
Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
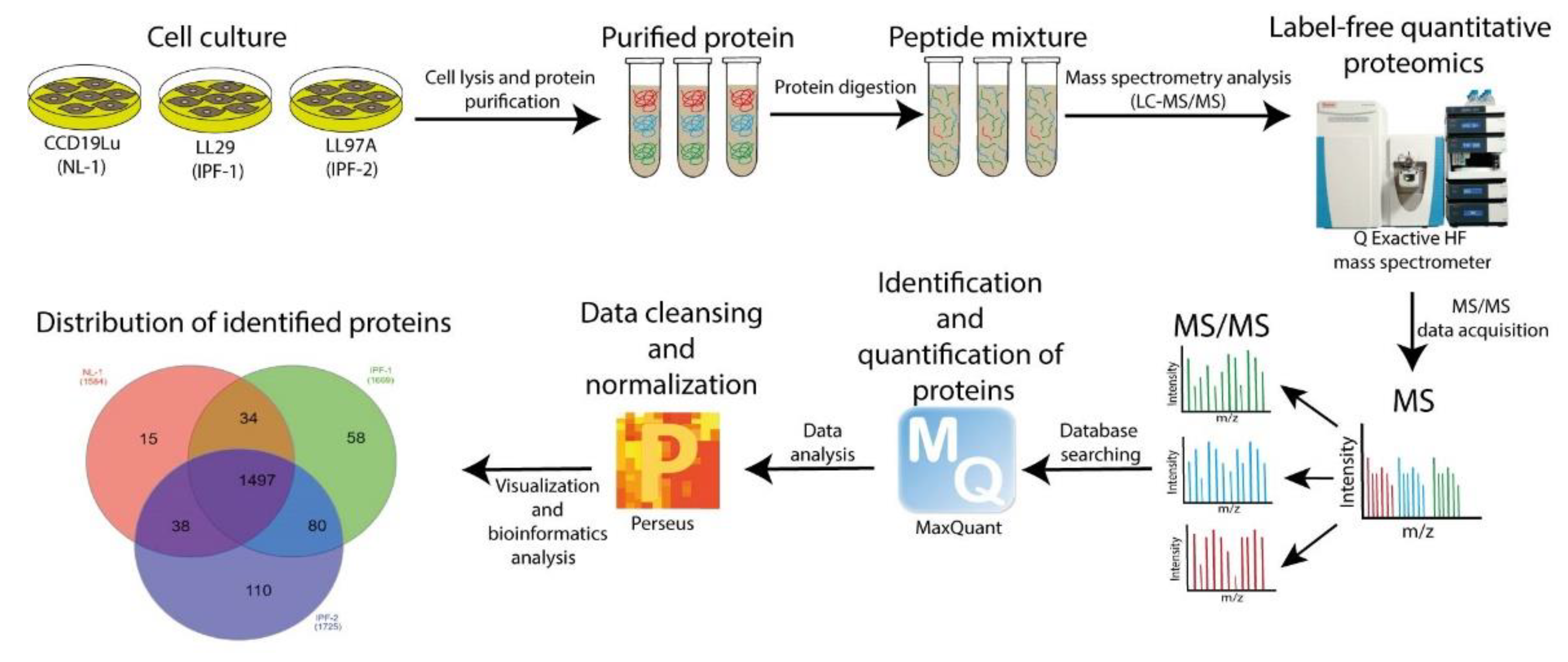
2.1. The Workflow of Proteomic Analysis
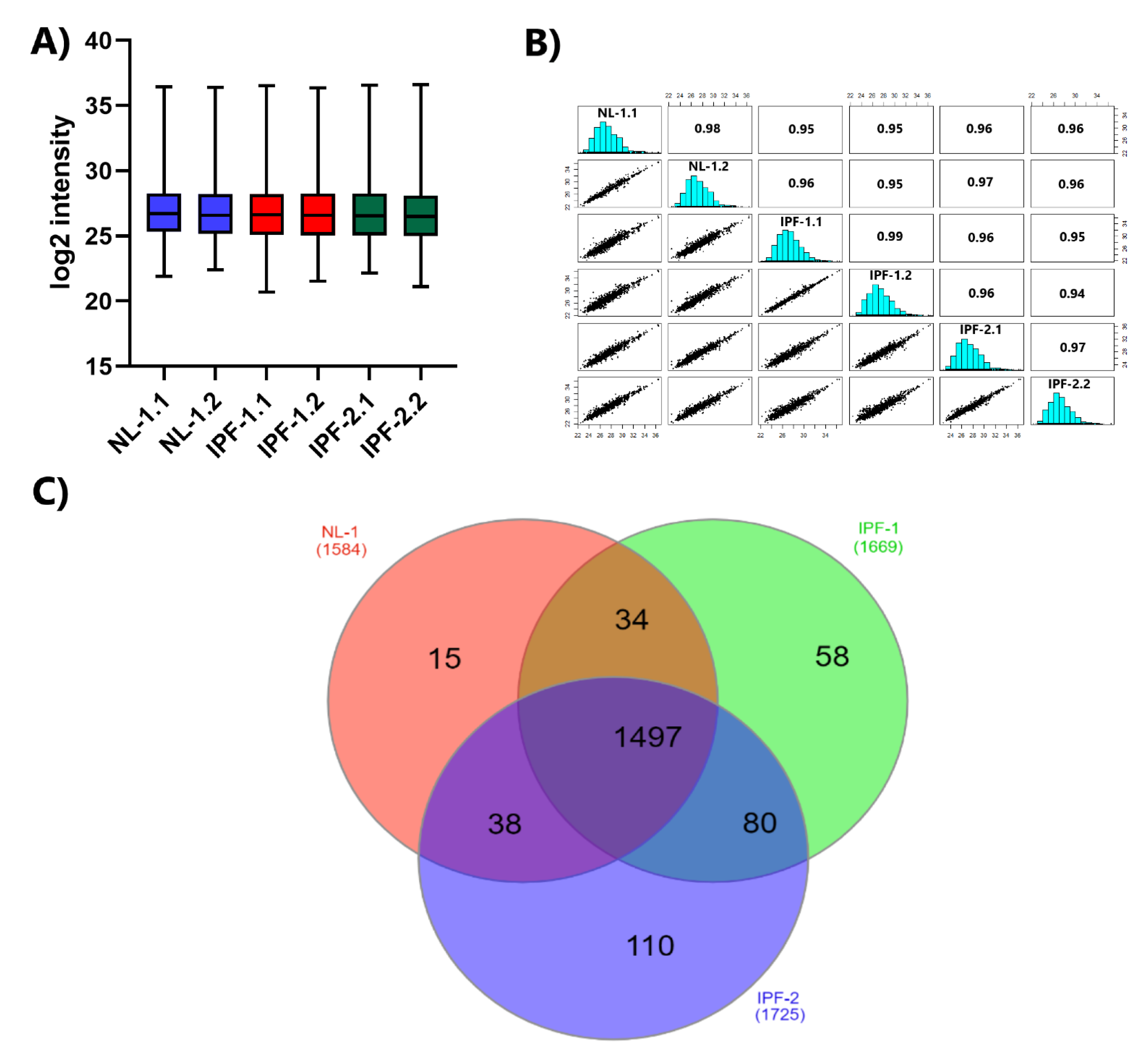
2.2. Reproducibility of LC-MS/MS Data
2.3. Distribution of Proteins Identified by Proteomic Analysis
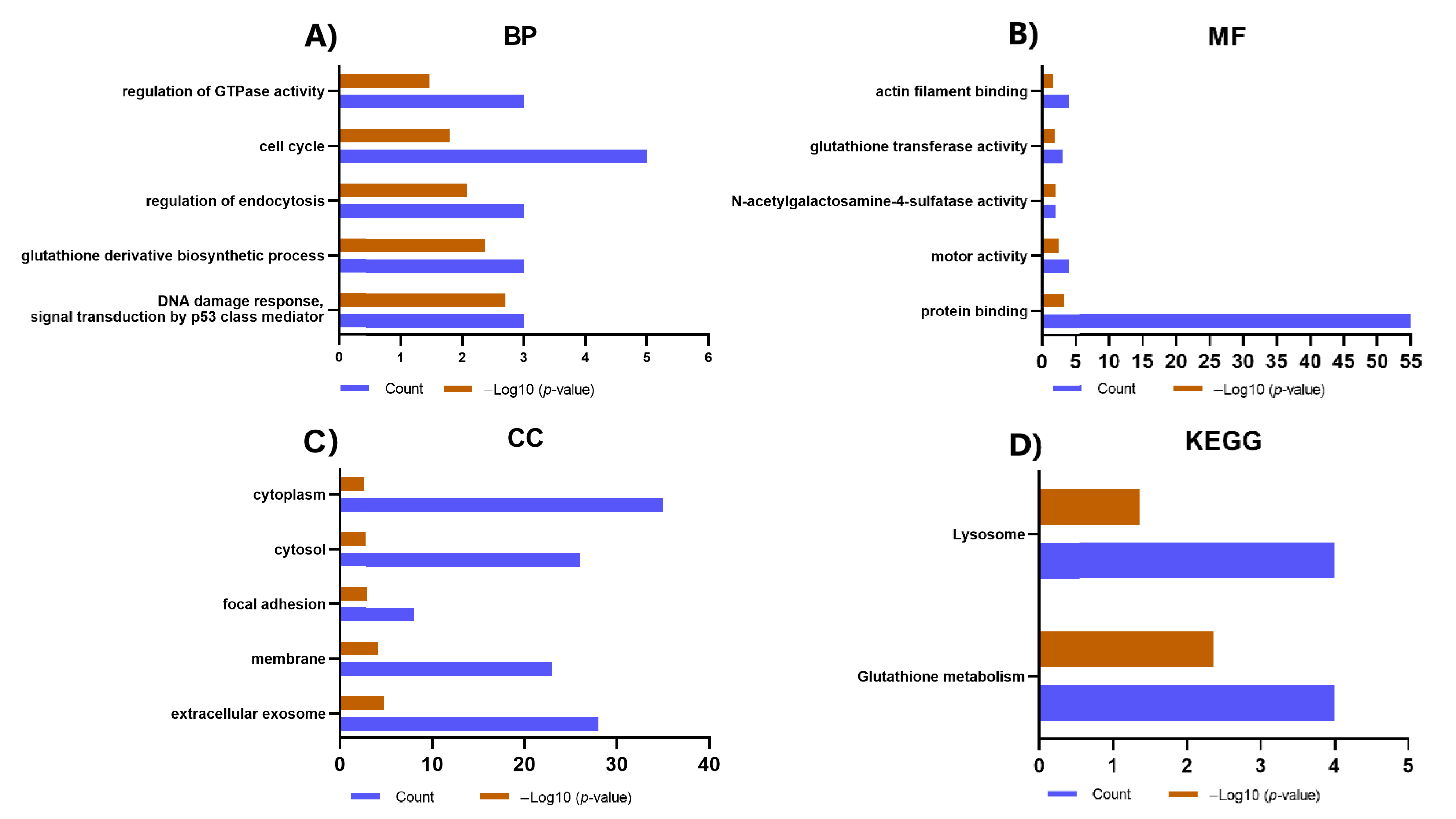
2.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of Proteins Identified Exclusively in the IPF-2 and IPF-1 Groups
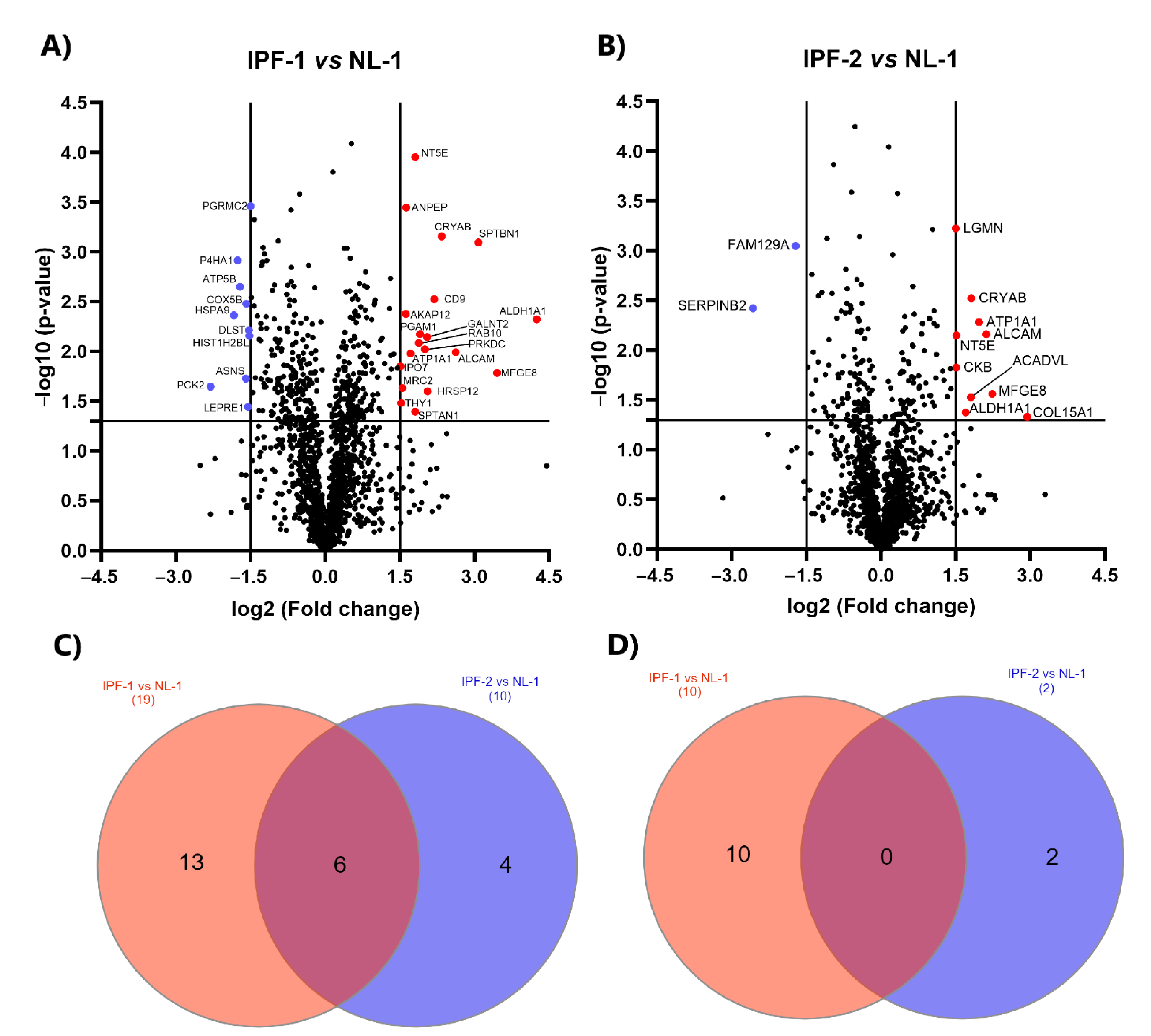
2.5. Identification of DEPs in IPF Cell Lines
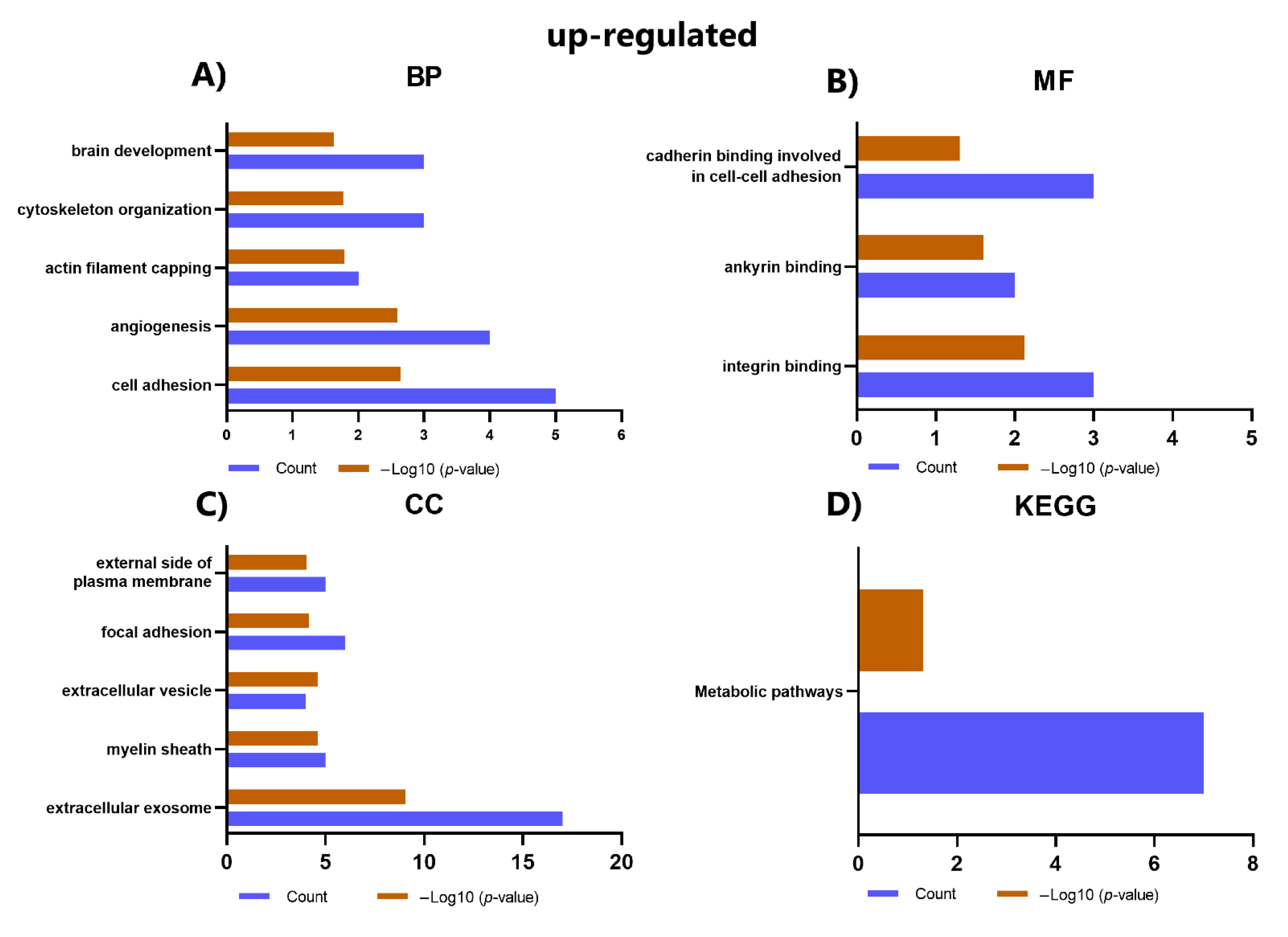
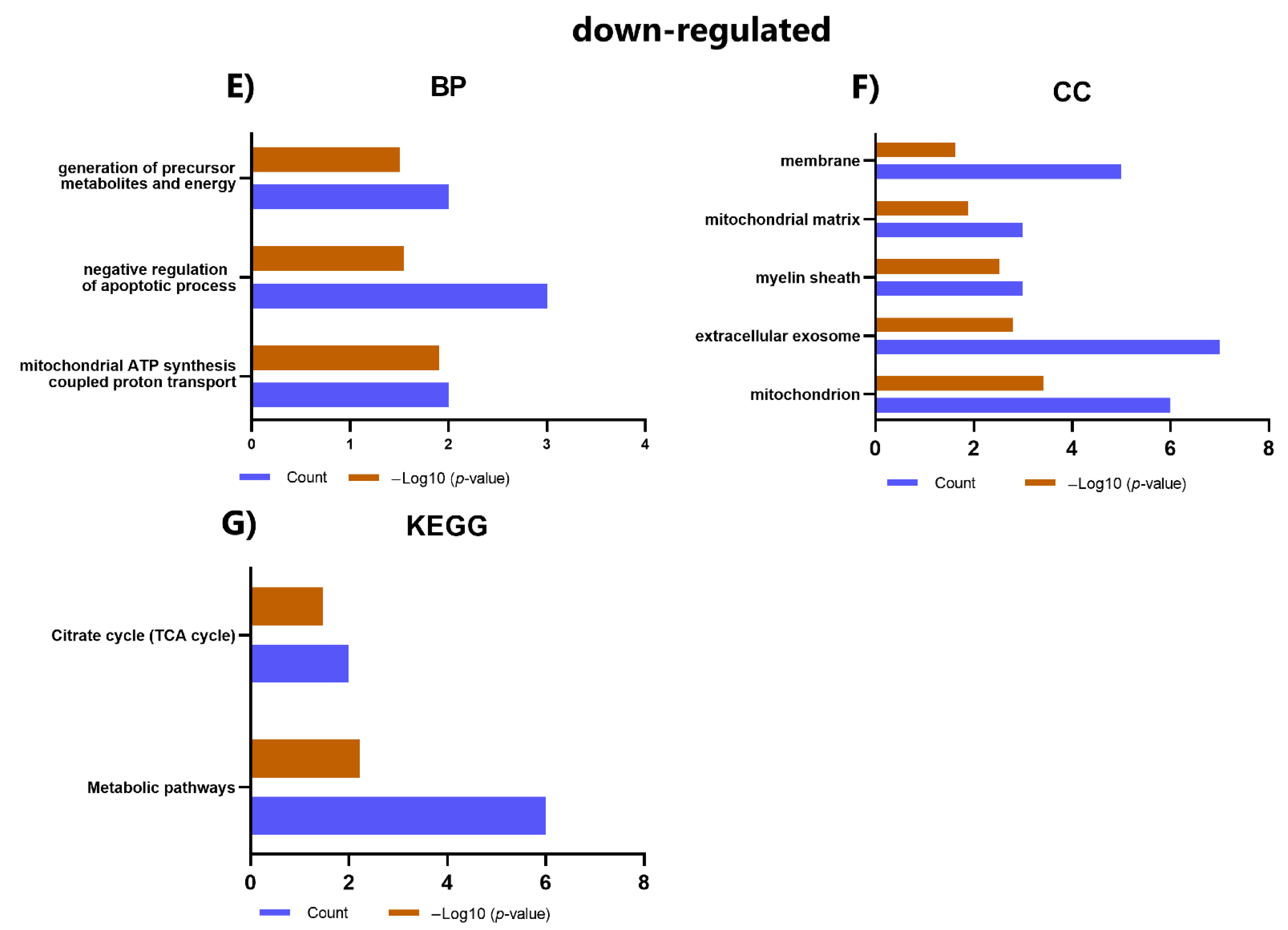
2.6. GO and KEGG Analysis of DEPs
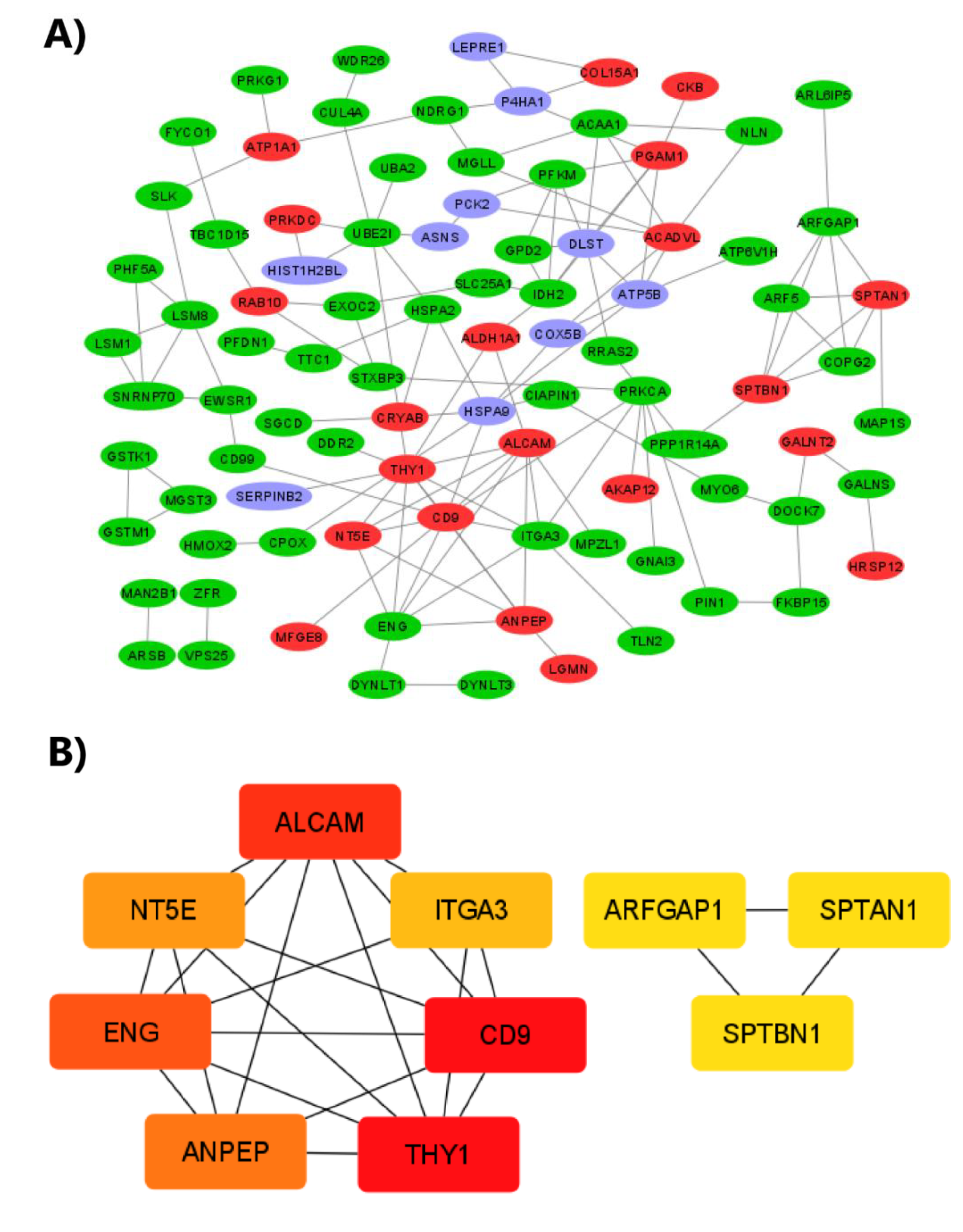
2.7. Protein–Protein Interaction (PPI) Network Construction and Analysis of Hub Proteins
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Protein Extraction and Quantification
4.3. Label-Free Quantitative Proteomic Analysis
4.3.1. Sample Preparation for Proteomic Analysis
4.3.2. LC-MS/MS Analysis
4.3.3. Proteomics Data Analysis
- Enzyme specificity was set to trypsin.
- Maximum missed cleavages were set to 2.
- Precursor ion mass tolerance was set to 10 ppm, and MS/MS tolerance was 0.6 Da.
- Protein modifications included carbamidomethylation (C) (fixed), oxidation (M) (variable).
- The false discovery rate (FDR) of peptides and proteins was 0.01.
4.4. Bioinformatics Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salvatore, M.; Ishikawa, G.; Padilla, M. Is It Idiopathic Pulmonary Fibrosis or Not? J. Am. Board Fam. Med. JABFM 2018, 31, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Enríquez, J.M.; Santos-Álvarez, J.C.; Ramírez-Hernández, A.A.; Reyes-Jiménez, E.; López-Martínez, A.; Pina-Canseco, S.; Aguilar-Ruiz, S.R.; Romero-Tlalolini, M.d.l.Á.; Castro-Sánchez, L.; Arellanes-Robledo, J.; et al. Proteomic Analysis Reveals Key Proteins in Extracellular Vesicles Cargo Associated with Idiopathic Pulmonary Fibrosis In Vitro. Biomedicines. 2021, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.; Swigris, J.; Pittrow, D.; Geier, S.; Klotsche, J.; Prasse, A.; Wirtz, H.; Koschel, D.; Andreas, S.; Claussen, M.; et al. Health related quality of life in patients with idiopathic pulmonary fibrosis in clinical practice: Insights-IPF registry. Respir. Res. 2017, 18, 139. [Google Scholar] [CrossRef]
- van Manen, M.J.; Geelhoed, J.J.; Tak, N.C.; Wijsenbeek, M.S. Optimizing quality of life in patients with idiopathic pulmonary fibrosis. Ther. Adv. Respir. Dis. 2017, 11, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Kathiriya, J.J.; Nakra, N.; Nixon, J.; Patel, P.S.; Vaghasiya, V.; Alhassani, A.; Tian, Z.; Allen-Gipson, D.; Davé, V. Galectin-1 inhibition attenuates profibrotic signaling in hypoxia-induced pulmonary fibrosis. Cell Death Discov. 2017, 3, 17010. [Google Scholar] [CrossRef]
- Plantier, L.; Renaud, H.; Respaud, R.; Marchand-Adam, S.; Crestani, B. Transcriptome of Cultured Lung Fibroblasts in Idiopathic Pulmonary Fibrosis: Meta-Analysis of Publically Available Microarray Datasets Reveals Repression of Inflammation and Immunity Pathways. Int. J. Mol. Sci. 2016, 17, 2091. [Google Scholar] [CrossRef]
- Chen, L.; Hou, J.; Fu, X.; Chen, X.; Wu, J.; Han, X. tPA promotes the proliferation of lung fibroblasts and activates the Wnt/β-catenin signaling pathway in idiopathic pulmonary fibrosis. Cell Cycle 2019, 18, 3137–3146. [Google Scholar] [CrossRef]
- Guillotin, D.; Taylor, A.R.; Platé, M.; Mercer, P.F.; Edwards, L.M.; Haggart, R.; Miele, G.; McAnulty, R.J.; Maher, T.M.; Hynds, R.E.; et al. Transcriptome analysis of IPF fibroblastic foci identifies key pathways involved in fibrogenesis. Thorax 2021, 76, 73–82. [Google Scholar] [CrossRef]
- Kimura, T.; Nojiri, T.; Hino, J.; Hosoda, H.; Miura, K.; Shintani, Y.; Inoue, M.; Zenitani, M.; Takabatake, H.; Miyazato, M.; et al. C-type natriuretic peptide ameliorates pulmonary fibrosis by acting on lung fibroblasts in mice. Respir. Res. 2016, 17, 19. [Google Scholar] [CrossRef]
- Sun, X.; Zhu, M.; Chen, X.; Jiang, X. MYH9 Inhibition Suppresses TGF-β1-Stimulated Lung Fibroblast-to-Myofibroblast Differentiation. Front. Pharmacol. 2020, 11, 573524. [Google Scholar] [CrossRef]
- Crestani, B.; Besnard, V.; Plantier, L.; Borensztajn, K.; Mailleux, A. Fibroblasts: The missing link between fibrotic lung diseases of different etiologies? Respir. Res. 2013, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Epstein Shochet, G.; Brook, E.; Israeli-Shani, L.; Edelstein, E.; Shitrit, D. Fibroblast paracrine TNF-α signaling elevates integrin A5 expression in idiopathic pulmonary fibrosis (IPF). Respir. Res. 2017, 18, 122. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.W.; Jo, H.-S.; Bae, S.; Seo, Y.; Song, P.; Song, M.; Yoon, J.H. Application of Proteomics in Cancer: Recent Trends and Approaches for Biomarkers Discovery. Front. Med. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Norman, K.C.; Moore, B.B.; Arnold, K.B.; O’Dwyer, D.N. Proteomics: Clinical and research applications in respiratory diseases. Respirology 2018, 23, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, H.; Gao, Y.; Liu, C.; Qiu, T.; Wu, H.; Cao, M.; Zhang, Y.; Ding, H.; Chen, J.; et al. Quantitative proteomic characterization of lung tissue in idiopathic pulmonary fibrosis. Clin. Proteom. 2019, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Niu, R.; Liu, Y.; Zhang, Y.; Zhang, Y.; Wang, H.; Wang, Y.; Wang, W.; Li, X. iTRAQ-Based Proteomics Reveals Novel Biomarkers for Idiopathic Pulmonary Fibrosis. PLoS ONE 2017, 12, e0170741. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.W.; Morrison, L.D.; Todd, J.L.; Snyder, L.D.; Thompson, J.W.; Soderblom, E.J.; Plonk, K.; Weinhold, K.J.; Townsend, R.; Minnich, A.; et al. Quantitative proteomics of bronchoalveolar lavage fluid in idiopathic pulmonary fibrosis. J. Proteome Res. 2015, 14, 1238–1249. [Google Scholar] [CrossRef]
- Saraswat, M.; Joenväärä, S.; Tohmola, T.; Sutinen, E.; Vartiainen, V.; Koli, K.; Myllärniemi, M.; Renkonen, R. Label-free plasma proteomics identifies haptoglobin-related protein as candidate marker of idiopathic pulmonary fibrosis and dysregulation of complement and oxidative pathways. Sci. Rep. 2020, 10, 7787. [Google Scholar] [CrossRef]
- Zhang, Y.; Xin, Q.; Wu, Z.; Wang, C.; Wang, Y.; Wu, Q.; Niu, R. Application of Isobaric Tags for Relative and Absolute Quantification (iTRAQ) Coupled with Two-Dimensional Liquid Chromatography/Tandem Mass Spectrometry in Quantitative Proteomic Analysis for Discovery of Serum Biomarkers for Idiopathic Pulmonary Fibrosis. Med. Sci. Monit. 2018, 24, 4146–4153. [Google Scholar] [CrossRef]
- Todd, J.L.; Neely, M.L.; Overton, R.; Durham, K.; Gulati, M.; Huang, H.; Roman, J.; Newby, L.K.; Flaherty, K.R.; Vinisko, R.; et al. Peripheral blood proteomic profiling of idiopathic pulmonary fibrosis biomarkers in the multicentre IPF-PRO Registry. Respir. Res. 2019, 20, 227. [Google Scholar] [CrossRef]
- Monaghan-Benson, E.; Wittchen, E.S.; Doerschuk, C.M.; Burridge, K. A Rnd3/p190RhoGAP pathway regulates RhoA activity in idiopathic pulmonary fibrosis fibroblasts. Mol. Biol. Cell 2018, 29, 2165–2175. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, T.; Zandbergen, M.; Wolterbeek, R.; Bruijn, J.A.; Baelde, H.J.; Scharpfenecker, M. Endoglin Promotes Myofibroblast Differentiation and Extracellular Matrix Production in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 7713. [Google Scholar] [CrossRef] [PubMed]
- Maring, J.A.; Trojanowska, M.; ten Dijke, P. Role of endoglin in fibrosis and scleroderma. Int. Rev. Cell Mol. Biol. 2012, 297, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.; Chrobak, I.; Bujor, A.; Hant, F.; Mummery, C.; Ten Dijke, P.; Trojanowska, M. Endoglin promotes TGF-β/Smad1 signaling in scleroderma fibroblasts. J. Cell. Physiol. 2011, 226, 3340–3348. [Google Scholar] [CrossRef]
- Kapur, N.K.; Qiao, X.; Paruchuri, V.; Mackey, E.E.; Daly, G.H.; Ughreja, K.; Morine, K.J.; Levine, J.; Aronovitz, M.J.; Hill, N.S.; et al. Reducing endoglin activity limits calcineurin and TRPC-6 expression and improves survival in a mouse model of right ventricular pressure overload. J. Am. Heart Assoc. 2014, 3, e000965. [Google Scholar] [CrossRef]
- Kasprzak, A.; Adamek, A. Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy. Int. J. Mol. Sci. 2018, 19, 3887. [Google Scholar] [CrossRef]
- Chen, K.; Mehta, J.L.; Li, D.; Joseph, L.; Joseph, J. Transforming Growth Factor β Receptor Endoglin Is Expressed in Cardiac Fibroblasts and Modulates Profibrogenic Actions of Angiotensin II. Circ. Res. 2004, 95, 1167–1173. [Google Scholar] [CrossRef]
- Oujo, B.; Muñoz-Félix, J.M.; Arévalo, M.; Núñez-Gómez, E.; Pérez-Roque, L.; Pericacho, M.; González-Núñez, M.; Langa, C.; Martínez-Salgado, C.; Perez-Barriocanal, F.; et al. L-Endoglin overexpression increases renal fibrosis after unilateral ureteral obstruction. PLoS ONE 2014, 9, e110365. [Google Scholar] [CrossRef]
- Liu, X.; Wong, S.S.; Taype, C.A.; Kim, J.; Shentu, T.-P.; Espinoza, C.R.; Finley, J.C.; Bradley, J.E.; Head, B.P.; Patel, H.H.; et al. Thy-1 interaction with Fas in lipid rafts regulates fibroblast apoptosis and lung injury resolution. Lab. Investig. J. Tech. methods Pathol. 2017, 97, 256–267. [Google Scholar] [CrossRef]
- Zhou, Y.; Hagood, J.S.; Murphy-Ullrich, J.E. Thy-1 expression regulates the ability of rat lung fibroblasts to activate transforming growth factor-beta in response to fibrogenic stimuli. Am. J. Pathol. 2004, 165, 659–669. [Google Scholar] [CrossRef]
- Schmidt, M.; Gutknecht, D.; Simon, J.C.; Schulz, J.-N.; Eckes, B.; Anderegg, U.; Saalbach, A. Controlling the Balance of Fibroblast Proliferation and Differentiation: Impact of Thy-1. J. Investig. Dermatol. 2015, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tang, R.-Z.; Ruan, J.; Zhu, X.-B.; Yang, Y. Up-regulation of THY1 attenuates interstitial pulmonary fibrosis and promotes lung fibroblast apoptosis during acute interstitial pneumonia by blockade of the WNT signaling pathway. Cell Cycle 2019, 18, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, Y.; Peng, X.; Yan, H.; Zhang, X.; Yin, L.; Yu, H. Transcription factor YY1 inhibits the expression of THY1 to promote interstitial pulmonary fibrosis by activating the HSF1/miR-214 axis. Aging 2020, 12, 8339–8351. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, G.; Hagood, J.S.; Sanders, Y.; Ramírez, R.; Becerril, C.; Segura, L.; Barrera, L.; Selman, M.; Pardo, A. Absence of Thy-1 results in TGF-β induced MMP-9 expression and confers a profibrotic phenotype to human lung fibroblasts. Lab. Investig. J. Tech. methods Pathol. 2011, 91, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.Y.; Breuer, R.; Wallach-Dayan, S.B. A Profibrotic Phenotype in Naïve and in Fibrotic Lung Myofibroblasts Is Governed by Modulations in Thy-1 Expression and Activation. Mediat. Inflamm. 2018, 2018, 4638437. [Google Scholar] [CrossRef] [PubMed]
- Hagood, J.S.; Prabhakaran, P.; Kumbla, P.; Salazar, L.; MacEwen, M.W.; Barker, T.H.; Ortiz, L.A.; Schoeb, T.; Siegal, G.P.; Alexander, C.B.; et al. Loss of fibroblast Thy-1 expression correlates with lung fibrogenesis. Am. J. Pathol. 2005, 167, 365–379. [Google Scholar] [CrossRef]
- Chen, S.; Sun, Y.; Jin, Z.; Jing, X. Functional and biochemical studies of CD9 in fibrosarcoma cell line. Mol. Cell. Biochem. 2011, 350, 89–99. [Google Scholar] [CrossRef]
- Tsujino, K.; Takeda, Y.; Arai, T.; Shintani, Y.; Inagaki, R.; Saiga, H.; Iwasaki, T.; Tetsumoto, S.; Jin, Y.; Ihara, S.; et al. Tetraspanin CD151 protects against pulmonary fibrosis by maintaining epithelial integrity. Am. J. Respir. Crit. Care Med. 2012, 186, 170–180. [Google Scholar] [CrossRef]
- Brosseau, C.; Colas, L.; Magnan, A.; Brouard, S. CD9 Tetraspanin: A New Pathway for the Regulation of Inflammation? Front. immunol. 2018, 9, 2316. [Google Scholar] [CrossRef]
- Jin, Y.; Tachibana, I.; Takeda, Y.; He, P.; Kang, S.; Suzuki, M.; Kuhara, H.; Tetsumoto, S.; Tsujino, K.; Minami, T.; et al. Statins decrease lung inflammation in mice by upregulating tetraspanin CD9 in macrophages. PLoS ONE 2013, 8, e73706. [Google Scholar] [CrossRef]
- Suzuki, M.; Tachibana, I.; Takeda, Y.; He, P.; Minami, S.; Iwasaki, T.; Kida, H.; Goya, S.; Kijima, T.; Yoshida, M.; et al. Tetraspanin CD9 Negatively Regulates Lipopolysaccharide-Induced Macrophage Activation and Lung Inflammation. J. Immunol. 2009, 182, 6485. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.P.; Itoh, M.N.; Takeda, Y.; Tsujino, K.; Kondo, Y.; Kumanogoh, A.; Mizuguchi, K. Integrative Analysis Reveals Common and Unique Roles of Tetraspanins in Fibrosis and Emphysema. Front. Genet. 2020, 11, 585998. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, C.; Tellez, A.; Jara, C.; Alarcón, S.; Torres, A.; Mendoza, P.; Podestá, L.; Flores, C.; Quezada, C.; Oyarzún, C.; et al. The TGF-β profibrotic cascade targets ecto-5′-nucleotidase gene in proximal tubule epithelial cells and is a traceable marker of progressive diabetic kidney disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165796. [Google Scholar] [CrossRef] [PubMed]
- Minor, M.; Alcedo, K.P.; Battaglia, R.A.; Snider, N.T. Cell type- and tissue-specific functions of ecto-5’-nucleotidase (CD73). Am. J. Physiol. Cell Physiol. 2019, 317, C1079–C1092. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Fernandez, P.; Wilder, T.; Yee, H.; Chiriboga, L.; Chan, E.S.L.; Cronstein, B.N. Ecto-5′-nucleotidase (CD73) -mediated extracellular adenosine production plays a critical role in hepatic fibrosis. FASEB J. 2008, 22, 2263–2272. [Google Scholar] [CrossRef]
- Wirsdörfer, F.; de Leve, S.; Cappuccini, F.; Eldh, T.; Meyer, A.V.; Gau, E.; Thompson, L.F.; Chen, N.-Y.; Karmouty-Quintana, H.; Fischer, U.; et al. Extracellular Adenosine Production by ecto-5’-Nucleotidase (CD73) Enhances Radiation-Induced Lung Fibrosis. Cancer Res. 2016, 76, 3045–3056. [Google Scholar] [CrossRef]
- Turiello, R.; Pinto, A.; Morello, S. CD73: A Promising Biomarker in Cancer Patients. Front. Pharmacol. 2020, 11, 609931. [Google Scholar] [CrossRef]
- Volmer, J.B.; Thompson, L.F.; Blackburn, M.R. Ecto-5’-nucleotidase (CD73)-mediated adenosine production is tissue protective in a model of bleomycin-induced lung injury. J. Immunol. 2006, 176, 4449–4458. [Google Scholar] [CrossRef]
- Colombo, E.A.; Spaccini, L.; Volpi, L.; Negri, G.; Cittaro, D.; Lazarevic, D.; Zirpoli, S.; Farolfi, A.; Gervasini, C.; Cubellis, M.V.; et al. Viable phenotype of ILNEB syndrome without nephrotic impairment in siblings heterozygous for unreported integrin alpha3 mutations. Orphanet J. Rare Dis. 2016, 11, 136. [Google Scholar] [CrossRef]
- Huang, Y.; Kong, Y.; Zhang, L.; He, T.; Zhou, X.; Yan, Y.; Zhang, L.; Zhou, D.; Lu, S.; Zhou, J.; et al. High Expression of ITGA3 Promotes Proliferation and Cell Cycle Progression and Indicates Poor Prognosis in Intrahepatic Cholangiocarcinoma. BioMed Res. Int. 2018, 2018, 2352139. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Bai, X.; Li, Y.; Ni, C.; Zhao, X.; Zhang, D. ITGA3 Is Associated With Immune Cell Infiltration and Serves as a Favorable Prognostic Biomarker for Breast Cancer. Front. Oncol. 2021, 11, 658547. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, N.; Margadant, C.; Kevelam, S.H.; Lilien, M.R.; Oosterveld, M.J.S.; Kreft, M.; van Eerde, A.M.; Pfundt, R.; Terhal, P.A.; van der Zwaag, B.; et al. Gain of glycosylation in integrin α3 causes lung disease and nephrotic syndrome. J. Clin. Investig. 2012, 122, 4375–4387. [Google Scholar] [CrossRef] [PubMed]
- Sreeja, J.S.; John, R.; Dharmapal, D.; Nellikka, R.K.; Sengupta, S. A Fresh Look at the Structure, Regulation, and Functions of Fodrin. Mol. Cell. Biol. 2020, 40, e00133-00120. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, A.; Brieger, A. The Role of Nonerythroid Spectrin αII in Cancer. J. Oncol. 2019, 2019, 7079604. [Google Scholar] [CrossRef] [PubMed]
- Derbala, M.H.; Guo, A.S.; Mohler, P.J.; Smith, S.A. The role of βII spectrin in cardiac health and disease. Life Sci. 2018, 192, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Yang, Y.; Sun, P.; Tian, Y.; Gao, F.; Wang, C.; Zong, T.; Li, M.; Zhang, Y.; Yu, T.; et al. βII spectrin (SPTBN1): Biological function and clinical potential in cancer and other diseases. Int. J. Biol. Sci. 2021, 17, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, K.; Ohuchida, K.; Sada, M.; Horioka, K.; Ulrich, C.D., III; Shindo, K.; Ohtsuka, T.; Takahata, S.; Mizumoto, K.; Oda, Y.; et al. CD166/ALCAM Expression Is Characteristic of Tumorigenicity and Invasive and Migratory Activities of Pancreatic Cancer Cells. PLoS ONE 2014, 9, e107247. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Cao, Q.; Yu, H.; Zheng, D.; Wang, Y.; Harris, D.C.H. Expression and Role of CD166 in the Chronic Kidney Disease. Iran J. Pediatr. 2015, 25, e543. [Google Scholar] [CrossRef]
- He, W.; Huang, L.; Li, M.; Yang, Y.; Chen, Z.; Shen, X. MiR-148b, MiR-152/ALCAM Axis Regulates the Proliferation and Invasion of Pituitary Adenomas Cells. Cell. Physiol. Biochem. 2017, 44, 792–803. [Google Scholar] [CrossRef]
- Dalerba, P.; Dylla, S.J.; Park, I.-K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef]
- Rosso, O.; Piazza, T.; Bongarzone, I.; Rossello, A.; Mezzanzanica, D.; Canevari, S.; Orengo, A.M.; Puppo, A.; Ferrini, S.; Fabbi, M. The ALCAM Shedding by the Metalloprotease ADAM17/TACE Is Involved in Motility of Ovarian Carcinoma Cells. Mol. Cancer Res. 2008, 5, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lin, H.; Qu, S.; Li, L.; Chen, K.; Yu, B.; Lin, G.; Wan, F.; Zhu, X. Downregulation of CD166 inhibits invasion, migration, and EMT in the radio-resistant human nasopharyngeal carcinoma cell line CNE-2R. Cancer Manag. Res. 2019, 11, 3593–3602. [Google Scholar] [CrossRef]
- Morgan, R.L.; Behbahani-Nejad, N.; Endres, J.; Amin, M.A.; Lepore, N.J.; Du, Y.; Urquhart, A.; Chung, K.C.; Fox, D.A. Localization, Shedding, Regulation and Function of Aminopeptidase N/CD13 on Fibroblast like Synoviocytes. PLoS ONE 2016, 11, e0162008. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.; Ghaffari, A.; Ghahary, A. Inhibitory effect of anti-aminopeptidase N/CD13 antibodies on fibroblast migration. Mol. Cell Biochem. 2010, 343, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.; Endres, J.; Behbahani-Nejad, N.; Phillips, K.; Ruth, J.H.; Friday, S.C.; Edhayan, G.; Lanigan, T.; Urquhart, A.; Chung, K.C.; et al. Expression and Function of Aminopeptidase N/CD13 Produced by Fibroblast-like Synoviocytes in Rheumatoid Arthritis: Role of CD13 in Chemotaxis of Cytokine-Activated T Cells Independent of Enzymatic Activity. Arthritis Rheumatol. 2015, 67, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Kadefors, M.; Rolandsson Enes, S.; Åhrman, E.; Michaliková, B.; Löfdahl, A.; Dellgren, G.; Scheding, S.; Westergren-Thorsson, G. CD105+CD90+CD13+ identifies a clonogenic subset of adventitial lung fibroblasts. Sci. Rep. 2021, 11, 24417. [Google Scholar] [CrossRef]
- Kong, S.L.; Chui, P.; Lim, B.; Salto-Tellez, M. Elucidating the molecular physiopathology of acute respiratory distress syndrome in severe acute respiratory syndrome patients. Virus Res. 2009, 145, 260–269. [Google Scholar] [CrossRef]
- Lu, C.; Amin, M.A.; Fox, D.A. CD13/Aminopeptidase N Is a Potential Therapeutic Target for Inflammatory Disorders. J. Immunol. 2020, 204, 3–11. [Google Scholar] [CrossRef]
- Khodadadi, E.; Maroufi, P.; Khodadadi, E.; Esposito, I.; Ganbarov, K.; Espsoito, S.; Yousefi, M.; Zeinalzadeh, E.; Kafil, H.S. Study of combining virtual screening and antiviral treatments of the SARS-CoV-2 (COVID-19). Microb. Pathog. 2020, 146, 104241. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Zlateva, K.; Van Ranst, M. Identification of six new polymorphisms in the human coronavirus 229E receptor gene (aminopeptidase N/CD13). Int. J. Infect. Dis. 2004, 8, 217–222. [Google Scholar] [CrossRef][Green Version]
- Gannon, J.; Fernandez-Rodriguez, J.; Alamri, H.; Feng, S.B.; Kalantari, F.; Negi, S.; Wong, A.H.Y.; Mazur, A.; Asp, L.; Fazel, A.; et al. ARFGAP1 is dynamically associated with lipid droplets in hepatocytes. PLoS ONE 2014, 9, e111309. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Jung, C.; Chandra, M.; Engel, A.; Kendall, A.K.; Jackson, L.P. The Glo3 GAP crystal structure supports the molecular niche model for ArfGAPs in COPI coats. Adv. Biol. Regul. 2021, 79, 100781. [Google Scholar] [CrossRef] [PubMed]
- Hsu, V.W. Role of ArfGAP1 in COPI vesicle biogenesis. Cell Logist 2011, 1, 55–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Meng, D.; Yang, Q.; Melick, C.H.; Park, B.C.; Hsieh, T.-S.; Curukovic, A.; Jeong, M.-H.; Zhang, J.; James, N.G.; Jewell, J.L. ArfGAP1 inhibits mTORC1 lysosomal localization and activation. EMBO J. 2021, 40, e106412. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Cox, J. Perseus: A Bioinformatics Platform for Integrative Analysis of Proteomics Data in Cancer Research. In Cancer Systems Biology; Humana Press: New York, NY, USA, 2018; Volume 1711, pp. 133–148. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl 4), S11. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Term | Count | p-Value | Proteins |
|---|---|---|---|---|
| BP | GO:0007155~cell adhesion | 5 | 6.23 × 105 | ALCAM, ITGA3, CD9, THY1, ENG |
| BP | GO:0043547~positive regulation of GTPase activity | 4 | 0.00273472 | THY1, SPTAN1, SPTBN1, ARFGAP1 |
| BP | GO:0006888~ER to Golgi vesicle-mediated transport | 3 | 0.0031085 | SPTAN1, SPTBN1, ARFGAP1 |
| BP | GO:0007010~cytoskeleton organization | 3 | 0.00314673 | THY1, SPTAN1, SPTBN1 |
| BP | GO:0051693~actin filament capping | 2 | 0.00694772 | SPTAN1, SPTBN1 |
| BP | GO:0017015~regulation of transforming growth factor-beta receptor signaling pathway | 2 | 0.01067099 | ITGA3, ENG |
| BP | GO:0030336~negative regulation of cell migration | 2 | 0.04979153 | THY1, ENG |
| MF | GO:0005178~integrin binding | 3 | 0.00134085 | ITGA3, CD9, THY1 |
| CC | GO:0009897~external side of plasma membrane | 6 | 2.52 × 108 | ALCAM, ITGA3, ANPEP, CD9, THY1, ENG |
| CC | GO:0005925~focal adhesion | 5 | 2.41 × 105 | ALCAM, ITGA3, CD9, THY1, ENG |
| CC | GO:0070062~extracellular exosome | 8 | 5.57 × 105 | NT5E, ALCAM, ITGA3, ANPEP, CD9, THY1, SPTAN1, SPTBN1 |
| CC | GO:0008091~spectrin | 2 | 0.00443689 | SPTAN1, SPTBN1 |
| CC | GO:1903561~extracellular vesicle | 2 | 0.02442875 | CD9, SPTAN1 |
| CC | GO:0005887~integral component of plasma membrane | 4 | 0.0275047 | ALCAM, ANPEP, CD9, THY1 |
| CC | GO:0009986~cell surface | 3 | 0.02767149 | NT5E, ITGA3, ENG |
| KEGG | hsa04640: Hematopoietic cell lineage | 3 | 0.00229492 | ITGA3, ANPEP, CD9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velázquez-Enríquez, J.M.; Ramírez-Hernández, A.A.; Navarro, L.M.S.; Reyes-Avendaño, I.; González-García, K.; Jiménez-Martínez, C.; Castro-Sánchez, L.; Sánchez-Chino, X.M.; Vásquez-Garzón, V.R.; Baltiérrez-Hoyos, R. Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines. Int. J. Mol. Sci. 2022, 23, 5032. https://doi.org/10.3390/ijms23095032
Velázquez-Enríquez JM, Ramírez-Hernández AA, Navarro LMS, Reyes-Avendaño I, González-García K, Jiménez-Martínez C, Castro-Sánchez L, Sánchez-Chino XM, Vásquez-Garzón VR, Baltiérrez-Hoyos R. Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines. International Journal of Molecular Sciences. 2022; 23(9):5032. https://doi.org/10.3390/ijms23095032
Chicago/Turabian StyleVelázquez-Enríquez, Juan Manuel, Alma Aurora Ramírez-Hernández, Luis Manuel Sánchez Navarro, Itayetzi Reyes-Avendaño, Karina González-García, Cristian Jiménez-Martínez, Luis Castro-Sánchez, Xariss Miryam Sánchez-Chino, Verónica Rocío Vásquez-Garzón, and Rafael Baltiérrez-Hoyos. 2022. "Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines" International Journal of Molecular Sciences 23, no. 9: 5032. https://doi.org/10.3390/ijms23095032
APA StyleVelázquez-Enríquez, J. M., Ramírez-Hernández, A. A., Navarro, L. M. S., Reyes-Avendaño, I., González-García, K., Jiménez-Martínez, C., Castro-Sánchez, L., Sánchez-Chino, X. M., Vásquez-Garzón, V. R., & Baltiérrez-Hoyos, R. (2022). Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines. International Journal of Molecular Sciences, 23(9), 5032. https://doi.org/10.3390/ijms23095032