

Yeast Mannan-Rich Fraction Modulates Endogenous Reactive Oxygen Species Generation and Antibiotic Sensitivity in Resistant E. coli

 , and
, and
Abstract
1. Introduction
2. Results
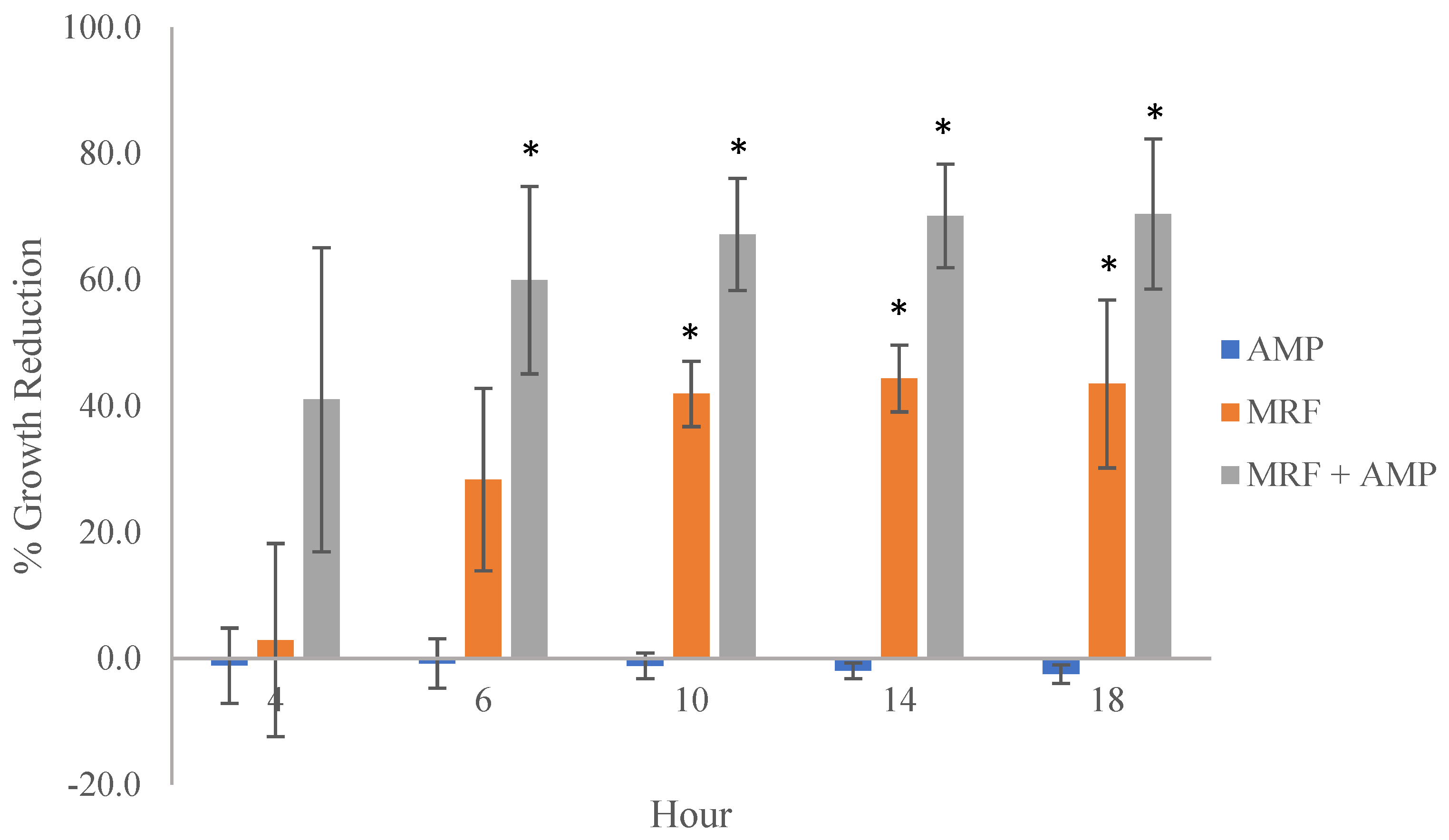
2.1. MRF-Mediated ROS Production Enhances Antibiotic Activity against E. coli
2.2. MRF Effects Translational Change in the TCA Cycle
2.3. Translational Impact of MRF on Other Central Metabolic Pathways
2.4. The Effect of MRF on NAD(H/+)
2.5. The Effect of MRF Supplementation on Kinetic Growth of E. coli
3. Discussion
4. Materials and Methods
4.1. Bacteria and Yeast Cell Wall Preparation
4.2. Inoculum Preparation and Storage
4.3. Growth and Maintenance of Recombinant E. coli
4.4. ROS Quantification
4.5. Preparation of Samples for Proteomic Analysis
4.6. Proteomic Analysis
4.7. NAD(H/+) Determination
4.8. Kinetic Growth Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat. Rev. Microbiol. 2006, 4, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Oyofo, B.A.; DeLoach, J.R.; Corrier, D.E.; Norman, J.O.; Ziprin, R.L.; Mollenhauer, H.H. Prevention of Salmonella typhimurium colonization of broilers with D-mannose. Poult. Sci. 1989, 68, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Ofek, I.; Beachey, E.H. Mannose binding and epithelial cell adherence of Escherichia coli. Infect. Immun. 1978, 22, 247–254. [Google Scholar] [CrossRef]
- Spring, P.; Wenk, C.; Connolly, A.; Kiers, A. A review of 733 published trials on Bio-Mos®, a mannan oligosaccharide, and Actigen®, a second generation mannose rich fraction, on farm and companion animals. J. Appl. Anim. Nutr. 2015, 3, e8. [Google Scholar] [CrossRef]
- Spring, P.; Wenk, C.; Dawson, K.A.; Newman, K.E. The effects of dietary mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of Salmonella-challenged broiler chicks. Poult. Sci. 2000, 79, 205–211. [Google Scholar] [CrossRef]
- Corrigan, A.; Fay, B.J.; Corcionivoschi, N.; Murphy, R.A. Effect of yeast mannan-rich fractions on reducing Campylobacter colonization in broiler chickens. J. Appl. Poult. Res. 2017, 26, 350–357. [Google Scholar] [CrossRef]
- McCaffrey, C.; Corrigan, A.; Moynagh, P.; Murphy, R. Effect of yeast cell wall supplementation on intestinal integrity, digestive enzyme activity and immune traits of broilers. Br. Poult. Sci. 2021, 62, 771–782. [Google Scholar] [CrossRef]
- Smith, H.; Grant, S.; Parker, J.; Murphy, R. Yeast cell wall mannan rich fraction modulates bacterial cellular respiration potentiating antibiotic efficacy. Sci. Rep. 2020, 10, 21880. [Google Scholar] [CrossRef]
- Belenky, P.; Jonathan, D.Y.; Porter, C.B.; Cohen, N.R.; Lobritz, M.A.; Ferrante, T.; Jain, S.; Korry, B.J.; Schwarz, E.G.; Walker, G.C.; et al. Bactericidal antibiotics induce toxic metabolic perturbations that lead to cellular damage. Cell Rep. 2015, 13, 968–980. [Google Scholar] [CrossRef]
- Van Acker, H.; Coenye, T. The role of reactive oxygen species in antibiotic-mediated killing of bacteria. Trends Microbiol. 2017, 25, 456–466. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gomez, K.; Flores, N.; Castaneda, H.M.; Martinez-Batallar, G.; Hernandez-Chavez, G.; Ramirez, O.T.; Gosset, G.; Encarnacion, S.; Bolivar, F. New insights into Escherichia coli metabolism: Carbon scavenging, acetate metabolism and carbon recycling responses during growth on glycerol. Microb. Cell Factories 2012, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Flores, N.; Escalante, A.; de Anda, R.; Báez-Viveros, J.L.; Merino, E.; Franco, B.; Georgellis, D.; Gosset, G.; Bolívar, F. New insights into the role of sigma factor RpoS as revealed in Escherichia coli strains lacking the phosphoenolpyruvate:carbohydrate phosphotransferase system. J. Mol. Microbiol. Biotechnol. 2008, 14, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Flores, N.; Flores, S.; Escalante, A.; de Anda, R.; Leal, L.; Malpica, R.; Georgellis, D.; Gosset, G.; Bolívar, F. Adaptation for fast growth on glucose by differential expression of central carbon metabolism and gal regulon genes in an Escherichia coli strain lacking the phosphoenolpyruvate:carbohydrate phosphotransferase system. Metab. Eng. 2005, 7, 70–87. [Google Scholar] [CrossRef]
- Olvera, L.; Mendoza-Vargas, A.; Flores, N.; Olvera, M.; Sigala, J.C.; Gosset, G.; Morett, E.; Bolívar, F. Transcription analysis of central metabolism genes in Escherichia coli. Possible roles of σ38 in their expression, as a response to carbon limitation. PLoS ONE 2009, 4, e7466. [Google Scholar] [CrossRef]
- Förster, A.H.; Gescher, J. Metabolic engineering of Escherichia coli for production of mixed-acid fermentation end products. Front. Bioeng. Biotechnol. 2014, 2. [Google Scholar] [CrossRef]
- Casey, P.G.; Gardiner, G.E.; Casey, G.; Bradshaw, B.; Lawlor, P.G.; Lynch, P.B.; Leonard, F.C.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F.; et al. A five-strain probiotic combination reduces pathogen shedding and alleviates disease signs in pigs challenged with Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2007, 73, 1858–1863. [Google Scholar] [CrossRef]
- Kim, Y.; Ingram, L.O.; Shanmugam, K.T. Dihydrolipoamide dehydrogenase mutation alters the NADH sensitivity of pyruvate dehydrogenase complex of Escherichia coli K-12. J. Bacteriol. 2008, 190, 3851–3858. [Google Scholar] [CrossRef]
- Cronan, J.E., Jr.; Laporte, D. Tricarboxylic acid cycle and glyoxylate bypass. EcoSal Plus 2005, 1. [Google Scholar] [CrossRef]
- Martínez, J.L.; Rojo, F. Metabolic regulation of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Li, A.H.; Sassetti, C.M. Metabolic regulation of mycobacterial growth and antibiotic sensitivity. PLoS Biol. 2011, 9, e1001065. [Google Scholar] [CrossRef] [PubMed]
- McBee, M.E.; Chionh, Y.H.; Sharaf, M.L.; Ho, P.; Cai, M.W.L.; Dedon, P.C. Production of superoxide in bacteria Is stress- and cell state-dependent: A gating-optimized flow cytometry method that minimizes ROS measurement artifacts with fluorescent dyes. Front. Microbiol. 2017, 8, 459. [Google Scholar] [CrossRef] [PubMed]
- Courcelle, J.; Hanawalt, P.C. RecA-dependent recovery of arrested DNA replication forks. Annu. Rev. Genet. 2003, 37, 611–646. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Thomsen, L.E.; Gaggero, C.; Mosseri, R.; Ingmer, H.; Cohen, S.N. SOS response induction by beta-lactams and bacterial defense against antibiotic lethality. Science 2004, 305, 1629–1631. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Akhova, A.V.; Tkachenko, A.G. ATP/ADP alteration as a sign of the oxidative stress development in Escherichia coli cells under antibiotic treatment. FEMS Microbiol. Lett. 2014, 353, 69–76. [Google Scholar] [CrossRef]
- Seshasayee, A.S.; Bertone, P.; Fraser, G.M.; Luscombe, N.M. Transcriptional regulatory networks in bacteria: From input signals to output responses. Curr. Opin. Microbiol. 2006, 9, 511–519. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef]
- Gruer, M.J.; Bradbury, A.J.; Guest, J.R. Construction and properties of aconitase mutants of Escherichia coli. Microbiology 1997, 143 Pt 6, 1837–1846. [Google Scholar] [CrossRef]
- Helling, R.B.; Kukora, J.S. Nalidixic acd-resistant mutants of Escherichia coli deficient in isocitrate dehydrogenase. J. Bacteriol. 1971, 105, 1224–1226. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Jung, J.; Jang, I.A.; Madsen, E.L.; Park, W. Role of glyoxylate shunt in oxidative stress response. J. Biol. Chem. 2016, 291, 11928–11938. [Google Scholar] [CrossRef] [PubMed]
- Eng, R.H.; Padberg, F.T.; Smith, S.M.; Tan, E.N.; Cherubin, C.E. Bactericidal effects of antibiotics on slowly growing and nongrowing bacteria. Antimicrob. Agents Chemother. 1991, 35, 1824–1828. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A. Glycolysis for microbiome generation. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Altman, E.; Sangurdekar, D.P.; Khodursky, A.B.; Eiteman, M.A. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl. Environ. Microbiol. 2006, 72, 3653–3661. [Google Scholar] [CrossRef]
- Bekker, M.; de Vries, S.; Ter Beek, A.; Hellingwerf, K.J.; de Mattos, M.J.T. Respiration of Escherichia coli can be fully uncoupled via the nonelectrogenic terminal cytochrome bd-II oxidase. J. Bacteriol. 2009, 191, 5510–5517. [Google Scholar] [CrossRef]
- Deckers-Hebestreit, G.; Altendorf, K. The F0F1-type ATP synthases of bacteria: Structure and function of the F0 complex. Annu. Rev. Microbiol. 1996, 50, 791–824. [Google Scholar] [CrossRef]
- Koebmann, B.J.; Westerhoff, H.V.; Snoep, J.L.; Nilsson, D.; Jensen, P.R. The glycolytic flux in Escherichia coli is controlled by the demand for ATP. J. Bacteriol. 2002, 184, 3909–3916. [Google Scholar] [CrossRef]
- Nakanishi-Matsui, M.; Sekiya, M.; Futai, M. ATP synthase from Escherichia coli: Mechanism of rotational catalysis, and inhibition with the ε subunit and phytopolyphenols. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 129–140. [Google Scholar] [CrossRef]
- Dadi, P.K.; Ahmad, M.; Ahmad, Z. Inhibition of ATPase activity of Escherichia coli ATP synthase by polyphenols. Int. J. Biol. Macromol. 2009, 45, 72–79. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E.; Davletshin, A.; Mastronicola, D.; Sarti, P.; Giuffrè, A. Cytochrome bd oxidase from Escherichia coli displays high catalase activity: An additional defense against oxidative stress. FEBS Lett. 2013, 587, 2214–2218. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkova, N.N.; Starkov, A.A. Metabolic ROS signaling: To immunity and beyond. Biochemistry 2020, 85, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- San, K.Y.; Bennett, G.N.; Berríos-Rivera, S.J.; Vadali, R.V.; Yang, Y.T.; Horton, E.; Rudolph, F.B.; Sariyar, B.; Blackwood, K. Metabolic engineering through cofactor manipulation and its effects on metabolic flux redistribution in Escherichia coli. Metab. Eng. 2002, 4, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Eiteman, M.A.; Altman, R.; Altman, E. High glycolytic flux improves pyruvate production by a metabolically engineered Escherichia coli strain. Appl. Environ. Microbiol. 2008, 74, 6649–6655. [Google Scholar] [CrossRef]
- Berríos-Rivera, S.J.; Bennett, G.N.; San, K.Y. Metabolic engineering of Escherichia coli: Increase of NADH availability by overexpressing an NAD(+)-dependent formate dehydrogenase. Metab. Eng. 2002, 4, 217–229. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, L.; Yang, F.; Lin, X.; Zhang, S.; Zhao, Z.K. Determining the extremes of the cellular NAD(H) level by using an Escherichia coli NAD(+)-auxotrophic mutant. Appl. Environ. Microbiol. 2011, 77, 6133–6140. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Collins, J.J. Role of reactive oxygen species in antibiotic action and resistance. Curr. Opin. Microbiol. 2009, 12, 482–489. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Citric Acid Cycle | MRF | AMP | MRF + AMP | Highest Mean Condition | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | Protein | PC | UP | PSM | SC (%) | Fold Change a | p-Value | Fold Change a | p-Value | Fold Change a | p-Value | |
| P0A9Q3 | ARCA | 4 | 3 | 41 | 27 | +0.52 | 0.19 | −0.01 | 0.92 | +0.78 | 0.03 | MRF + AMP |
| Q8X722 | ICD | 8 | 8 | 62 | 23 | −0.20 | 0.30 | −0.13 | 0.40 | +0.72 | 0.03 | MRF + AMP |
| A0A0H3JCM9 | GLTA | 2 | 2 | 17 | 6 | +0.37 | 0.22 | +0.46 | 0.17 | +1.65 | 0.05 | MRF + AMP |
| Q8X957 | ACNB | 9 | 9 | 127 | 21 | +0.04 | 0.73 | −0.01 | 0.92 | +0.62 | 0.02 | MRF + AMP |
| P0AFG5 | SUCA | 3 | 3 | 40 | 7 | +0.27 | 0.14 | +0.65 | 0.06 | +1.59 | 0.00 | MRF + AMP |
| Q8XDQ0 | FRDA | 6 | 6 | 47 | 12 | +0.26 | 0.18 | −0.12 | 0.52 | +0.83 | 0.01 | MRF + AMP |
| Q8X609 | ACEB | 3 | 3 | 43 | 19 | −0.26 | 0.10 | −0.51 | 0.01 | +0.15 | 0.16 | MRF +AMP |
| P61891 | MDH | 3 | 2 | 23 | 21 | +0.60 | 0.02 | −0.05 | 0.54 | +0.54 | 0.02 | MRF |
| Q8XDS0 | ASPA | 4 | 4 | 31 | 12 | −0.27 | 0.23 | −0.31 | 0.15 | +1.02 | 0.06 | MRF + AMP |
| Q8X743 | PPC | 6 | 6 | 108 | 18 | +0.52 | 0.17 | +0.31 | 0.21 | +0.45 | 0.12 | MRF |
| Other Metabolic Pathways | MRF | AMP | MRF + AMP | Highest Mean Condition | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | Protein | PC | UP | PSM | SC (%) | Fold Change a | p-Value | Fold Change a | p-Value | Fold Change a | p-Value | |
| Acetyl-coenzyme A | ||||||||||||
| P0AC99 | SATP | 1 | 1 | 3 | 5 | 0.03 | 0.92 | 0.88 | 0.18 | 4.14 | 0.04 | MRF + AMP |
| EMP Pathway | ||||||||||||
| P0A860 | TPIA | 2 | 2 | 15 | 5 | 3.20 | 0.01 | −0.12 | 0.61 | 3.08 | 0.08 | MRF |
| P62709 | GPMA | 7 | 7 | 39 | 31 | 0.93 | 0.04 | 0.16 | 0.50 | 0.4 | 0.19 | MRF |
| Q8XDE9 | GPMI | 9 | 9 | 78 | 24 | 0.85 | 0.05 | 0.16 | 0.09 | 1.09 | 0.08 | MRF + AMP |
| P0AD62 | PYKF | 12 | 12 | 79 | 27 | 0.41 | 0.03 | 0.06 | 0.52 | 0.68 | 0.00 | MRF + AMP |
| Q8X7H3 | FBAB | 5 | 5 | 35 | 21 | 0.91 | 0.02 | −0.30 | 0.14 | 0.3 | 0.14 | MRF |
| Q8XCF0 | FBP | 4 | 4 | 23 | 17 | 0.69 | 0.02 | 0.13 | 0.05 | 0.92 | 0.14 | MRF + AMP |
| Q8X926 | PGL | 3 | 3 | 12 | 15 | 0.64 | 0.11 | 0.3 | 0.13 | 0.79 | 0.03 | MRF + AMP |
| P0AB69 | PNTB | 3 | 3 | 15 | 7 | 0.20 | 0.34 | 0.03 | 0.84 | 1.27 | 0.03 | MRF + AMP |
| P0A797 | PFKA | 7 | 7 | 36 | 20 | 1.17 | 0.02 | 0.38 | 0.08 | 1.86 | 0.16 | MRF + AMP |
| Q8XE22 | PFKB | 3 | 3 | 17 | 12 | 1.08 | 0.02 | −0.19 | 0.13 | 0.34 | 0.35 | MRF |
| Q8XDE9 | GPMI | 9 | 9 | 78 | 24 | 0.85 | 0.05 | 0.16 | 0.09 | 1.09 | 0.08 | MRF + AMP |
| OxPhos Pathway | ||||||||||||
| P0AFC9 | NUOB | 2 | 2 | 12 | 6 | 1.41 | 0.01 | 1.33 | 0.09 | 4.43 | 0.09 | MRF + AMP |
| Q8XCX1 | NUOF | 2 | 2 | 11 | 5 | −0.44 | 0.03 | −0.19 | 0.03 | 0.3 | 0.03 | MRF + AMP |
| Q8XCX2 | NUOG | 7 | 7 | 34 | 9 | 0.03 | 0.83 | 0.07 | 0.91 | 1.65 | 0.03 | MRF + AMP |
| P0ABJ0 | CYOB | 1 | 1 | 7 | 1 | −0.70 | 0.06 | −0.57 | 0.02 | 0.01 | 0.86 | MRF + AMP |
| P58646 | ATPC | 3 | 3 | 39 | 23 | 0.25 | 0.53 | −0.74 | 0.03 | −0.54 | 0.15 | MRF |
| P0ABA8 | ATPG | 7 | 7 | 55 | 23 | −0.17 | 0.33 | −0.17 | 0.33 | 1.21 | 0.01 | MRF + AMP |
| P0ABB2 | ATPA | 9 | 8 | 62 | 27 | −0.06 | 0.46 | −0.36 | 0.02 | 0.2 | 0.30 | MRF + AMP |
| P0ABA2 | ATPF | 3 | 3 | 14 | 6 | 2.03 | 0.06 | −0.05 | 0.02 | 2.03 | 0.00 | MRF |
| P0ABB6 | ATPD | 14 | 14 | 104 | 40 | 0.14 | 0.02 | −0.12 | 0.01 | 1.17 | 0.16 | MRF + AMP |
| P0A7G8 | RECA | 3 | 3 | 14 | 6 | 8.70 | 0.06 | 1.58 | 0.06 | 1.72 | 0.00 | MRF |
| Stress and Signaling | ||||||||||||
| P0AGD2 | SODC | 4 | 4 | 31 | 28 | 1.07 | 0.83 | 1.03 | 0.91 | 2.65 | 0.03 | MRF + AMP |
| Q8XBT4 | AHPF | 9 | 9 | 49 | 17 | 0.35 | 0.07 | 0.09 | 0.50 | 1.39 | 0.01 | MRF + AMP |
| P0ACG0 | H-NS | 5 | 5 | 13 | 28 | 1.20 | 0.21 | 0.35 | 0.12 | 1.15 | 0.02 | MRF |
| P0ACJ2 | LRP | 4 | 4 | 19 | 12 | 0.56 | 0.05 | 0.31 | 0.06 | 0.93 | 0.10 | MRF + AMP |
| P58162 | DSBD | 1 | 1 | 8 | 3 | 2.69 | 0.00 | 0.9 | 0.01 | 2.79 | 0.03 | MRF + AMP |
| P58320 | DSBG | 3 | 3 | 29 | 14 | 0.68 | 0.02 | −0.03 | 0.69 | 1.21 | 0.09 | MRF + AMP |
| P0C0V1 | DEGP | 5 | 5 | 30 | 12 | 0.15 | 0.18 | 0.29 | 0.05 | 0.48 | 0.05 | MRF + AMP |
| Q8XE55 | PPID | 15 | 15 | 95 | 26 | 0.26 | 0.30 | −0.23 | 0.05 | 0.77 | 0.00 | MRF + AMP |
| Control | AMP | MRF | AMP + MRF | |
|---|---|---|---|---|
| NADH | 1.720 ± 0.266 | 1.758 ± 0.119 | 2.700 ± 0.176 ab | 2.553 ± 0.201 ab |
| NAD+ | 5.095 ± 0.022 | 4.910 ± 0.336 | 6.280 ± 0.004 ab | 6.387 ± 0.312 ab |
| NADH/NAD+ | 0.340 ± 0.051 | 0.358 ± 0.039 | 0.427 ± 0.028 a | 0.397 ± 0.026 |
| NAD+/NADH | 2.983 ± 0.418 | 2.820 ± 0.292 | 2.352 ± 0.151 | 2.524 ± 0.161 |
| Total NAD(H/+) | 6.82 | 6.67 | 8.98 ab | 8.94 ab |
| % Change in total NAD(H/+) | - | −2.2 | 31.8 | 31.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, H.; Grant, S.; Meleady, P.; Henry, M.; O’Gorman, D.; Clynes, M.; Murphy, R. Yeast Mannan-Rich Fraction Modulates Endogenous Reactive Oxygen Species Generation and Antibiotic Sensitivity in Resistant E. coli. Int. J. Mol. Sci. 2023, 24, 218. https://doi.org/10.3390/ijms24010218
Smith H, Grant S, Meleady P, Henry M, O’Gorman D, Clynes M, Murphy R. Yeast Mannan-Rich Fraction Modulates Endogenous Reactive Oxygen Species Generation and Antibiotic Sensitivity in Resistant E. coli. International Journal of Molecular Sciences. 2023; 24(1):218. https://doi.org/10.3390/ijms24010218
Chicago/Turabian StyleSmith, Helen, Sharon Grant, Paula Meleady, Michael Henry, Donal O’Gorman, Martin Clynes, and Richard Murphy. 2023. "Yeast Mannan-Rich Fraction Modulates Endogenous Reactive Oxygen Species Generation and Antibiotic Sensitivity in Resistant E. coli" International Journal of Molecular Sciences 24, no. 1: 218. https://doi.org/10.3390/ijms24010218
APA StyleSmith, H., Grant, S., Meleady, P., Henry, M., O’Gorman, D., Clynes, M., & Murphy, R. (2023). Yeast Mannan-Rich Fraction Modulates Endogenous Reactive Oxygen Species Generation and Antibiotic Sensitivity in Resistant E. coli. International Journal of Molecular Sciences, 24(1), 218. https://doi.org/10.3390/ijms24010218