
Seroma after Simple Mastectomy in Breast Cancer—The Role of CD4+ T Helper Cells and the Evidence as a Possible Specific Immune Process


 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Patient Cohort
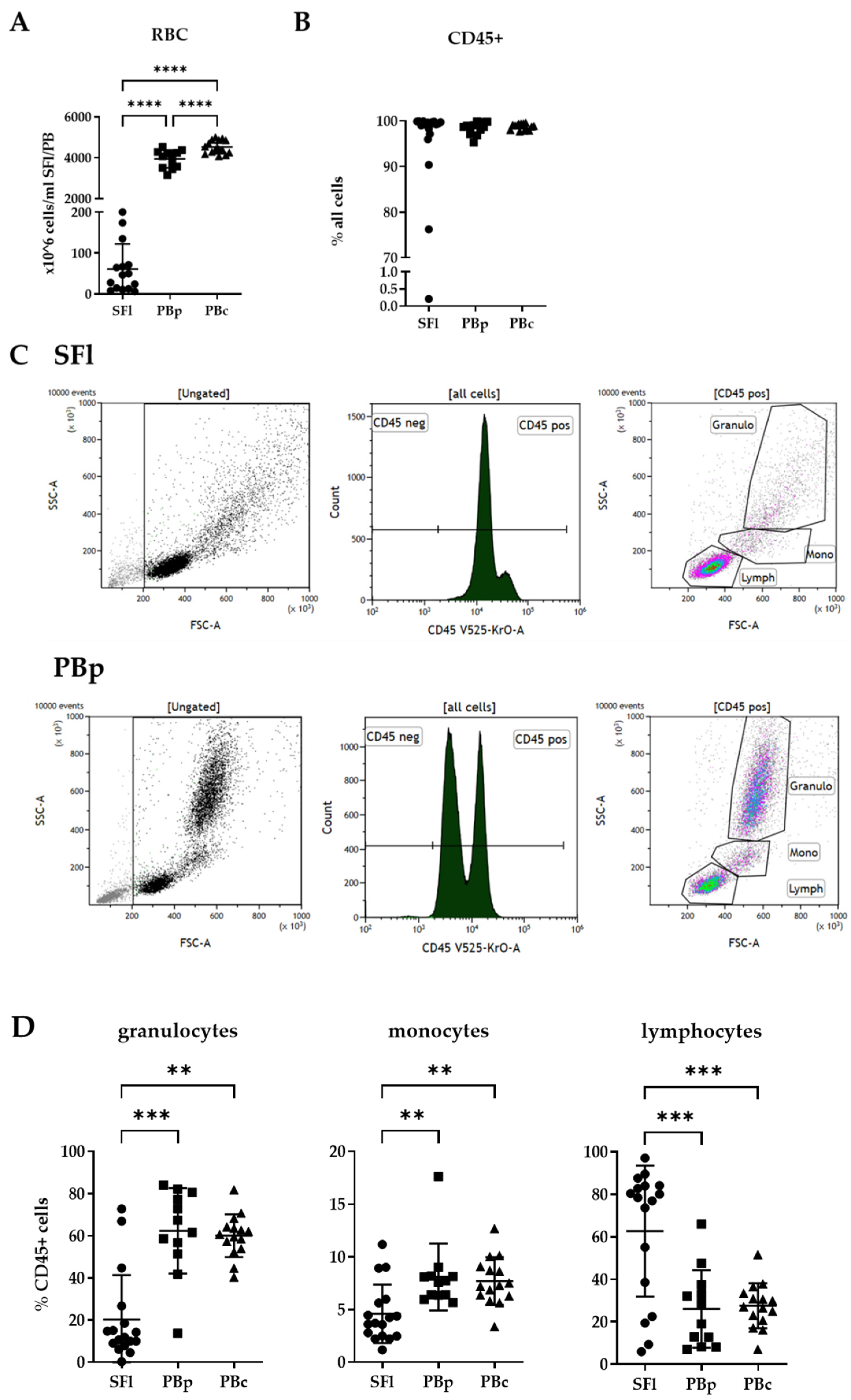
2.2. Basic Cell Composition in Seroma Fluids
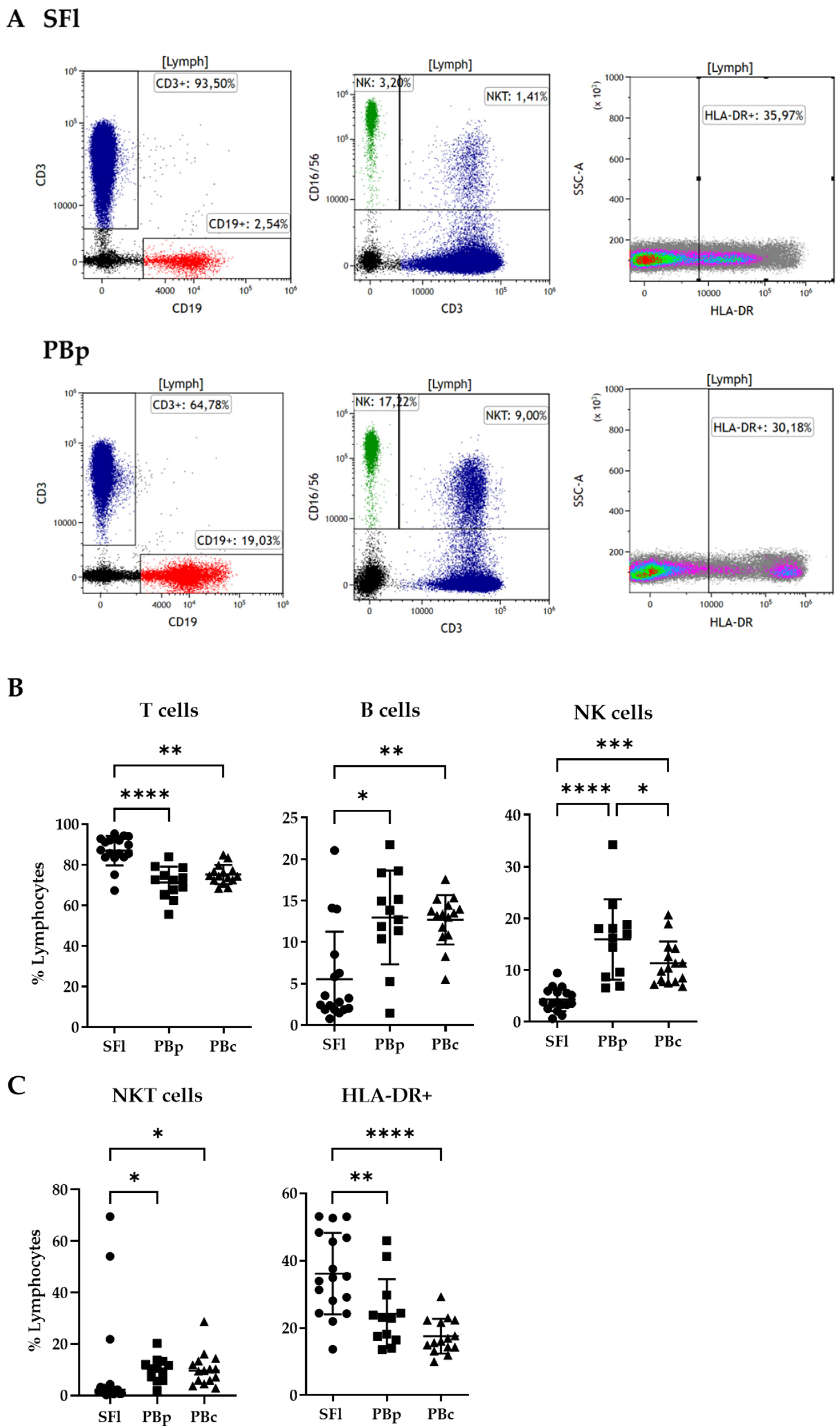
2.3. Lymphocyte Composition in Seroma Liquid and PB Analyzed by Flow Cytometry
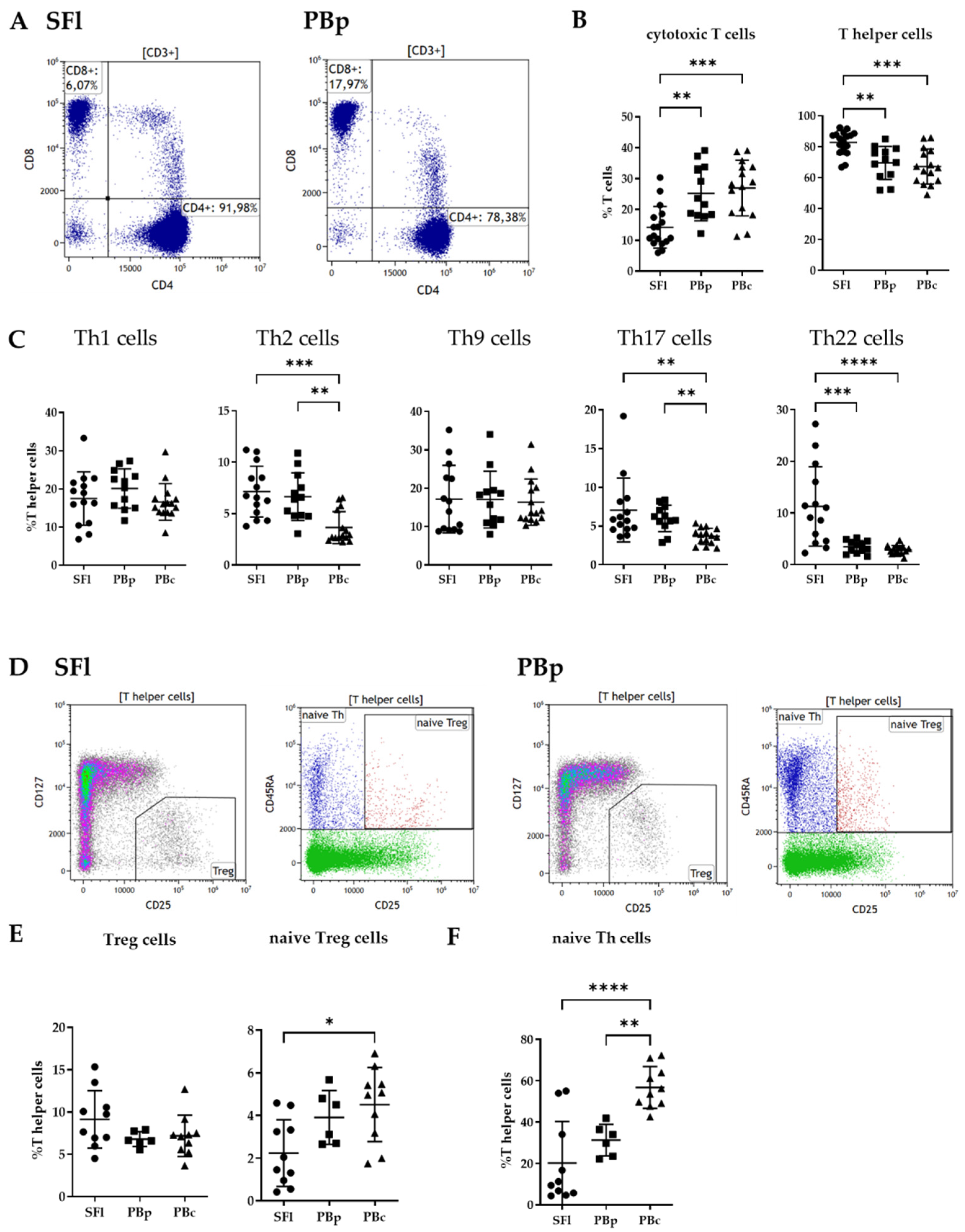
2.4. T-Cell Identification
2.4.1. Cytotoxic vs. T-Helper Cell Analysis
2.4.2. T Helper Cell Subpopulation Analysis
2.4.3. Treg Subpopulation
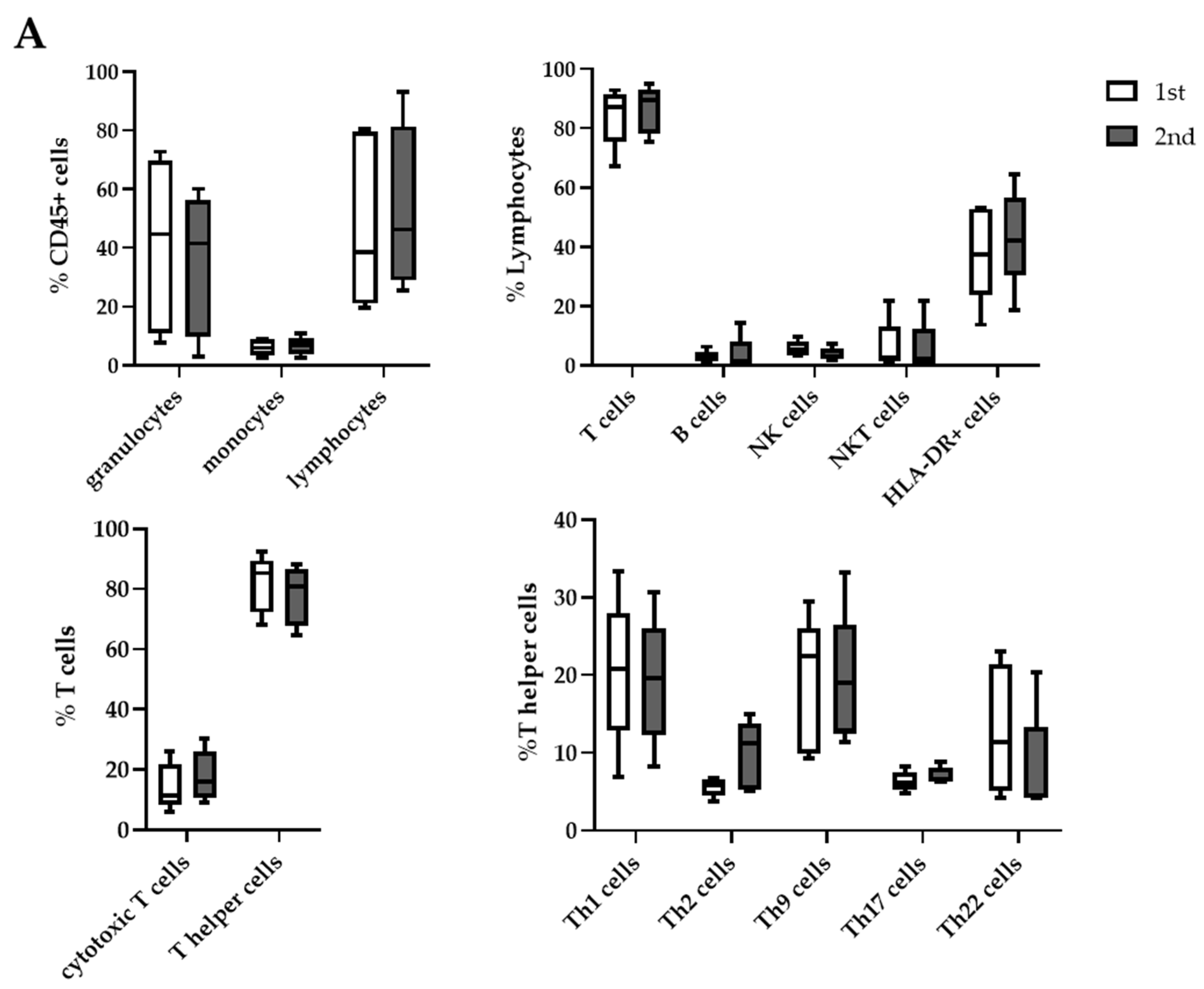
2.5. Differences in Cell Distribution between First and Second Aspiration
3. Discussion
4. Materials and Methods
4.1. Patient Cohort
4.2. Sample Processing
4.3. Flow Cytometry Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Gaithy, Z.K.; Ayuob, N.N. Vascular and cellular events in post-mastectomy seroma: An immunohistochemical study. Cell Immunol. 2012, 272, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Andeweg, C.S.; Schriek, M.J.; Heisterkamp, J.; Roukema, J.A. Seroma formation in two cohorts after axillary lymph node dissection in breast cancer surgery: Does timing of drain removal matter? Breast J. 2011, 17, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Carless, P.A.; Henry, D.A. Systematic review and meta-analysis of the use of fibrin sealant to prevent seroma formation after breast cancer surgery. Br. J. Surg. 2006, 93, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Olsen, M.A.; Margenthaler, J.A. Indications for readmission following mastectomy for breast cancer: An assessment of patient and operative factors. Breast J. 2020, 26, 1966–1972. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Basu, S.; Shukla, V.K. Seroma formation after breast cancer surgery: What we have learned in the last two decades. J Breast Cancer 2012, 15, 373–380. [Google Scholar] [CrossRef]
- Kuroi, K.; Shimozuma, K.; Taguchi, T.; Imai, H.; Yamashiro, H.; Ohsumi, S.; Saito, S. Pathophysiology of seroma in breast cancer. Breast Cancer 2005, 12, 288–293. [Google Scholar] [CrossRef]
- Kumar, S.; Lal, B.; Misra, M.C. Post-mastectomy seroma: A new look into the aetiology of an old problem. J. R. Coll. Surg. Edinb. 1995, 40, 292–294. [Google Scholar]
- Persa, O.D.; Knuever, J.; Rose, A.; Mauch, C.; Schlaak, M. Predicting risk for seroma development after axillary or inguinal sentinel lymph node biopsy in melanoma patients. Int. J. Dermatol. 2019, 58, 185–189. [Google Scholar] [CrossRef]
- Hansson, E.; Edvinsson, A.C.; Elander, A.; Kolby, L.; Hallberg, H. First-year complications after immediate breast reconstruction with a biological and a synthetic mesh in the same patient: A randomized controlled study. J. Surg. Oncol. 2021, 123, 80–88. [Google Scholar] [CrossRef]
- Heidemann, L.N.; Gunnarsson, G.L.; Salzberg, C.A.; Sorensen, J.A.; Thomsen, J.B. Complications following Nipple-Sparing Mastectomy and Immediate Acellular Dermal Matrix Implant-based Breast Reconstruction-A Systematic Review and Meta-analysis. Plast. Reconstr. Surg. Glob. Open 2018, 6, e1625. [Google Scholar] [CrossRef]
- Hashemi, E.; Kaviani, A.; Najafi, M.; Ebrahimi, M.; Hooshmand, H.; Montazeri, A. Seroma formation after surgery for breast cancer. World J. Surg. Oncol. 2004, 2, 44. [Google Scholar] [CrossRef] [PubMed]
- Mancaux, A.; Naepels, P.; Mychaluk, J.; Abboud, P.; Merviel, P.; Fauvet, R. Prevention of seroma post-mastectomy by surgical padding technique. Gynecol. Obstet. Fertil. 2015, 43, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Andrades, P.; Prado, A. Composition of postabdominoplasty seroma. Aesthetic Plast. Surg. 2007, 31, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Watt-Boolsen, S.; Nielsen, V.B.; Jensen, J.; Bak, S. Postmastectomy seroma. A study of the nature and origin of seroma after mastectomy. Dan. Med. Bull. 1989, 36, 487–489. [Google Scholar]
- Montalto, E.; Mangraviti, S.; Costa, G.; Carrega, P.; Morandi, B.; Pezzino, G.; Bonaccorsi, I.; Cancellieri, A.; Mingari, M.C.; Mesiti, M.; et al. Seroma fluid subsequent to axillary lymph node dissection for breast cancer derives from an accumulation of afferent lymph. Immunol. Lett. 2010, 131, 67–72. [Google Scholar] [CrossRef]
- Farber, D.; Netea, M.G.; Radbruch, A.; Rajewsky, K.; Zinkernagel, R.M. Immunological memory: Lessons from the past and a look to the future. Nat. Rev. Immunol. 2016, 16, 124–128. [Google Scholar] [CrossRef]
- Danilova, N. The evolution of adaptive immunity. Adv. Exp. Med. Biol. 2012, 738, 218–235. [Google Scholar]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4+ T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef]
- Mitchison, N.A. The discovery of T cell-B cell cooperation. Front. Immunol. 2014, 5, 377. [Google Scholar] [CrossRef]
- Ye, J.; Wang, Y.; Wang, Z.; Ji, Q.; Huang, Y.; Zeng, T.; Hu, H.; Ye, D.; Wan, J.; Lin, Y. Circulating Th1, Th2, Th9, Th17, Th22, and Treg Levels in Aortic Dissection Patients. Mediat. Inflamm. 2018, 2018, 5697149. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. Heterogeneity and plasticity of T helper cells. Cell Res. 2010, 20, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Cantor, H. CD4 T-cell Subsets and Tumor Immunity: The Helpful and the Not-so-Helpful. Cancer Immunol. Res. 2014, 2, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.-F.; Huan, J.; Qin, X.-J. Potential risk factors for the development of seroma following mastectomy with axillary dissection. Mol. Clin. Oncol. 2015, 3, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.M.; Harper, J.L.; Dragun, A.E.; Ashenafi, M.S.; Sinha, D.; Li, J.; Cole, D.J.; Jenrette, J.M. Incidence and prognostic factors for seroma development after MammoSite breast brachytherapy. Brachytherapy 2008, 7, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Storek, J.; Ferrara, S.; Isacescu, V. Blood B cell subpopulations. The effect of narrow versus wide forward scatter x side scatter gating. J. Immunol. Methods 1992, 156, 129–133. [Google Scholar] [CrossRef]
- Comans-Bitter, W.; de Groot, R.; Beemd, R.V.D.; Neijens, H.J.; Hop, W.C.; Groeneveld, K.; Hooijkaas, H.; Van Dongen, J. Immunophenotyping of blood lymphocytes in childhoodReference values for lymphocyte subpopulations. J. Pediatr. 1997, 130, 388–393. [Google Scholar] [CrossRef]
- Schatorjé, E.J.H.; Gemen, E.F.A.; Driessen, G.J.A.; Leuvenink, J.; Van Hout, R.W.N.M.; De Vries, E. Paediatric Reference Values for the Peripheral T cell Compartment. Scand. J. Immunol. 2012, 75, 436–444. [Google Scholar] [CrossRef]
- Christian Neumann, A.S.; Nicole, P.; Anna, F.-M.; Michaela, N.; Julia, M.; Anne, R.; Tobias, S. T Helper Cells: A Complete Workflow for Cell Preparation, Isolation, Stimulation, Polarization, and Analysis; Miltenyi Biotec GmbH: Gütersloh, Germany, 2015. [Google Scholar]
- Yu, N.; Li, X.; Song, W.; Li, D.; Yu, D.; Zeng, X.; Li, M.; Leng, X.; Li, X. CD4(+)CD25 (+)CD127 (low/-) T cells: A More Specific Treg Population in Human Peripheral Blood. Inflammation 2012, 35, 1773–1780. [Google Scholar] [CrossRef]
- Moradi, B.; Schnatzer, P.; Hagmann, S.; Rosshirt, N.; Gotterbarm, T.; Kretzer, J.P.; Thomsen, M.; Lorenz, H.M.; Zeifang, F.; Tretter, T. CD4(+)CD25(+)/highCD127low/(-) regulatory T cells are enriched in rheumatoid arthritis and osteoarthritis joints—analysis of frequency and phenotype in synovial membrane, synovial fluid and peripheral blood. Arthritis Res. Ther. 2014, 16, R97. [Google Scholar] [CrossRef]
- Booth, N.J.; McQuaid, A.J.; Sobande, T.; Kissane, S.; Agius, E.; Jackson, S.E.; Salmon, M.; Falciani, F.; Yong, K.; Rustin, M.H.; et al. Different Proliferative Potential and Migratory Characteristics of Human CD4+ Regulatory T Cells That Express either CD45RA or CD45RO. J. Immunol. 2010, 184, 4317–4326. [Google Scholar] [CrossRef] [PubMed]
- Daae, L.N.; Halvorsen, S.; Mathisen, P.M.; Mironska, K. A comparison between haematological parameters in ’capillary’ and venous blood from healthy adults. Scand. J. Clin. Lab. Investig. 1988, 48, 723–726. [Google Scholar]
- Tippalagama, R.; Singhania, A.; Dubelko, P.; Arlehamn, C.S.L.; Crinklaw, A.; Pomaznoy, M.; Seumois, G.; Desilva, A.D.; Premawansa, S.; Vidanagama, D.; et al. HLA-DR Marks Recently Divided Antigen-Specific Effector CD4 T Cells in Active Tuberculosis Patients. J. Immunol. 2021, 207, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Camous, X.; Pera, A.; Solana, R.; Larbi, A. NK Cells in Healthy Aging and Age-Associated Diseases. J. Biomed. Biotechnol. 2012, 2012, 195956. [Google Scholar] [CrossRef]
- Solana, R.; Mariani, E. NK and NK/T cells in human senescence. Vaccine 2000, 18, 1613–1620. [Google Scholar] [CrossRef]
- Provinciali, M.; Moresi, R.; Donnini, A.; Lisa, R.M. Reference values for CD4+ and CD8+ T lymphocytes with naive or memory phenotype and their association with mortality in the elderly. Gerontology 2009, 55, 314–321. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, W.-W.; Kim, S.H.; Kang, Y.; Lee, N.; Shin, M.S.; Kang, S.W.; Kang, I. Age-associated alteration in naive and memory Th17 cell response in humans. Clin. Immunol. 2011, 140, 84–91. [Google Scholar] [CrossRef]
- Besler, E.; Harmancioglu, O. Examination of the Effects of Celecoxib on Postmastectomy Seroma and Wound Healing. Med. Bull. Sisli Etfal Hosp. 2018, 52, 212–219. [Google Scholar] [CrossRef]
- Doulabi, H.; Masoumi, E.; Rastin, M.; Foolady Azarnaminy, A.; Esmaeili, S.A.; Mahmoudi, M. The role of Th22 cells, from tissue repair to cancer progression. Cytokine 2022, 149, 155749. [Google Scholar] [CrossRef]
- Gregor, C.E.; Foeng, J.; Comerford, I.; McColl, S.R. Chemokine-Driven CD4(+) T Cell Homing: New Concepts and Recent Advances. Adv. Immunol. 2017, 135, 119–181. [Google Scholar]
- Morandi, B.; Bonaccorsi, I.; Mesiti, M.; Conte, R.; Carrega, P.; Costa, G.; Iemmo, R.; Martini, S.; Ferrone, S.; Cantoni, C.; et al. Characterization of Human Afferent Lymph Dendritic Cells from Seroma Fluids. J. Immunol. 2013, 191, 4858–4866. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, D.; Mazzotta, M.; Pizzuti, L.; Krasniqi, E.; Gamucci, T.; Natoli, C.; Grassadonia, A.; Tinari, N.; Tomao, S.; Sperduti, I.; et al. Neoadjuvant Immune-Checkpoint Blockade in Triple-Negative Breast Cancer: Current Evidence and Literature-Based Meta-Analysis of Randomized Trials. Cancers 2020, 12, 2497. [Google Scholar] [CrossRef]
- Watts, T.H. Stepping up Th1 immunity to control phagosomal bacteria. Trends Immunol. 2021, 42, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Characteristics | Subgroups | n (Number) |
|---|---|---|
| Histopathological type | ||
| NST Invasive lobular mucinous apocrine only DCIS solid papillary | 11 3 1 1 1 1 | |
| Focality (2 bilateral carcinomas) | ||
| unifocal multifocal multicentric | 12 2 4 | |
| Hormone receptor status (without DCIS, 2 bilateral carcinomas) | ||
| HR + ER-/PR- | 14 3 | |
| Her-2/neu (without DCIS; 2 bilateral carcinomas) | ||
| positive negative | 2 15 | |
| Ki67 (without DCIS; 2 bilateral carcinomas) | ||
| <20% ≥20% | 8 9 | |
| Tumor size (2 bilateral carcinomas) | ||
| ypT0 pTis (y)pT1a (y)pT1b (y)pT1c (y)pT2 (y)pT3 (y)pT4 | 3 1 1 2 2 5 4 0 | |
| Axillary nodal status (2 bilateral carcinomas) | ||
| (y)pN0 (y)pN1 (y)pN2 | 12 3 2 | |
| Grading (without DCIS; 2 bilateral carcinomas) | ||
| G1 G2 G3 | 2 11 4 |
| Panel 1 (all Beckman Coulter) | Fluorescent Label |
|---|---|
| anti-human CD45 antibody | Chromium Orange |
| anti-human CD3 antibody | APC-AF750 |
| anti-human CD19 antibody | ECD |
| anti-human CD4 antibody | APC |
| anti-human CD8 antibody | Pacific Blue |
| anti-human CD16 antibody | PE |
| anti-human CD56 antibody | PE |
| anti-human HLA-DR antibody | PC7 |
| Panel 2 (all Miltenyi Biotec) | |
| anti-human CD4 antibody | VioBlue |
| anti-human CD183 (CXCR3) antibody | VioBright FITC |
| anti-human CD194 (CXCR4) antibody | PE-Vio 770 |
| anti-human CD196 (CCR6) antibody | PE |
| anti-human CCR10 antibody | APC |
| Panel 1 (all Beckman Coulter) | |
| anti-human CD45 antibody | Chromium Orange |
| anti-human CD3 antibody | APC-AF750 |
| anti-human CD4 antibody | APC |
| anti-human CD127 antibody | FITC |
| anti-human CD25 antibody | PE |
| anti-human CD45RA antibody | ECD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pochert, N.; Schneider, M.; Ansorge, N.; Strieder, A.; Sagasser, J.; Reiger, M.; Traidl-Hoffmann, C.; Neumann, A.; Jeschke, U.; Dannecker, C.; et al. Seroma after Simple Mastectomy in Breast Cancer—The Role of CD4+ T Helper Cells and the Evidence as a Possible Specific Immune Process. Int. J. Mol. Sci. 2022, 23, 4848. https://doi.org/10.3390/ijms23094848
Pochert N, Schneider M, Ansorge N, Strieder A, Sagasser J, Reiger M, Traidl-Hoffmann C, Neumann A, Jeschke U, Dannecker C, et al. Seroma after Simple Mastectomy in Breast Cancer—The Role of CD4+ T Helper Cells and the Evidence as a Possible Specific Immune Process. International Journal of Molecular Sciences. 2022; 23(9):4848. https://doi.org/10.3390/ijms23094848
Chicago/Turabian StylePochert, Nicole, Mariella Schneider, Nadine Ansorge, Annamarie Strieder, Jacqueline Sagasser, Matthias Reiger, Claudia Traidl-Hoffmann, Avidan Neumann, Udo Jeschke, Christian Dannecker, and et al. 2022. "Seroma after Simple Mastectomy in Breast Cancer—The Role of CD4+ T Helper Cells and the Evidence as a Possible Specific Immune Process" International Journal of Molecular Sciences 23, no. 9: 4848. https://doi.org/10.3390/ijms23094848
APA StylePochert, N., Schneider, M., Ansorge, N., Strieder, A., Sagasser, J., Reiger, M., Traidl-Hoffmann, C., Neumann, A., Jeschke, U., Dannecker, C., Kühn, T., & Ditsch, N. (2022). Seroma after Simple Mastectomy in Breast Cancer—The Role of CD4+ T Helper Cells and the Evidence as a Possible Specific Immune Process. International Journal of Molecular Sciences, 23(9), 4848. https://doi.org/10.3390/ijms23094848