24108961
APA StyleMaqueda, J. J., Santos, M., Ferreira, M., Marinho, S., Rocha, S., Rocha, M., Saraiva, N., Bonito, N., Carvalho, J., & Oliveira, C. (2023). NGS Data Repurposing Allows Detection of tRNA Fragments as Gastric Cancer Biomarkers in Patient-Derived Extracellular Vesicles. International Journal of Molecular Sciences, 24(10), 8961. https://doi.org/10.3390/ijms24108961