

Emerging Mutual Regulatory Roles between m6A Modification and microRNAs


Abstract
1. Introduction
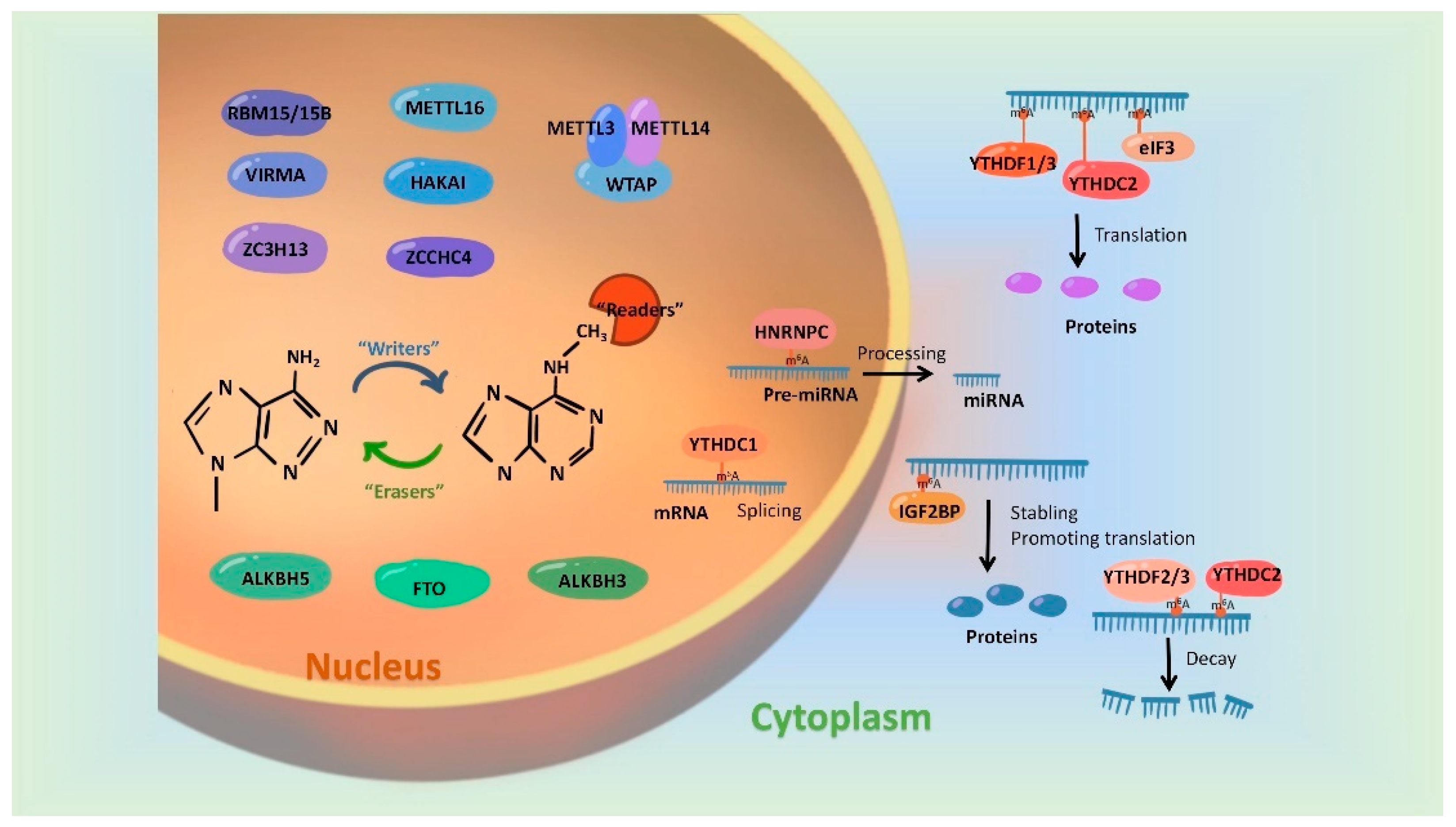
2. Dynamic Regulation of m6A Modification
2.1. “Writers”-m6A Methyltransferases
2.2. “Erasers”-m6A Demethylases
2.3. “Readers”-m6A Methyl-Binding Proteins
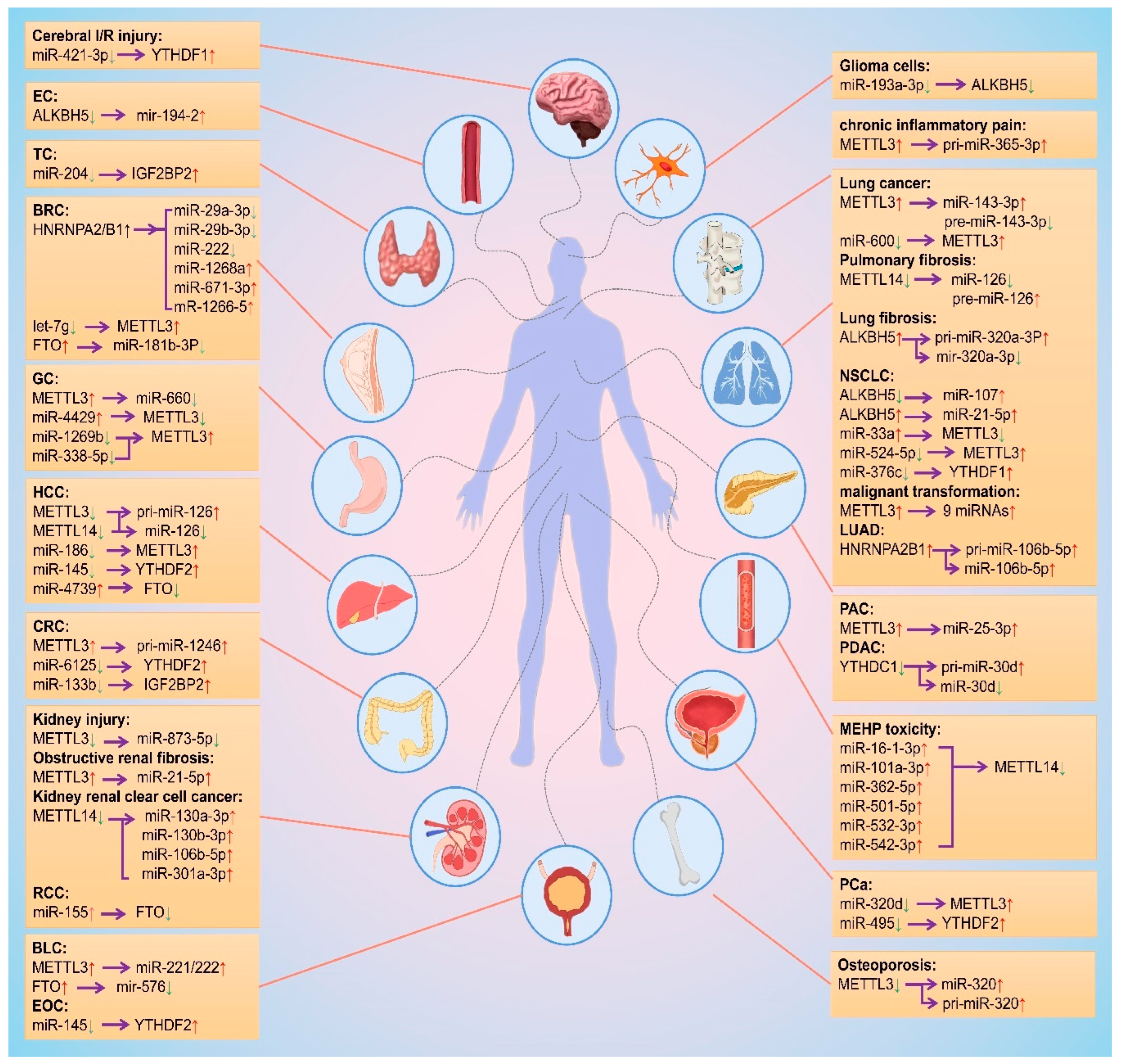
3. Mutual Regulatory Mechanisms between m6A Modification and miRNAs
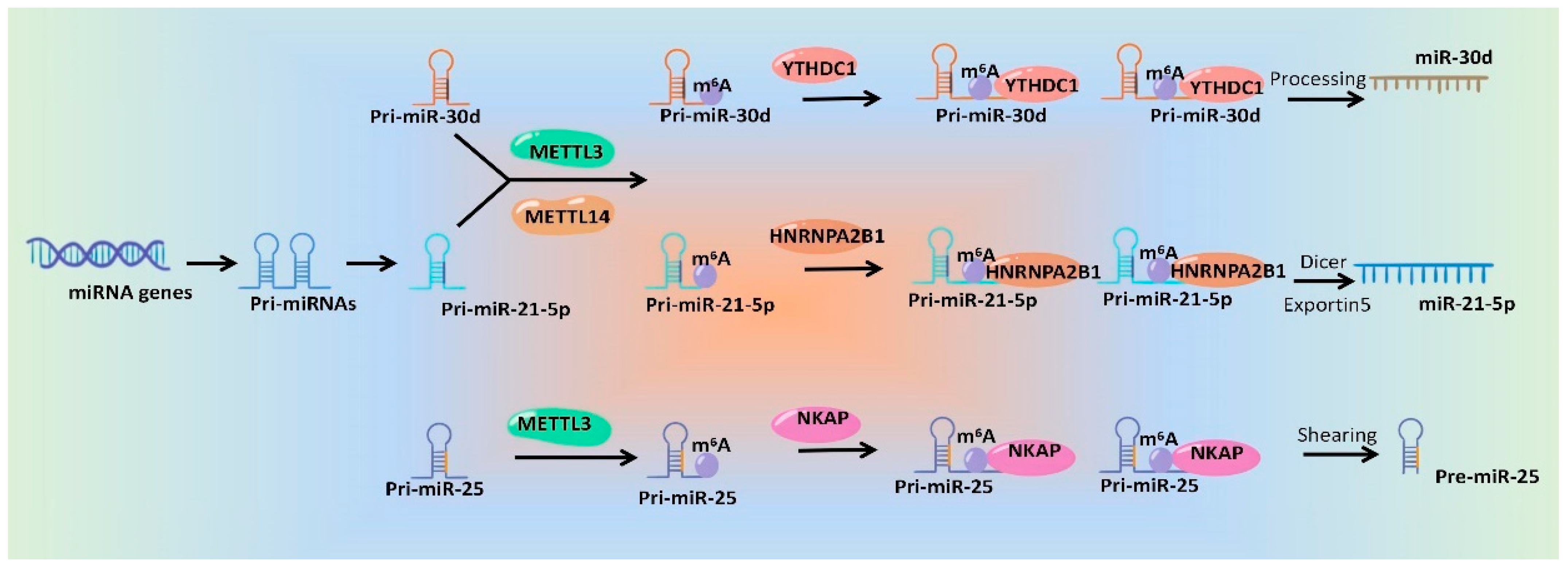
3.1. m6A Modifications Involves in miRNA Generation and Function
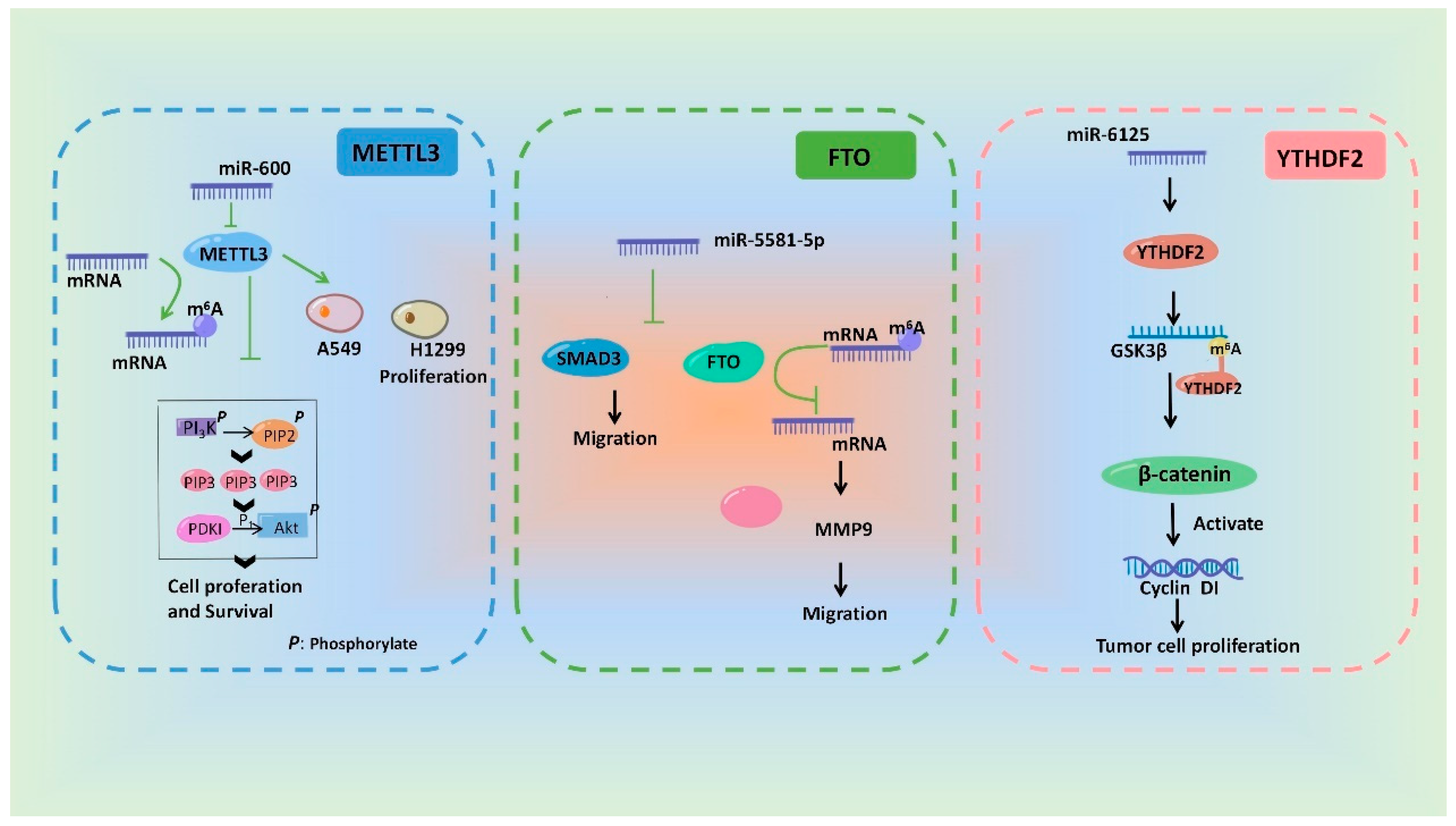
3.2. miRNAs Regulate the m6A Modification
4. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, J.; Wan, Z.; Tang, M.; Lin, Z.; Jiang, S.; Ji, L.; Gorshkov, K.; Mao, Q.; Xia, S.; Cen, D.; et al. N6-methyladenosine-modified CircRNA-SORE sustains sorafenib resistance in hepatocellular carcinoma by regulating β-catenin signaling. Mol. Cancer 2020, 19, 163. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lv, W.; Li, T.; Zhang, S.; Wang, H.; Li, X.; Wang, L.; Ma, D.; Zang, Y.; Shen, J.; et al. Dynamic regulation and functions of mRNA m6A modification. Cancer Cell Int. 2022, 22, 48. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m6A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Chen, J. The Biogenesis and Precise Control of RNA m6A Methylation. Trends Genet. 2020, 36, 44–52. [Google Scholar] [CrossRef]
- Sun, T.; Wu, R.; Ming, L. The role of m6A RNA methylation in cancer. Biomed. Pharmacother. 2019, 112, 108613. [Google Scholar] [CrossRef]
- Wei, W.; Ji, X.; Guo, X.; Ji, S. Regulatory Role of N6-methyladenosine (m6A) Methylation in RNA Processing and Human Diseases. J. Cell. Biochem. 2017, 118, 2534–2543. [Google Scholar] [CrossRef]
- Zepecki, J.P.; Karambizi, D.; Fajardo, J.E.; Snyder, K.M.; Guetta-Terrier, C.; Tang, O.Y.; Chen, J.S.; Sarkar, A.; Fiser, A.; Toms, S.A.; et al. miRNA-mediated loss of m6A increases nascent translation in glioblastoma. PLoS Genet. 2021, 17, e1009086. [Google Scholar] [CrossRef]
- Wiener, D.; Schwartz, S. The epitranscriptome beyond m6A. Nat. Rev. Genet. 2021, 22, 119–131. [Google Scholar] [CrossRef]
- Lin, C.; Ma, M.; Zhang, Y.; Li, L.; Long, F.; Xie, C.; Xiao, H.; Liu, T.; Tian, B.; Yang, K.; et al. Correction to: The N6-methyladenosine modification of circALG1 promotes the metastasis of colorectal cancer mediated by the miR-342-5p/PGF signalling pathway. Mol. Cancer 2022, 21, 80. [Google Scholar] [CrossRef]
- Khan, A.; Rehman, H.U.; Habib, U.; Ijaz, U. m6A-Finder: Detecting m6A methylation sites from RNA transcriptomes using physical and statistical properties based features. Comput. Biol. Chem. 2022, 97, 107640. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, Z.; Tao, B.; Yi, C.; Lin, Z.; Li, Y.; Shao, W.; Lin, J.; Chen, J. m6A target microRNAs in serum for cancer detection. Mol. Cancer 2021, 20, 170. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Chen, X.; Zhang, J.; Zhu, J. Novel insights into the interplay between m6A modification and noncoding RNAs in cancer. Mol. Cancer 2020, 19, 121. [Google Scholar] [CrossRef]
- Tang, F.; Chen, L.; Gao, H.; Xiao, D.; Li, X. m6A: An Emerging Role in Programmed Cell Death. Front. Cell Dev. Biol. 2022, 10, 817112. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, H.; Wang, Y.; Yuan, G.; Yu, X.; Jiang, H.; Wu, Q.; Yang, B.; Hu, Z.; Shi, F.; et al. MiR-103-3p targets the m6A methyltransferase METTL14 to inhibit osteoblastic bone formation. Aging Cell 2021, 20, e13298. [Google Scholar] [CrossRef]
- Yan, R.; Dai, W.; Wu, R.; Huang, H.; Shu, M. Therapeutic targeting m6A-guided miR-146a-5p signaling contributes to the melittin-induced selective suppression of bladder cancer. Cancer Lett. 2022, 534, 215615. [Google Scholar] [CrossRef]
- Han, B.; Chu, C.; Su, X.; Zhang, N.; Zhou, L.; Zhang, M.; Yang, S.; Shi, L.; Zhao, B.; Niu, Y.; et al. N6-methyladenosine-dependent primary microRNA-126 processing activated PI3K-AKT-mTOR pathway drove the development of pulmonary fibrosis induced by nanoscale carbon black particles in rats. Nanotoxicology 2020, 14, 1–20. [Google Scholar] [CrossRef]
- Chen, S.; Dong, J.; Luo, X.; Nie, Z.; Lu, S.; Liu, H.; Liu, J. Interaction between m6A and ncRNAs and Its Association with Diseases. Cytogenet. Genome Res. 2022, 162, 171–187. [Google Scholar] [CrossRef]
- Zeng, C.; Huang, W.; Li, Y.; Weng, H. Roles of METTL3 in cancer: Mechanisms and therapeutic targeting. J. Hematol. Oncol. 2020, 13, 117. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Kong, S.; Tao, M.; Ju, S. The potential role of RNA N6-methyladenosine in Cancer progression. Mol. Cancer 2020, 19, 88. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, J.; Wei, J.; Lu, C. Role of m6A methyltransferase component VIRMA in multiple human cancers. Cancer Cell Int. 2021, 21, 172. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Ren, H.; Ma, L.; Guo, J.; Mao, D.; Lu, Z.; Lu, L.; Yan, D. Role of Hakai in m6A modification pathway in Drosophila. Nat. Commun. 2021, 12, 2159. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, H.; Peng, H.; Yan, J.; Han, L.; Ye, G. ZC3H13 Inhibits the Progression of Hepatocellular Carcinoma through m6A-PKM2-Mediated Glycolysis and Enhances Chemosensitivity. J. Oncol. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Zhang, Z.; Mei, Y.; Hou, M. Knockdown RBM15 Inhibits Colorectal Cancer Cell Proliferation and Metastasis Via N6-Methyladenosine (m6A) Modification of MyD88 mRNA. Cancer Biother Radiopharm 2021. [Google Scholar] [CrossRef]
- Mendel, M.; Chen, K.; Homolka, D.; Gos, P.; Pandey, R.R.; McCarthy, A.A.; Pillai, R.S. Methylation of Structured RNA by the m6A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol. Cell 2018, 71, 986–1000. [Google Scholar] [CrossRef]
- van Tran, N.; Ernst, F.G.M.; Hawley, B.R.; Zorbas, C.; Ulryck, N.; Hackert, P.; Bohnsack, K.E.; Bohnsack, M.T.; Jaffrey, S.R.; Graille, M.; et al. The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112. Nucleic Acids Res. 2019, 47, 7719–7733. [Google Scholar] [CrossRef]
- Ma, H.; Wang, X.; Cai, J.; Dai, Q.; Natchiar, S.K.; Lv, R.; Chen, K.; Lu, Z.; Chen, H.; Shi, Y.G.; et al. N(6-)Methyladenosine methyltransferase ZCCHC4 mediates ribosomal RNA methylation. Nat. Chem. Biol. 2019, 15, 88–94. [Google Scholar] [CrossRef]
- Kim, H.; Jang, S.; Lee, Y. The m6A(m)-independent role of FTO in regulating WNT signaling pathways. Life Sci. Alliance 2022, 5, e202101250. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Feng, X.; Zhang, H.; Luo, Y.; Huang, J.; Lin, M.; Jin, J.; Ding, X.; Wu, S.; Huang, H.; et al. METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy 2019, 15, 1419–1437. [Google Scholar] [CrossRef] [PubMed]
- Mathiyalagan, P.; Adamiak, M.; Mayourian, J.; Sassi, Y.; Liang, Y.; Agarwal, N.; Jha, D.; Zhang, S.; Kohlbrenner, E.; Chepurko, E.; et al. FTO-Dependent N6-Methyladenosine Regulates Cardiac Function During Remodeling and Repair. Circulation 2019, 139, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, G.; Zhu, X.; Peng, T.; Lv, Y. Critical roles of FTO-mediated mRNA m6A demethylation in regulating adipogenesis and lipid metabolism: Implications in lipid metabolic disorders. Genes Dis. 2022, 9, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wei, J.; Cui, X.; Yu, C.; Ni, W.; Bungert, J.; Wu, L.; He, C.; Qian, Z. Post-translational modification of RNA m6A demethylase ALKBH5 regulates ROS-induced DNA damage response. Nucleic Acids Res. 2021, 49, 5779–5797. [Google Scholar] [CrossRef]
- Shen, D.; Suo, F.; Song, Q.; Chang, J.; Zhang, T.; Hong, J.; Zheng, Y.; Liu, H. Development of formaldehyde dehydrogenase-coupled assay and antibody-based assays for ALKBH5 activity evaluation. J. Pharm. Biomed. Anal. 2019, 162, 9–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Q.; Kaboli, P.J.; Shen, J.; Li, M.; Wu, X.; Yin, J.; Zhang, H.; Wu, Y.; Lin, L.; et al. m1A Regulated Genes Modulate PI3K/AKT/mTOR and ErbB Pathways in Gastrointestinal Cancer. Transl. Oncol. 2019, 12, 1323–1333. [Google Scholar] [CrossRef]
- Chen, Z.; Qi, M.; Shen, B.; Luo, G.; Wu, Y.; Li, J.; Lu, Z.; Zheng, Z.; Dai, Q.; Wang, H. Transfer RNA demethylase ALKBH3 promotes cancer progression via induction of tRNA-derived small RNAs. Nucleic Acids Res. 2019, 47, 2533–2545. [Google Scholar] [CrossRef]
- Wu, Y.; Zhan, S.; Xu, Y.; Gao, X. RNA modifications in cardiovascular diseases, the potential therapeutic targets. Life Sci. 2021, 278, 119565. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Kasowitz, S.D.; Homolka, D.; Leu, N.A.; Shaked, J.T.; Ruthel, G.; Jain, D.; Lin, H.; Keeney, S.; Luo, M.; et al. YTHDC2 is essential for pachytene progression and prevents aberrant microtubule-driven telomere clustering in male meiosis. Cell Rep. 2021, 37, 110110. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.; Ott, S.; Joseph, B.; Park, E.S.; Dai, W.; Kleiner, R.E.; Claridge-Chang, A.; Lai, E.C. A neural m6A/Ythdf pathway is required for learning and memory in Drosophila. Nat. Commun. 2021, 12, 1458. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xie, R.; Wei, Q. Network analysis of miRNA targeting m6A-related genes in patients with esophageal cancer. PeerJ 2021, 9, e11893. [Google Scholar] [CrossRef]
- Widagdo, J.; Anggono, V.; Wong, J.J. The multifaceted effects of YTHDC1-mediated nuclear m6A recognition. Trends Genet. 2022, 38, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, S.L. The roles of hnRNP A2/B1inRNA biology and disease. WIREs RNA 2021, 12, e1612. [Google Scholar] [CrossRef]
- Gomes-Duarte, A.; Lacerda, R.; Menezes, J.; Romão, L. eIF3: A factor for human health and disease. RNA Biol. 2018, 15, 26–34. [Google Scholar] [CrossRef]
- Coots, R.A.; Liu, X.; Mao, Y.; Dong, L.; Zhou, J.; Wan, J.; Zhang, X.; Qian, S. m6A Facilitates eIF4F-Independent mRNA Translation. Mol. Cell 2017, 68, 504–514. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Sun, L.; Wan, A.; Zhou, Z.; Chen, D.; Liang, H.; Liu, C.; Yan, S.; Niu, Y.; Lin, Z.; Zhan, S.; et al. RNA-binding protein RALY reprogrammes mitochondrial metabolism via mediating miRNA processing in colorectal cancer. Gut 2021, 70, 1698–1712. [Google Scholar] [CrossRef]
- Zhou, K.I.; Parisien, M.; Dai, Q.; Liu, N.; Diatchenko, L.; Sachleben, J.R.; Pan, T. N6-Methyladenosine Modification in a Long Noncoding RNA Hairpin Predisposes Its Conformation to Protein Binding. J. Mol. Biol. 2016, 428, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhou, K.I.; Parisien, M.; Dai, Q.; Diatchenko, L.; Pan, T. N 6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017, 45, 6051–6063. [Google Scholar] [CrossRef] [PubMed]
- Edupuganti, R.R.; Geiger, S.; Lindeboom, R.G.H.; Shi, H.; Hsu, P.J.; Lu, Z.; Wang, S.; Baltissen, M.P.A.; Jansen, P.W.T.C.; Rossa, M.; et al. N6-methyladenosine (m6A) recruits and repels proteins to regulate mRNA homeostasis. Nat. Struct. Mol. Biol. 2017, 24, 870–878. [Google Scholar] [CrossRef]
- Dai, D.; Wang, H.; Zhu, L.; Jin, H.; Wang, X. N6-methyladenosine links RNA metabolism to cancer progression. Cell Death Dis. 2018, 9, 113–124. [Google Scholar] [CrossRef]
- Afonso-Grunz, F.; Müller, S. Principles of miRNA–mRNA interactions: Beyond sequence complementarity. Cell. Mol. Life Sci. 2015, 72, 3127–3141. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, R.; Li, M.; Ye, H.; Wu, C.; Wang, C.; Li, S.; Tan, L.; Mai, D.; Li, G.; et al. Excessive miR-25-3p maturation via N6-methyladenosine stimulated by cigarette smoke promotes pancreatic cancer progression. Nat. Commun. 2019, 10, 1858. [Google Scholar] [CrossRef]
- Zailaie, S.A.; Sergi, C.M. MiR-126 in Hepatocellular Carcinoma and Cholangiocellular Carcinoma: A Reappraisal with an in situ Detection of miR-126. Ann. Clin. Lab. Sci. 2022, 52, 73–85. [Google Scholar]
- Wang, J.; Ishfaq, M.; Xu, L.; Xia, C.; Chen, C.; Li, J. METTL3/m6A/miRNA-873-5p Attenuated Oxidative Stress and Apoptosis in Colistin-Induced Kidney Injury by Modulating Keap1/Nrf2 Pathway. Front. Pharmacol. 2019, 10, 517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Y.; Peng, Y.; Xu, H.; Zhou, X. METTL3 regulates inflammatory pain by modulating m6A-dependent pri-miR-365-3p processing. FASEB J. 2020, 34, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, J.; Yang, X.; Yu, H.; Zhou, R.; Lu, H.; Yuan, W.; Lu, J.; Zhou, Z.; Lu, Q.; et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol. Cancer 2019, 18, 110. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, W.; Li, R.; Liu, Y. miRNA-93-5p in exosomes derived from M2 macrophages improves lipopolysaccharide-induced podocyte apoptosis by targeting Toll-like receptor 4. Bioengineered 2022, 13, 7683–7696. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Lv, L.; Zhan, Y.; Ma, Y.; Feng, J.; He, Y.; Wen, Y.; Zhang, Y.; Pu, Q.; Ji, F.; et al. METTL3/N6-methyladenosine/ miR-21-5p promotes obstructive renal fibrosis by regulating inflammation through SPRY1/ERK/NF-κB pathway activation. J. Cell. Mol. Med. 2021, 25, 7660–7674. [Google Scholar] [CrossRef]
- Li, S.; Lu, X.; Zheng, D.; Chen, W.; Li, Y.; Li, F. Methyltransferase-like 3 facilitates lung cancer progression by accelerating m6A methylation-mediated primary miR-663 processing and impeding SOCS6 expression. J. Cancer Res. Clin. Oncol. 2022, 148, 3485–3499. [Google Scholar] [CrossRef]
- Wang, H.; Deng, Q.; Lv, Z.; Ling, Y.; Hou, X.; Chen, Z.; Dinglin, X.; Ma, S.; Li, D.; Wu, Y.; et al. N6-methyladenosine induced miR-143-3p promotes the brain metastasis of lung cancer via regulation of VASH1. Mol. Cancer 2019, 18, 181. [Google Scholar] [CrossRef]
- Yan, G.; Yuan, Y.; He, M.; Gong, R.; Lei, H.; Zhou, H.; Wang, W.; Du, W.; Ma, T.; Liu, S.; et al. m6A Methylation of Precursor-miR-320/RUNX2 Controls Osteogenic Potential of Bone Marrow-Derived Mesenchymal Stem Cells. Mol. Ther. Nucleic Acids 2020, 19, 421–436. [Google Scholar] [CrossRef]
- Peng, W.; Li, J.; Chen, R.; Gu, Q.; Yang, P.; Qian, W.; Ji, D.; Wang, Q.; Zhang, Z.; Tang, J.; et al. Upregulated METTL3 promotes metastasis of colorectal Cancer via miR-1246/SPRED2/MAPK signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 393. [Google Scholar] [CrossRef]
- Gu, S.; Sun, D.; Dai, H.; Zhang, Z. N6-methyladenosine mediates the cellular proliferation and apoptosis via microRNAs in arsenite-transformed cells. Toxicol. Lett. 2018, 292, 1–11. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, H.; Chen, Q.; Wan, Z.; Gao, X.; Qian, W. Identification of METTL14 in Kidney Renal Clear Cell Carcinoma Using Bioinformatics Analysis. Dis. Markers 2019, 2019, 5648783. [Google Scholar] [CrossRef] [PubMed]
- Berulava, T.; Rahmann, S.; Rademacher, K.; Klein-Hitpass, L.; Horsthemke, B. N6-Adenosine Methylation in MiRNAs. PLoS ONE 2015, 10, e118438. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ye, S.; Zhang, N.; Zheng, S.; Liu, H.; Zhou, K.; Wang, L.; Cao, Y.; Sun, P.; Wang, T. The FTO/miR-181b-3p/ARL5B signaling pathway regulates cell migration and invasion in breast cancer. Cancer Commun. 2020, 40, 484–500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yan, K.; Liu, J.; Gao, L.; Jiang, X.; Fan, Y. FTO promotes tumour proliferation in bladder cancer via the FTO/miR-576/CDK6 axis in an m6A-dependent manner. Cell Death Discov. 2021, 7, 329. [Google Scholar] [CrossRef]
- Jin, D.; Guo, J.; Wu, Y.; Yang, L.; Wang, X.; Du, J.; Dai, J.; Chen, W.; Gong, K.; Miao, S.; et al. m6A demethylase ALKBH5 inhibits tumor growth and metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2–mediated YAP activity in NSCLC. Mol. Cancer 2020, 19, 40. [Google Scholar] [CrossRef]
- Chen, P.; Li, S.; Zhang, K.; Zhao, R.; Cui, J.; Zhou, W.; Liu, Y.; Zhang, L.; Cheng, Y. N6-methyladenosine demethylase ALKBH5 suppresses malignancy of esophageal cancer by regulating microRNA biogenesis and RAI1 expression. Oncogene 2021, 40, 5600–5612. [Google Scholar] [CrossRef]
- Wang, H.; Song, X.; Song, C.; Wang, X.; Cao, H. m6A-seq analysis of microRNAs reveals that the N6-methyladenosine modification of miR-21–5p affects its target expression. Arch. Biochem. Biophys. 2021, 711, 109023. [Google Scholar] [CrossRef]
- Sun, W.; Li, Y.; Ma, D.; Liu, Y.; Xu, Q.; Cheng, D.; Li, G.; Ni, C. ALKBH5 promotes lung fibroblast activation and silica-induced pulmonary fibrosis through miR-320a-3p and FOXM1. Cell Mol. Biol. Lett. 2022, 27, 26. [Google Scholar] [CrossRef]
- Hou, Y.; Zhang, Q.; Pang, W.; Hou, L.; Liang, Y.; Han, X.; Luo, X.; Wang, P.; Zhang, X.; Li, L.; et al. YTHDC1-mediated augmentation of miR-30d in repressing pancreatic tumorigenesis via attenuation of RUNX1-induced transcriptional activation of Warburg effect. Cell Death Differ. 2021, 28, 3105–3124. [Google Scholar] [CrossRef]
- Jiang, L.; Lin, W.; Zhang, C.; Ash, P.E.A.; Verma, M.; Kwan, J.; van Vliet, E.; Yang, Z.; Cruz, A.L.; Boudeau, S.; et al. Interaction of tau with HNRNPA2B1 and N6-methyladenosine RNA mediates the progression of tauopathy. Mol. Cell 2021, 81, 4209–4227. [Google Scholar] [CrossRef]
- Klinge, C.M.; Piell, K.M.; Tooley, C.S.; Rouchka, E.C. HNRNPA2/B1 is upregulated in endocrine-resistant LCC9 breast cancer cells and alters the miRNA transcriptome when overexpressed in MCF-7 cells. Sci. Rep. 2019, 9, 9430. [Google Scholar] [CrossRef] [PubMed]
- Luo, K. Signaling Cross Talk between TGF-beta/Smad and Other Signaling Pathways. Cold Spring Harb. Perspect. Biol. 2017, 9, a022137. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Liu, F.; Wang, D.; Fang, C.; Tang, X.; Guo, B.; Shi, Z.; Dong, Z.; Guo, D.; Yue, J.; et al. Exosomal Transfer of miR-185 Is Controlled by hnRNPA2B1 and Impairs Re-endothelialization After Vascular Injury. Front. Cell Dev. Biol. 2021, 9, 619444. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Xu, Y.; Zhang, K.; Jin, L.; Liu, X. HNRNPA2B1 inhibited SFRP2 and activated Wnt-beta/catenin via m6A-mediated miR-106b-5p processing to aggravate stemness in lung adenocarcinoma. Pathol. Res. Pract. 2022, 233, 153794. [Google Scholar] [CrossRef]
- Li, K.; Gao, S.; Ma, L.; Sun, Y.; Peng, Z.; Wu, J.; Du, N.; Ren, H.; Tang, S.; Sun, X. Stimulation of Let-7 Maturation by Metformin Improved the Response to Tyrosine Kinase Inhibitor Therapy in an m6A Dependent Manner. Front. Oncol. 2022, 11, 731561. [Google Scholar] [CrossRef]
- Müller, S.; Glaß, M.; Singh, A.K.; Haase, J.; Bley, N.; Fuchs, T.; Lederer, M.; Dahl, A.; Huang, H.; Chen, J.; et al. IGF2BP1 promotes SRF-dependent transcription in cancer in a m6A- and miRNA-dependent manner. Nucleic Acids Res. 2019, 47, 375–390. [Google Scholar] [CrossRef]
- He, X.; Shu, Y. RNA N6-methyladenosine modification participates in miR-660/E2F3 axis-mediated inhibition of cell proliferation in gastric cancer. Pathol.-Res. Pract. 2019, 215, 152393. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, Y.; Shu, Y.; He, J.; Gao, W. Interaction between N6-methyladenosine (m6A) modification and noncoding RNAs in cancer. Mol. Cancer 2020, 19, 84. [Google Scholar] [CrossRef]
- Ma, J.Z.; Yang, F.; Zhou, C.C.; Liu, F.; Yuan, J.H.; Wang, F.; Wang, T.T.; Xu, Q.G.; Zhou, W.P.; Sun, S.H. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N6 -methyladenosine-dependent primary MicroRNA processing. Hepatology 2017, 65, 529–543. [Google Scholar] [CrossRef]
- Kang, J.; Huang, X.; Dong, W.; Zhu, X.; Li, M.; Cui, N. MicroRNA-1269b inhibits gastric cancer development through regulating methyltransferase-like 3 (METTL3). Bioengineered 2021, 12, 1150–1160. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, Z.; Xia, C. Long non-coding RNA LINC00240 promotes gastric cancer progression via modulating miR-338-5p/METTL3 axis. Bioengineered 2021, 12, 9678–9691. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhang, Y.; Mao, Y.; Mou, J.; Zhao, J.; Xue, Q.; Wang, D.; Huang, J.; Gao, S.; Gao, Y. MiR-33a suppresses proliferation of NSCLC cells via targeting METTL3 mRNA. Biochem. Biophys. Res. Commun. 2017, 482, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Huo, B.; Shi, X. miR-600 inhibits lung cancer via downregulating the expression of METTL3. Cancer Manag. Res. 2019, 11, 1177–1187. [Google Scholar] [CrossRef]
- Xie, H.; Yao, J.; Wang, Y.; Ni, B. Exosome-transmitted circVMP1 facilitates the progression and cisplatin resistance of non-small cell lung cancer by targeting miR-524-5p-METTL3/SOX2 axis. Drug Deliv. 2022, 29, 1257–1271. [Google Scholar] [CrossRef]
- He, H.; Wu, W.; Sun, Z.; Chai, L. MiR-4429 prevented gastric cancer progression through targeting METTL3 to inhibit m6A-caused stabilization of SEC62. Biochem. Biophys. Res. Commun. 2019, 517, 581–587. [Google Scholar] [CrossRef]
- Cai, X.; Wang, X.; Cao, C.; Gao, Y.; Zhang, S.; Yang, Z.; Liu, Y.; Zhang, X.; Zhang, W.; Ye, L. HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tumor suppressor let-7g. Cancer Lett. 2018, 415, 11–19. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Z.; Li, J.; Zhu, J.; Ren, Z.; Zhang, D.; Zhao, W.; Fan, Y.; Zhang, D.; Sun, R. Cross talk between RNA N6-methyladenosine methyltransferase-like 3 and miR-186 regulates hepatoblastoma progression through Wnt/β-catenin signalling pathway. Cell Prolif. 2020, 53, e12768. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, F.; Zhong, Q.; Hou, J. METTL3-mediated m6A modification of KIF3C-mRNA promotes prostate cancer progression and is negatively regulated by miR-320d. Aging 2021, 13, 22332–22344. [Google Scholar] [CrossRef]
- Park, M.H.; Jeong, E.; Choudhury, M. Mono-(2-Ethylhexyl)phthalate Regulates Cholesterol Efflux via MicroRNAs Regulated m6A RNA Methylation. Chem. Res. Toxicol. 2020, 33, 461–469. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, Q.; Lin, J.; Zhang, L.; Zhang, C.; Chen, H.; Qian, J.; Luo, C. miRNA-193a-3p Regulates the AKT2 Pathway to Inhibit the Growth and Promote the Apoptosis of Glioma Cells by Targeting ALKBH5. Front. Oncol. 2021, 11, 600451. [Google Scholar] [CrossRef]
- Liu, L.; Gu, M.; Ma, J.; Wang, Y.; Li, M.; Wang, H.; Yin, X.; Li, X. CircGPR137B/miR-4739/FTO feedback loop suppresses tumorigenesis and metastasis of hepatocellular carcinoma. Mol. Cancer 2022, 21, 149. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Xie, L.; Wang, P.; Zhuang, C. MiR-155 regulates m6A level and cell progression by targeting FTO in clear cell renal cell carcinoma. Cell. Signal. 2022, 91, 110217. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Tang, X.; Lu, M.; Sun, S.; Xie, S.; Cai, J.; Zan, J. microRNA-421-3p prevents inflammatory response in cerebral ischemia/reperfusion injury through targeting m6A Reader YTHDF1 to inhibit p65 mRNA translation. Int. Immunopharmacol. 2020, 88, 106937. [Google Scholar] [CrossRef]
- Zhou, J.; Xiao, D.; Qiu, T.; Li, J.; Liu, Z. Loading MicroRNA-376c in Extracellular Vesicles Inhibits Properties of Non-Small Cell Lung Cancer Cells by Targeting YTHDF1. Technol. Cancer Res. Treat. 2020, 19, 1180565672. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.; Feng, G.; Gao, S.; Wang, Y.; Zhang, S.; Liu, Y.; Ye, L.; Li, Y.; Zhang, X. MicroRNA-145 Modulates N6-Methyladenosine Levels by Targeting the 3′-Untranslated mRNA Region of the N6-Methyladenosine Binding YTH Domain Family 2 Protein. J. Biol. Chem. 2017, 292, 3614–3623. [Google Scholar] [CrossRef]
- Li, J.; Wu, L.; Pei, M.; Zhang, Y. YTHDF2, a protein repressed by miR-145, regulates proliferation, apoptosis, and migration in ovarian cancer cells. J. Ovarian Res. 2020, 13, 111. [Google Scholar] [CrossRef]
- Du, C.; Lv, C.; Feng, Y.; Yu, S. Activation of the KDM5A/miRNA-495/YTHDF2/m6A-MOB3B axis facilitates prostate cancer progression. J. Exp. Clin. Cancer Res. 2020, 39, 223. [Google Scholar] [CrossRef]
- Li, H.; Zhang, N.; Jiao, X.; Wang, C.; Sun, W.; He, Y.; Ren, G.; Huang, S.; Li, M.; Chang, Y.; et al. Downregulation of microRNA-6125 promotes colorectal cancer growth through YTHDF2-dependent recognition of N6-methyladenosine-modified GSK3β. Clin. Transl. Med. 2021, 11, e602. [Google Scholar] [CrossRef]
- Hao, J.; Hu, H.; Jiang, Z.; Yu, X.; Li, C.; Chen, L.; Xia, Y.; Liu, D.; Wang, D. microRNA-670 modulates Igf2bp1 expression to regulate RNA methylation in parthenogenetic mouse embryonic development. Sci. Rep. 2020, 10, 4782. [Google Scholar] [CrossRef]
- Ye, M.; Dong, S.; Hou, H.; Zhang, T.; Shen, M. Oncogenic Role of Long Noncoding RNAMALAT1 in Thyroid Cancer Progression through Regulation of the miR-204/IGF2BP2/m6A-MYC Signaling. Mol. Ther.-Nucleic Acids 2021, 23, 1–12. [Google Scholar] [CrossRef]
- Yao, B.; Zhang, Q.; Yang, Z.; An, F.; Nie, H.; Wang, H.; Yang, C.; Sun, J.; Chen, K.; Zhou, J.; et al. CircEZH2/miR-133b/IGF2BP2 aggravates colorectal cancer progression via enhancing the stability of m6A-modified CREB1 mRNA. Mol. Cancer 2022, 21, 140. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hao, Y.; Zhang, Y.; Li, M.; Wang, M.; Han, W.; Wu, Y.; Lv, Y.; Hao, J.; Wang, L.; et al. m6A RNA Methylation Is Regulated by MicroRNAs and Promotes Reprogramming to Pluripotency. Cell Stem Cell 2015, 16, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ma, X.; Ying, Y.; Wang, W.; Shen, H.; Wang, S.; Xie, H.; Yi, J.; Zhan, W.; Li, J.; et al. SMAD3 and FTO are involved in miR-5581-3p-mediated inhibition of cell migration and proliferation in bladder cancer. Cell Death Discov. 2022, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Nagahashi, M.; Shimada, Y.; Ichikawa, H.; Kameyama, H.; Takabe, K.; Okuda, S.; Wakai, T. Next generation sequencing-based gene panel tests for the management of solid tumors. Cancer Sci. 2019, 110, 6–15. [Google Scholar] [CrossRef]
- Sarkar, A.; Gasperi, W.; Begley, U.; Nevins, S.; Huber, S.M.; Dedon, P.C.; Begley, T.J. Detecting the epitranscriptome. Wiley Interdiscip. Rev. RNA 2021, 12, e1663. [Google Scholar] [CrossRef]
- Wei, J.; Yu, X.; Yang, L.; Liu, X.; Gao, B.; Huang, B.; Dou, X.; Liu, J.; Zou, Z.; Cui, X.L.; et al. FTO mediates LINE1 m6A demethylation and chromatin regulation in mESCs and mouse development. Science 2022, 376, 968–973. [Google Scholar] [CrossRef]
- Selmi, T.; Lanzuolo, C. Driving Chromatin Organisation through N6-methyladenosine Modification of RNA: What Do We Know and What Lies Ahead? Genes 2022, 13, 340. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regulatory Proteins of m6A | m6A on RNAs | miRNAs Affected by m6A | Mechanisms of m6A Mediating miRNAs | Diseases | Ref. |
|---|---|---|---|---|---|
| METTL14↓ | pri-miR-126↑ | miR-126↓ | M6A modification reduced recognition of pri-miR-126 by DGCR8 | Pulmonary fibrosis | [18] |
| METTL3↓ | - | 70% miRNAs↓ | METTL3 catalyzed m6A modification on pri- miRNAs, thus promoting them for recognition and processing by DGCR8 | - | [57] |
| METTL3↑ | - | miR-25-3p↑ | METTL3 catalyzed the formation of m6A modification on pri-miR-25 to promote its processing via NKAP | Pancreatic cancer | [59] |
| METTL3↓METTL14↓ | pri-miR-126↑ | miR-126↓ | Reduction of m6A modification weakened recognition of pri-miR-126 by DGCR8 | Hepatocellular carcinoma | [60] |
| METTL3↓ | - | miR-873-5p↓ | METTL3 interacted with DGCR8 and positively modulates maturation of miR-873-5p | Kidney injury | [61] |
| METTL3↑ | pri-miR-365-3p↑ | - | METTL3 positively affected the interaction between DGCR8 and pri-miR-365 | Chronic inflammatory pain | [62] |
| METTL3↑ | pri-miR-221/222↑ | - | METTL3 positively modulated the pri-miR221/222 processing through interacting with DGCR8 | Bladder cancer | [63] |
| METTL3↑ | - | miR-21-5p↑ | Up-regulation of METTL3 promoted maturation of miRNA-21-5p | Obstructive renal fibrosis | [65] |
| METTL3↑ | pre-miR-143-3p↓ | miR-143-3p↑ | Knockdown of dicer can abolish METTL3-induced up-regulation of miR-143-3p | Lung cancer | [67] |
| METTL3↓ | pre-miR-320↑ | miR-320↑ | METTL3 enhanced the m6A methylations on pre-miR-320 | Osteoporosis | [68] |
| METTL14↓ | - | miR-130a-3p↑ miR-130b-3p↑ miR-106b-5p↑ miR-301a-3p↑ | MiRNA levels negative correlation with METTL14 mRNA | Kidney renal clear cell carcinoma | [71] |
| METTL3↑ | pri-miR-1246↑ | - | METTL3 methylated and promoted pri-miR-1246 maturation | Colorectal cancer | [69] |
| METTL3↑ | Total RNAs | 9 miRNAs↑ | miRNA level positive correlation with METTL3 level | Malignant transformation | [70] |
| METTL3↑ | E2F3 mRNA | miR-660↓ | m6A-modified E2F3 mRNA was required for the interaction between miR-660 and METTL3 | Gastric cancer | [87] |
| ALKBH5↓ | - | miR-107↑ | miR-107 level negative correlation with ALKBH5 | Non-small cell lung cancer | [75] |
| ALKBH5↓ | - | miR-194-2↑ | ALKBH5 inhibited miR-194-2 biogenesis through an m6A/DGCR8-dependent manner | Esophageal cancer malignancy | [76] |
| ALKBH5↑ | - | miR-21-5p↑ | ALKBH5 changed the miR-21-5p silencing potency by altered the m6A abundance | Non-small cell lung cancer cells | [77] |
| ALKBH5↑ | Pri-miR-320a-3p↑ | miR-320a-3p↓ | ALKBH5 promoted lung fibrosis via the miR-320a-3p/FOXM1 axis or targeting FOXM1 | Lung fibrosis | [78] |
| FTO↓ | miRNAs | 42 miRNAs↑9 miRNAs↓ | FTO affected the modification of m6A on mature miRNA | - | [72] |
| FTO↑ | - | miR-181b-3p↓ | FTO promoting breast cancer via the FTO/miR-181b-3p/ARL5B pathway | Breast cancer | [73] |
| FTO↑ | - | miR-576↓ | FTO promoted bladder cancer cell proliferation via the FTO/miR-576/CDK6 pathways | Bladder cancer | [74] |
| HNRNPA2B1↑ | - | miR-29a-3↓miR-29b-3↓miR-222↓ miR-1266-5↑miR-1268a↑miR-671-3p↑ | HNRNPA2/B1 promoteed DROSHA processing to pre-miRNAs | Breast cancer | [81] |
| HNRNPA2B1↑ | pri-miR-106b-5p↑ | MiR-106b-5p↑ | HNRNPA2B1 inhibited SFRP2 and activated Wnt-β/catenin via m6A-mediated miR-106b-5p processing to aggravate steaminess in lung adenocarcinoma | Lung adenocarcino-ma | [84] |
| IGF2BP1 ↑ | - | miR-22-3p↓miR-125-5p↓miR-181-5p↓, etc. | IGF2BP1 impaired the miRNA-directed down regulation of SRF expression | Ovarian, liver and lung cancer | [86] |
| YTHDC1↓ | Pri-miR-30d↑ | miR-30d↓ | YTHDC1 induced miR-30d function as a tumor suppressor gene by negatively regulating RUNX1 and its downstream glycolytic genes including HK1 and SLC2A1 | Pancreatic ductal Adenocarcin-oma | [79] |
| miRNAs | Regulatory Proteins of m6A | Mechanisms of miRNAs Regulate m6A Level | Diseases | Refs. |
|---|---|---|---|---|
| miR-33a↑ | METTL3↓ | MiR-33a targeted to the 3′-UTR of METTL3 mRNA | Non-small cell lung cancer | [92] |
| miR-600↓ | METTL3↑ | MiR-600 inhibited the expression of METTL3 | Lung cancer | [93] |
| miR-4429↑ | METTL3↓ | MiR-4429 targeted and inhibited METTL3 level | Gastric cancer | [95] |
| let-7g↓ | METTL3↑ | Suppressing let-7g thus up-regulating METTL3 | Breast cancer | [96] |
| miR-186↓ | METTL3↑ | Low level of miR-186 led to high expression of METTL3 | Hepatocellular carcinoma | [97] |
| miR-320d↓ | METTL3↑ | MiR-320d inhibits KIF3C expression by targeting METTL3 | Prostate cancer | [98] |
| miR-1269b↓mir-338-5p↓ | METTL3↑ | METTL3 was the direct target of miR-1269b and miR-338-5p | Gastric cancer | [90,91] |
| miR-524-5P↓ | METTL3↑ | CircVMP1 plays an oncogenic role by targeting miR-524-5p-METTL3/SOX2 axis in A549/DDP and H1299/DDP cells. | Non-small cell lung cancer | [94] |
| miR-16-1-3p↑miR-101a-3p↑miR-362-5p↑miR-501-5p↑ miR-532-3p↑miR-542-3p↑ | METTL14↓ | MiRNAs were all predicted to regulate METTL14 mRNA | MEHP toxicity | [99] |
| miR-193a-3p↓ | ALKBH5↓ | MiR-193a-3p targeted to ALKBH5 by suppressing the AKT2 pathway | Glioma cells | [100] |
| miR-4739↑ | FTO↓ | CircGPR137B inhibits HCC tumorigenesis and metastasis through the circGPR137B/miR-4739/FTO feedback loop | Hepatocellular carcinoma | [101] |
| miR-155↑ | FTO↓ | MiR-155 directly targeted FTO and negatively regulated its expression and increased m6A level | Renal cell carcinoma | [102] |
| miR-421-3p↓ | YTHDF1↑ | MiR-421-3p suppresses inflammatory response during cerebral I/R injury through regulating YTHDF1/P65 | Cerebral I/R injury | [103] |
| miR-376c↓ | YTHDF1↑ | YTHDF1 overexpression reversed the inhibitory role of miR-376c released by EC-Evs in NSCLC cells | Non-small cell lung cancer | [104] |
| miR-145↓ | YTHDF2↑ | MiR-145 decreased the luciferase activities of 3′-UTR of YTHDF2 mRNA | Hepatocellular carcinoma | [105] |
| miR-145↓ | YTHDF2↑ | YTHDF2 was the direct target gene of miR-145 | Epithelial ovarian cancer | [106] |
| miR-6125↓ | YTHDF2↑ | MiR-6125 targeted the 3′-UTR of YTHDF2 and downregulated the YTHDF2 protein | Colorectal cancer | [108] |
| miR-495↓ | YTHDF2↑ | MiR-495 can inhibit the proliferation, invasion and migration of PCa cells by targeting YTHDF2 | Prostate cancer | [107] |
| miR-204↓ | IGF2BP2↑ | IGF2BP2 was a target of miR-204. | Thyroid cancer | [110] |
| miR-133b↓ | IGF2BP2↑ | CircEZH2 interacted with IGF2BP2 and served as a sponge of miR-133b | Colorectal cancer | [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, Z.; Mou, Y.; Zhang, N.; Liu, X.; He, Z.; Gu, S. Emerging Mutual Regulatory Roles between m6A Modification and microRNAs. Int. J. Mol. Sci. 2023, 24, 773. https://doi.org/10.3390/ijms24010773
Mei Z, Mou Y, Zhang N, Liu X, He Z, Gu S. Emerging Mutual Regulatory Roles between m6A Modification and microRNAs. International Journal of Molecular Sciences. 2023; 24(1):773. https://doi.org/10.3390/ijms24010773
Chicago/Turabian StyleMei, Zongqin, Yahao Mou, Nan Zhang, Xiaoyu Liu, Zuoshun He, and Shiyan Gu. 2023. "Emerging Mutual Regulatory Roles between m6A Modification and microRNAs" International Journal of Molecular Sciences 24, no. 1: 773. https://doi.org/10.3390/ijms24010773
APA StyleMei, Z., Mou, Y., Zhang, N., Liu, X., He, Z., & Gu, S. (2023). Emerging Mutual Regulatory Roles between m6A Modification and microRNAs. International Journal of Molecular Sciences, 24(1), 773. https://doi.org/10.3390/ijms24010773