
Biological Insight and Recent Advancement in the Treatment of Neuroblastoma

Abstract
1. Introduction
2. Immunosuppressive Pathways in Neuroblastoma

{kind=link}
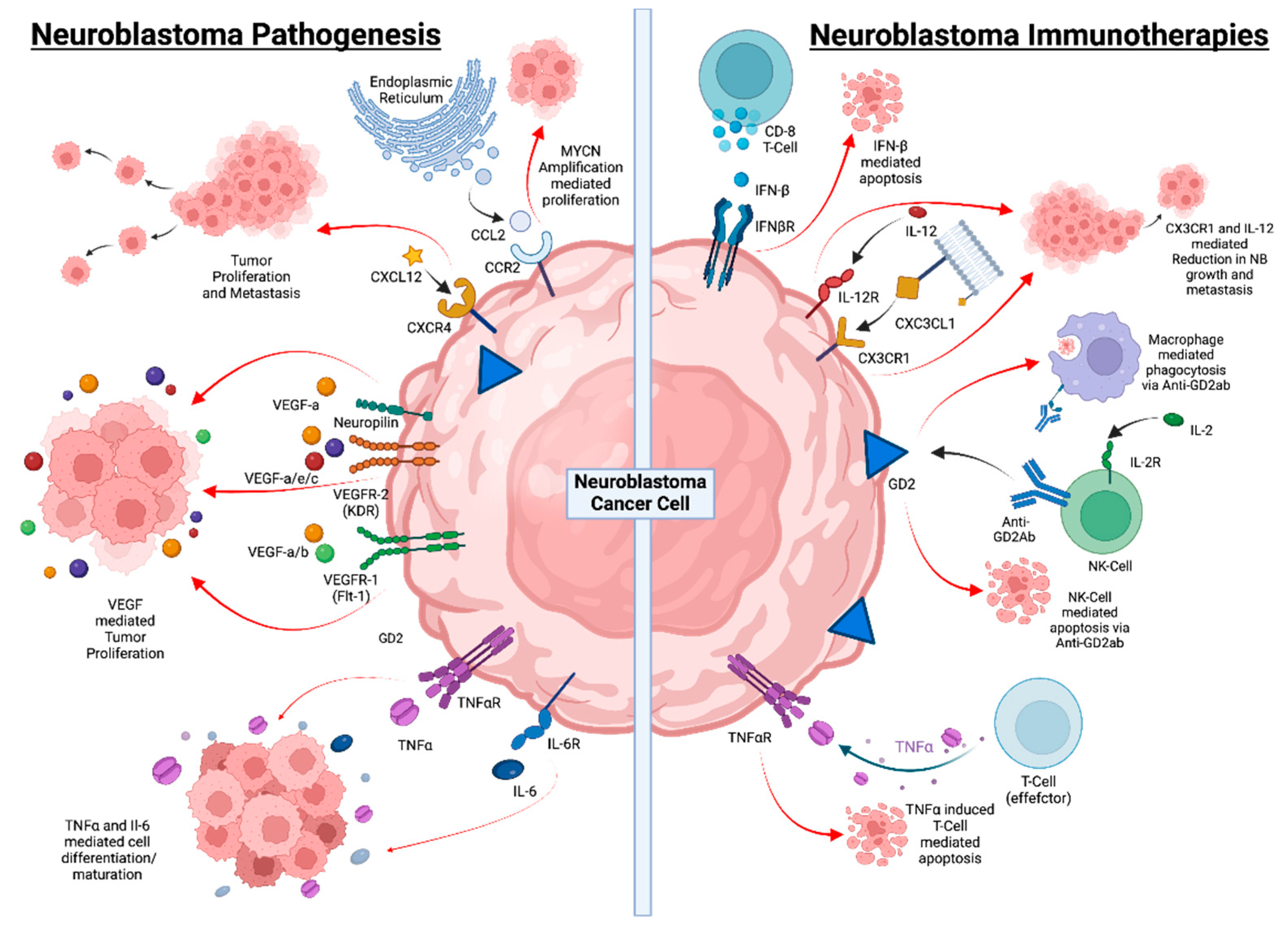{kind=link}
| Cytokine | Immunologic Function | Interaction with Immune Checkpoint Inhibitor (ICI) |
|---|---|---|
| TNFα | A proinflammatory cytokine primarily produced by activated macrophages, T lymphocytes, and NK cells. TNF plays major roles in bone remodeling, infection control, and leukocyte trafficking [37]. | TNF has been shown to induce resistance to immunotherapies and acts as a negative biomarker for prognosis. TNFα increases the expression of PD-L1 in tumor cells. Studies have also shown that a TNF-β blockade combined with ICI, such as anti-PD1, has a better therapeutic effect than ICI therapy alone [37]. |
| IFNγ | A proinflammatory cytokine primarily produced by NK cells, activated T lymphocytes, B-cells, and antigen presenting (AP) cells. IFNγ has many immunomodulatory roles including antiviral and antitumor functioning [37]. | IFNγ has an antitumor mechanism targeted by ICIs. It increases tumor immunogenicity, suppresses cancer cell proliferation, increases NK cell cytotoxic functioning, and recruit’s tumor-reactive T cells [37]. Clinic studies have reported increased IFNγ levels following anti-PD-1 ICI therapy and improved prognosis. Further, IFNγ has been shown to be a positive biomarker for successful ICI therapy [37]. |
| IL-6 | IL-6 has pro and anti-inflammatory properties. Its function is involved in cell survival and growth, immune system regulation, and carcinogenesis. Importantly, it has been shown to promote tumor transmission [44]. | IL-6 has been shown to have a negative role in immunotherapy. It has been reported that increased levels of IL-6 induce the production of myeloid-derived suppressor cells (MDSCs), which promote an immunosuppressive TME. Studies have shown that combining anti-IL6 with ICI treatment, such as anti-PD-1 immunotherapy, fosters increased anti-tumor activity and improved prognosis [44]. |
| IL-8 | A proinflammatory cytokine produced by macrophages. Its primary roles are to activate neutrophils stimulated by cellular stresses and stimulate endothelial cell proliferation. IL-8 levels have been shown to reflect tumor burden [44]. | The interaction between IL-8 and ICI therapy is unclear. However, studies have reported that increased levels of IL-8 are correlated with longer overall survival (OS) in non-small-cell lung cancer (NSCLC) patients treated with nivolumab, and anti-PD-1 ICI [44]. |
| TGF-β | A proinflammatory cytokine produced by leukocytes and stromal cells. It serves many functions, including driving the differentiation of T helper 17 (Th17) cells and regulating cell growth, proliferation, and apoptosis [32,45]. | TGF-β inhibits early cancer cells by inducing cell-cycle arrest and apoptosis. However, TGF-β has been shown to have cancer-promoting properties in later stages. Mouse models used to study urothelial cancer have demonstrated that combining anti-PD-1 therapy with the TGF-β antibody reduced the TGF-β pathway and induced tumor infiltration by cytotoxic T cells, resulting in tumor suppression. Similarly, another study has shown that combing ICI therapy with the TGF-β antibody resulted in improved prognosis compared with monotherapy [37]. |
| IL-17 | IL-17 has pro- and anti-inflammatory properties. It induces neutrophil-mediated inflammation while also suppressing autoimmune diseases [46,47]. Importantly, studies investigating colorectal cancer have shown that IL-17 exhibits protumoral properties, especially during the early stages [47]. | IL-17 signaling has been shown to promote carcinogenesis. Further, studies have shown that inhibiting signaling from this cytokine slows down oncogenesis initiation, suggesting that IL-17 inhibition may be used to halt the early stages of tumor growth. Importantly, studies utilizing mouse models have found that increased IL-17 levels were correlated with high PD-L1 expression. Further, combining IL-17 and a PD-1 blockade induced higher levels of cytotoxic T cells and tumor regression [48]. |
| Il-21 | IL-21 has pro- and anti-inflammatory properties. It regulates various immune cells such as NK and cytotoxic T cells while hindering the pro-inflammatory mechanisms of macrophages. IL-21 also exhibits anti-tumor properties [49]. | Studies have found that IL-21 hinders tumor development, especially during early stages [50]. Additionally, more recent studies using mouse models have demonstrated that combining an immune checkpoint blockade, such as anti PD-L1, with IL-21 administration increased antitumor activity, characterized by increased CD8+ T cell proliferation and by increased infiltration by, and memory of, effector T cells [50]. |
3. Immunotherapy Targets in Neuroblastoma
4. Role of MYCN in Neuroblastoma
5. Are Tumors in Children Any Different? Knowing How the Immune System Differs in Children
6. Challenges and Future Directions for NB Treatment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skertich, N.J.; Chu, F.; Tarhoni, I.A.M.; Szajek, S.; Borgia, J.A.; Madonna, M.B. Expression of Immunomodulatory Checkpoint Molecules in Drug-Resistant Neuroblastoma: An Exploratory Study. Cancers 2022, 14, 751. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M. Recent advances in neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M.; Matthay, K.K. Molecular biology of neuroblastoma. J. Clin. Oncol. 1999, 17, 2264–2279. [Google Scholar] [CrossRef]
- Tolbert, V.P.; Matthay, K.K. Neuroblastoma: Clinical and biological approach to risk stratification and treatment. Cell Tissue Res. 2018, 372, 195–209. [Google Scholar] [CrossRef]
- Chung, C.; Boterberg, T.; Lucas, J.; Panoff, J.; Valteau-Couanet, D.; Hero, B.; Bagatell, R.; Hill-Kayser, C.E. Neuroblastoma. Pediatr. Blood Cancer 2021, 68 (Suppl. 2). [Google Scholar] [CrossRef]
- Nguyen, R.; Thiele, C.J. Immunotherapy approaches targeting neuroblastoma. Curr. Opin. Pediatr. 2021, 33, 19–25. [Google Scholar] [CrossRef]
- Lin, K.S.; Uemura, S.; Thwin, K.K.M.; Nakatani, N.; Ishida, T.; Yamamoto, N.; Tamura, A.; Saito, A.; Mori, T.; Hasegawa, D.; et al. Minimal residual disease in high-risk neuroblastoma shows a dynamic and disease burden-dependent correlation between bone marrow and peripheral blood. Transl. Oncol. 2021, 14, 101019. [Google Scholar] [CrossRef]
- Cheung, N.K.; Dyer, M.A. Neuroblastoma: Developmental biology, cancer genomics and immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef]
- Cohn, S.L.; Pearson, A.D.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (INRG) classification system: An INRG Task Force report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef]
- Brodeur, G.M. Neuroblastoma: Biological insights into a clinical enigma. Nat. Rev. Cancer 2003, 3, 203–216. [Google Scholar] [CrossRef]
- Wienke, J.; Dierselhuis, M.P.; Tytgat, G.A.M.; Künkele, A.; Nierkens, S.; Molenaar, J.J. The immune landscape of neuroblastoma: Challenges and opportunities for novel therapeutic strategies in pediatric oncology. Eur. J. Cancer 2021, 144, 123–150. [Google Scholar] [CrossRef] [PubMed]
- Raffaghello, L.; Prigione, I.; Airoldi, I.; Camoriano, M.; Morandi, F.; Bocca, P.; Gambini, C.; Ferrone, S.; Pistoia, V. Mechanisms of immune evasion of human neuroblastoma. Cancer Lett. 2005, 228, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Blavier, L.; Yang, R.M.; DeClerck, Y.A. The Tumor Microenvironment in Neuroblastoma: New Players, New Mechanisms of Interaction and New Perspectives. Cancers 2020, 12, 2912. [Google Scholar] [CrossRef] [PubMed]
- Masih, K.E.; Wei, J.S.; Milewski, D.; Khan, J. Exploring and Targeting the Tumor Immune Microenvironment of Neuroblastoma. J. Cell. Immunol. 2021, 3, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Casey, D.L.; Cheung, N.-K.V. Immunotherapy of Pediatric Solid Tumors: Treatments at a Crossroads, with an Emphasis on Antibodies. Cancer Immunol. Res. 2020, 8, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Sarioglu, F.C.; Salman, M.; Guleryuz, H.; Ozer, E.; Cecen, E.; Ince, D.; Olgun, N. Radiological staging in neuroblastoma: Computed tomography or magnetic resonance imaging? Pol. J. Radiol. 2019, 84, e46–e53. [Google Scholar] [CrossRef]
- Brodeur, G.M.; Hogarty, M.D.; Mosse, Y.P.; Maris, J.M. Neuroblastoma. In Pizzo and Poplack’s Principles and Practice of Pediatric Oncology; Blaney, S.M.A.P., Helman, L.J., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2021; Volume 8. [Google Scholar]
- Leonard, M.P.; Gearhart, J.P.; Jeffs, R.D. Chemoimmunotherapy in conjunction with surgery: Strategies for management of murine neuroblastoma. J. Pediatr. Surg. 1991, 26, 1224–1229. [Google Scholar] [CrossRef]
- Smith, V.; Foster, J. High-Risk Neuroblastoma Treatment Review. Children 2018, 5, 114. [Google Scholar] [CrossRef]
- Yalçin, B.; Kremer, L.C.; van Dalen, E.C. High-dose chemotherapy and autologous haematopoietic stem cell rescue for children with high-risk neuroblastoma. Cochrane Database Syst. Rev. 2015, 2015, Cd006301. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Cheng, Y.; Hu, A.; Li, D.; Wang, X.; Guo, Y.; Zhou, Y.; Chen, G.; Bao, B.; et al. Long-Term Survival of Neuroblastoma Patients Receiving Surgery, Chemotherapy, and Radiotherapy: A Propensity Score Matching Study. J. Clin. Med. 2023, 12, 754. [Google Scholar] [CrossRef] [PubMed]
- PDQ Pediatric Treatment Editorial Board. Neuroblastoma Treatment (PDQ®): Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute (US): Bethesda, MD, USA, 2002. [Google Scholar]
- Kayano, D.; Kinuya, S. Iodine-131 metaiodobenzylguanidine therapy for neuroblastoma: Reports so far and future perspective. Sci. World J. 2015, 2015, 189135. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Rangarajan, V.; Shah, S.; Puranik, A.; Purandare, N. MIBG (metaiodobenzylguanidine) theranostics in pediatric and adult malignancies. Br. J. Radiol. 2018, 91, 20180103. [Google Scholar] [CrossRef] [PubMed]
- Gay, A.N.; Chang, S.; Rutland, L.; Yu, L.; Byeseda, S.; Naik-Mathuria, B.; Cass, D.L.; Russell, H.; Olutoye, O.O. Granulocyte colony stimulating factor alters the phenotype of neuroblastoma cells: Implications for disease-free survival of high-risk patients. J. Pediatr. Surg. 2008, 43, 837–842. [Google Scholar] [CrossRef]
- Reynolds, C.P.; Matthay, K.K.; Villablanca, J.G.; Maurer, B.J. Retinoid therapy of high-risk neuroblastoma. Cancer Lett. 2003, 197, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Majzner, R.G.; Sondel, P.M. Immunotherapy of Neuroblastoma: Facts and Hopes. Clin. Cancer Res. 2022, 28, 3196–3206. [Google Scholar] [CrossRef]
- Matthay, K.K.; Villablanca, J.G.; Seeger, R.C.; Stram, D.O.; Harris, R.E.; Ramsay, N.K.; Swift, P.; Shimada, H.; Black, C.T.; Brodeur, G.M.; et al. Treatment of High-Risk Neuroblastoma with Intensive Chemotherapy, Radiotherapy, Autologous Bone Marrow Transplantation, and 13-cis-Retinoic Acid. N. Engl. J. Med. 1999, 341, 1165–1173. [Google Scholar] [CrossRef]
- Kumar, A.; Taghi Khani, A.; Sanchez Ortiz, A.; Swaminathan, S. GM-CSF: A Double-Edged Sword in Cancer Immunotherapy. Front. Immunol. 2022, 13, 901277. [Google Scholar] [CrossRef] [PubMed]
- Stepan-Giljevic, J.; Jakovljevic, G.; Bonevski, A.; Cvrlje, F.J.; Rajacic, N.; Pavlovic, M.; Kranjcec, I. 300 Immunotherapy in high risk neuroblastoma—Treatment algorithm of complications. Arch. Dis. Child. 2021, 106, A127. [Google Scholar] [CrossRef]
- Szanto, C.L.; Cornel, A.M.; Vijver, S.V.; Nierkens, S. Monitoring Immune Responses in Neuroblastoma Patients during Therapy. Cancers 2020, 12, 519. [Google Scholar] [CrossRef]
- Mina, M.; Boldrini, R.; Citti, A.; Romania, P.; D’Alicandro, V.; De Ioris, M.; Castellano, A.; Furlanello, C.; Locatelli, F.; Fruci, D. Tumor-infiltrating T lymphocytes improve clinical outcome of therapy-resistant neuroblastoma. Oncoimmunology 2015, 4, e1019981. [Google Scholar] [CrossRef] [PubMed]
- Tilak, T.; Sherawat, S.; Agarwala, S.; Gupta, R.; Vishnubhatla, S.; Bakhshi, S. Circulating T-regulatory cells in neuroblastoma: A pilot prospective study. Pediatr. Hematol. Oncol. 2014, 31, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.X.; Shimada, H.; Ikegaki, N. Clinical Relevance of CD4 Cytotoxic T Cells in High-Risk Neuroblastoma. Front. Immunol. 2021, 12, 650427. [Google Scholar] [CrossRef]
- Pelosi, A.; Fiore, P.F.; Di Matteo, S.; Veneziani, I.; Caruana, I.; Ebert, S.; Munari, E.; Moretta, L.; Maggi, E.; Azzarone, B. Pediatric Tumors-Mediated Inhibitory Effect on NK Cells: The Case of Neuroblastoma and Wilms’ Tumors. Cancers 2021, 13, 2374. [Google Scholar] [CrossRef]
- Wang, M.; Zhai, X.; Li, J.; Guan, J.; Xu, S.; Li, Y.; Zhu, H. The Role of Cytokines in Predicting the Response and Adverse Events Related to Immune Checkpoint Inhibitors. Front. Immunol. 2021, 12, 670391. [Google Scholar] [CrossRef]
- Fultang, L.; Gamble, L.D.; Gneo, L.; Berry, A.M.; Egan, S.A.; De Bie, F.; Yogev, O.; Eden, G.L.; Booth, S.; Brownhill, S.; et al. Macrophage-Derived IL1β and TNFα Regulate Arginine Metabolism in Neuroblastoma. Cancer Res. 2019, 79, 611–624. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front. Immunol. 2020, 10, 3038. [Google Scholar] [CrossRef]
- Main, E.K.; Lampson, L.A.; Hart, M.K.; Kornbluth, J.; Wilson, D.B. Human neuroblastoma cell lines are susceptible to lysis by natural killer cells but not by cytotoxic T lymphocytes. J. Immunol. 1985, 135, 242–246. [Google Scholar] [CrossRef]
- Spel, L.; Boelens, J.J.; van der Steen, D.M.; Blokland, N.J.; van Noesel, M.M.; Molenaar, J.J.; Heemskerk, M.H.; Boes, M.; Nierkens, S. Natural killer cells facilitate PRAME-specific T-cell reactivity against neuroblastoma. Oncotarget 2015, 6, 35770–35781. [Google Scholar] [CrossRef]
- Bottino, C.; Dondero, A.; Bellora, F.; Moretta, L.; Locatelli, F.; Pistoia, V.; Moretta, A.; Castriconi, R. Natural killer cells and neuroblastoma: Tumor recognition, escape mechanisms, and possible novel immunotherapeutic approaches. Front. Immunol. 2014, 5, 56. [Google Scholar] [CrossRef]
- Veneziani, I.; Fruci, D.; Compagnone, M.; Pistoia, V.; Rossi, P.; Cifaldi, L. The BET-bromodomain inhibitor JQ1 renders neuroblastoma cells more resistant to NK cell-mediated recognition and killing by downregulating ligands for NKG2D and DNAM-1 receptors. Oncotarget 2019, 10, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, Y.; Amano, Y.; Kanata, K.; Hasegwa, H.; Matsui, T.; Kakutani, T.; Koyauchi, T.; Tanahashi, M.; Niwa, H.; Yokomura, K.; et al. Impact of early inflammatory cytokine elevation after commencement of PD-1 inhibitors to predict efficacy in patients with non-small cell lung cancer. Med. Oncol. 2019, 36, 33. [Google Scholar] [CrossRef]
- Muñoz-Fernández, M.A.; Cano, E.; O’Donnell, C.A.; Doyle, J.; Liew, F.Y.; Fresno, M. Tumor necrosis factor-alpha (TNF-alpha), interferon-gamma, and interleukin-6 but not TNF-beta induce differentiation of neuroblastoma cells: The role of nitric oxide. J. Neurochem. 1994, 62, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontology 2000 2015, 69, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chai, W.; Yang, W.; Han, W.; Mou, W.; Xi, Y.; Chen, X.; Wang, H.; Wang, W.; Qin, H.; et al. The increased IL-17-producing γδT cells promote tumor cell proliferation and migration in neuroblastoma. Clin. Immunol. 2020, 211, 108343. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, R.; Wang, B.; Lian, J.; Yao, Y.; Sun, H.; Zhang, C.; Fang, L.; Guan, X.; Shi, J.; et al. Blocking IL-17A enhances tumor response to anti-PD-1 immunotherapy in microsatellite stable colorectal cancer. J. Immunother. Cancer 2021, 9, e001895. [Google Scholar] [CrossRef] [PubMed]
- Jian, L.; Li, C.; Wang, X.; Sun, L.; Ma, Z.; Zhao, J. IL-21 impairs pro-inflammatory activity of M1-like macrophages exerting anti-inflammatory effects on rheumatoid arthritis. Autoimmunity 2022, 55, 75–85. [Google Scholar] [CrossRef]
- Lewis, K.E.; Selby, M.J.; Masters, G.; Valle, J.; Dito, G.; Curtis, W.R.; Garcia, R.; Mink, K.A.; Waggie, K.S.; Holdren, M.S.; et al. Interleukin-21 combined with PD-1 or CTLA-4 blockade enhances antitumor immunity in mouse tumor models. Oncoimmunology 2017, 7, e1377873. [Google Scholar] [CrossRef]
- Sait, S.; Modak, S. Anti-GD2 immunotherapy for neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 889–904. [Google Scholar] [CrossRef]
- Whittle, S.B.; Smith, V.; Doherty, E.; Zhao, S.; McCarty, S.; Zage, P.E. Overview and recent advances in the treatment of neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 369–386. [Google Scholar] [CrossRef]
- Ren, X.; Guo, S.; Guan, X.; Kang, Y.; Liu, J.; Yang, X. Immunological Classification of Tumor Types and Advances in Precision Combination Immunotherapy. Front. Immunol. 2022, 13, 790113. [Google Scholar] [CrossRef] [PubMed]
- Baxter, D. Active and passive immunization for cancer. Hum. Vaccines Immunother. 2014, 10, 2123–2129. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef]
- Raffaghello, L.; Prigione, I.; Airoldi, I.; Camoriano, M.; Levreri, I.; Gambini, C.; Pende, D.; Steinle, A.; Ferrone, S.; Pistoia, V. Downregulation and/or release of NKG2D ligands as immune evasion strategy of human neuroblastoma. Neoplasia 2004, 6, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Bao, R.; Spranger, S.; Hernandez, K.; Zha, Y.; Pytel, P.; Luke, J.J.; Gajewski, T.F.; Volchenboum, S.L.; Cohn, S.L.; Desai, A.V. Immunogenomic determinants of tumor microenvironment correlate with superior survival in high-risk neuroblastoma. J. Immunother. Cancer 2021, 9, e002417. [Google Scholar] [CrossRef]
- Mody, R.; Naranjo, A.; Van Ryn, C.; Yu, A.L.; London, W.B.; Shulkin, B.L.; Parisi, M.T.; Servaes, S.E.; Diccianni, M.B.; Sondel, P.M.; et al. Irinotecan-temozolomide with temsirolimus or dinutuximab in children with refractory or relapsed neuroblastoma (COG ANBL1221): An open-label, randomised, phase 2 trial. Lancet Oncol. 2017, 18, 946–957. [Google Scholar] [CrossRef]
- Kanold, J.; Paillard, C.; Tchirkov, A.; Lang, P.; Kelly, A.; Halle, P.; Isfan, F.; Merlin, E.; Marabelle, A.; Rochette, E.; et al. NK cell immunotherapy for high-risk neuroblastoma relapse after haploidentical HSCT. Pediatr. Blood Cancer 2012, 59, 739–742. [Google Scholar] [CrossRef]
- Federico, S.M.; McCarville, M.B.; Shulkin, B.L.; Sondel, P.M.; Hank, J.A.; Hutson, P.; Meagher, M.; Shafer, A.; Ng, C.Y.; Leung, W.; et al. A Pilot Trial of Humanized Anti-GD2 Monoclonal Antibody (hu14.18K322A) with Chemotherapy and Natural Killer Cells in Children with Recurrent/Refractory Neuroblastoma. Clin. Cancer Res. 2017, 23, 6441–6449. [Google Scholar] [CrossRef]
- Modak, S.; Le Luduec, J.B.; Cheung, I.Y.; Goldman, D.A.; Ostrovnaya, I.; Doubrovina, E.; Basu, E.; Kushner, B.H.; Kramer, K.; Roberts, S.S.; et al. Adoptive immunotherapy with haploidentical natural killer cells and Anti-GD2 monoclonal antibody m3F8 for resistant neuroblastoma: Results of a phase I study. Oncoimmunology 2018, 7, e1461305. [Google Scholar] [CrossRef]
- Gröbner, S.N.; Worst, B.C.; Weischenfeldt, J.; Buchhalter, I.; Kleinheinz, K.; Rudneva, V.A.; Johann, P.D.; Balasubramanian, G.P.; Segura-Wang, M.; Brabetz, S.; et al. The landscape of genomic alterations across childhood cancers. Nature 2018, 555, 321–327. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science 2015, 348, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Haen, S.P.; Löffler, M.W.; Rammensee, H.G.; Brossart, P. Towards new horizons: Characterization, classification and implications of the tumour antigenic repertoire. Nat. Rev. Clin. Oncol. 2020, 17, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Yarmarkovich, M.; Marshall, Q.F.; Warrington, J.M.; Premaratne, R.; Farrel, A.; Groff, D.; Li, W.; di Marco, M.; Runbeck, E.; Truong, H.; et al. Cross-HLA targeting of intracellular oncoproteins with peptide-centric CARs. Nature 2021, 599, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Mosse, Y.P.; Laudenslager, M.; Khazi, D.; Carlisle, A.J.; Winter, C.L.; Rappaport, E.; Maris, J.M. Germline PHOX2B mutation in hereditary neuroblastoma. Am. J. Hum. Genet. 2004, 75, 727–730. [Google Scholar] [CrossRef]
- Marabelle, A.; Kohrt, H.; Caux, C.; Levy, R. Intratumoral immunization: A new paradigm for cancer therapy. Clin. Cancer Res. 2014, 20, 1747–1756. [Google Scholar] [CrossRef]
- Sagiv-Barfi, I.; Czerwinski, D.K.; Levy, S.; Alam, I.S.; Mayer, A.T.; Gambhir, S.S.; Levy, R. Eradication of spontaneous malignancy by local immunotherapy. Sci. Transl. Med. 2018, 10, eaan4488. [Google Scholar] [CrossRef]
- Morris, Z.S.; Guy, E.I.; Francis, D.M.; Gressett, M.M.; Werner, L.R.; Carmichael, L.L.; Yang, R.K.; Armstrong, E.A.; Huang, S.; Navid, F.; et al. In Situ Tumor Vaccination by Combining Local Radiation and Tumor-Specific Antibody or Immunocytokine Treatments. Cancer Res. 2016, 76, 3929–3941. [Google Scholar] [CrossRef]
- Voeller, J.; Erbe, A.K.; Slowinski, J.; Rasmussen, K.; Carlson, P.M.; Hoefges, A.; VandenHeuvel, S.; Stuckwisch, A.; Wang, X.; Gillies, S.D.; et al. Combined innate and adaptive immunotherapy overcomes resistance of immunologically cold syngeneic murine neuroblastoma to checkpoint inhibition. J. Immunother. Cancer 2019, 7, 344. [Google Scholar] [CrossRef]
- Melder, R.J.; Koenig, G.C.; Witwer, B.P.; Safabakhsh, N.; Munn, L.L.; Jain, R.K. During angiogenesis, vascular endothelial growth factor regulate natural killer cell adhesion to tumor endothelium. Nat. Med. 1996, 2, 992–997. [Google Scholar] [CrossRef]
- Sköldenberg, E.G.; Larsson, A.; Jakobson, A.; Hedborg, F.; Kogner, P.; Christofferson, R.H.; Azarbayjani, F. The angiogenic growth factors HGF and VEGF in serum and plasma from neuroblastoma patients. Anticancer Res. 2009, 29, 3311–3319. [Google Scholar]
- Bose, S.; Cho, J. Role of chemokine CCL2 and its receptor CCR2 in neurodegenerative diseases. Arch. Pharm. Res. 2013, 36, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Metelitsa, L.S.; Wu, H.W.; Wang, H.; Yang, Y.; Warsi, Z.; Asgharzadeh, S.; Groshen, S.; Wilson, S.B.; Seeger, R.C. Natural killer T cells infiltrate neuroblastomas expressing the chemokine CCL2. J. Exp. Med. 2004, 199, 1213–1221. [Google Scholar] [CrossRef]
- Guyon, A. CXCL12 chemokine and its receptors as major players in the interactions between immune and nervous systems. Front. Cell. Neurosci. 2014, 8, 65. [Google Scholar] [CrossRef]
- Eier, R.; Mühlethaler-Mottet, A.; Flahaut, M.; Coulon, A.; Fusco, C.; Louache, F.; Auderset, K.; Bourloud, K.B.; Daudigeos, E.; Ruegg, C.; et al. The chemokine receptor CXCR4 strongly promotes neuroblastoma primary tumour and metastatic growth, but not invasion. PLoS ONE 2007, 2, e1016. [Google Scholar] [CrossRef]
- Raza, S.; Rajak, S.; Tewari, A.; Gupta, P.; Chattopadhyay, N.; Sinha, R.A.; Chakravarti, B. Multifaceted role of chemokines in solid tumors: From biology to therapy. Semin. Cancer Biol. 2022, 86, 1105–1121. [Google Scholar] [CrossRef]
- Bule, P.; Aguiar, S.I.; Aires-Da-Silva, F.; Dias, J.N.R. Chemokine-Directed Tumor Microenvironment Modulation in Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 9804. [Google Scholar] [CrossRef] [PubMed]
- Hibi, M.; Murakami, M.; Saito, M.; Hirano, T.; Taga, T.; Kishimoto, T. Molecular cloning and expression of an IL-6 signal transducer, gp130. Cell 1990, 63, 1149–1157. [Google Scholar] [CrossRef]
- Ara, T.; Song, L.; Shimada, H.; Keshelava, N.; Russell, H.V.; Metelitsa, L.S.; Groshen, S.G.; Seeger, R.C.; DeClerck, Y.A. Interleukin-6 in the bone marrow microenvironment promotes the growth and survival of neuroblastoma cells. Cancer Res. 2009, 69, 329–337. [Google Scholar] [CrossRef]
- Zeng, Y.; Huebener, N.; Fest, S.; Weixler, S.; Schroeder, U.; Gaedicke, G.; Xiang, R.; Schramm, A.; Eggert, A.; Reisfeld, R.A.; et al. Fractalkine (CX3CL1)- and interleukin-2-enriched neuroblastoma microenvironment induces eradication of metastases mediated by T cells and natural killer cells. Cancer Res. 2007, 67, 2331–2338. [Google Scholar] [CrossRef]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New insights into the regulation of T cells by gamma(c) family cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef]
- Vicari, A.P.; Chiodoni, C.; Vaure, C.; Aït-Yahia, S.; Dercamp, C.; Matsos, F.; Reynard, O.; Taverne, C.; Merle, P.; Colombo, M.P.; et al. Reversal of tumor-induced dendritic cell paralysis by CpG immunostimulatory oligonucleotide and anti-interleukin 10 receptor antibody. J. Exp. Med. 2002, 196, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Brignole, C.; Marimpietri, D.; Pastorino, F.; Di Paolo, D.; Pagnan, G.; Loi, M.; Piccardi, F.; Cilli, M.; Tradori-Cappai, A.; Arrigoni, G.; et al. Anti-IL-10R antibody improves the therapeutic efficacy of targeted liposomal oligonucleotides. J. Control. Release 2009, 138, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Milo, R.; Swoveland, P.; Johnson, K.P.; Panitch, H.; Dhib-Jalbut, S. Interferon beta-1b reduces interferon gamma-induced antigen-presenting capacity of human glial and B cells. J. Neuroimmunol. 1995, 61, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Rosati, S.F.; Williams, R.F.; Nunnally, L.C.; McGee, M.C.; Sims, T.L.; Tracey, L.; Zhou, J.; Fan, M.; Ng, C.Y.; Nathwani, A.C.; et al. IFN-beta sensitizes neuroblastoma to the antitumor activity of temozolomide by modulating O6-methylguanine DNA methyltransferase expression. Mol. Cancer Ther. 2008, 7, 3852–3858. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, S.S.; Clarke, S.; Veschi, V.; Thiele, C.J. Targeting MYCN in Pediatric and Adult Cancers. Front. Oncol. 2020, 10, 623679. [Google Scholar] [CrossRef]
- Seeger, R.C.; Brodeur, G.M.; Sather, H.; Dalton, A.; Siegel, S.E.; Wong, K.Y.; Hammond, D. Association of multiple copies of the N-myc oncogene with rapid progression of neuroblastomas. N. Engl. J. Med. 1985, 313, 1111–1116. [Google Scholar] [CrossRef]
- Wei, J.S.; Kuznetsov, I.B.; Zhang, S.; Song, Y.K.; Asgharzadeh, S.; Sindiri, S.; Wen, X.; Patidar, R.; Najaraj, S.; Walton, A.; et al. Clinically Relevant Cytotoxic Immune Cell Signatures and Clonal Expansion of T-Cell Receptors in High-Risk MYCN-Not-Amplified Human Neuroblastoma. Clin. Cancer Res. 2018, 24, 5673–5684. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, X.; Basu, M.; Dong, C.; Zheng, P.; Liu, Y.; Sandler, A.D. MYCN Amplification Is Associated with Repressed Cellular Immunity in Neuroblastoma: An In Silico Immunological Analysis of TARGET Database. Front. Immunol. 2017, 8, 1473. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, Y.; Wang, L.; Zhang, H.; Liu, H.; Liu, Y. Cellular components in tumor microenvironment of neuroblastoma and the prognostic value. PeerJ 2019, 7, e8017. [Google Scholar] [CrossRef]
- Layer, J.P.; Kronmüller, M.T.; Quast, T.; van den Boorn-Konijnenberg, D.; Effern, M.; Hinze, D.; Althoff, K.; Schramm, A.; Westermann, F.; Peifer, M.; et al. Amplification of N-Myc is associated with a T-cell-poor microenvironment in metastatic neuroblastoma restraining interferon pathway activity and chemokine expression. Oncoimmunology 2017, 6, e1320626. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Elements of cancer immunity and the cancer-immune set point. Nature 2017, 541, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Künkele, A.; Taraseviciute, A.; Finn, L.S.; Johnson, A.J.; Berger, C.; Finney, O.; Chang, C.A.; Rolczynski, L.S.; Brown, C.; Mgebroff, S.; et al. Preclinical Assessment of CD171-Directed CAR T-cell Adoptive Therapy for Childhood Neuroblastoma: CE7 Epitope Target Safety and Product Manufacturing Feasibility. Clin. Cancer Res. 2017, 23, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Simon, F. Immunomodulatory cytokines: Directing and controlling immune activation. Arthritis Res. Ther. 2011, 13, O14. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef] [PubMed]
- Hutzen, B.; Ghonime, M.; Lee, J.; Mardis, E.R.; Wang, R.; Lee, D.A.; Cairo, M.S.; Roberts, R.D.; Cripe, T.P.; Cassady, K.A. Immunotherapeutic Challenges for Pediatric Cancers. Mol. Ther. -Oncolytics 2019, 15, 38–48. [Google Scholar] [CrossRef]
- Kugel, C.H., 3rd; Douglass, S.M.; Webster, M.R.; Kaur, A.; Liu, Q.; Yin, X.; Weiss, S.A.; Darvishian, F.; Al-Rohil, R.N.; Ndoye, A.; et al. Age Correlates with Response to Anti-PD1, Reflecting Age-Related Differences in Intratumoral Effector and Regulatory T-Cell Populations. Clin. Cancer Res. 2018, 24, 5347–5356. [Google Scholar] [CrossRef]
- Richards, R.M.; Sotillo, E.; Majzner, R.G. CAR T Cell Therapy for Neuroblastoma. Front. Immunol. 2018, 9, 2380. [Google Scholar] [CrossRef]
- Williams, E.L.; Dunn, S.N.; James, S.; Johnson, P.W.; Cragg, M.S.; Glennie, M.J.; Gray, J.C. Immunomodulatory monoclonal antibodies combined with peptide vaccination provide potent immunotherapy in an aggressive murine neuroblastoma model. Clin. Cancer Res. 2013, 19, 3545–3555. [Google Scholar] [CrossRef]
- Tierney, J.F.; Vogle, A.; Poirier, J.; Min, I.M.; Finnerty, B.; Zarnegar, R.; Pappas, S.G.; Scognamiglio, T.; Ghai, R.; Gattuso, P.; et al. Expression of programmed death ligand 1 and 2 in adrenocortical cancer tissues: An exploratory study. Surgery 2019, 165, 196–201. [Google Scholar] [CrossRef]
- DuBois, S.G.; Kalika, Y.; Lukens, J.N.; Brodeur, G.M.; Seeger, R.C.; Atkinson, J.B.; Haase, G.M.; Black, C.T.; Perez, C.; Shimada, H.; et al. Metastatic sites in stage IV and IVS neuroblastoma correlate with age, tumor biology, and survival. J. Pediatr. Hematol. Oncol. 1999, 21, 181–189. [Google Scholar] [CrossRef]
- Newman Jenna, H.; Chesson, C.B.; Herzog Nora, L.; Bommareddy Praveen, K.; Aspromonte Salvatore, M.; Pepe, R.; Estupinian, R.; Aboelatta Mones, M.; Buddhadev, S.; Tarabichi, S.; et al. Intratumoral injection of the seasonal flu shot converts immunologically cold tumors to hot and serves as an immunotherapy for cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Melaiu, O.; Chierici, M.; Lucarini, V.; Jurman, G.; Conti, L.A.; De Vito, R.; Boldrini, R.; Cifaldi, L.; Castellano, A.; Furlanello, C.; et al. Cellular and gene signatures of tumor-infiltrating dendritic cells and natural-killer cells predict prognosis of neuroblastoma. Nat. Commun. 2020, 11, 5992. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.; Sun, J.; Zuo, P.; Da, M.; Mo, X.; Fang, Y. Verification of genetic differences and immune cell infiltration subtypes in the neuroblastoma tumour microenvironment during immunotherapy. World J. Surg. Oncol. 2022, 20, 169. [Google Scholar] [CrossRef] [PubMed]


| Treatment | Indication | Clinical Uses and Benefits | Risks and Side Effects | Agents |
|---|---|---|---|---|
| Surgery | First-line treatment to remove tumor | Results from surgery are often used to determine disease stage if imaging is insufficient and there is a need for additional treatment [17]. For example, if the tumor is in its early stages, it may be possible to completely resect the mass. In other cases, the tumor may not be able to be removed completely, or may have metastasized, requiring the patient to undergo additional treatment such as chemotherapy [18]. | Surgical risks largely depend on tumor location, duration of surgery, and the patient’s overall health. Major complications are rare but can include anesthetic errors; bleeding; infection; and damage to organs, nerves, or blood vessels. The risk for complications increases if the tumor is growing into nerves or blood vessels [18]. | N/A |
| Chemotherapy | L1, L2, M, MS First line in cases where the cancer has metastasized too far to be completely removed by surgery | Chemotherapy is often used in conjunction with surgery [19]. Chemotherapy may be used prior to or after surgery. For patients in high-risk groups, a combination of chemotherapy drugs may be used, as well as other therapy regimes. Factors considered are location of the tumor, age, patient health, and others [20]. | There are many risks that must be considered and discussed with patients and their families before initiation of chemotherapy [21]. Chemotherapy drugs can be damaging to various cells in the body, leading to side effects. The most common are hair loss, sores, loss of appetite, nausea, vomiting, and GI upset. More severe side effects include leukopenia causing infection; thrombocytopenia leading to excessive bruising and bleeding; and anemia [22]. | Cyclophosphamide Cisplatin Vincristine Doxorubicin Etoposide Topotecan Melphalan Busulfan Thiotepa |
| Radiation Therapy | M, MS Only used in patients considered high-risk | Radiation therapy is necessary for cases where the cancer has spread and is not removable with surgery and chemotherapy alone. It also may be used in patients with emergency, life-threatening symptoms to rapidly reduce the tumor’s size [23]. Two radiotherapy options are available. External beam radiation therapy focuses a radiation beam onto the tumor from a machine source, known as total body irradiation. Iodine meta-iodebenzylfuanidine (MIBG) radiotherapy, which is chemically similar to norepinephrine (NE), may be initially injected into the blood to detect neuroblastoma or to directly deliver radiation to the tumor [24]. | Radiation therapy is avoided when possible due to the high risk of various short and long-term side effects. These include skin burns, hair loss and other skin reactions, nausea and GI upset. Long-term effects include bone damage; growth arrest; hypothyroidism; cardiovascular and respiratory problems; and cellular damage, especially to DNA [22]. MIBG therapy typically results in milder side effects due to its local mechanism of action [25]. | External beam radiation therapy MIBG radiotherapy |
| High-Dose Chemotherapy and Stem Cell Transplant (STC) | M, MS used in high-risk patients unlikely to be respond to other treatments | This treatment is a combination of chemotherapy, usually higher doses, and bone marrow stem cell transplantation to replace the bone marrow cells injured by the chemotherapy [21]. Many patients have a second stem cell transplant several months apart. The patient is given a medication called filgrastim (G-CSF) to induce bone marrow cell proliferation, which is then collected and later used for the transplant [26]. | High-dose chemotherapy and stem cell transplantation is a complex procedure and comes with several short- and long-term effects. High-dose chemotherapy is severely toxic to the body’s cells and can lead to anemia, bleeding, GI upset, mouth sores, loss of appetite, and hair loss. Long-term effects include damage to various organs including the liver, heart, or lungs; hypothyroidism; poor hormone control; infertility; osteopenia; and higher risk for infections and other cancers such as leukemia [21]. | High doses of chemotherapy agents listed above |
| Retinoid Therapy | M, MS Used after completing high-dose chemotherapy and STC | This treatment uses retinoids. Retinoids are chemical compounds that are structurally related to vitamin A and work as differentiating agents, meaning that they are thought to induce cancer cell differentiation to normal cells [27]. Retinoid therapy is recommended for high-risk neuroblastoma patients that have completed high-dose chemotherapy [28] | The side effects of retinoid therapy are less severe than the other neuroblastoma treatments. The most often reported side effects are cracked lips, joint and muscle pain, and epistaxis [29]. | Isotretinoin (13-cis-retinoic acid) |
| Cytokine | Function | Role in Neuroblastoma |
|---|---|---|
| VEGF | VEGF acts as a pro-inflammatory cytokine by increasing endothelial cell permeability, by inducing the expression of endothelial cell adhesion molecules, and via its ability to act as a monocyte chemo-attractant [71]. | Studies looking at the expression of several markers in NB xenografts have shown that some angiogenic factors including VEGF-A, -B and -C are associated with advanced NB stage [72]. |
| CCL2 | Chemokine CCL2 (also known as monocyte chemo-attractant protein-1, MCP-1) is one of the vital chemokines that control the migration and infiltration of monocytes/macrophages [73]. | The infiltration of neuroblastoma cells by invariant NKT (iNKT) cells was found to correlate with the expression of the chemokine CCL2 by the tumor [74]. |
| CXCL12 | CXCL12 acts through its receptors CXCR4 and CXCR7. CXCR4 stimulation leads to the activation of numerous signaling pathways depending on the associated cell types, while CXCR7 has mainly been shown to be involved in scavenging CXCL12, although it can activate a MAP kinase pathway through β-arrestin in several systems [75]. | CXCL12 and CXCR4 have been demonstrated to be overexpressed in NB cell lines in addition to primary metastatic NB. This hints at the role of CXCL12 in its connection to autocrine/paracrine signaling of tumor growth instead of the development of metastatic pathways [76]. |
| CXC3CL1 | CXC3CL1 is an unusual chemokine expressed on the cell surface and acting as adhesion molecule by binding to its receptor CX3CR1.7 and is also expressed in a variety of human tissues and cell lines, where it mediates leukocyte migration and adhesion [77]. | It has been shown in animal models that CX3CL1 is able to inhibit NB growth and eradicate metastasis when used in combination with IL-12 through the attraction of immune cells to the tumor site [78]. |
| IL-6 | IL-6 and VEGF are the best characterized cytokines to stimulate tumor growth and metastasis, while others, such as IFN-γ, can exert anti-NB activity by inducing tumor cell apoptosis and inhibiting angiogenesis [79]. | IL-6 is introduced into the bone marrow by the bone marrow stromal cells (BMSC) which promotes the growth and survival of neuroblastoma cells [80]. |
| IL-7 | IL-7 is a cytokine that stimulates proliferation of all cells in the lymphoid lineage (B, T and NK cells) [81]. | A study using a humanized mouse model of metastatic NB showed that the combinatory therapy of human γδ T cells, hu14.18 anti-GD2 antibody, and Fc-IL-7 was able to increase the survival rate of the subject animals [82]. |
| IL-10 | IL-10 is an immunosuppressive cytokine consistently expressed in the tumor microenvironment. Studies carried out in different tumor models have demonstrated that blocking the IL-10R relieves immunosuppression in the tumor microenvironment and reinstates immune response directed at malignant cells [83]. | This response was observed in an NB model wherein an antibody targeting the IL-10 receptor was used in combination with liposomal oligonucleotides to enhance the immune response. The observed immune response was larger compared with the use of oligonucleotides alone [84]. |
| IFN-β | Interferon-beta reduces myeloid dendritic concentrations in peripheral blood. It also alters the function of dendritic cells and other APCs to downregulate antigen presentation and the ability of APCs to stimulate T cell responses [85]. | IFN-β was found to increase the sensitivity of NB cells to the cytotoxic effects of the chemotherapy drug temozolomide (TMZ) through the mitigation of DNA repair enzyme (MGMT) expression [86]. |
| Identifier | Study Title | Phase | Start Date | Patients Enrolled | Status | Primary Aims |
|---|---|---|---|---|---|---|
| NCT02311621 | Engineered Neuroblastoma Cellular Immunotherapy (ENCIT)-01 | 1 | 2014 | 65 | Active, not recruiting | This is a phase 1 study designed to determine the maximum tolerated dose of the CAR+ T cells designed to recognize CD171 in patients with neuroblastoma. |
| NCT02914405 | Phase I Study of 131-I mIBG Followed by Nivolumab and Dinutuximab Beta Antibodies in Children with Relapsed/Refractory Neuroblastoma (MiniVan) | 1 | 2016 | 36 | Recruiting | To determine the safety and tolerability of the novel combination of 131-I-MIBG, ch14.18/CHO and nivolumab in pediatric patients. This is assessed by the nature, frequency, severity, and timing of adverse events, including serious adverse events and immune-related adverse events, during the administration of ch14.18/CHO. |
| NCT033633 | Naxitamab for High-Risk Neuroblastoma Patients with Primary Refractory Disease or Incomplete Response to Salvage Treatment in Bone and/or Bone Marrow | 2 | 2017 | 122 | Recruiting | The purpose of this study is to test the safety and efficacy of the combined therapy of naxitamab, a humanized monoclonal antibody targeting GD2 and GM-CSF in high-risk neuroblastoma patients. |
| NCT03294954 | GD2 Specific CAR and Interleukin-15 Expressing Autologous NKT Cells to Treat Children with Neuroblastoma (GINAKIT2) | 1 | 2017 | 36 | Active, not recruiting | The purpose of this study is to find the largest effective and safe dose of GD2-CAR NKT cells (GINAKIT cells), to evaluate their effect on the tumor, how long they can be detected in the patient’s blood and what affect they have on the patient’s neuroblastoma. |
| NCT04049864 | DNA Vaccination Against Neuroblastoma | 1 | 2019 | 12 | Recruiting | This is pilot open-label study to evaluate the safety and immunogenicity of a DNA vaccine strategy in relapsed neuroblastoma patients following chemotherapy and HSC transplantation. |
| NCT02395666 | Preventative Trial of Difluoromethylornithine (DFMO) in High-Risk Patients with Neuroblastoma That is in Remission | 2 | 2020 | 140 | Active, not recruiting | To evaluate the preventative activity of DFMO as a single agent in patients that are in remission based on event-free survival (EFS). |
| NCT04637503 | 4SCAR-T Therapy Targeting GD2, PSMA and CD276 for Treating Neuroblastoma | 1 and 2 | 2020 | 100 | Recruiting | The purpose of this clinical study is to assess the feasibility, safety and efficacy of the combinational GD2, PSMA and CD276 4SCAR-T cell therapy against NB. |
| NCT05027386 | Apatinib Mesylate Combined with IT Regimen for the Treatment of Recurrent or Refractory Pediatric Neuroblastoma | 2 | 2021 | 62 | Recruiting | The enrolled patients diagnosed with recurrent or refractory pediatric neuroblastoma received apatinib combined with IT regimen chemotherapy, the treatment includes a combination therapy phase and monotherapy maintenance phase. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera, Z.; Escutia, C.; Madonna, M.B.; Gupta, K.H. Biological Insight and Recent Advancement in the Treatment of Neuroblastoma. Int. J. Mol. Sci. 2023, 24, 8470. https://doi.org/10.3390/ijms24108470
Rivera Z, Escutia C, Madonna MB, Gupta KH. Biological Insight and Recent Advancement in the Treatment of Neuroblastoma. International Journal of Molecular Sciences. 2023; 24(10):8470. https://doi.org/10.3390/ijms24108470
Chicago/Turabian StyleRivera, Zoriamin, Carlos Escutia, Mary Beth Madonna, and Kajal H. Gupta. 2023. "Biological Insight and Recent Advancement in the Treatment of Neuroblastoma" International Journal of Molecular Sciences 24, no. 10: 8470. https://doi.org/10.3390/ijms24108470
APA StyleRivera, Z., Escutia, C., Madonna, M. B., & Gupta, K. H. (2023). Biological Insight and Recent Advancement in the Treatment of Neuroblastoma. International Journal of Molecular Sciences, 24(10), 8470. https://doi.org/10.3390/ijms24108470