


Prevention of Noise-Induced Hearing Loss In Vivo: Continuous Application of Insulin-like Growth Factor 1 and Its Effect on Inner Ear Synapses, Auditory Function and Perilymph Proteins

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
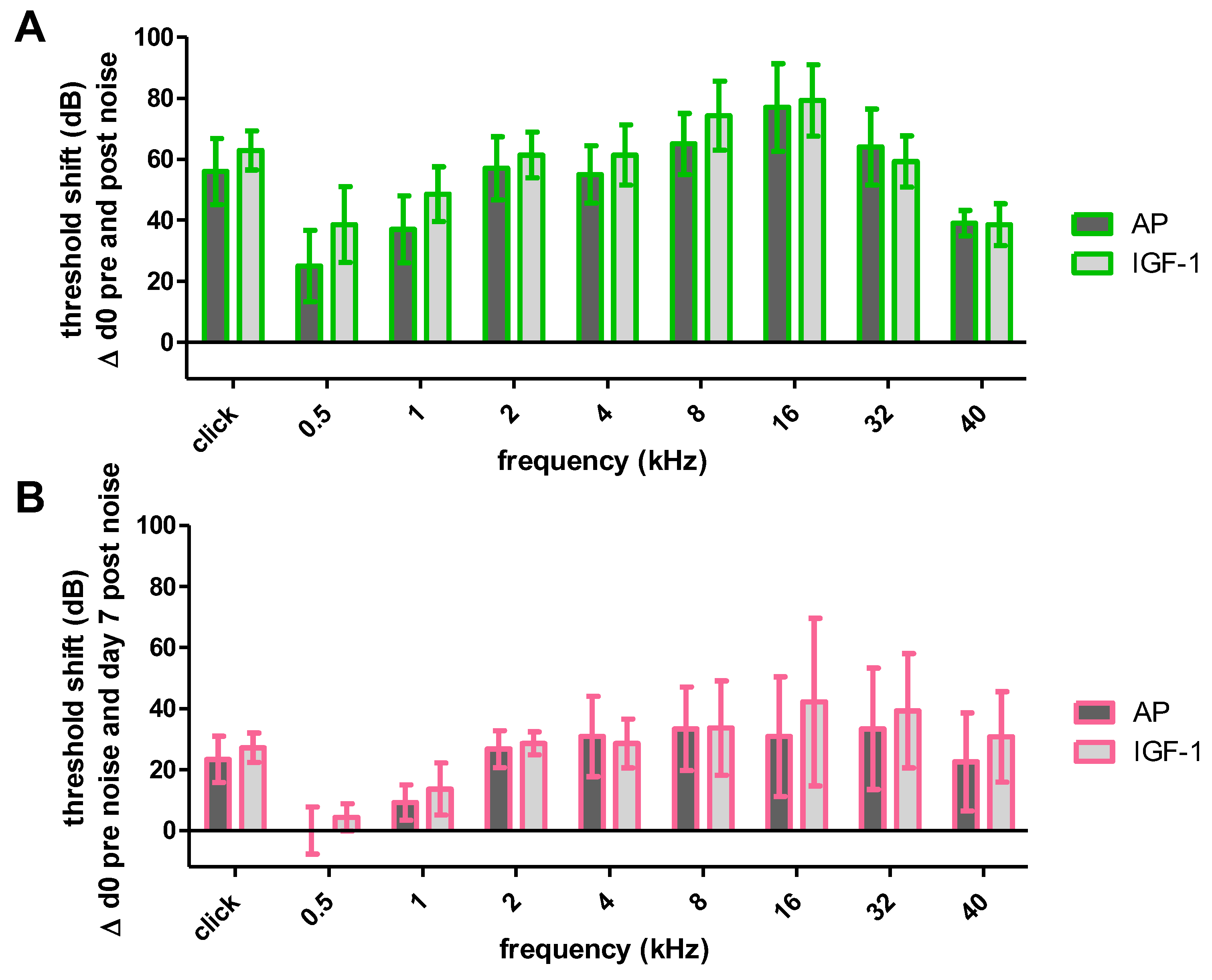
2.1. Acoustically Evoked Auditory Brainstem Response (AABR)-Measurements
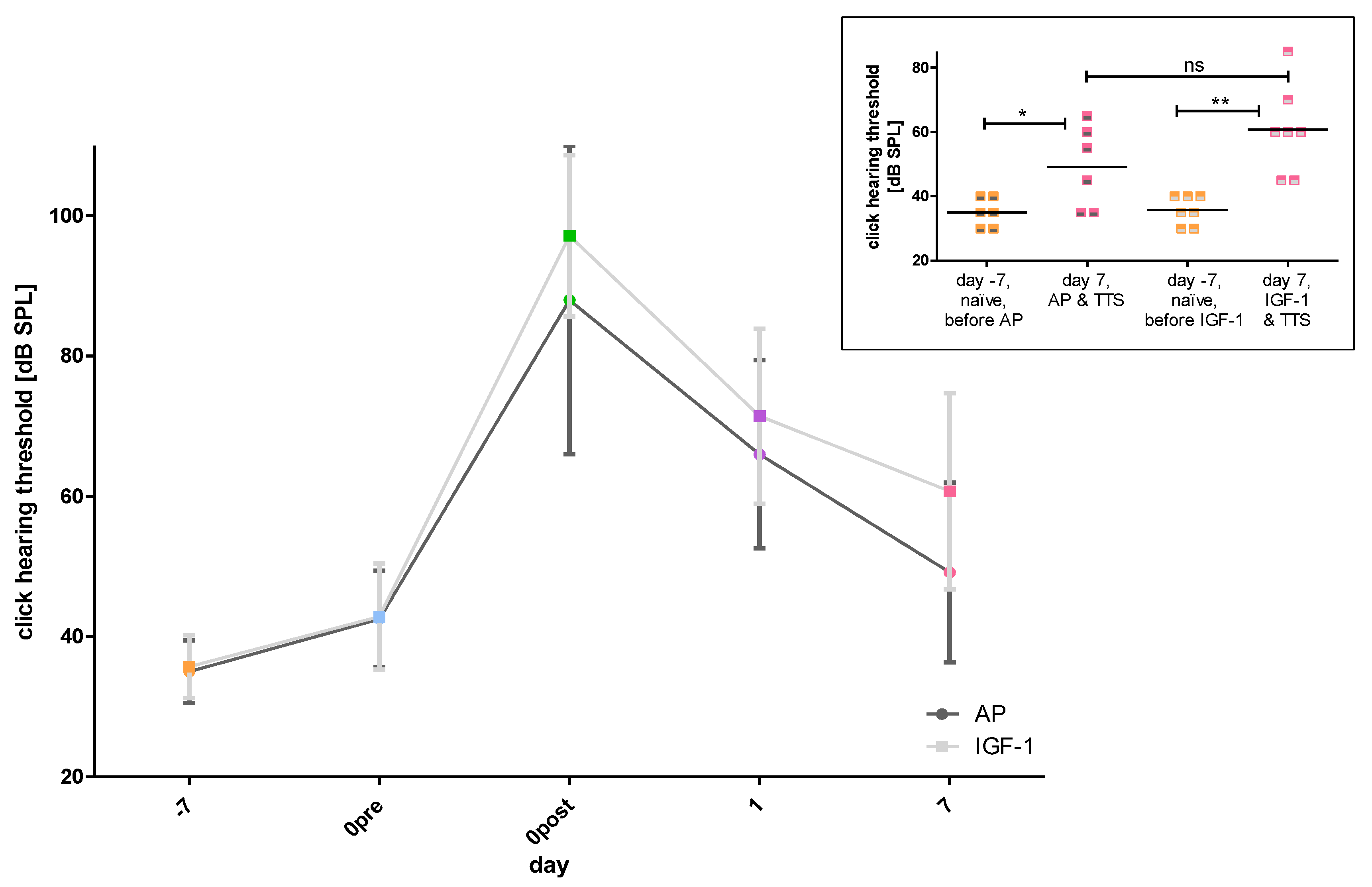
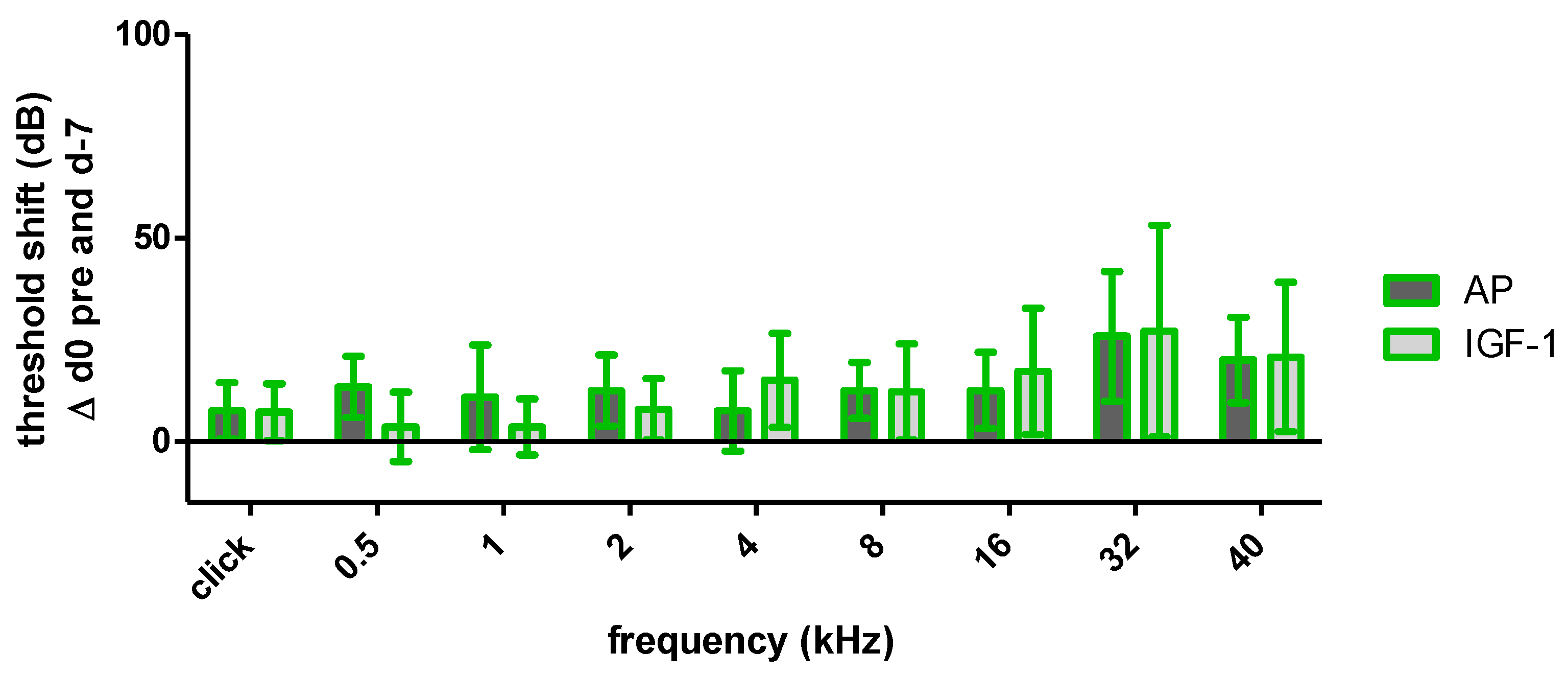
2.1.1. Impact of the Implantation on Hearing Threshold
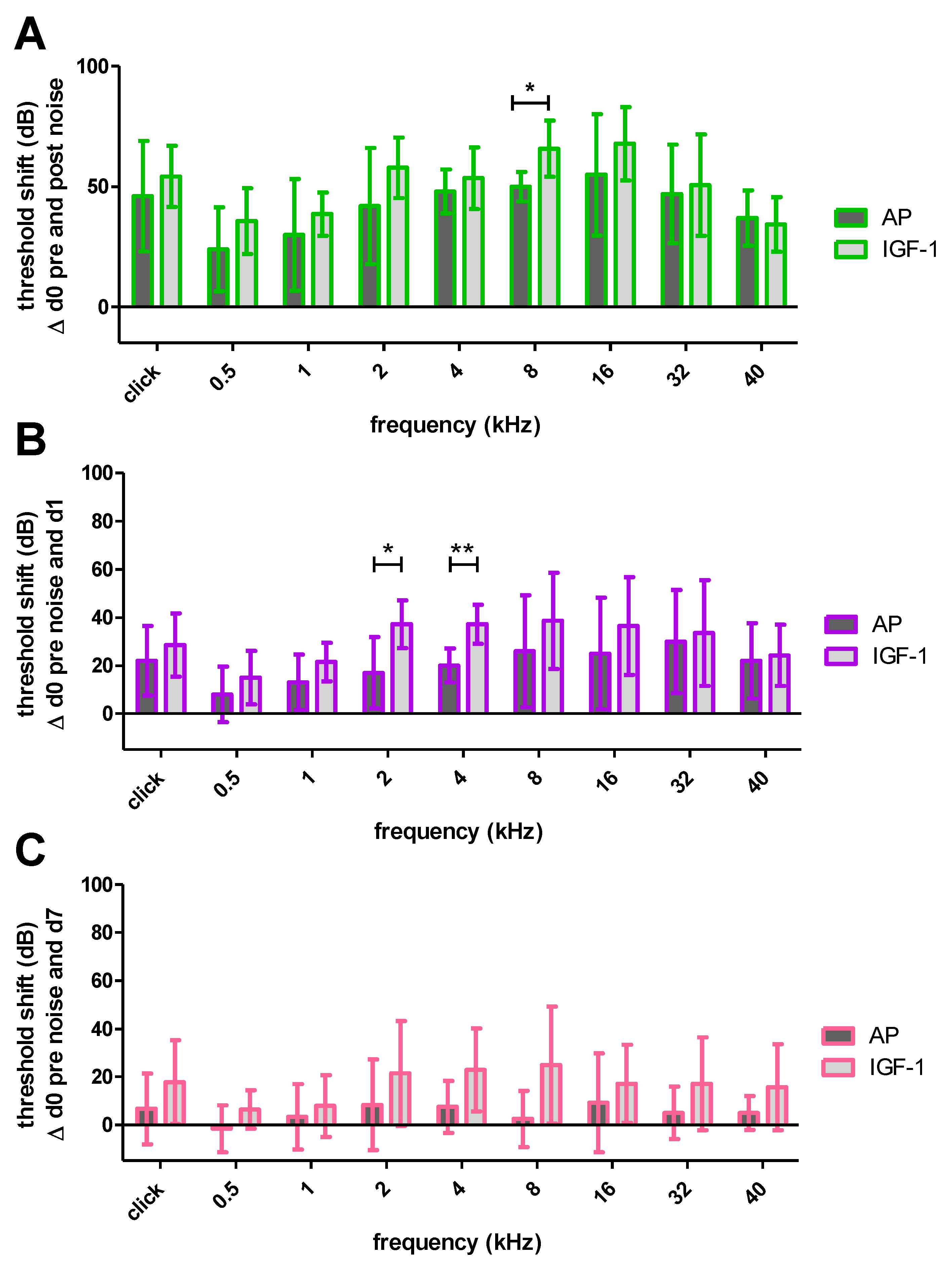
2.1.2. Impact of Noise Trauma and Treatment on Hearing Threshold
2.1.3. Impact of Noise Trauma on Contralateral Ears
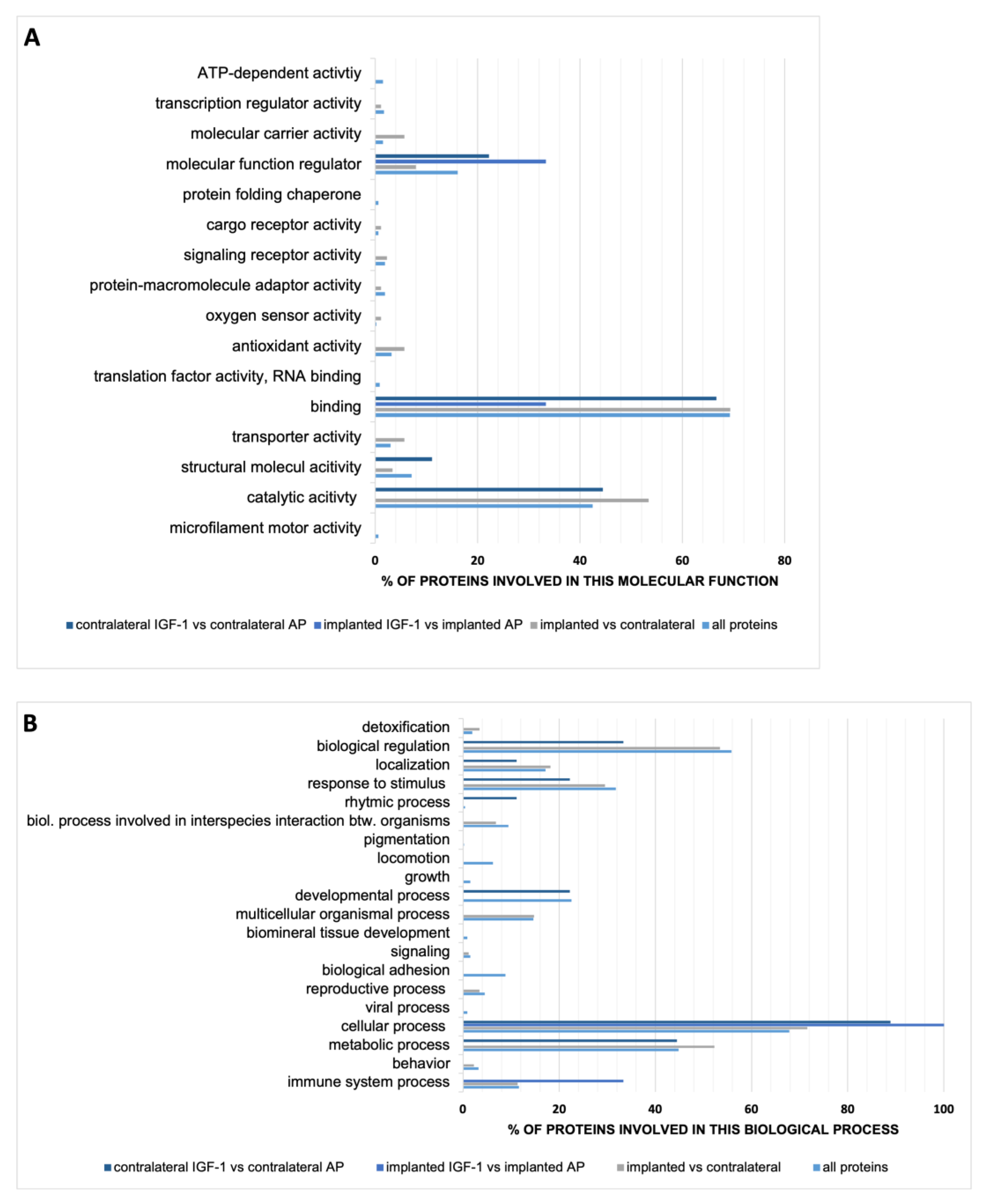
2.2. Perilymph
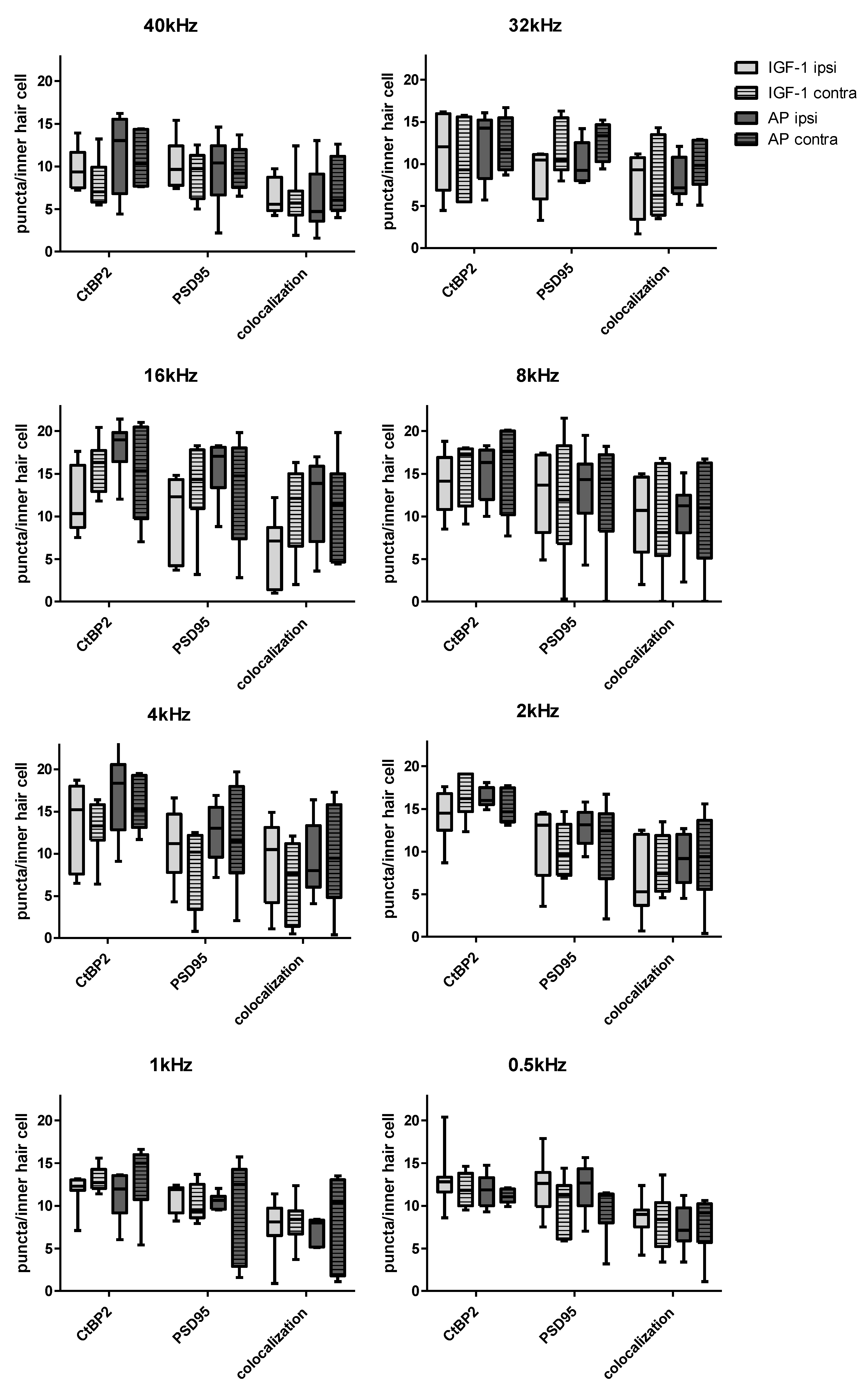
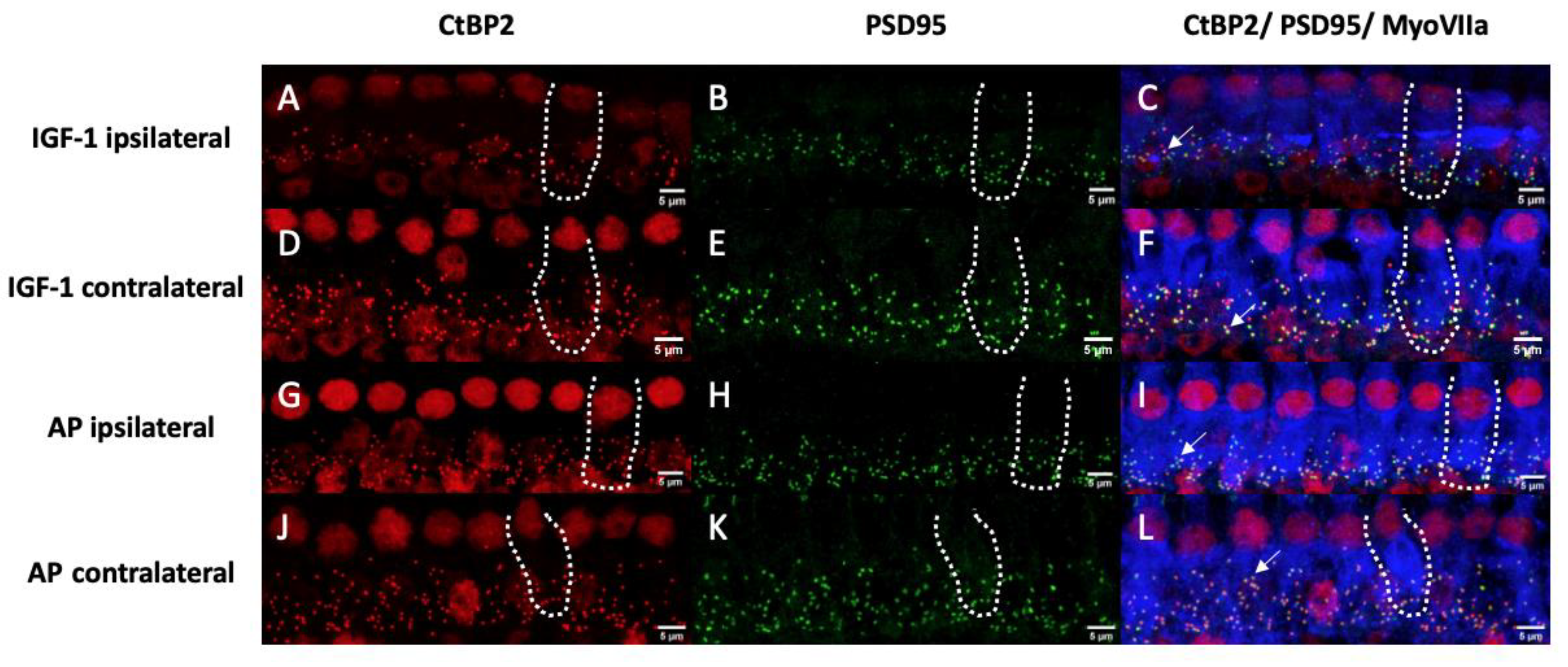
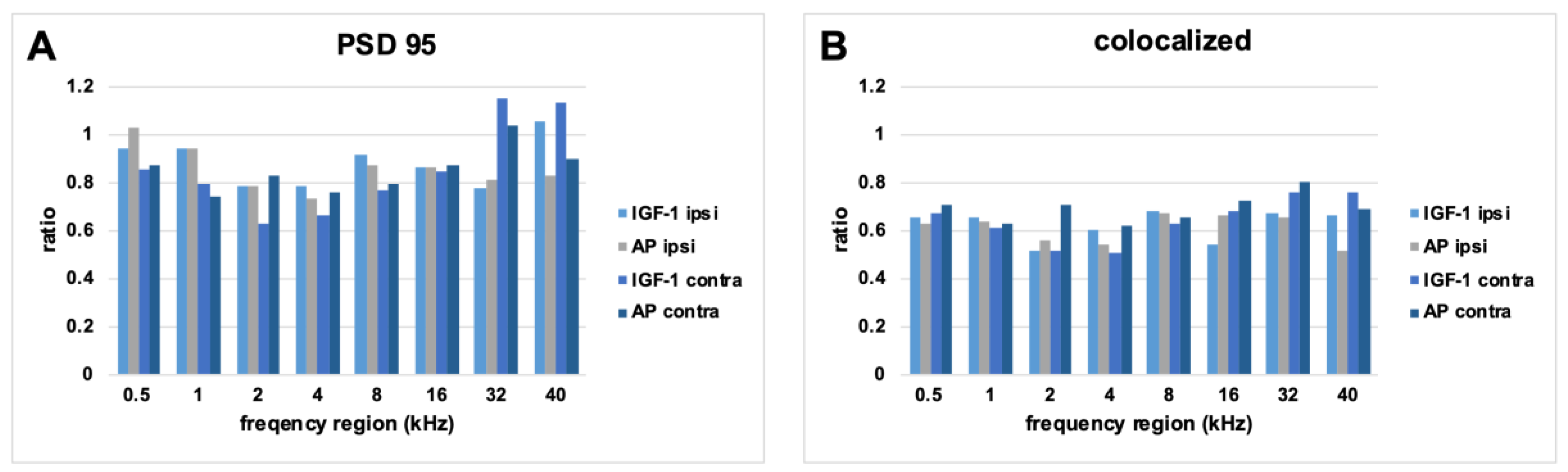
2.3. Histological Examination of the Inner Hair Cell’s Ribbon Synapses
3. Discussion
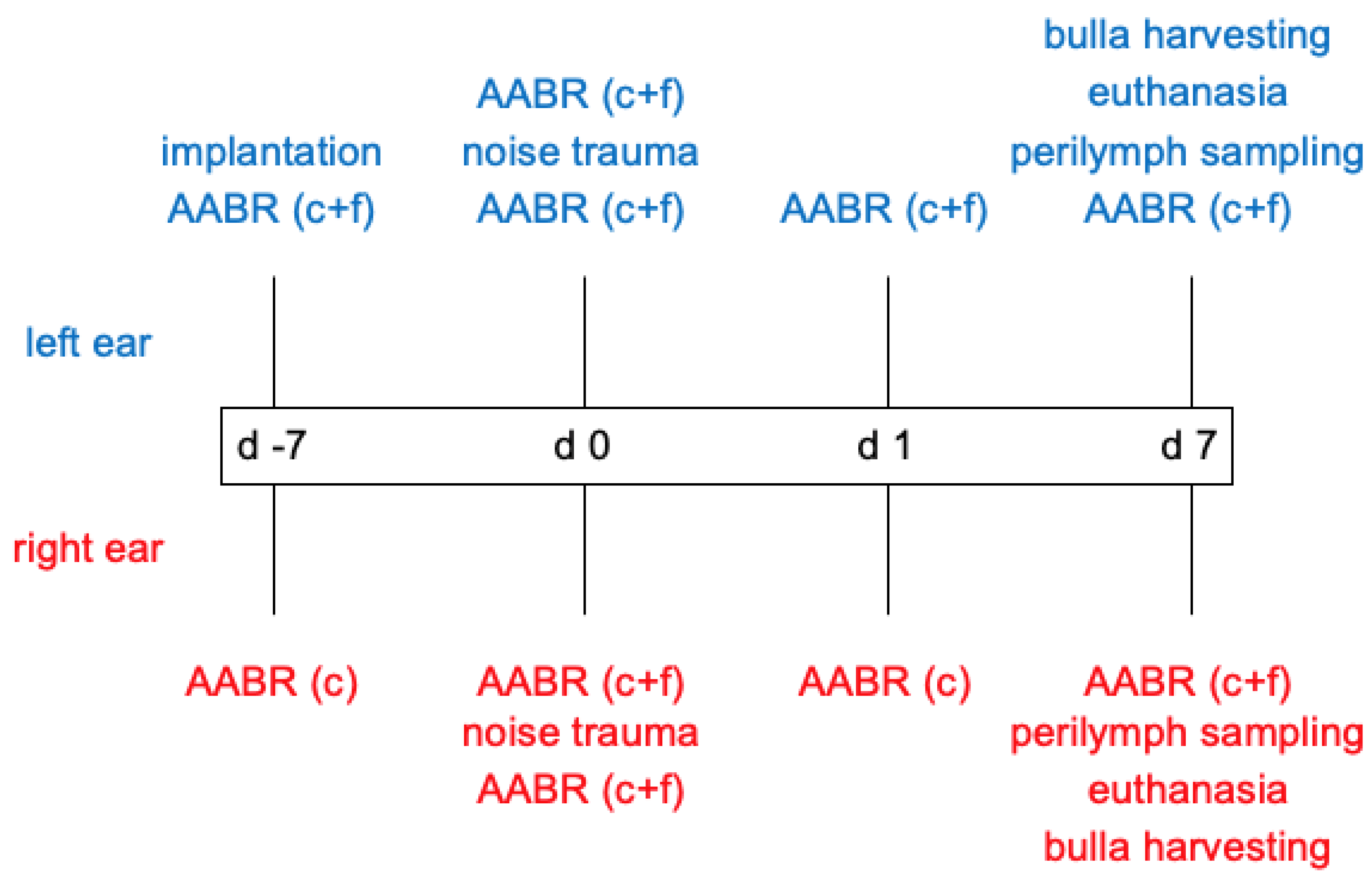
4. Materials and Methods
4.1. Pump Preparation
4.2. Animals
4.3. Acoustically Evoked Auditory Brainstem Response (AABR)—Measurement
4.4. Hook Delivery Device (HDD) Implantation
4.5. Noise Trauma
4.6. Perilymph Sampling and Analysis
4.7. Tissue Preparation
4.8. Synapse Quantification
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.I.; Nelson, R.Y.; Concha-Barrientos, M.; Fingerhut, M. The global burden of occupational noise-induced hearing loss. Am. J. Ind. Med. 2005, 48, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Al-Omari, A.; Al-Khalaf, H.; Hussien, N.M. Association of flying time with hearing loss in military pilots. Saudi J. Med. Med. Sci. 2018, 6, 155. [Google Scholar] [CrossRef] [PubMed]
- Neitzel, R.L.; Long, R.N.; Sun, K.; Sayler, S.; von Thaden, T.L. Injury Risk and Noise Exposure in Firefighter Training Operations. Ann. Occup. Hyg. 2016, 60, 405–420. [Google Scholar] [CrossRef][Green Version]
- Win, K.N.; Balalla, N.B.P.; Lwin, M.Z.; Lai, A. Noise-Induced Hearing Loss in the Police Force. Saf. Health Work 2015, 6, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; John, A.B.; Kimball, S.; Fruits, T. Prevalence of tinnitus and noise-induced hearing loss in dentists. Noise Health 2016, 18, 347–354. [Google Scholar] [CrossRef]
- Fredriksson, S.; Kim, J.L.; Torén, K.; Magnusson, L.; Kähäri, K.; Söderberg, M.; Persson Waye, K. Working in preschool increases the risk of hearing-related symptoms: A cohort study among Swedish women. Int. Arch. Occup. Environ. Health 2019, 92, 1179–1190. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Freitas, M.A.; Neves, M.P.; Silva, M.V. Evaluation of the noise exposure of symphonic orchestra musicians. Noise Health 2014, 16, 40–46. [Google Scholar] [CrossRef]
- Schink, T.; Kreutz, G.; Busch, V.; Pigeot, I.; Ahrens, W. Incidence and relative risk of hearing disorders in professional musicians. Occup. Environ. Med. 2014, 71, 472–476. [Google Scholar] [CrossRef]
- Murphy, W.J.; Eichwald, J.; Meinke, D.K.; Chadha, S.; Iskander, J. CDC Grand Rounds: Promoting Hearing Health Across the Lifespan. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 243–246. [Google Scholar] [CrossRef]
- Basner, M.; Babisch, W.; Davis, A.; Brink, M.; Clark, C.; Janssen, S.; Stansfeld, S. Auditory and non-auditory effects of noise on health. Lancet 2014, 383, 1325–1332. [Google Scholar] [CrossRef]
- Spoendlin, H. Degeneration behaviour of the cochlear nerve. Arch. Klin. Exp. Ohren. Nasen. Kehlkopfheilkd. 1971, 200, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Dodson, H.C.; Mohuiddin, A. Response of spiral ganglion neurones to cochlear hair cell destruction in the guinea pig. J. Neurocytol. 2000, 29, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Ylikoski, J. Correlative studies on the cochlear pathology and hearing loss in guinea-pigs after intoxication with ototoxic antibiotics. Acta Otolaryngol. 1974, 326, 1–62. [Google Scholar]
- Kujawa, S.G.; Liberman, M.C. Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hear. Res. 2015, 330, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Liberman, M.C.; Kujawa, S.G. Cochlear synaptopathy in acquired sensorineural hearing loss: Manifestations and mechanisms. Hear. Res. 2017, 349, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shi, L.; Liu, L.; Yin, S.; Aiken, S.; Wang, J. Noise-induced Cochlear Synaptopathy and Signal Processing Disorders. Neuroscience 2019, 407, 41–52. [Google Scholar] [CrossRef]
- Wei, M.; Wang, W.; Liu, Y.; Mao, X.; Chen, T.S.; Lin, P. Protection of Cochlear Ribbon Synapses and Prevention of Hidden Hearing Loss. Neural Plast. 2020, 2020, 8815990. [Google Scholar] [CrossRef]
- Monaghan, J.J.M.; Garcia-Lazaro, J.A.; McAlpine, D.; Schaette, R. Hidden Hearing Loss Impacts the Neural Representation of Speech in Background Noise. Curr. Biol. 2020, 30, 4710–4721.e4. [Google Scholar] [CrossRef]
- Gupta, A.; Koochakzadeh, S.; Nguyen, S.A.; Brennan, E.A.; Meyer, T.A.; Lambert, P.R. Pharmacological Prevention of Noise-induced Hearing Loss: A Systematic Review. Otol. Neurotol. 2021, 42, 2–9. [Google Scholar] [CrossRef]
- Le Prell, C.G.; Hammill, T.L.; Murphy, W.J. Noise-induced hearing loss: Translating risk from animal models to real-world environments. J. Acoust. Soc. Am. 2019, 146, 3646–3651. [Google Scholar] [CrossRef]
- Sha, S.H.; Schacht, J. Emerging therapeutic interventions against noise-induced hearing loss. Expert Opin. Investig. Drugs 2017, 26, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Ren, T.; Nuttall, A.L. The electrochemical and fluorescence detection of nitric oxide in the cochlea and its increase following loud sound. Hear. Res. 2002, 164, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Ohlemiller, K.K.; Wright, J.S.; Dugan, L.L. Early elevation of cochlear oxygen species following reactive noise exposure. Audiol. Neuro-Otol. 1999, 4, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; He, Q.; Lai, H.; Wang, J. Oxidative stress in spiral ganglion cells of pigmented and albino guinea pigs exposed to impulse noise. Acta Otolaryngol. 2011, 131, 914–920. [Google Scholar] [CrossRef]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear Hear. 2006, 27, 1–19. [Google Scholar] [CrossRef]
- Pak, J.H.; Kim, Y.; Yi, J.; Chung, J.W. Antioxidant therapy against oxidative damage of the inner ear: Protection and preconditioning. Antioxidants 2020, 9, 1076. [Google Scholar] [CrossRef]
- Scheper, V.; Leifholz, M.; von der Leyen, H.; Keller, M.; Denkena, U.; Koch, A.; Karch, A.; Miller, J.; Lenarz, T. ACEMg-mediated hearing preservation in cochlear implant patients receiving different electrode lengths (PROHEARING): Study protocol for a randomized controlled trial. Trials 2016, 17, 394. [Google Scholar] [CrossRef]
- Miller, J.M.; Brown, J.N.; Schacht, J. 8-Iso-prostaglandin F2α, a product of noise exposure, reduces inner ear blood flow. Audiol. Neuro-Otol. 2003, 8, 207–221. [Google Scholar] [CrossRef]
- Miller, J.M.; Dengerink, H. Control of inner ear blood flow. Am. J. Otolaryngol. Head Neck Med. Surg. 1988, 9, 302–316. [Google Scholar] [CrossRef]
- Shin, S.A.; Lyu, A.R.; Jeong, S.H.; Kim, T.H.; Park, M.J.; Park, Y.H. Acoustic trauma modulates cochlear blood flow and vasoactive factors in a rodent model of noise-induced hearing loss. Int. J. Mol. Sci. 2019, 20, 5316. [Google Scholar] [CrossRef]
- Bailey-Downs, L.C.; Mitschelen, M.; Sosnowska, D.; Toth, P.; Pinto, J.T.; Ballabh, P.; Valcarcel-Ares, M.N.; Farley, J.; Koller, A.; Henthorn, J.C.; et al. Liver-specific knockdown of IGF-1 decreases vascular oxidative stress resistance by impairing the Nrf2-dependent antioxidant response: A novel model of vascular aging. Journals Gerontol. Ser. A Biol. Sci. Med. Sci. 2012, 67, 313–329. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, M.; Castilla-Cortázar, I.; Díaz-Sanchez, M.; Navarro, I.; Puche, J.E.; Castilla, A.; Díaz Casares, A.; Clavijo, E.; González-Barón, S. Antioxidant effects of insulin-like growth factor-I (IGF-I) in rats with advanced liver cirrhosis. BMC Gastroenterol. 2005, 5, 7. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li, X.D.; Hao, Z.H.; Xu, D. Insulin-like growth factor-1 improves diabetic cardiomyopathy through antioxidative and anti-inflammatory processes along with modulation of Akt/GSK-3β signaling in rats. Korean J. Physiol. Pharmacol. 2016, 20, 613. [Google Scholar] [CrossRef] [PubMed]
- Yamahara, K.; Nishimura, K.; Ogita, H.; Ito, J.; Nakagawa, T.; Furuta, I.; Kita, T.; Omori, K.; Yamamoto, N. Hearing preservation at low frequencies by insulin-like growth factor 1 in a guinea pig model of cochlear implantation. Hear. Res. 2018, 368, 92–108. [Google Scholar] [CrossRef]
- Yamahara, K.; Asaka, N.; Kita, T.; Kishimoto, I.; Matsunaga, M.; Yamamoto, N.; Omori, K.; Nakagawa, T. Insulin-like growth factor 1 promotes cochlear synapse regeneration after excitotoxic trauma in vitro. Hear. Res. 2019, 374, 5–12. [Google Scholar] [CrossRef]
- Iwai, K.; Nakagawa, T.; Endo, T.; Matsuoka, Y.; Kita, T.; Kim, T.S.; Tabata, Y.; Ito, J. Cochlear protection by local insulin-like growth factor-1 application using biodegradable hydrogel. Laryngoscope 2006, 116, 529–533. [Google Scholar] [CrossRef]
- Lee, K.Y.; Nakagawa, T.; Okano, T.; Hori, R.; Ono, K.; Tabata, Y.; Lee, S.H.; Ito, J. Novel therapy for hearing loss: Delivery of insulin-like growth factor 1 to the cochlea using gelatin hydrogel. Otol. Neurotol. 2007, 28, 976–981. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, Y.Y.; Chen, H.C.; Kuo, C.Y.; Liao, A.H.; Chou, Y.L.; Hung, C.L.; Shih, C.P.; Wang, C.H. Ultrasound microbubbles enhance the efficacy of insulin-like growth factor-1 therapy for the treatment of noise-induced hearing loss. Molecules 2021, 26, 3626. [Google Scholar] [CrossRef]
- Fujiwara, T.; Hato, N.; Nakagawa, T.; Tabata, Y.; Yoshida, T.; Komobuchi, H.; Takeda, S.; Hyodo, J.; Hakuba, N.; Gyo, K. Insulin-like growth factor I treatment via hydrogels rescues cochlear hair cells from ischemic injury. Neuroreport 2008, 19, 1585–1588. [Google Scholar] [CrossRef]
- Nakagawa, T.; Ogino-Nishimura, E.; Hiraumi, H.; Sakamoto, T.; Yamamoto, N.; Ito, J. Audiometric outcomes of topical IGF1 treatment for sudden deafness refractory to systemic steroids. Otol. Neurotol. 2012, 33, 941–946. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sakamoto, T.; Hiraumi, H.; Kikkawa, Y.S.; Yamamoto, N.; Hamaguchi, K.; Ono, K.; Yamamoto, M.; Tabata, Y.; Teramukai, S.; et al. Topical insulin-like growth factor 1 treatment using gelatin hydrogels for glucocorticoid-resistant sudden sensorineural hearing loss: A prospective clinical trial. BMC Med. 2010, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Kling, R.N.; Demers, P.A.; Alamgir, H.; Davies, H.W. Noise exposure and serious injury to active sawmill workers in British Columbia. Occup. Environ. Med. 2012, 69, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Malfeld, K.; Baumhoff, P.; Volk, H.A.; Lenarz, T.; Scheper, V. Local Long-Term Inner Ear Drug Delivery in Normal Hearing Guinea Pig—An Animal Model to Develop Preventive Treatment for Noise-Induced Hearing Loss. Biomolecules 2022, 12, 1427. [Google Scholar] [CrossRef] [PubMed]
- Hickman, T.T.; Hashimoto, K.; Liberman, L.D.; Liberman, M.C. Synaptic migration and reorganization after noise exposure suggests regeneration in a mature mammalian cochlea. Sci. Rep. 2020, 10, 19945. [Google Scholar] [CrossRef]
- Shinomori, Y.; Spack, D.S.; Jones, D.D.; Kimura, R.S. Volumetric and dimensional analysis of the guinea pig inner ear. Ann. Otol. Rhinol. Laryngol. 2001, 110, 91–98. [Google Scholar] [CrossRef]
- Furman, A.C.; Kujawa, S.G.; Charles Liberman, M. Noise-induced cochlear neuropathy is selective for fibers with low spontaneous rates. J. Neurophysiol. 2013, 110, 577–586. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.; Shi, L.; Almuklass, A.; He, T.; Aiken, S.; Bance, M.; Yin, S.; Wang, J. Silent Damage of Noise on Cochlear Afferent Innervation in Guinea Pigs and the Impact on Temporal Processing. PLoS ONE 2012, 7, e0171448. [Google Scholar] [CrossRef]
- Brigande, J.V.; Heller, S. Quo vadis, hair cell regeneration? Nat. Neurosci. 2009, 12, 679–685. [Google Scholar] [CrossRef]
- Sukhanov, S.; Higashi, Y.; Shai, S.Y.; Vaughn, C.; Mohler, J.; Li, Y.; Song, Y.H.; Titterington, J.; Delafontaine, P. IGF-1 reduces inflammatory responses, suppresses oxidative stress, and decreases atherosclerosis progression in ApoE-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2684–2690. [Google Scholar] [CrossRef]
- Higashi, Y.; Pandey, A.; Goodwin, B.; Delafontaine, P. Insulin-like growth factor-1 regulates glutathione peroxidase expression and activity in vascular endothelial cells: Implications for atheroprotective actions of insulin-like growth factor-1. Biochim. Biophys. Acta Mol. Basis Dis. 2013, 1832, 391–399. [Google Scholar] [CrossRef]
- El Ayadi, A.; Zigmond, M.J.; Smith, A.D. IGF-1 protects dopamine neurons against oxidative stress: Association with changes in phosphokinases. Exp. Brain Res. 2016, 234, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.; Machulik, A.; Moller, R.; Fuchs, J.; Amarjargal, N.; Ungethüm, U.; Kuban, R.J.; Szczepek, A.J.; Haupt, H.; Mazurek, B. mRNA expression of members of the IGF system in the organ of Corti, the modiolus and the stria vascularis of newborn rats. Growth Factors 2008, 26, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Siddle, K. Signalling by insulin and IGF receptors: Supporting acts and new players. J. Mol. Endocrinol. 2011, 47, R1–R10. [Google Scholar] [CrossRef] [PubMed]
- García-Mato, Á.; Cervantes, B.; Murillo-Cuesta, S.; De La Rosa, L.R.; Varela-Nieto, I. Insulin-like growth factor 1 signaling in mammalian hearing. Genes 2021, 12, 1553. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Yamamoto, N.; Nakagawa, T.; Ito, J. Insulin-like growth factor 1 induces the transcription of Gap43 and Ntn1 during hair cell protection in the neonatal murine cochlea. Neurosci. Lett. 2014, 560, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Yamamoto, N.; Nakagawa, T.; Ito, J. Insulin-like growth factor 1 inhibits hair cell apoptosis and promotes the cell cycle of supporting cells by activating different downstream cascades after pharmacological hair cell injury in neonatal mice. Mol. Cell. Neurosci. 2013, 56, 29–38. [Google Scholar] [CrossRef]
- Chen, J.; Yuan, H.; Talaska, A.E.; Hill, K.; Sha, S.H. Increased Sensitivity to Noise-Induced Hearing Loss by Blockade of Endogenous PI3K/Akt Signaling. JARO J. Assoc. Res. Otolaryngol. 2015, 16, 347–356. [Google Scholar] [CrossRef]
- Kurioka, T.; Matsunobu, T.; Niwa, K.; Tamura, A.; Satoh, Y.; Shiotani, A. Activated protein C rescues the cochlea from noise-induced hearing loss. Brain Res. 2014, 1583, 201–210. [Google Scholar] [CrossRef]
- Maeda, Y.; Fukushima, K.; Omichi, R.; Kariya, S.; Nishizaki, K. Time Courses of Changes in Phospho- and Total- MAP Kinases in the Cochlea after Intense Noise Exposure. PLoS ONE 2013, 8, e58775. [Google Scholar] [CrossRef]
- Kurioka, T.; Matsunobu, T.; Satoh, Y.; Niwa, K.; Endo, S.; Fujioka, M.; Shiotani, A. ERK2 mediates inner hair cell survival and decreases susceptibility to noise-induced hearing loss. Sci. Rep. 2015, 5, 16839. [Google Scholar] [CrossRef]
- Russo, V.C.; Kobayashi, K.; Najdovska, S.; Baker, N.L.; Werther, G.A. Neuronal protection from glucose deprivation via modulation of glucose transport and inhibition of apoptosis: A role for the insulin-like growth factor system. Brain Res. 2004, 1009, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Lai, L.; Ye, Y.; Zheng, Y. Glucose Protects Cochlear Hair Cells Against Oxidative Stress and Attenuates Noise-Induced Hearing Loss in Mice. Neurosci. Bull. 2021, 37, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Varela-Nieto, I.; Morales-Garcia, J.A.; Vigil, P.; Diaz-Casares, A.; Gorospe, I.; Sánchez-Galiano, S.; Cañon, S.; Camarero, G.; Contreras, J.; Cediel, R.; et al. Trophic effects of insulin-like growth factor-I (IGF-I) in the inner ear. Hear. Res. 2004, 196, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Yin, Y.; Chang, R.; Gong, X.; He, X.; Xie, Z. Effects of insulin-like growth factor 1 on synaptic excitability in cultured rat hippocampal neurons. Exp. Neurol. 2007, 205, 222–229. [Google Scholar] [CrossRef]
- Song, F.; Gan, B.; Wang, N.; Wang, Z.; Xu, A. Hidden hearing loss is associated with loss of ribbon synapses of cochlea inner hair cells. Biosci. Rep. 2021, 41, bsr20201637. [Google Scholar] [CrossRef]
- Lin, H.W.; Furman, A.C.; Kujawa, S.G.; Liberman, M.C. Primary neural degeneration in the guinea pig cochlea after reversible noise-induced threshold shift. JARO J. Assoc. Res. Otolaryngol. 2011, 12, 605–616. [Google Scholar] [CrossRef]
- Shi, L.; Liu, K.; Wang, H.; Zhang, Y.; Hong, Z.; Wang, M.; Wang, X.; Jiang, X.; Yang, S. Noise induced reversible changes of cochlear ribbon synapses contribute to temporary hearing loss in mice. Acta Otolaryngol. 2015, 135, 1093–1102. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, N.; Yan, K.; Liu, X.; Zhang, Y.; Hong, Z.; Wang, M.; Yin, Q.; Wu, F.; Lei, Y.; et al. Inner hair cell ribbon synapse plasticity might be molecular basis of temporary hearing threshold shifts in mice. Int. J. Clin. Exp. Pathol. 2015, 8, 8680–8691. [Google Scholar]
- Wu, C.C.; Brugeaud, A.; Seist, R.; Lin, H.C.; Yeh, W.H.; Petrillo, M.; Coppola, G.; Edge, A.S.B.; Stankovic, K.M. Altered expression of genes regulating inflammation and synaptogenesis during regrowth of afferent neurons to cochlear hair cells. PLoS ONE 2020, 15, e0238578. [Google Scholar] [CrossRef]
- Gao, L.; Kita, T.; Katsuno, T.; Yamamoto, N.; Omori, K.; Nakagawa, T. Insulin-Like Growth Factor 1 on the Maintenance of Ribbon Synapses in Mouse Cochlear Explant Cultures. Front. Cell. Neurosci. 2020, 14, 571155. [Google Scholar] [CrossRef]
- Michanski, S.; Smaluch, K.; Maria Steyer, A.; Chakrabarti, R.; Setz, C.; Oestreicher, D.; Fischer, C.; Möbius, W.; Moser, T.; Vogl, C.; et al. Mapping developmental maturation of inner hair cell ribbon synapses in the apical mouse cochlea. Proc. Natl. Acad. Sci. USA 2019, 116, 6415–6424. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, H.A.; Pich, A.; Schröder, A.; Scheper, V.; Lilli, G.; Reuter, G.; Lenarz, T. Proteome Analysis of Human Perilymph Using an Intraoperative Sampling Method. J. Proteome Res. 2017, 16, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, H.A.; Pich, A.; Prenzler, N.K.; Lenarz, T.; Harre, J.; Staecker, H.; Durisin, M.; Warnecke, A. Personalized Proteomics for Precision Diagnostics in Hearing Loss: Disease-Specific Analysis of Human Perilymph by Mass Spectrometry. ACS Omega 2021, 6, 21241–21254. [Google Scholar] [CrossRef]
- Warnecke, A.; Prenzler, N.K.; Schmitt, H.; Daemen, K.; Keil, J.; Dursin, M.; Lenarz, T.; Falk, C.S. Defining the Inflammatory Microenvironment in the Human Cochlea by Perilymph Analysis: Toward Liquid Biopsy of the Cochlea. Front. Neurol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Palmer, J.C.; Lord, M.S.; Pinyon, J.L.; Wise, A.K.; Lovell, N.H.; Carter, P.M.; Enke, Y.L.; Housley, G.D.; Green, R.A. Comparing perilymph proteomes across species. Laryngoscope 2018, 128, E47–E52. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, Y.J.; Li, H.; Kim, H.; Suh, M.W.; Han, D.; Oh, S.H. In-depth proteome of perilymph in guinea pig model. Proteomics 2021, 21, 2000138. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Yamashita, D.; Irino, Y.; Kitamoto, J.; Fukuda, Y.; Inokuchi, G.; Hasegawa, S.; Otsuki, N.; Yoshida, M.; Nibu, K.I. Metabolomic profiling in inner ear fluid by gas chromatography/mass spectrometry in guinea pig cochlea. Neurosci. Lett. 2015, 606, 188–193. [Google Scholar] [CrossRef]
- Pirttilä, K.; Videhult, P.; Jakob, P.; Mikael, H.; Mikael, E.; Göran, H.; Arvidsson, T.; Pettersson, C. An LCMS - based untargeted metabolomics protocol for cochlear perilymph: Highlighting metabolic effects of hydrogen gas on the inner ear of noise exposed Guinea pigs. Metabolomics 2019, 15, 138. [Google Scholar] [CrossRef]
- Jongkamonwiwat, N.; Ramirez, M.A.; Edassery, S.; Wong, A.C.Y.; Yu, J.; Abbott, T.; Pak, K.; Ryan, A.F.; Savas, J.N. Noise Exposures Causing Hearing Loss Generate Proteotoxic Stress and Activate the Proteostasis Network. Cell Rep. 2020, 33, 108431. [Google Scholar] [CrossRef]
- Miao, L.; Zhang, J.; Yin, L.; Pu, Y. TMT-based quantitative proteomics reveals cochlear protein profile alterations in mice with noise-induced hearing loss. Int. J. Environ. Res. Public Health 2022, 19, 382. [Google Scholar] [CrossRef]
- Guo, H.L.; Chen, G.; Song, Z.L.; Sun, J.; Gao, X.H.; Han, Y.X. COL6A3 promotes cellular malignancy of osteosarcoma by activating the PI3K/AKT pathway. Rev. Assoc. Med. Bras. 2020, 66, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Yeo, S.W.; Troy, F.A. Expression of polysialylated neural cell adhesion molecules on adult stem cells after neuronal differentiation of inner ear spiral ganglion neurons. Biochem. Biophys. Res. Commun. 2014, 453, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Rønn, L.C.B.; Berezin, V.; Bock, E. The neural cell adhesion molecule in synaptic plasticity and ageing. Int. J. Dev. Neurosci. 2000, 18, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Ishikura, S.; Usami, N.; Araki, M.; Hara, A. Structural and functional characterization of rabbit and human L-gulonate 3-dehydrogenase. J. Biochem. 2005, 137, 303–314. [Google Scholar] [CrossRef]
- Rebillard, G.; Bryant, G.M. Effects of in vivo perfusion of glutamate dehydrogenase in the guinea pig cochlea on the VIIIth nerve compound action potential. Brain Res. 1989, 494, 379–382. [Google Scholar] [CrossRef]
- Ji, L.; Lee, H.J.; Wan, G.; Wang, G.P.; Zhang, L.; Sajjakulnukit, P.; Schacht, J.; Lyssiotis, C.A.; Corfas, G. Auditory metabolomics, an approach to identify acute molecular effects of noise trauma. Sci. Rep. 2019, 9, 9273. [Google Scholar] [CrossRef]
- Vecchio, L.M.; Sullivan, P.; Dunn, A.R.; Bermejo, M.K.; Fu, R.; Masoud, S.T.; Gregersen, E.; Urs, N.M.; Nazari, R.; Jensen, P.H.; et al. Enhanced tyrosine hydroxylase activity induces oxidative stress, causes accumulation of autotoxic catecholamine metabolites, and augments amphetamine effects in vivo. J. Neurochem. 2021, 158, 960–979. [Google Scholar] [CrossRef]
- Ma, Y.; Su, K.N.; Pfau, D.; Rao, V.S.; Wu, X.; Hu, X.; Leng, L.; Du, X.; Piecychna, M.; Bedi, K.; et al. Cardiomyocyte d-dopachrome tautomerase protects against heart failure. JCI Insight 2019, 4, e128900. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Qiu, P.; Luo, H. Proteomic profiling of proteins associated with methamphetamine-induced neurotoxicity in different regions of rat brain. Neurochem. Int. 2008, 52, 256–264. [Google Scholar] [CrossRef]
- Shigyo, M.; Kuboyama, T.; Sawai, Y.; Tada-Umezaki, M.; Tohda, C. Extracellular vimentin interacts with insulin-like growth factor 1 receptor to promote axonal growth. Sci. Rep. 2015, 5, 12055. [Google Scholar] [CrossRef]
- Laron, Z. Somatomedin-1 (recombinant insulin-like growth factor-1): Clinical pharmacology and potential treatment of endocrine and metabolic disorders. BioDrugs 1999, 11, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Klinger; Anin; Silbergeld; Eshet; Laron. Development of hyperandrogenism during treatment with insulin-like growth factor-I (IGF-I) in female patients with Laron syndrome. Clin. Endocrinol. 1998, 48, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Sahan, O.B.; Gunel-Ozcan, A. Hepatocyte Growth Factor and Insulin-like Growth Factor-1 based Cellular Therapies for Oxidative Stress Injury. Curr. Stem Cell Res. Ther. 2021, 16, 771–791. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Han, Z.; Naveena, K.; Lei, G.; Qiu, S.; Li, X.; Li, T.; Shi, X.; Zhuang, W.; Li, Y.; et al. ROS-Responsive Nanoparticle as a Berberine Carrier for OHC-Targeted Therapy of Noise-Induced Hearing Loss. ACS Appl. Mater. Interfaces 2021, 13, 7102–7114. [Google Scholar] [CrossRef]
- Soubry, A.; Il’yasova, D.; Sedjo, R.; Wang, F.; Byers, T.; Rosen, C.; Yashin, A.; Ukraintseva, S.; Haffner, S.; D’Agostino, R. Increase in circulating levels of IGF-1 and IGF-1/IGFBP-3 molar ratio over a decade is associated with colorectal adenomatous polyps. Int. J. Cancer 2012, 131, 512–517. [Google Scholar] [CrossRef][Green Version]
- Scheper, V.; Paasche, G.; Miller, J.M.; Warnecke, A.; Berkingali, N.; Lenarz, T.; Stöver, T. Effects of delayed treatment with combined GDNF and continuous electrical stimulation on spiral ganglioncell survival in deafened guinea pigs. J. Neurosci. Res. 2009, 87, 1389–1399. [Google Scholar] [CrossRef]
- Whitlon, D.S.; Young, H.; Barna, M.; Depreux, F.; Richter, C.P. Hearing differences in Hartley guinea pig stocks from two breeders. Hear. Res. 2019, 379, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Stieglitz, F.; Gerhard, R.; Hönig, R.; Giehl, K.; Pich, A. TcdB of Clostridioides difficile Mediates RAS-Dependent Necrosis in Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 4258. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Palmowski, P.; Watson, R.; Europe-Finner, G.N.; Karolczak-Bayatti, M.; Porter, A.; Treumann, A.; Taggart, M.J. The Generation of a Comprehensive Spectral Library for the Analysis of the Guinea Pig Proteome by SWATH-MS. Proteomics 2019, 19, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Green, S.H. Functional role of neurotrophin-3 in synapse regeneration by spiral ganglion neurons on inner hair cells after excitotoxic trauma in vitro. J. Neurosci. 2011, 31, 7938–7949. [Google Scholar] [CrossRef] [PubMed]
- Malfeld, K.; Armbrecht, N.; Volk, H.A.; Lenarz, T.; Scheper, V. In Situ 3D-Imaging of the Inner Ear Synapses with a Cochlear Implant. Life 2021, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.; Wendler, O.; Frischknecht, R.; Shigemoto, R.; Schulze, H.; Enz, R. Localization of group II and III metabotropic glutamate receptors at pre- and postsynaptic sites of inner hair cell ribbon synapses. FASEB J. 2019, 33, 13734–13746. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Protein Name | ↑/↓ | p-Value | Ratio (Logarithm to Base 2) |
|---|---|---|---|---|
| H0VIY0 | Collagen type VI alpha 3 chain | ↑ | 0.029 | 2.003 |
| H0VRB7 | Ig-like domain-containing protein | ↑ | 0.022 | 2.282 |
| H0UXT4 | Neural cell adhesion molecule 2 | ↑ | 0.002 | 2.654 |
| UniProt ID | Protein Name | ↑/↓ | p-Value | Ratio (Logarithm to Base 2) |
|---|---|---|---|---|
| H0VIY0 | Collagen type VI alpha 3 chain | ↑ | 0.003 | 1.768 |
| H0V0H2 | Crystallin lambda 1 | ↑ | 0.01 | 1.549 |
| A0A286XVA7 | Proteasome subunit beta | ↓ | 0.012 | −1.021 |
| H0VFS3 | Inosine-5’-monophosphate dehydrogenase | ↓ | 0.008 | −2.219 |
| A0A286Y4U7 * | D-dopachrome decarboxylase | ↑ | 0.034 | 1.422 |
| A0A286Y3H7 | Pyridoxal kinase | ↑ | 0.007 | 1.301 |
| A0A286XP66 | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein gamma | ↓ | 0.016 | −0.843 |
| H0UVC5 | Vimentin | ↓ | 0.022 | −0.809 |
| H0UT45 * | Alpha-2-macroglobulin | ↑ | 0.044 | 0.851 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malfeld, K.; Armbrecht, N.; Pich, A.; Volk, H.A.; Lenarz, T.; Scheper, V. Prevention of Noise-Induced Hearing Loss In Vivo: Continuous Application of Insulin-like Growth Factor 1 and Its Effect on Inner Ear Synapses, Auditory Function and Perilymph Proteins. Int. J. Mol. Sci. 2023, 24, 291. https://doi.org/10.3390/ijms24010291
Malfeld K, Armbrecht N, Pich A, Volk HA, Lenarz T, Scheper V. Prevention of Noise-Induced Hearing Loss In Vivo: Continuous Application of Insulin-like Growth Factor 1 and Its Effect on Inner Ear Synapses, Auditory Function and Perilymph Proteins. International Journal of Molecular Sciences. 2023; 24(1):291. https://doi.org/10.3390/ijms24010291
Chicago/Turabian StyleMalfeld, Kathrin, Nina Armbrecht, Andreas Pich, Holger A. Volk, Thomas Lenarz, and Verena Scheper. 2023. "Prevention of Noise-Induced Hearing Loss In Vivo: Continuous Application of Insulin-like Growth Factor 1 and Its Effect on Inner Ear Synapses, Auditory Function and Perilymph Proteins" International Journal of Molecular Sciences 24, no. 1: 291. https://doi.org/10.3390/ijms24010291
APA StyleMalfeld, K., Armbrecht, N., Pich, A., Volk, H. A., Lenarz, T., & Scheper, V. (2023). Prevention of Noise-Induced Hearing Loss In Vivo: Continuous Application of Insulin-like Growth Factor 1 and Its Effect on Inner Ear Synapses, Auditory Function and Perilymph Proteins. International Journal of Molecular Sciences, 24(1), 291. https://doi.org/10.3390/ijms24010291