
Flavonoid Nobiletin Attenuates Cyclophosphamide-Induced Cystitis in Mice through Mechanisms That Involve Inhibition of IL-1β Induced Connexin 43 Upregulation and Gap Junction Communication in Urothelial Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
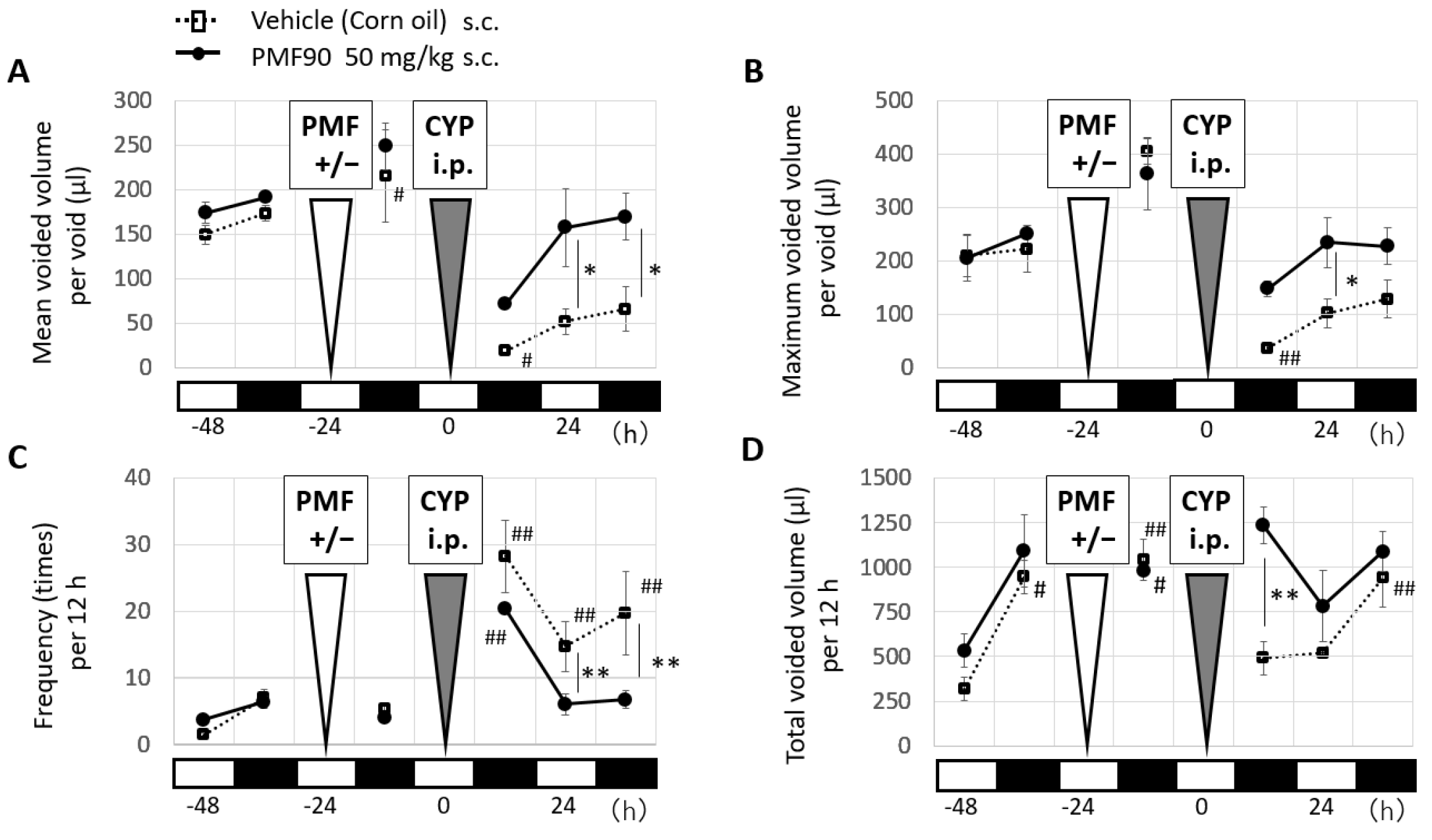
2.1. Prophylactic Treatment with PMF90 Improved Urinary Symptoms in the CYP-Induced Cystitis Mouse Model
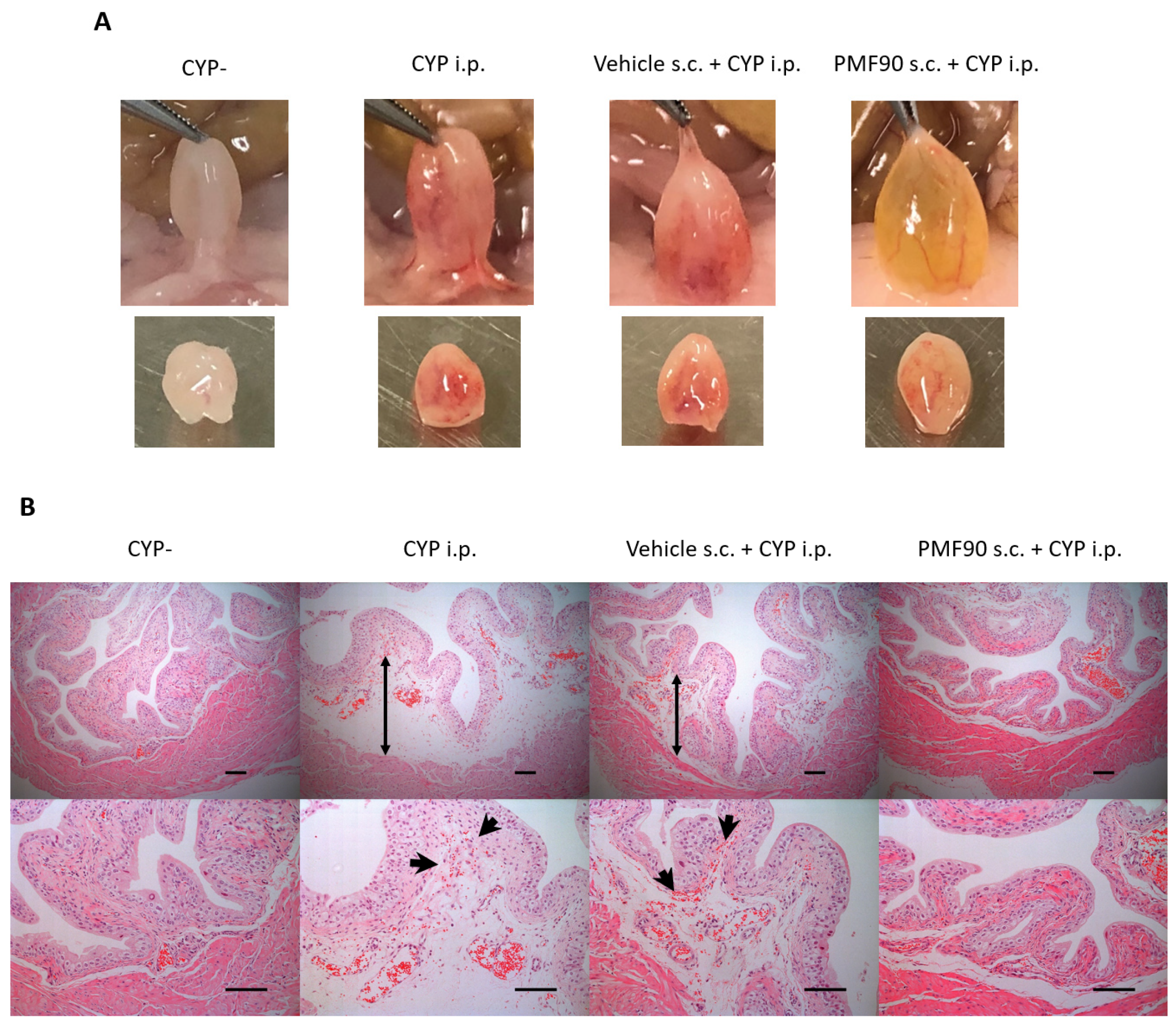
2.2. Prophylactic Administration of PMF90 Suppressed Bladder Inflammation in CYP-Treated Mice
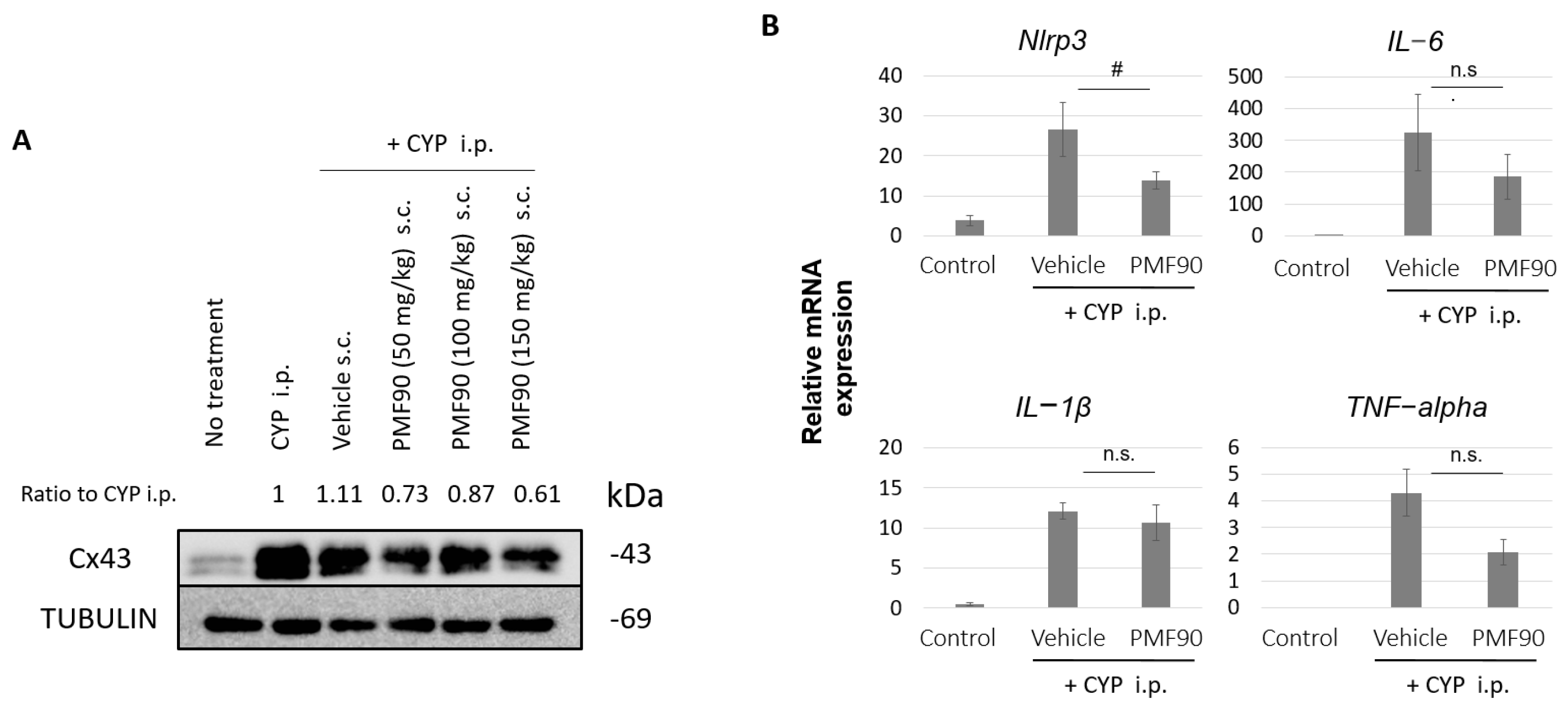
2.3. Prophylactic PMF90 Administration Attenuated the Upregulation of Cx43 Expression and Activation of Inflammation-Related Genes in the Bladder Mucosa of CYP-Treated Mice
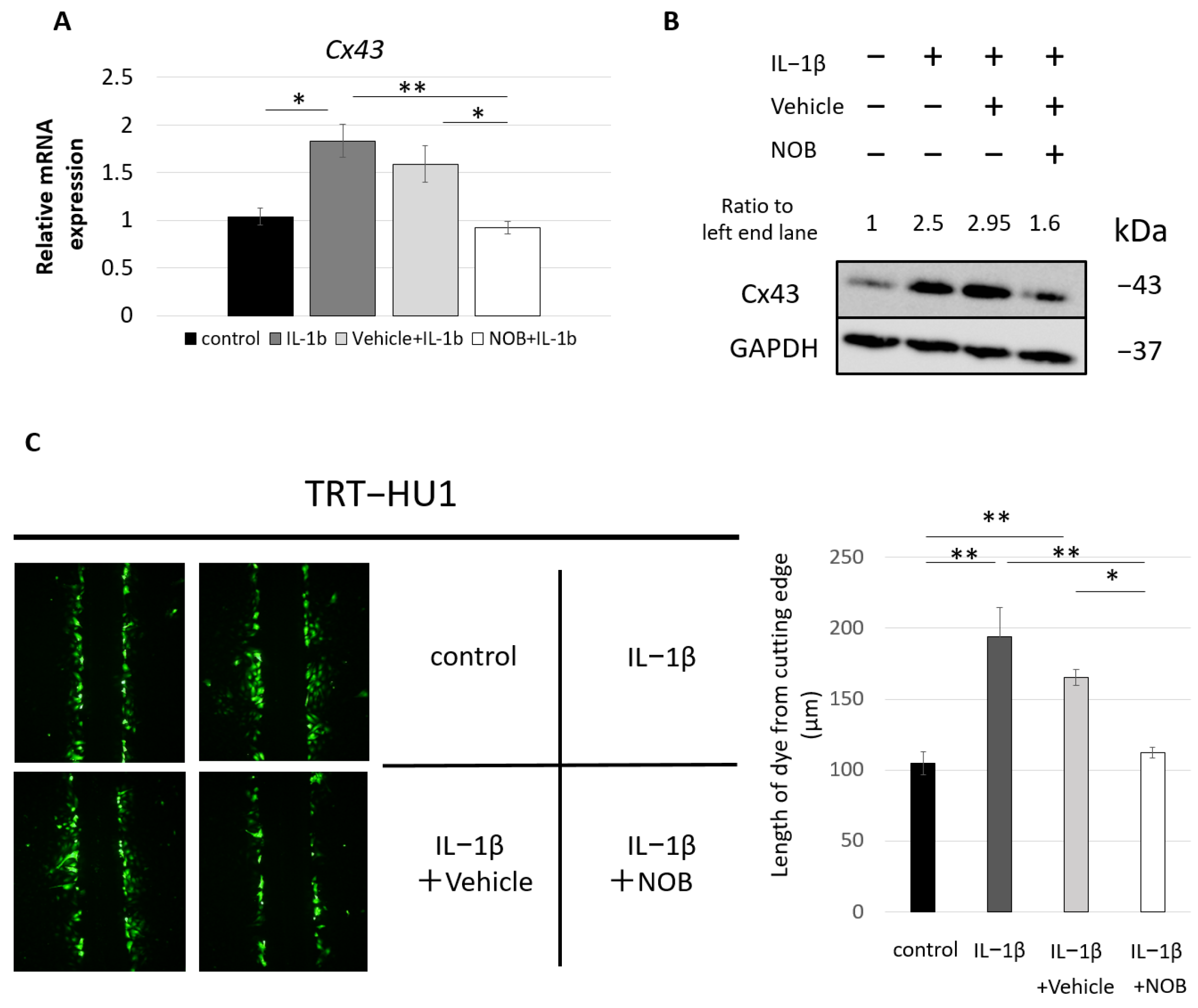
2.4. In Vitro Treatment with NOB Prevented IL-1β-Induced Upregulation of Cx43 Expression in Urothelial Cell Evaluation of Mechanosensitive Urothelial ATP Release in uCx43KO Mice
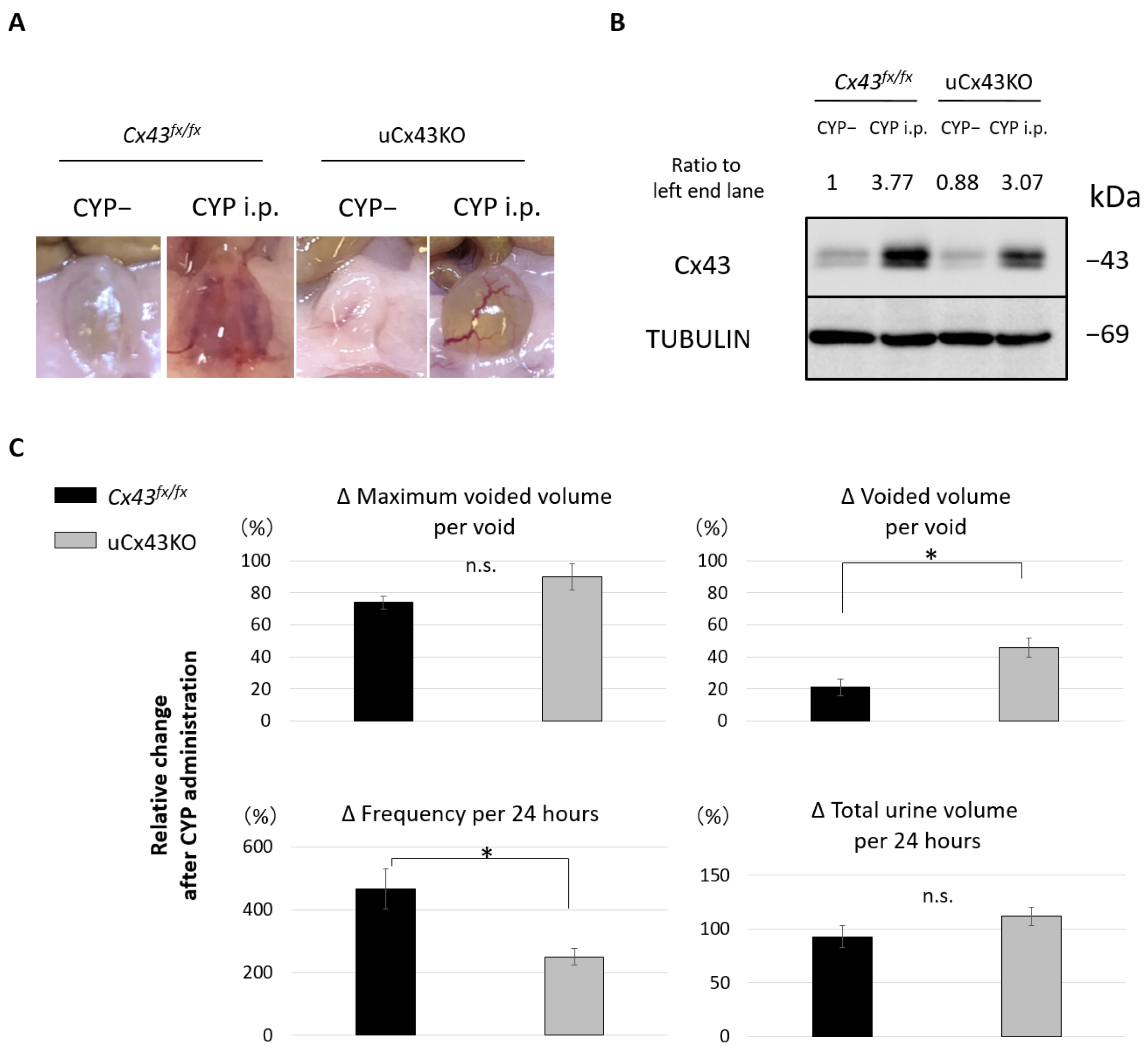
2.5. Effect of CYP Administration on Urothelium-Specific Cx43 KO Mice (uCx43KO Mice)
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Model of CYP-Induced Cystitis
4.3. Treatment with PMF90
4.4. Micturition Analysis
4.5. Tissue Harvesting
4.6. Hematoxylin and Eosin Staining of Mouse Bladder
4.7. Urothelial Cells and Cell Culture
4.8. IL-1 β Stimulation and Pre-Treatment of Urothelial Cells with Nobiletin
4.9. Scrape-Loading and Dye-Transfer Assay
4.10. Real-Time Quantitative PCR (qPCR)
4.11. Immunoblotting
4.12. Drugs/Reagents
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di, X.P.; Luo, D.Y.; Jin, X.; Zhao, W.Y.; Li, H.; Wang, K.J. Efficacy and safety comparison of pharmacotherapies for interstitial cystitis and bladder pain syndrome: A systematic review and Bayesian network meta-analysis. Int. Urogynecol. J. 2021, 32, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, N.; Homma, Y.; Tomoe, H.; Otsuka, A.; Kitta, T.; Masumori, N.; Akiyama, Y.; Niimi, A.; Mitsui, T.; Nanri, M.; et al. Efficacy and safety of intravesical instillation of KRP-116D (50% dimethyl sulfoxide solution) for interstitial cystitis/bladder pain syndrome in Japanese patients: A multicenter, randomized, double-blind, placebo-controlled, clinical study. Int. J. Urol. 2021, 28, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Homma, Y.; Akiyama, Y.; Tomoe, H.; Furuta, A.; Ueda, T.; Maeda, D.; Lin, A.T.; Kuo, H.C.; Lee, M.H.; Oh, S.J.; et al. Clinical guidelines for interstitial cystitis/bladder pain syndrome. Int. J. Urol. 2020, 27, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Hanno, P.M.; Erickson, D.; Moldwin, R.; Faraday, M.M. Diagnosis and treatment of interstitial cystitis/bladder pain syndrome: AUA guideline amendment. J. Urol. 2015, 193, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Hillelsohn, J.H.; Rais-Bahrami, S.; Friedlander, J.I.; Okhunov, Z.; Kashan, M.; Rosen, L.; Moldwin, R.M. Fulguration for Hunner ulcers: Long-term clinical outcomes. J. Urol. 2012, 188, 2238–2241. [Google Scholar] [CrossRef]
- Ryu, J.; Pak, S.; Song, M.; Chun, J.Y.; Hong, S.; Choo, M.S. Elimination of Hunner’s Ulcers by Fulguration in Patients With Interstitial Cystitis: Is It Effective and Long Lasting? Korean J. Urol. 2013, 54, 767–771. [Google Scholar] [CrossRef]
- Kajiwara, M.; Inoue, S.; Kobayashi, K.; Ohara, S.; Teishima, J.; Matsubara, A. Therapeutic efficacy of narrow band imaging-assisted transurethral electrocoagulation for ulcer-type interstitial cystitis/painful bladder syndrome. Int. J. Urol. 2014, 21 (Suppl. 1), 57–60. [Google Scholar] [CrossRef]
- Chennamsetty, A.; Khourdaji, I.; Goike, J.; Killinger, K.A.; Girdler, B.; Peters, K.M. Electrosurgical management of Hunner ulcers in a referral center’s interstitial cystitis population. Urology 2015, 85, 74–78. [Google Scholar] [CrossRef]
- Glemain, P.; Rivière, C.; Lenormand, L.; Karam, G.; Bouchot, O.; Buzelin, J.M. Prolonged hydrodistention of the bladder for symptomatic treatment of interstitial cystitis: Efficacy at 6 months and 1 year. Eur. Urol. 2002, 41, 79–84. [Google Scholar] [CrossRef]
- El-Hefnawy, A.S.; Makharita, M.Y.; Abed, A.; Amr, Y.M.; Salah El-Badry, M.; Shaaban, A.A. Anesthetic Bladder Hydrodistention Is Superior to Superior Hypogastric Plexus Neurolysis in Treatment of Interstitial Cystitis-bladder Pain Syndrome: A Prospective Randomized Trial. Urology 2015, 85, 1039–1044. [Google Scholar] [CrossRef]
- Niimi, A.; Nomiya, A.; Yamada, Y.; Suzuki, M.; Fujimura, T.; Fukuhara, H.; Kume, H.; Igawa, Y.; Homma, Y. Hydrodistension with or without fulguration of hunner lesions for interstitial cystitis: Long-term outcomes and prognostic predictors. Neurourol. Urodyn. 2016, 35, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Hoke, T.P.; Goldstein, H.; Saks, E.K.; Vakili, B. Hydrodistention of the bladder for the treatment of bladder pain syndrome/interstitial cystitis (BPS/IC). Neurourol. Urodyn. 2017, 36, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.S.; Santiago-Lastra, Y.; Qin, Y.; Stoffel, J.T.; Clemens, J.Q.; Cameron, A.P. The effects of cystoscopy and hydrodistention on symptoms and bladder capacity in interstitial cystitis/bladder pain syndrome. Neurourol. Urodyn. 2018, 37, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Abelleyra Lastoria, D.A.; Raison, N.; Aydin, A.; Khan, S.; Dasgupta, P.; Ahmed, K. Comparing surgical interventions for interstitial cystitis: A systematic review. Low Urin. Tract. Symptoms 2022. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Jhang, J.F.; Lee, Y.K.; Kuo, H.C. Low-Energy Shock Wave Plus Intravesical Instillation of Botulinum Toxin A for Interstitial Cystitis/Bladder Pain Syndrome: Pathophysiology and Preliminary Result of a Novel Minimally Invasive Treatment. Biomedicines 2022, 10, 396. [Google Scholar] [CrossRef]
- Evans, M.; Judy, W.V.; Wilson, D.; Rumberger, J.A.; Guthrie, N. Randomized, double-blind, placebo-controlled, clinical study on the effect of Diabetinol((R)) on glycemic control of subjects with impaired fasting glucose. Diabetes Metab. Syndr. Obes. 2015, 8, 275–286. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.T.; Li, H.B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients 2020, 12, 907. [Google Scholar] [CrossRef]
- Jang, S.E.; Ryu, K.R.; Park, S.H.; Chung, S.; Teruya, Y.; Han, M.J.; Woo, J.T.; Kim, D.H. Nobiletin and tangeretin ameliorate scratching behavior in mice by inhibiting the action of histamine and the activation of NF-kappaB, AP-1 and p38. Int. Immunopharmacol. 2013, 17, 502–507. [Google Scholar] [CrossRef]
- Lee, Y.S.; Cha, B.Y.; Saito, K.; Yamakawa, H.; Choi, S.S.; Yamaguchi, K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Nobiletin improves hyperglycemia and insulin resistance in obese diabetic ob/ob mice. Biochem. Pharmacol. 2010, 79, 1674–1683. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Ohizumi, Y. Beneficial Effects of Citrus-Derived Polymethoxylated Flavones for Central Nervous System Disorders. Nutrients 2021, 13, 145. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef] [PubMed]
- Boucher, M.; Meen, M.; Codron, J.P.; Coudore, F.; Kemeny, J.L.; Eschalier, A. Cyclophosphamide-induced cystitis in freely-moving conscious rats: Behavioral approach to a new model of visceral pain. J. Urol. 2000, 164, 203–208. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Yoshimura, N.; Huang, C.C.; Chiang, P.H.; Chancellor, M.B. Intravesical botulinum toxin a administration produces analgesia against acetic acid induced bladder pain responses in rats. J. Urol. 2004, 172, 1529–1532. [Google Scholar] [CrossRef] [PubMed]
- Boudes, M.; Uvin, P.; Kerselaers, S.; Vennekens, R.; Voets, T.; De Ridder, D. Functional characterization of a chronic cyclophosphamide-induced overactive bladder model in mice. Neurourol. Urodyn. 2011, 30, 1659–1665. [Google Scholar] [CrossRef]
- Auge, C.; Chene, G.; Dubourdeau, M.; Desoubzdanne, D.; Corman, B.; Palea, S.; Lluel, P.; Vergnolle, N.; Coelho, A.M. Relevance of the cyclophosphamide-induced cystitis model for pharmacological studies targeting inflammation and pain of the bladder. Eur. J. Pharmacol. 2013, 707, 32–40. [Google Scholar] [CrossRef]
- Homan, T.; Tsuzuki, T.; Dogishi, K.; Shirakawa, H.; Oyama, T.; Nakagawa, T.; Kaneko, S. Novel mouse model of chronic inflammatory and overactive bladder by a single intravesical injection of hydrogen peroxide. J. Pharmacol. Sci. 2013, 121, 327–337. [Google Scholar] [CrossRef]
- Birder, L.; Andersson, K.E. Animal Modelling of Interstitial Cystitis/Bladder Pain Syndrome. Int. Neurourol. J. 2018, 22, S3–s9. [Google Scholar] [CrossRef]
- Okinami, T.; Imamura, M.; Nishikawa, N.; Negoro, H.; Sugino, Y.; Yoshimura, K.; Kanematsu, A.; Hashitani, H.; Ogawa, O. Altered detrusor gap junction communications induce storage symptoms in bladder inflammation: A mouse cyclophosphamide-induced model of cystitis. PLoS ONE 2014, 9, e104216. [Google Scholar] [CrossRef]
- Cajas, Y.N.; Cañón-Beltrán, K.; Ladrón de Guevara, M.; Millán de la Blanca, M.G.; Ramos-Ibeas, P.; Gutiérrez-Adán, A.; Rizos, D.; González, E.M. Antioxidant Nobiletin Enhances Oocyte Maturation and Subsequent Embryo Development and Quality. Int. J. Mol. Sci. 2020, 21, 5340. [Google Scholar] [CrossRef]
- Kono, J.; Ueda, M.; Sengiku, A.; Suadicani, S.O.; Ogawa, O.; Negoro, H. Urothelium-Specific Deletion of Connexin 43 in the Mouse Urinary Bladder Alters Distension-Induced ATP Release and Voiding Behavior. Int. J. Mol. Sci. 2021, 22, 1594. [Google Scholar] [CrossRef]
- Negoro, H.; Kanematsu, A.; Doi, M.; Suadicani, S.O.; Matsuo, M.; Imamura, M.; Okinami, T.; Nishikawa, N.; Oura, T.; Matsui, S.; et al. Involvement of urinary bladder Connexin 43 and the circadian clock in coordination of diurnal micturition rhythm. Nat. Commun. 2012, 3, 809. [Google Scholar] [CrossRef] [PubMed]
- Sengiku, A.; Ueda, M.; Kono, J.; Sano, T.; Nishikawa, N.; Kunisue, S.; Tsujihana, K.; Liou, L.S.; Kanematsu, A.; Shimba, S.; et al. Circadian coordination of ATP release in the urothelium via connexin 43 hemichannels. Sci. Rep. 2018, 8, 1996. [Google Scholar] [CrossRef] [PubMed]
- Birder, L.A.; Kullmann, F.A. Role of neurogenic inflammation in local communication in the visceral mucosa. Semin. Immunopathol. 2018, 40, 261–279. [Google Scholar] [CrossRef]
- Birder, L.A. Urinary bladder, cystitis and nerve/urothelial interactions. Auton. Neurosci. 2014, 182, 89–94. [Google Scholar] [CrossRef]
- Negoro, H.; Urban-Maldonado, M.; Liou, L.S.; Spray, D.C.; Thi, M.M.; Suadicani, S.O. Pannexin 1 channels play essential roles in urothelial mechanotransduction and intercellular signaling. PLoS ONE 2014, 9, e106269. [Google Scholar] [CrossRef]
- Li, W.; Wang, X.; Niu, X.; Zhang, H.; He, Z.; Wang, Y.; Zhi, W.; Liu, F. Protective Effects of Nobiletin Against Endotoxic Shock in Mice Through Inhibiting TNF-α, IL-6, and HMGB1 and Regulating NF-κB Pathway. Inflammation 2016, 39, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wu, D.; Huang, L.; Jiang, C.; Pan, T.; Kang, X.; Pan, J. Nobiletin Inhibits IL-1β-Induced Inflammation in Chondrocytes via Suppression of NF-κB Signaling and Attenuates Osteoarthritis in Mice. Front. Pharmacol. 2019, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guo, Y.; Qiao, Y.; Zhang, J.; Jiang, P. Nobiletin Ameliorates NLRP3 Inflammasome-Mediated Inflammation Through Promoting Autophagy via the AMPK Pathway. Mol. Neurobiol. 2020, 57, 5056–5068. [Google Scholar] [CrossRef]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Inflammasome activation and regulation: Toward a better understanding of complex mechanisms. Cell Discov. 2020, 6, 36. [Google Scholar] [CrossRef]
- Chen, L.; Wang, H.; Ge, S.; Tai, S. IL-6/STAT3 pathway is involved in the regulation of autophagy in chronic non-bacterial prostatitis cells, and may be affected by the NLRP3 inflammasome. Ultrastruct. Pathol. 2021, 45, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Su, Y.; Chen, X.; Chen, Y.; Luo, P.; Lin, F.; Zhang, J. Rapamycin-induced autophagy attenuates hormone-imbalance-induced chronic non-bacterial prostatitis in rats via the inhibition of NLRP3 inflammasome-mediated inflammation. Mol. Med. Rep. 2019, 19, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M., Jr.; Hill, H.M.; Wood, C.M.; Edmondson, A.T.; Dumas, A.; Foo, W.C.; Oelsen, J.M.; Rac, G.; Purves, J.T. The NLRP3 Inflammasome Mediates Inflammation Produced by Bladder Outlet Obstruction. J. Urol. 2016, 195, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M., Jr.; Hirshman, N.A.; Inouye, B.M.; Jin, H.; Stanton, E.W.; Yun, C.E.; Davis, L.G.; Routh, J.C.; Purves, J.T. NLRP3 Promotes Diabetic Bladder Dysfunction and Changes in Symptom-Specific Bladder Innervation. Diabetes 2019, 68, 430–440. [Google Scholar] [CrossRef]
- Hughes, F.M., Jr.; Sexton, S.J.; Jin, H.; Govada, V.; Purves, J.T. Bladder fibrosis during outlet obstruction is triggered through the NLRP3 inflammasome and the production of IL-1β. Am. J. Physiol. Renal. Physiol. 2017, 313, F603–F610. [Google Scholar] [CrossRef]
- Lütolf, R.; Hughes, F.M., Jr.; Inouye, B.M.; Jin, H.; McMains, J.C.; Pak, E.S.; Hannan, J.L.; Purves, J.T. NLRP3/IL-1β mediates denervation during bladder outlet obstruction in rats. Neurourol. Urodyn. 2018, 37, 952–959. [Google Scholar] [CrossRef]
- Hughes, F.M., Jr.; Vivar, N.P.; Kennis, J.G.; Pratt-Thomas, J.D.; Lowe, D.W.; Shaner, B.E.; Nietert, P.J.; Spruill, L.S.; Purves, J.T. Inflammasomes are important mediators of cyclophosphamide-induced bladder inflammation. Am. J. Physiol. Renal. Physiol. 2014, 306, F299–F308. [Google Scholar] [CrossRef]
- Chen, L.; He, P.L.; Yang, J.; Yang, Y.F.; Wang, K.; Amend, B.; Stenzl, A.; Zhang, Y.M.; Wang, Z.L.; Xing, S.S.; et al. NLRP3/IL1beta inflammasome associated with the aging bladder triggers bladder dysfunction in female rats. Mol. Med. Rep. 2019, 19, 2960–2968. [Google Scholar] [CrossRef]
- Verma, V.; Gupta, S.; Kumar, P.; Yadav, S.; Dhanda, R.S.; Gaind, R.; Arora, R.; Frimodt-Moller, N.; Yadav, M. Involvement of NLRP3 and NLRC4 Inflammasome in Uropathogenic E. coli Mediated Urinary Tract Infections. Front. Microbiol. 2019, 10, 2020. [Google Scholar] [CrossRef]
- Shih, H.J.; Chang, C.Y.; Lai, C.H.; Huang, C.J. Therapeutic effect of modulating the NLRP3-regulated transforming growth factor-beta signaling pathway on interstitial cystitis/bladder pain syndrome. Biomed. Pharmacother. 2021, 138, 111522. [Google Scholar] [CrossRef]
- Korkmaz, A.; Topal, T.; Oter, S. Pathophysiological aspects of cyclophosphamide and ifosfamide induced hemorrhagic cystitis; implication of reactive oxygen and nitrogen species as well as PARP activation. Cell Biol. Toxicol. 2007, 23, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Malley, S.E.; Vizzard, M.A. Changes in urinary bladder cytokine mRNA and protein after cyclophosphamide-induced cystitis. Physiol. Genomics 2002, 9, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.; Laxton, V.; Srivastava, S.; Chan, Y.W.; Tse, G. The role of gap junctions in inflammatory and neoplastic disorders (Review). Int. J. Mol. Med. 2017, 39, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Cliff, C.L.; Williams, B.M.; Chadjichristos, C.E.; Mouritzen, U.; Squires, P.E.; Hills, C.E. Connexin 43: A Target for the Treatment of Inflammation in Secondary Complications of the Kidney and Eye in Diabetes. Int. J. Mol. Sci. 2022, 23, 600. [Google Scholar] [CrossRef]
- Swartzendruber, J.A.; Nicholson, B.J.; Murthy, A.K. The Role of Connexin 43 in Lung Disease. Life 2020, 10, 363. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, J.; Gao, K.; Chi, Y.; Mitsui, T.; Ihara, T.; Sawada, N.; Kamiyama, M.; Fan, J.; Takeda, M. AMPK Suppresses Connexin 43 Expression in the Bladder and Ameliorates Voiding Dysfunction in Cyclophosphamide-induced Mouse Cystitis. Sci. Rep. 2016, 6, 19708. [Google Scholar] [CrossRef]
- Willebrords, J.; Crespo Yanguas, S.; Maes, M.; Decrock, E.; Wang, N.; Leybaert, L.; Kwak, B.R.; Green, C.R.; Cogliati, B.; Vinken, M. Connexins and their channels in inflammation. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 413–439. [Google Scholar] [CrossRef]
- Peng, B.; Xu, C.; Wang, S.; Zhang, Y.; Li, W. The Role of Connexin Hemichannels in Inflammatory Diseases. Biology 2022, 11, 237. [Google Scholar] [CrossRef]
- Final report on the safety assessment of Glycyrrhetinic Acid, Potassium Glycyrrhetinate, Disodium Succinoyl Glycyrrhetinate, Glyceryl Glycyrrhetinate, Glycyrrhetinyl Stearate, Stearyl Glycyrrhetinate, Glycyrrhizic Acid, Ammonium Glycyrrhizate, Dipotassium Glycyrrhizate, Disodium Glycyrrhizate, Trisodium Glycyrrhizate, Methyl Glycyrrhizate, and Potassium Glycyrrhizinate. Int. J. Toxicol. 2007, 26 (Suppl. 2), 79–112. [CrossRef]
- Deutch, M.R.; Grimm, D.; Wehland, M.; Infanger, M.; Krüger, M. Bioactive Candy: Effects of Licorice on the Cardiovascular System. Foods 2019, 8, 495. [Google Scholar] [CrossRef]
- Roza, J.M.; Xian-Liu, Z.; Guthrie, N. Effect of citrus flavonoids and tocotrienols on serum cholesterol levels in hypercholesterolemic subjects. Altern. Ther. Health Med. 2007, 13, 44–48. [Google Scholar] [PubMed]
- Yamada, S.; Shirai, M.; Ono, K.; Teruya, T.; Yamano, A.; Tae Woo, J. Beneficial effects of a nobiletin-rich formulated supplement of Sikwasa (C. depressa) peel on cognitive function in elderly Japanese subjects; A multicenter, randomized, double-blind, placebo-controlled study. Food Sci. Nutr. 2021, 9, 6844–6853. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Heine, S.; Hahn, A. A combination of palm oil tocotrienols and citrus peel polymethoxylated flavones does not influence elevated LDL cholesterol and high-sensitivity C-reactive protein levels. Eur. J. Clin. Nutr. 2015, 69, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ji, M.; DiDonato, J.A.; Rackley, R.R.; Kuang, M.; Sadhukhan, P.C.; Mauney, J.R.; Keay, S.K.; Freeman, M.R.; Liou, L.S.; et al. An hTERT-immortalized human urothelial cell line that responds to anti-proliferative factor. In Vitro Cell Dev. Biol. Anim. 2011, 47, 2–9. [Google Scholar] [CrossRef] [PubMed][Green Version]
- el-Fouly, M.H.; Trosko, J.E.; Chang, C.C. Scrape-loading and dye transfer. A rapid and simple technique to study gap junctional intercellular communication. Exp. Cell Res. 1987, 168, 422–430. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kono, J.; Ueda, M.; Sengiku, A.; Suadicani, S.O.; Woo, J.T.; Kobayashi, T.; Ogawa, O.; Negoro, H. Flavonoid Nobiletin Attenuates Cyclophosphamide-Induced Cystitis in Mice through Mechanisms That Involve Inhibition of IL-1β Induced Connexin 43 Upregulation and Gap Junction Communication in Urothelial Cells. Int. J. Mol. Sci. 2022, 23, 5037. https://doi.org/10.3390/ijms23095037
Kono J, Ueda M, Sengiku A, Suadicani SO, Woo JT, Kobayashi T, Ogawa O, Negoro H. Flavonoid Nobiletin Attenuates Cyclophosphamide-Induced Cystitis in Mice through Mechanisms That Involve Inhibition of IL-1β Induced Connexin 43 Upregulation and Gap Junction Communication in Urothelial Cells. International Journal of Molecular Sciences. 2022; 23(9):5037. https://doi.org/10.3390/ijms23095037
Chicago/Turabian StyleKono, Jin, Masakatsu Ueda, Atsushi Sengiku, Sylvia O. Suadicani, Je Tae Woo, Takashi Kobayashi, Osamu Ogawa, and Hiromitsu Negoro. 2022. "Flavonoid Nobiletin Attenuates Cyclophosphamide-Induced Cystitis in Mice through Mechanisms That Involve Inhibition of IL-1β Induced Connexin 43 Upregulation and Gap Junction Communication in Urothelial Cells" International Journal of Molecular Sciences 23, no. 9: 5037. https://doi.org/10.3390/ijms23095037
APA StyleKono, J., Ueda, M., Sengiku, A., Suadicani, S. O., Woo, J. T., Kobayashi, T., Ogawa, O., & Negoro, H. (2022). Flavonoid Nobiletin Attenuates Cyclophosphamide-Induced Cystitis in Mice through Mechanisms That Involve Inhibition of IL-1β Induced Connexin 43 Upregulation and Gap Junction Communication in Urothelial Cells. International Journal of Molecular Sciences, 23(9), 5037. https://doi.org/10.3390/ijms23095037