
Inhibitory Effects of Ursolic Acid on the Stemness and Progression of Human Breast Cancer Cells by Modulating Argonaute-2

Abstract
1. Introduction
2. Results
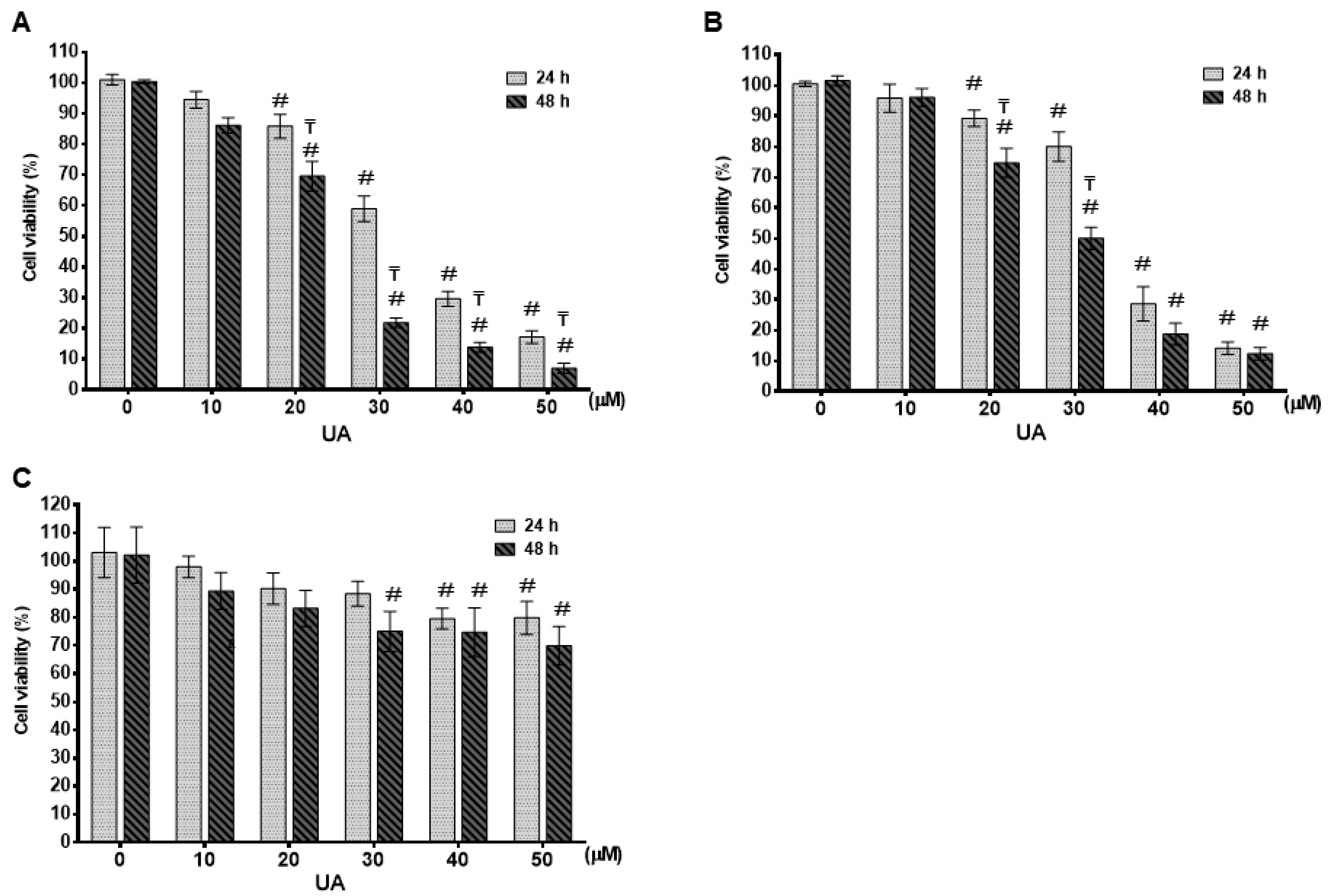
2.1. UA Inhibited Cell Proliferation in Breast Cancer Cell Lines
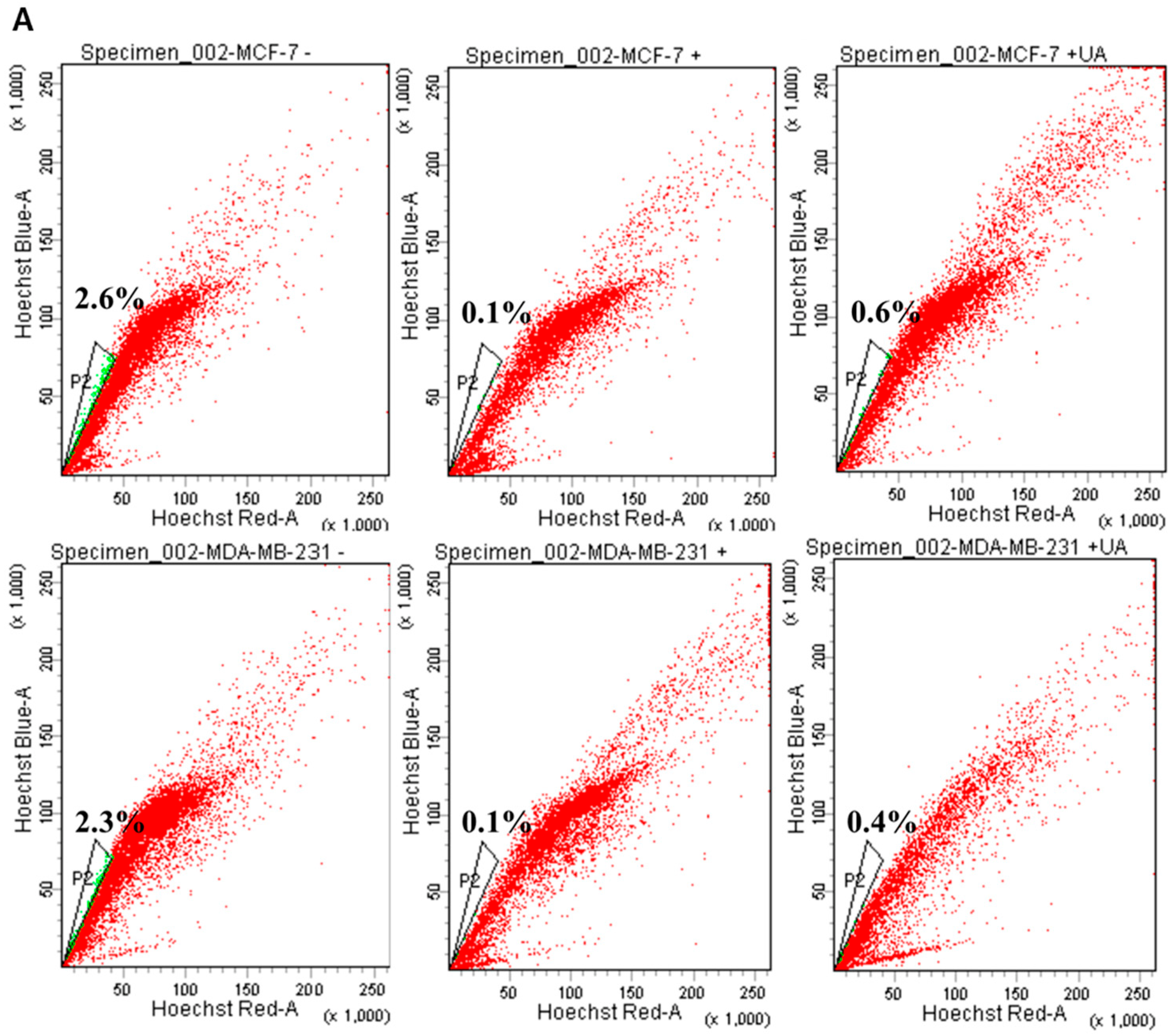
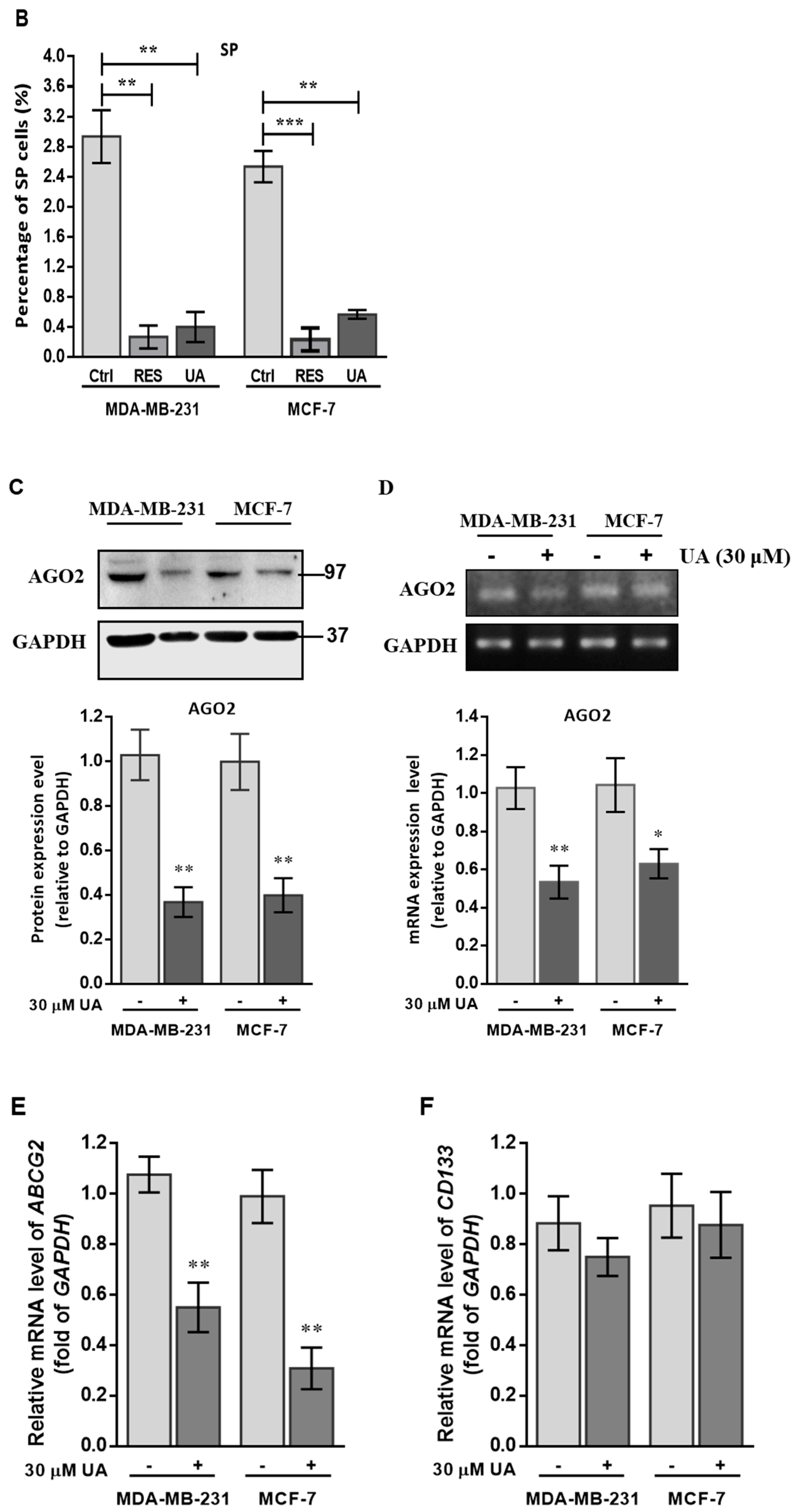
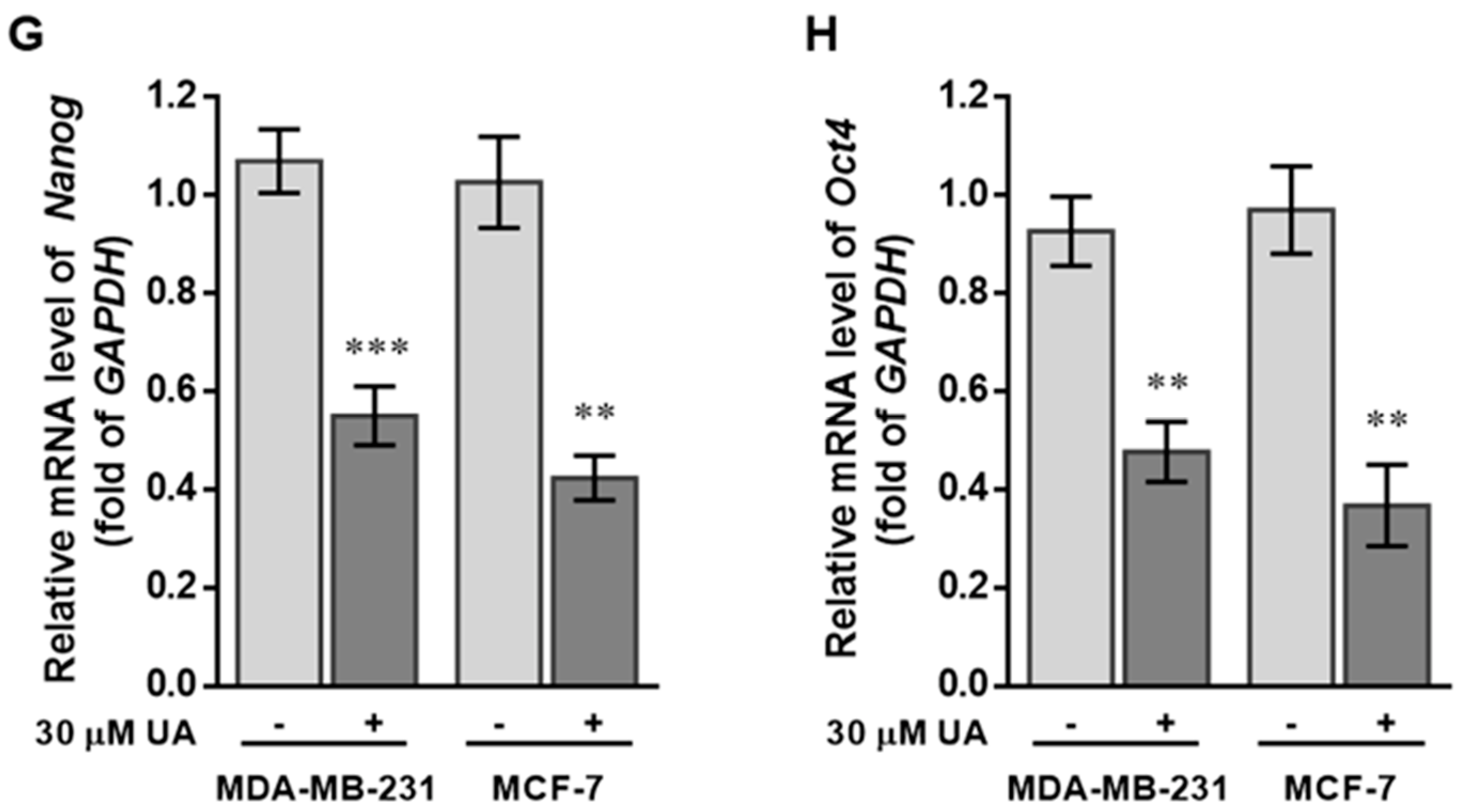
2.2. UA Decreases the Fraction of Breast Cancer Stem Cells
2.3. Sustained Exposure to UA Suppresses Migration and Invasion of Breast Cancer
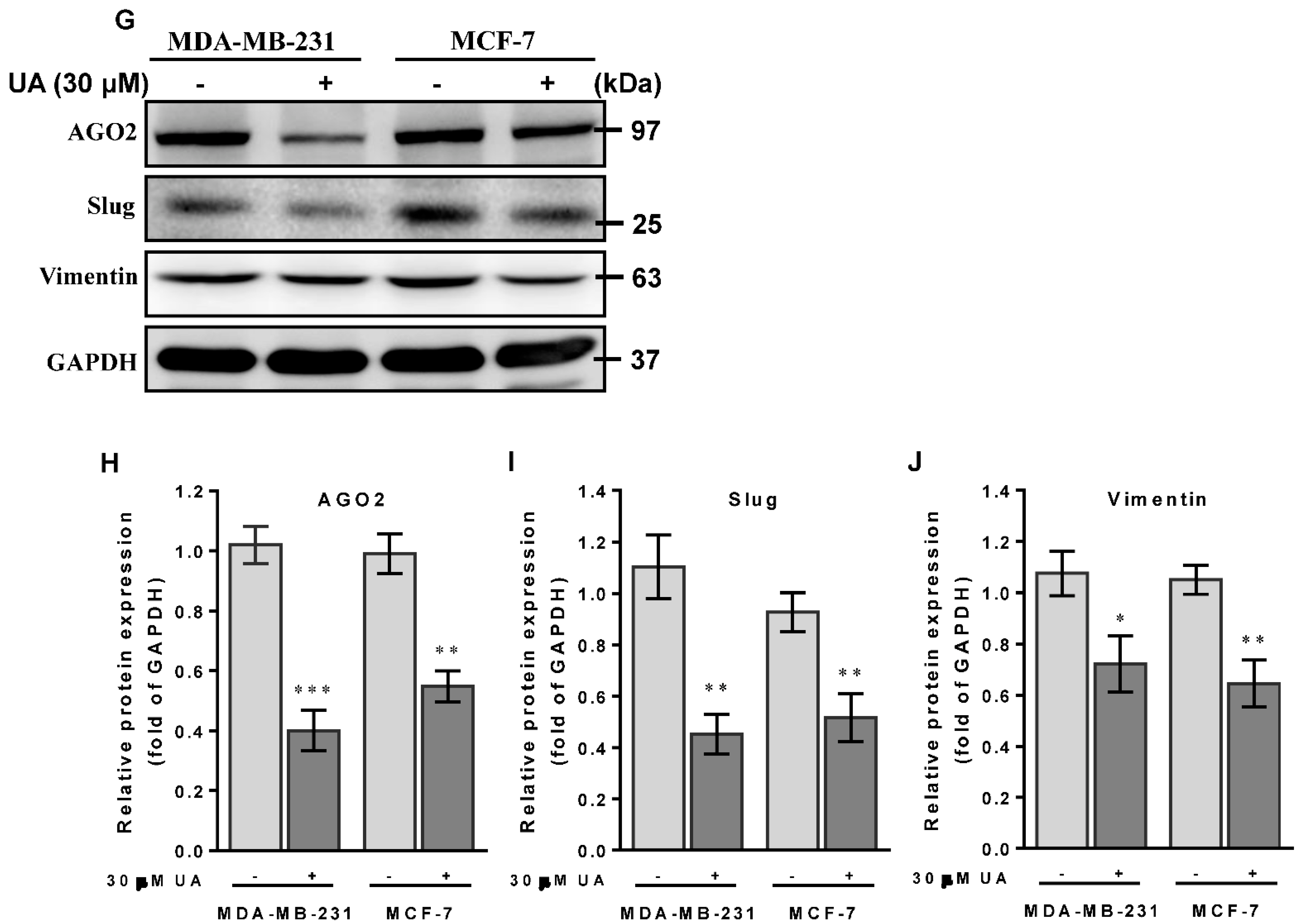
2.4. UA Impedes Tumor Cell Migration and Invasion by Inhibiting EMT-Related miRNAs and Proteins
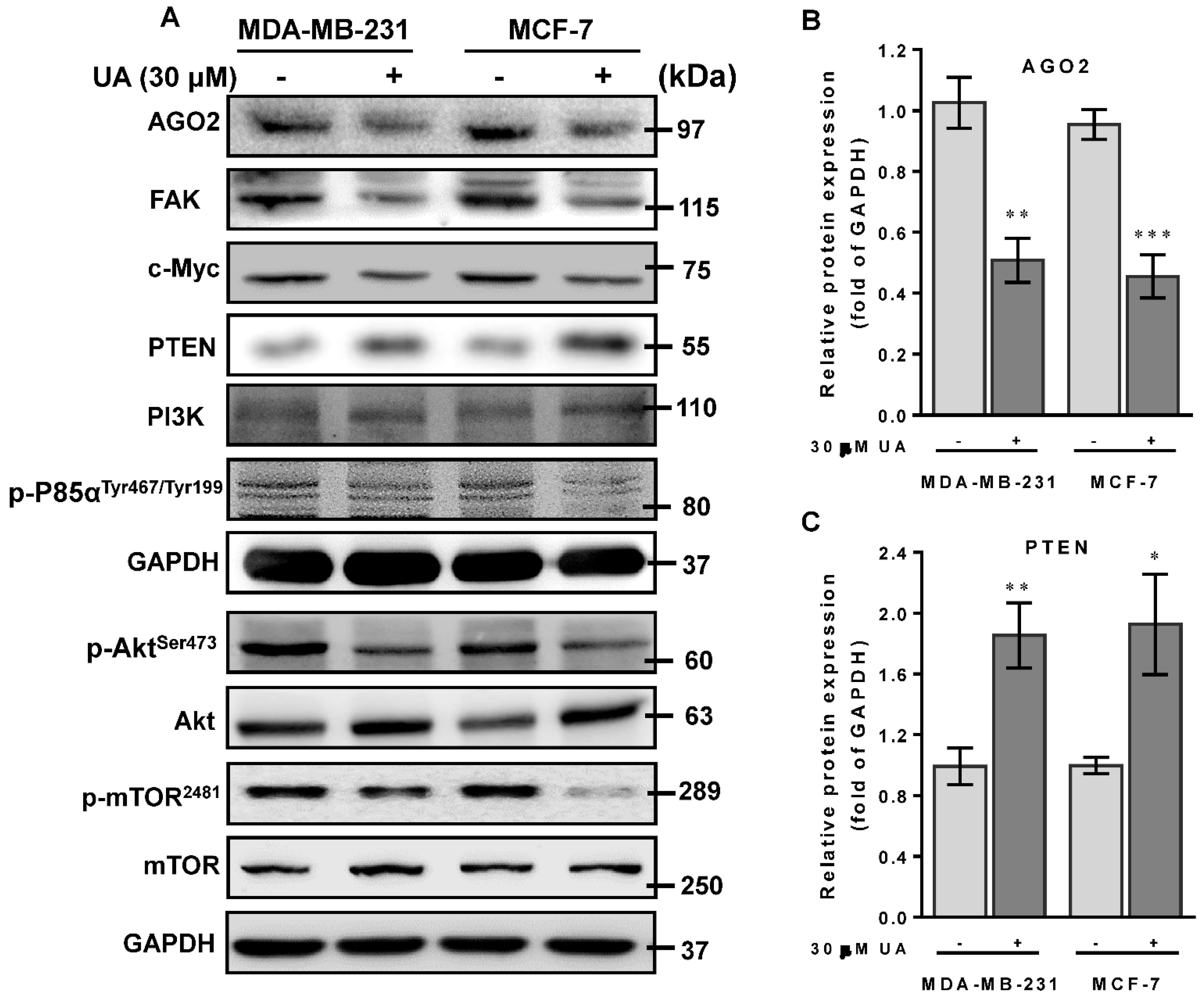
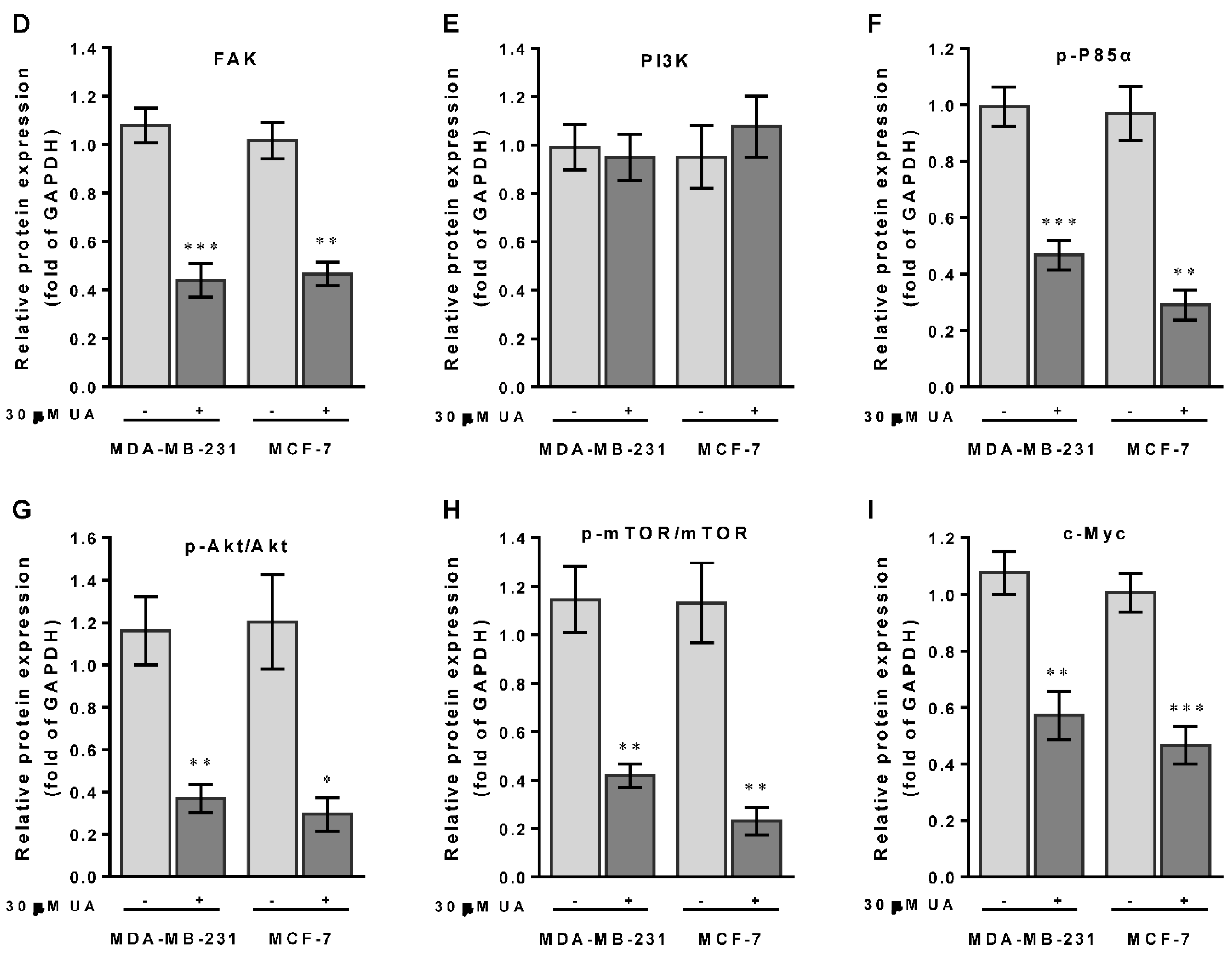
2.5. UA Enhances PTEN to Downmodulate the FAK/PI3K/Akt/mTOR Signaling Pathway
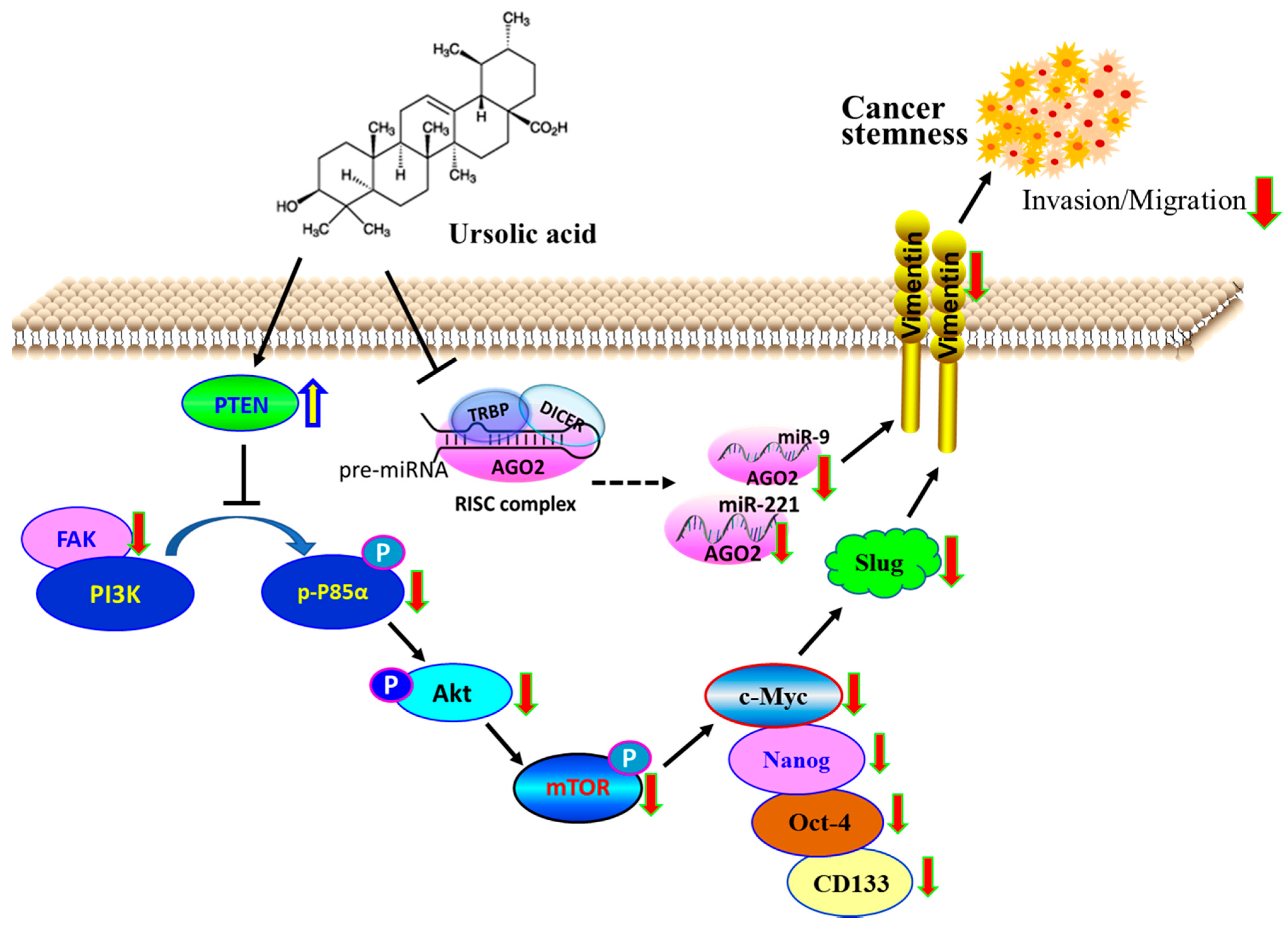
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Characterization and Isolation of CSC
4.3. Cell Viability
4.4. Matrigel Invasion and Migration Assays
4.5. RNA Extraction, cDNA Transcription and Real-Time PCR Analysis
4.6. Western Blot Analysis
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| UA | ursolic acid |
| AGO2 | Argonaute-2 |
| CSC | cancer stem cell |
| SP | side population |
| ABCG2 | ATP-binding cassette superfamily G member 2 |
| EMT | epithelial-to-mesenchymal transition |
| miRNA | microRNA |
| PTEN | phosphatase and tensin homologue on chromosome 10 |
| qRT-PCR | quantitative reverse transcription PCR |
References
- Chiang, A.C.; Massague, J. Molecular basis of metastasis. N. Engl. J. Med. 2008, 359, 2814–2823. [Google Scholar] [CrossRef] [PubMed]
- Hunter, K.W.; Crawford, N.P.; Alsarraj, J. Mechanisms of metastasis. Breast Cancer Res. 2008, 10 (Suppl. 1), S2. [Google Scholar] [CrossRef] [PubMed]
- Yokota, J. Tumor progression and metastasis. Carcinogenesis 2000, 21, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Turnquist, H.; Jackson, J.; Sgagias, M.; Yan, Y.; Gong, M.; Dean, M.; Sharp, J.G.; Cowan, K. The multidrug resistance transporter ABCG2 (breast cancer resistance protein 1) effluxes Hoechst 33342 and is overexpressed in hematopoietic stem cells. Clin. Cancer Res. 2002, 8, 22–28. [Google Scholar]
- Cheng, C.W.; Yu, J.C.; Hsieh, Y.H.; Liao, W.L.; Shieh, J.C.; Yao, C.C.; Lee, H.J.; Chen, P.M.; Wu, P.E.; Shen, C.Y. Increased Cellular Levels of MicroRNA-9 and MicroRNA-221 Correlate with Cancer Stemness and Predict Poor Outcome in Human Breast Cancer. Cell. Physiol. Biochem. 2018, 48, 2205–2218. [Google Scholar] [CrossRef]
- Cheng, C.W.; Liao, W.L.; Chen, P.M.; Yu, J.C.; Shiau, H.P.; Hsieh, Y.H.; Lee, H.J.; Cheng, Y.C.; Wu, P.E.; Shen, C.Y. MiR-139 Modulates Cancer Stem Cell Function of Human Breast Cancer through Targeting CXCR4. Cancers 2021, 13, 2582. [Google Scholar] [CrossRef]
- Theunissen, T.W.; Jaenisch, R. Molecular control of induced pluripotency. Cell. Stem Cell. 2014, 14, 720–734. [Google Scholar] [CrossRef]
- Romero-Cordoba, S.L.; Salido-Guadarrama, I.; Rodriguez-Dorantes, M.; Hidalgo-Miranda, A. miRNA biogenesis: Biological impact in the development of cancer. Cancer Biol. Ther. 2014, 15, 1444–1455. [Google Scholar] [CrossRef]
- Lee, J.H.; Schutte, D.; Wulf, G.; Fuzesi, L.; Radzun, H.J.; Schweyer, S.; Engel, W.; Nayernia, K. Stem-cell protein Piwil2 is widely expressed in tumors and inhibits apoptosis through activation of Stat3/Bcl-XL pathway. Hum. Mol. Genet. 2006, 15, 201–211. [Google Scholar] [CrossRef]
- Chen, Z.; Che, Q.; He, X.; Wang, F.; Wang, H.; Zhu, M.; Sun, J.; Wan, X. Stem cell protein Piwil1 endowed endometrial cancer cells with stem-like properties via inducing epithelial-mesenchymal transition. BMC Cancer. 2015, 15, 811. [Google Scholar] [CrossRef]
- Ciuffreda, L.; Falcone, I.; Incani, U.C.; Del Curatolo, A.; Conciatori, F.; Matteoni, S.; Vari, S.; Vaccaro, V.; Cognetti, F.; Milella, M. PTEN expression and function in adult cancer stem cells and prospects for therapeutic targeting. Adv. Biol. Regul. 2014, 56, 66–80. [Google Scholar] [CrossRef]
- Montales, M.T.; Rahal, O.M.; Kang, J.; Rogers, T.J.; Prior, R.L.; Wu, X.; Simmen, R.C. Repression of mammosphere formation of human breast cancer cells by soy isoflavone genistein and blueberry polyphenolic acids suggests diet-mediated targeting of cancer stem-like/progenitor cells. Carcinogenesis 2012, 33, 652–660. [Google Scholar] [CrossRef]
- Mulholland, D.J.; Kobayashi, N.; Ruscetti, M.; Zhi, A.; Tran, L.M.; Huang, J.; Gleave, M.; Wu, H. Pten loss and RAS/MAPK activation cooperate to promote EMT and metastasis initiated from prostate cancer stem/progenitor cells. Cancer Res. 2012, 72, 1878–1889. [Google Scholar] [CrossRef]
- Wu, P.P.; Zhang, K.; Lu, Y.J.; He, P.; Zhao, S.Q. In vitro and in vivo evaluation of the antidiabetic activity of ursolic acid derivatives. Eur. J. Med. Chem. 2014, 80, 502–508. [Google Scholar] [CrossRef]
- Kim, K.D.; Jung, H.Y.; Ryu, H.G.; Kim, B.; Jeon, J.; Yoo, H.Y.; Park, C.H.; Choi, B.H.; Hyun, C.K.; Kim, K.T.; et al. Betulinic acid inhibits high-fat diet-induced obesity and improves energy balance by activating AMPK. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Erdmann, J.; Kujacinski, M.; Wicinski, M. Beneficial Effects of Ursolic Acid and Its Derivatives-Focus on Potential Biochemical Mechanisms in Cardiovascular Conditions. Nutrients 2021, 13, 3900. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choi, J.K.; Jeong, N.H.; Yoo, J.; Ha, Y.S.; Lee, B.; Choi, H.; Park, P.H.; Shin, T.Y.; Kwon, T.K.; et al. Anti-inflammatory effects of ursolic acid-3-acetate on human synovial fibroblasts and a murine model of rheumatoid arthritis. Int. Immunopharmacol. 2017, 49, 118–125. [Google Scholar] [CrossRef]
- Dhakal, H.; Kim, M.J.; Lee, S.; Choi, Y.A.; Kim, N.; Kwon, T.K.; Khang, D.; Kim, S.H. Ursolic acid inhibits FcepsilonRI-mediated mast cell activation and allergic inflammation. Int. Immunopharmacol. 2021, 99, 107994. [Google Scholar] [CrossRef]
- Lorincz, A.; Mihaly, J.; Wacha, A.; Nemeth, C.; Besztercei, B.; Gyulavari, P.; Varga, Z.; Petak, I.; Bota, A. Combination of multifunctional ursolic acid with kinase inhibitors for anti-cancer drug carrier vesicles. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 131, 112481. [Google Scholar] [CrossRef]
- Jin, H.; Pi, J.; Yang, F.; Jiang, J.; Wang, X.; Bai, H.; Shao, M.; Huang, L.; Zhu, H.; Yang, P.; et al. Folate-Chitosan Nanoparticles Loaded with Ursolic Acid Confer Anti-Breast Cancer Activities in vitro and in vivo. Sci. Rep. 2016, 6, 0782. [Google Scholar] [CrossRef]
- Liu, T.J.; Sun, B.C.; Zhao, X.L.; Zhao, X.M.; Sun, T.; Gu, Q.; Yao, Z.; Dong, X.Y.; Zhao, N.; Liu, N. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene 2013, 32, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Yousefnia, S.; Seyed Forootan, F.; Seyed Forootan, S.; Nasr Esfahani, M.H.; Gure, A.O.; Ghaedi, K. Mechanistic Pathways of Malignancy in Breast Cancer Stem Cells. Front. Oncol. 2020, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Weissman, I.L. Translating stem and progenitor cell biology to the clinic: Barriers and opportunities. Science 2000, 287, 1442–1446. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, G.J.; Visvader, J.E. Insights into the cell of origin in breast cancer and breast cancer stem cells. Asia-Pac. J. Clin. Oncol. 2010, 6, 89–97.21. [Google Scholar] [CrossRef] [PubMed]
- Alberti, L.; Losi, L.; Leyvraz, S.; Benhattar, J. Different Effects of BORIS/CTCFL on Stemness Gene Expression, Sphere Formation and Cell Survival in Epithelial Cancer Stem Cells. PLoS ONE 2015, 10, e0132977. [Google Scholar] [CrossRef]
- Hollestelle, A.; Peeters, J.K.; Smid, M.; Timmermans, M.; Verhoog, L.C.; Westenend, P.J.; Heine, A.A.; Chan, A.; Sieuwerts, A.M.; Wiemer, E.A.C.; et al. Loss of E-cadherin is not a necessity for epithelial to mesenchymal transition in human breast cancer. Breast Cancer Res. Treat. 2013, 138, 47–57. [Google Scholar] [CrossRef]
- Vuoriluoto, K.; Haugen, H.; Kiviluoto, S.; Mpindi, J.P.; Nevo, J.; Gjerdrum, C.; Tiron, C.; Lorens, J.B.; Ivaska, J. Vimentin regulates EMT induction by Slug and oncogenic H-Ras and migration by governing Axl expression in breast cancer. Oncogene 2011, 30, 1436–1448. [Google Scholar] [CrossRef]
- Bleau, A.M.; Hambardzumyan, D.; Ozawa, T.; Fomchenko, E.I.; Huse, J.T.; Brennan, C.W.; Holland, E.C. PTEN/PI3K/Akt pathway regulates the side population phenotype and ABCG2 activity in glioma tumor stem-like cells. Cell. Stem Cell. 2009, 4, 226–235. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, B.; Chen, X.; Li, W.; Dong, P. AGO2 involves the malignant phenotypes and FAK/PI3K/AKT signaling pathway in hypopharyngeal-derived FaDu cells. Oncotarget 2017, 8, 54735–54746. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/beta-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Nowak, I.; Sarshad, A.A. Argonaute Proteins Take Center Stage in Cancers. Cancers 2021, 13, 788. [Google Scholar] [CrossRef]
- Hanina, S.A.; Mifsud, W.; Down, T.A.; Hayashi, K.; O’Carroll, D.; Lao, K.; Miska, E.A.; Surani, M.A. Genome-wide identification of targets and function of individual MicroRNAs in mouse embryonic stem cells. PLoS Genet. 2010, 6, e1001163. [Google Scholar] [CrossRef]
- Takahashi, R.U.; Miyazaki, H.; Ochiya, T. The role of microRNAs in the regulation of cancer stem cells. Front. Genet. 2014, 4, 295. [Google Scholar] [CrossRef]
- Khan, A.Q.; Ahmed, E.I.; Elareer, N.R.; Junejo, K.; Steinhoff, M.; Uddin, S. Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells 2019, 8, 840. [Google Scholar] [CrossRef]
- Wu, B.; Wang, X.; Chi, Z.F.; Hu, R.; Zhang, R.; Yang, W.; Liu, Z.G. Ursolic acid-induced apoptosis in K562 cells involving upregulation of PTEN gene expression and inactivation of the PI3K/Akt pathway. Arch. Pharm. Res. 2012, 35, 543–548. [Google Scholar] [CrossRef]
- Yeh, C.T.; Wu, C.H.; Yen, G.C. Ursolic acid, a naturally occurring triterpenoid, suppresses migration and invasion of human breast cancer cells by modulating c-Jun N-terminal kinase, Akt and mammalian target of rapamycin signaling. Mol. Nutr. Food. Res. 2010, 54, 1285–1295. [Google Scholar] [CrossRef]
- Meng, Y.; Lin, Z.M.; Ge, N.; Zhang, D.L.; Huang, J.; Kong, F. Ursolic Acid Induces Apoptosis of Prostate Cancer Cells via the PI3K/Akt/mTOR Pathway. Am. J. Chin. Med. 2015, 43, 1471–1486. [Google Scholar] [CrossRef]
- Obayashi, M.; Yoshida, M.; Tsunematsu, T.; Ogawa, I.; Sasahira, T.; Kuniyasu, H.; Imoto, I.; Abiko, Y.; Xu, D.; Fukunaga, S.; et al. microRNA-203 suppresses invasion and epithelial-mesenchymal transition induction via targeting NUAK1 in head and neck cancer. Oncotarget 2016, 7, 8223–8239. [Google Scholar] [CrossRef]
- Sternburg, E.L.; Estep, J.A.; Nguyen, D.K.; Li, Y.; Karginov, F.V. Antagonistic and cooperative AGO2-PUM interactions in regulating mRNAs. Sci. Rep. 2018, 8, 15316. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, H.; Lou, J.R.; Zheng, J.; Zhu, H.; Popescu, N.I.; Lupu, F.; Lind, S.E.; Ding, W.Q. MicroRNA-19 (miR-19) regulates tissue factor expression in breast cancer cells. J. Biol. Chem. 2011, 286, 1429–1435. [Google Scholar] [CrossRef]
- Katz, S.; Cussigh, D.; Urban, N.; Blomfield, I.; Guillemot, F.; Bally-Cuif, L.; Coolen, M. A Nuclear Role for miR-9 and Argonaute Proteins in Balancing Quiescent and Activated Neural Stem Cell States. Cell Rep. 2016, 17, 1383–1398. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.L.; Huang, Y.; Li, L.F.; Zhu, H.L.; Gao, H.X.; Liu, H.; Lv, S.Q.; Xu, Z.H.; Zheng, L.N.; Liu, T.; et al. Argonaute 2 promotes angiogenesis via the PTEN/VEGF signaling pathway in human hepatocellular carcinoma. Acta Pharmacol. Sin. 2015, 36, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Grigorieva, I.; Steele, R.; Hoover, R.G.; Ray, R.B. PTEN transcriptionally modulates c-myc gene expression in human breast carcinoma cells and is involved in cell growth regulation. Gene 1999, 235, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.L.; Shui, Y.M.; Jiang, W.; Huang, E.Y.; Shou, Q.Y.; Ji, X.; He, B.C.; Lv, G.Y.; He, T.C. Hypoxia pathway and hypoxia-mediated extensive extramedullary hematopoiesis are involved in ursolic acid’s anti-metastatic effect in 4T1 tumor bearing mice. Oncotarget 2016, 7, 71802–71816. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D. Ursolic Acid Decreases the Proliferation of MCF-7 Cell-Derived Breast Cancer Stem-Like Cells by Modulating the ERK and PI3K/AKT Signaling Pathways. Prev. Nutr. Food Sci. 2021, 26, 434–444. [Google Scholar] [CrossRef]
- Zhang, R.X.; Li, Y.; Tian, D.D.; Liu, Y.; Nian, W.; Zou, X.; Chen, Q.Z.; Zhou, L.Y.; Deng, Z.L.; He, B.C. Ursolic acid inhibits proliferation and induces apoptosis by inactivating Wnt/beta-catenin signaling in human osteosarcoma cells. Int. J. Oncol. 2016, 49, 1973–1982. [Google Scholar] [CrossRef]
- Kang, D.Y.; Sp, N.; Lee, J.M.; Jang, K.J. Antitumor Effects of Ursolic Acid through Mediating the Inhibition of STAT3/PD-L1 Signaling in Non-Small Cell Lung Cancer Cells. Biomedicines 2021, 9, 297. [Google Scholar] [CrossRef]
- Liao, W.L.; Lin, J.Y.; Shieh, J.C.; Yeh, H.F.; Hsieh, Y.H.; Cheng, Y.C.; Lee, H.J.; Shen, C.Y.; Cheng, C.W. Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 21, 172. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences Used for the mRNA Transcript | PCR Product | |
|---|---|---|---|
| AGO2 | Forward: | 5′-TCCACCTAGACCCGACTTT-3′ | 155 bp |
| Reverse: | 5′-GTTCCACGATTTCCCTGTT-3′ | ||
| ABCG2 | Forward: | 5′-TCGGCTTGCAACAACTATG-3′ | 128 bp |
| Reverse: | 5′-TCCAGACACACCACGGATAA-3′ | ||
| Nanog | Forward: | 5′-TAGCAATGGTGTGACGCAGAAG-3′ | 116 bp |
| Reverse: | 5′-TCTGGTTGCTCCACATTGGAAGG-3′ | ||
| CD133 | Forward: | 5′-ACAATTCACCAGCAACGAGTCC-3′ | 63 bp |
| Reverse: | 5′-GACGCTTTGGTATAGAGTGCTCAG-3′ | ||
| Oct4 | Forward: | 5′-GAGGCAACCTGGAGAATTTGTTCC-3′ | 64 bp |
| Reverse: | 5′-ATGTGGCTGATCTGCTGCAGTG-3′ | ||
| GAPDH | Forward: | 5′-GAAGGTGAAGGTCGGAGTC-3′ | 226 bp |
| Reverse: | 5′-GAAGATGGTGATGGGATTTC-3′ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, W.-L.; Liu, Y.-F.; Ying, T.-H.; Shieh, J.-C.; Hung, Y.-T.; Lee, H.-J.; Shen, C.-Y.; Cheng, C.-W. Inhibitory Effects of Ursolic Acid on the Stemness and Progression of Human Breast Cancer Cells by Modulating Argonaute-2. Int. J. Mol. Sci. 2023, 24, 366. https://doi.org/10.3390/ijms24010366
Liao W-L, Liu Y-F, Ying T-H, Shieh J-C, Hung Y-T, Lee H-J, Shen C-Y, Cheng C-W. Inhibitory Effects of Ursolic Acid on the Stemness and Progression of Human Breast Cancer Cells by Modulating Argonaute-2. International Journal of Molecular Sciences. 2023; 24(1):366. https://doi.org/10.3390/ijms24010366
Chicago/Turabian StyleLiao, Wen-Ling, Yu-Fan Liu, Tsung-Ho Ying, Jia-Ching Shieh, Yueh-Tzu Hung, Huei-Jane Lee, Chen-Yang Shen, and Chun-Wen Cheng. 2023. "Inhibitory Effects of Ursolic Acid on the Stemness and Progression of Human Breast Cancer Cells by Modulating Argonaute-2" International Journal of Molecular Sciences 24, no. 1: 366. https://doi.org/10.3390/ijms24010366
APA StyleLiao, W.-L., Liu, Y.-F., Ying, T.-H., Shieh, J.-C., Hung, Y.-T., Lee, H.-J., Shen, C.-Y., & Cheng, C.-W. (2023). Inhibitory Effects of Ursolic Acid on the Stemness and Progression of Human Breast Cancer Cells by Modulating Argonaute-2. International Journal of Molecular Sciences, 24(1), 366. https://doi.org/10.3390/ijms24010366