
Myoglobin Interaction with Lactate Rapidly Releases Oxygen: Studies on Binding Thermodynamics, Spectroscopy, and Oxygen Kinetics

 ,
,
Abstract
1. Introduction
2. Results and Discussion
3. Materials & Methods
3.1. Materials
3.2. Preparation of Mb
3.3. Ligand Binding Studies
3.4. Oxygen Kinetic Studies
3.5. Time-Resolved Spectroscopic Studies
3.6. Molecular Docking
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| deoxy-Mb | deoxygenated myoglobin |
| LAC | sodium lactate |
| Mb | myoglobin |
| oxy-Mb | oxygenated myoglobin |
References
- Juel, C.; Halestrap, A.P. Lactate transport in skeletal muscle—role and regulation of the monocarboxylate transporter. J. Physiol. 1999, 517, 633–642. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of Biochemistry Life at the Molecular Level; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.D.; Enerback, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Lactate as a fulcrum of metabolism. Redox Biol. 2020, 35, 101454. [Google Scholar] [CrossRef] [PubMed]
- Proia, P.; Di Liegro, C.M.; Schiera, G.; Fricano, A.; Di Liegro, I. Lactate as a Metabolite and a Regulator in the Central Nervous System. Int. J. Mol. Sci. 2016, 17, 1450. [Google Scholar] [CrossRef]
- Henderson, G.C.; Horning, M.A.; Wallis, G.A.; Brooks, G.A. Pyruvate metabolism in working human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E366. [Google Scholar] [CrossRef]
- Rogatzki, M.J.; Ferguson, B.S.; Goodwin, M.L.; Gladden, L.B. Lactate is always the end product of glycolysis. Front. Neurosci. 2015, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Khacho, M.; Tarabay, M.; Patten, D.; Khacho, P.; MacLaurin, J.G.; Guadagno, J.; Bergeron, R.; Cregan, S.P.; Harper, M.E.; Park, D.S.; et al. Acidosis overrides oxygen deprivation to maintain mitochondrial function and cell survival. Nat. Commun. 2014, 5, 3550. [Google Scholar] [CrossRef]
- Davis, J.A. Anaerobic threshold: Review of the concept and directions for future research. Med. Sci. Sports Exerc. 1985, 17, 6–21. [Google Scholar] [CrossRef]
- Foucher, C.D.; Tubben, R.E. Lactic Acidosis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Luft, F.C. Lactic acidosis update for critical care clinicians. J. Am. Soc. Nephrol. 2001, 12 (Suppl. 17), S15–S19. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; W.H. Freeman: New York, NY, USA, 2017. [Google Scholar]
- Gertz, E.W.; Wisneski, J.A.; Neese, R.; Bristow, J.D.; Searle, G.L.; Hanlon, J.T. Myocardial lactate metabolism: Evidence of lactate release during net chemical extraction in man. Circulation 1981, 63, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Diakos, N.A.; Navankasattusas, S.; Abel, E.D.; Rutter, J.; McCreath, L.; Ferrin, P.; McKellar, S.H.; Miller, D.V.; Park, S.Y.; Richardson, R.S.; et al. Evidence of Glycolysis Up-Regulation and Pyruvate Mitochondrial Oxidation Mismatch During Mechanical Unloading of the Failing Human Heart: Implications for Cardiac Reloading and Conditioning. JACC Basic Transl. Sci. 2016, 1, 432–444. [Google Scholar] [CrossRef]
- Neubauer, S.; Horn, M.; Cramer, M.; Harre, K.; Newell, J.B.; Peters, W.; Pabst, T.; Ertl, G.; Hahn, D.; Ingwall, J.S.; et al. Myocardial phosphocreatine-to-ATP ratio is a predictor of mortality in patients with dilated cardiomyopathy. Circulation 1997, 96, 2190–2196. [Google Scholar] [CrossRef]
- Wittenberg, B.A.; Wittenberg, J.B.; Caldwell, P.R. Role of myoglobin in the oxygen supply to red skeletal muscle. J. Biol. Chem. 1975, 250, 9038–9043. [Google Scholar] [CrossRef]
- Cole, R.P. Skeletal muscle function in hypoxia: Effect of alteration of intracellular myoglobin. Respir. Physiol. 1983, 53, 1–14. [Google Scholar] [CrossRef]
- Driedzic, W.R. The fish heart as a model system for the study of myoglobin. Comp. Biochem. Physiol. A Comp. Physiol. 1983, 76, 487–493. [Google Scholar] [CrossRef]
- Taylor, D.J.; Matthews, P.M.; Radda, G.K. Myoglobin-dependent oxidative metabolism in the hypoxic rat heart. Respir. Physiol. 1986, 63, 275–283. [Google Scholar] [CrossRef]
- Wittenberg, B.A.; Wittenberg, J.B. Myoglobin-mediated oxygen delivery to mitochondria of isolated cardiac myocytes. Proc. Natl. Acad. Sci. USA 1987, 84, 7503–7507. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, E.Y.; Chupyrkina, A.A.; Pevzner, I.B.; Isaev, N.K.; Zorov, D.B. Myoglobin causes oxidative stress, increase of NO production and dysfunction of kidney’s mitochondria. Biochim. Biophys. Acta 2009, 1792, 796–803. [Google Scholar] [CrossRef]
- Park, J.W.; Piknova, B.; Dey, S.; Noguchi, C.T.; Schechter, A.N. Compensatory mechanisms in myoglobin deficient mice preserve NO homeostasis. Nitric. Oxide 2019, 90, 10–14. [Google Scholar] [CrossRef]
- Nakamura, M.; Nakamura, S. Conversion of metmyoglobin to NO myoglobin in the presence of nitrite and reductants. Biochim. Biophys. Acta 1996, 1289, 329–335. [Google Scholar] [CrossRef]
- Eich, R.F.; Li, T.; Lemon, D.D.; Doherty, D.H.; Curry, S.R.; Aitken, J.F.; Mathews, A.J.; Johnson, K.A.; Smith, R.D.; Phillips, G.N., Jr.; et al. Mechanism of NO-induced oxidation of myoglobin and hemoglobin. Biochemistry 1996, 35, 6976–6983. [Google Scholar] [CrossRef] [PubMed]
- Moller, J.K.; Skibsted, L.H. Nitric oxide and myoglobins. Chem. Rev. 2002, 102, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Cossins, A.; Berenbrink, M. Physiology: Myoglobin’s new clothes. Nature 2008, 454, 416–417. [Google Scholar] [CrossRef]
- Chintapalli, S.V.; Anishkin, A.; Adams, S.H. Exploring the entry route of palmitic acid and palmitoylcarnitine into myoglobin. Arch. Biochem. Biophys. 2018, 655, 56–66. [Google Scholar] [CrossRef]
- Chintapalli, S.V.; Bhardwaj, G.; Patel, R.; Shah, N.; Patterson, R.L.; van Rossum, D.B.; Anishkin, A.; Adams, S.H. Molecular dynamic simulations reveal the structural determinants of Fatty Acid binding to oxy-myoglobin. PLoS ONE 2015, 10, e0128496. [Google Scholar] [CrossRef]
- Chintapalli, S.V.; Jayanthi, S.; Mallipeddi, P.L.; Gundampati, R.; Suresh Kumar, T.K.; van Rossum, D.B.; Anishkin, A.; Adams, S.H. Novel Molecular Interactions of Acylcarnitines and Fatty Acids with Myoglobin. J. Biol. Chem. 2016, 291, 25133–25143. [Google Scholar] [CrossRef] [PubMed]
- Gloster, J.; Harris, P. Fatty acid binding to cytoplasmic proteins of myocardium and red and white skeletal muscle in the rat. A possible new role for myoglobin. Biochem. Biophys. Res. Commun. 1977, 74, 506–513. [Google Scholar] [CrossRef]
- Gotz, F.M.; Hertel, M.; Groschel-Stewart, U. Fatty acid binding of myoglobin depends on its oxygenation. Biol. Chem. Hoppe-Seyler 1994, 375, 387–392. [Google Scholar] [CrossRef]
- Jue, T.; Shih, L.; Chung, Y. Differential Interaction of Myoglobin with Select Fatty Acids of Carbon Chain Lengths C8 to C16. Lipids 2017, 52, 711–727. [Google Scholar] [CrossRef]
- Jue, T.; Simond, G.; Wright, T.J.; Shih, L.; Chung, Y.; Sriram, R.; Kreutzer, U.; Davis, R.W. Effect of fatty acid interaction on myoglobin oxygen affinity and triglyceride metabolism. J. Physiol. Biochem. 2016, 73, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Sriram, R.; Kreutzer, U.; Shih, L.; Jue, T. Interaction of fatty acid with myoglobin. FEBS Lett. 2008, 582, 3643–3649. [Google Scholar] [CrossRef]
- Schlater, A.E.; De Miranda, M.A., Jr.; Frye, M.A.; Trumble, S.J.; Kanatous, S.B. Changing the paradigm for myoglobin: A novel link between lipids and myoglobin. J. Appl. Physiol. 2014, 117, 307–315. [Google Scholar] [CrossRef]
- Tichivangana, J.Z.; Morrissey, P.A. Metmyoglobin and inorganic metals as pro-oxidants in raw and cooked muscle systems. Meat Sci. 1985, 15, 107–116. [Google Scholar] [CrossRef]
- Galaris, D.; Sevanian, A.; Cadenas, E.; Hochstein, P. Ferrylmyoglobin-catalyzed linoleic acid peroxidation. Arch. Biochem. Biophys. 1990, 281, 163–169. [Google Scholar] [CrossRef]
- Chan, W.K.; Faustman, C.; Yin, M.; Decker, E.A. Lipid oxidation induced by oxymyoglobin and metmyoglobin with involvement of H(2)O(2) and superoxide anion. Meat Sci. 1997, 46, 181–190. [Google Scholar] [CrossRef]
- Baron, C.P.; Skibsted, L.H.; Andersen, H.J. Concentration effects in myoglobin-catalyzed peroxidation of linoleate. J. Agric. Food Chem. 2002, 50, 883–888. [Google Scholar] [CrossRef]
- Vuletich, J.L.; Osawa, Y.; Aviram, M. Enhanced lipid oxidation by oxidatively modified myoglobin: Role of protein-bound heme. Biochem. Biophys. Res. Commun. 2000, 269, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Baron, C.P.; Andersen, H.J. Myoglobin-induced lipid oxidation. A review. J. Agric. Food Chem. 2002, 50, 3887–3897. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.L.; Wankhade, U.D.; Ono-Moore, K.D.; Chintapalli, S.V.; Fox, R.; Rutkowsky, J.M.; Willis, B.J.; Tolentino, T.; Lloyd, K.C.K.; Adams, S.H. On the potential role of globins in brown adipose tissue: A novel conceptual model and studies in myoglobin knockout mice. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E47–E62. [Google Scholar] [CrossRef] [PubMed]
- Rassaf, T.; Flogel, U.; Drexhage, C.; Hendgen-Cotta, U.; Kelm, M.; Schrader, J. Nitrite reductase function of deoxymyoglobin: Oxygen sensor and regulator of cardiac energetics and function. Circ. Res. 2007, 100, 1749–1754. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Furuichi, Y.; Takakura, H.; Hashimoto, T.; Hanai, Y.; Jue, T.; Masuda, K. Interaction between myoglobin and mitochondria in rat skeletal muscle. J. Appl. Physiol. 2013, 114, 490–497. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Edwards, D.L.; Criddle, R.S. The interaction of myoglobin with mitochondrial structural protein. Biochemistry 1966, 5, 588–591. [Google Scholar] [CrossRef]
- Postnikova, G.B.; Tselikova, S.V.; Shekhovtsova, E.A. Myoglobin and mitochondria: Oxymyoglobin interacts with mitochondrial membrane during deoxygenation. Biochemistry 2009, 74, 1211–1218. [Google Scholar] [CrossRef]
- Postnikova, G.B.; Shekhovtsova, E.A. Fluorescence studies on the interaction of myoglobin with mitochondria. Biochemistry 2012, 77, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bocedi, A.; Bolli, A.; Fasano, M.; Notari, S.; Polticelli, F. Allosteric modulation of monomeric proteins. Biochem. Mol. Biol. Educ. 2005, 33, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Giardina, B.; Ascenzi, P.; Clementi, M.E.; De Sanctis, G.; Rizzi, M.; Coletta, M. Functional modulation by lactate of myoglobin. A monomeric allosteric hemoprotein. J. Biol. Chem. 1996, 271, 16999–17001. [Google Scholar] [CrossRef]
- Turnbull, W.B.; Daranas, A.H. On the value of c: Can low affinity systems be studied by isothermal titration calorimetry? J. Am. Chem. Soc. 2003, 125, 14859–14866. [Google Scholar] [CrossRef]
- Kamatchi, S. Studies on pH-Controlled Transition of Myoglobin Capsules from Hollow to Multilayered Structures. J. Adsorpt. Sci. Technol. 2015, 33, 10. [Google Scholar] [CrossRef]
- Butler, I.B.; Schoonen, M.A.; Rickard, D.T. Removal of dissolved oxygen from water: A comparison of four common techniques. Talanta 1994, 41, 211–215. [Google Scholar] [CrossRef]
- Phillips, S.E. Structure and refinement of oxymyoglobin at 1.6 A resolution. J. Mol. Biol. 1980, 142, 531–554. [Google Scholar] [CrossRef]
- Collman, J.P.; Brauman, J.I.; Doxsee, K.M. Carbon monoxide binding to iron porphyrins. Proc. Natl. Acad. Sci. USA 1979, 76, 6035–6039. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.; Jones, C.M.; Henry, E.R.; Hofrichter, J.; Eaton, W.A. Conformational relaxation and ligand binding in myoglobin. Biochemistry 1994, 33, 5128–5145. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.P.; Hoekstra, J.W. Oxidation of nitrogen oxides by bound dioxygen in hemoproteins. J. Inorg. Biochem. 1981, 14, 351–358. [Google Scholar] [CrossRef]
- Olson, J.S.; Mathews, A.J.; Rohlfs, R.J.; Springer, B.A.; Egeberg, K.D.; Sligar, S.G.; Tame, J.; Renaud, J.P.; Nagai, K. The role of the distal histidine in myoglobin and haemoglobin. Nature 1988, 336, 265–266. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.S.; Soman, J.; Phillips, G.N., Jr. Ligand pathways in myoglobin: A review of Trp cavity mutations. IUBMB Life 2007, 59, 552–562. [Google Scholar] [CrossRef]
- Scott, E.E.; Gibson, Q.H. Ligand migration in sperm whale myoglobin. Biochemistry 1997, 36, 11909–11917. [Google Scholar] [CrossRef]
- Scott, E.E.; Gibson, Q.H.; Olson, J.S. Mapping the pathways for O2 entry into and exit from myoglobin. J. Biol. Chem. 2001, 276, 5177–5188. [Google Scholar] [CrossRef]
- Tian, W.D.; Sage, J.T.; Champion, P.M. Investigations of ligand association and dissociation rates in the “open” and “closed” states of myoglobin. J. Mol. Biol. 1993, 233, 155–166. [Google Scholar] [CrossRef]
- Yang, F.; Phillips, G.N., Jr. Crystal structures of CO-, deoxy- and met-myoglobins at various pH values. J. Mol. Biol. 1996, 256, 762–774. [Google Scholar] [CrossRef]
- Tuesdale, G.A.; Downing, A.L. Solubility of oxygen in water. Nature 1954, 173, 1236. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.H.; Sondergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pKa Predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Ono-Moore, K.D.; Olfert, I.M.; Rutkowsky, J.M.; Chintapalli, S.V.; Willis, B.J.; Blackburn, M.L.; Williams, D.K.; O’Reilly, J.; Tolentino, T.; Lloyd, K.C.K.; et al. Metabolic physiology and skeletal muscle phenotypes in male and female myoglobin knockout mice. Am. J. Physiology. Endocrinol. Metab. 2021, 321, E63–E79. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
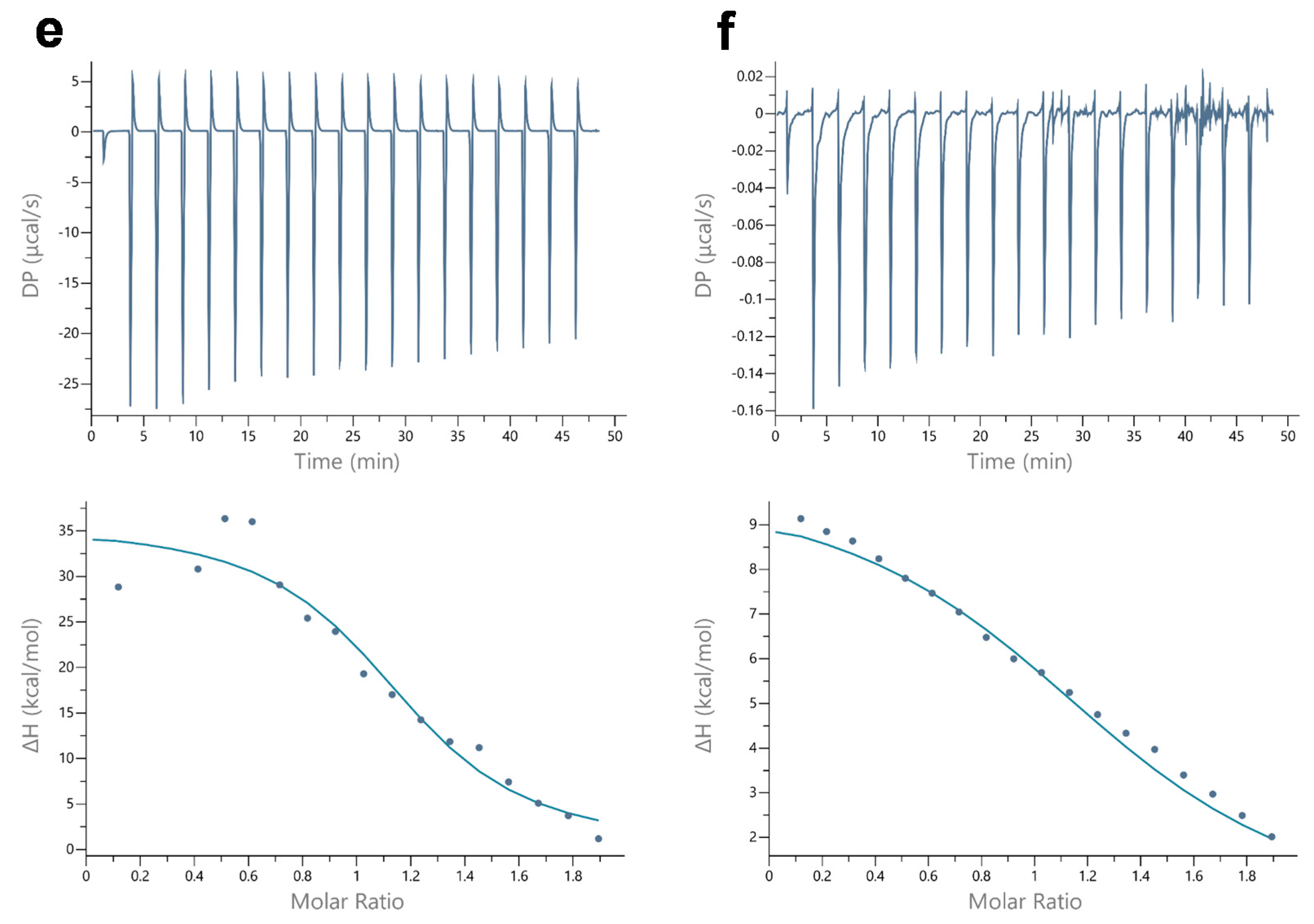
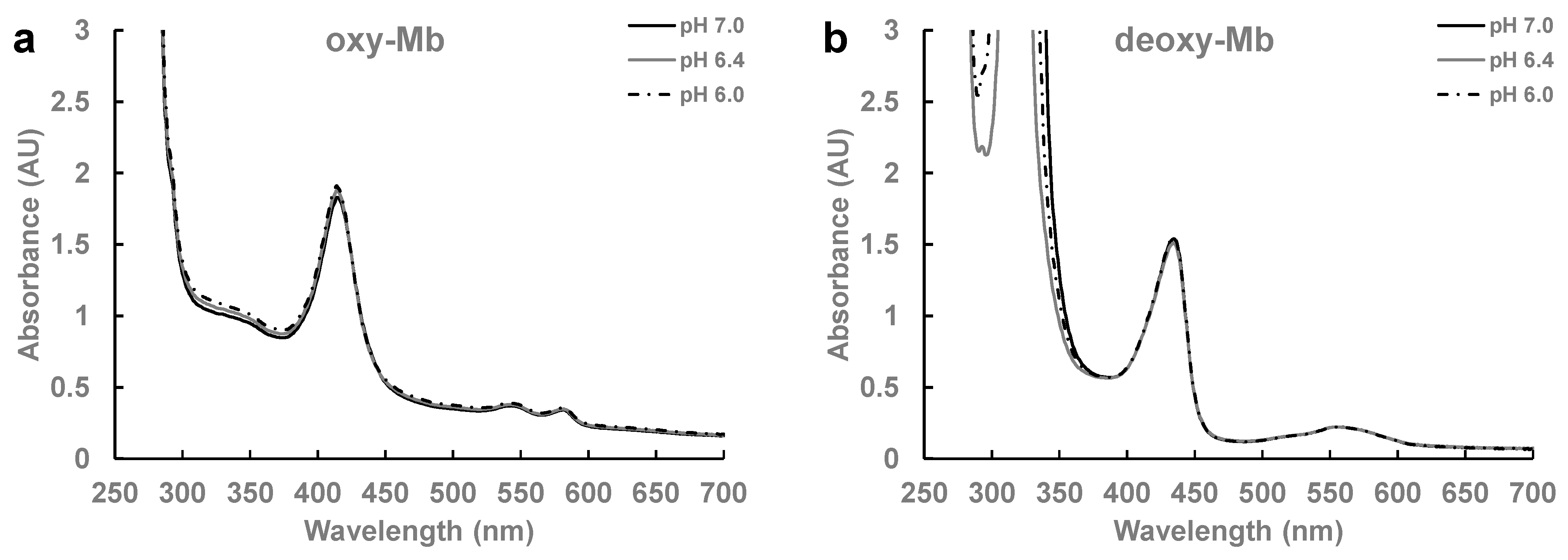
| Thermal Properties | pH 7.0 | pH 6.4 | pH 6.0 | |||
|---|---|---|---|---|---|---|
| oxy-Mb | deoxy-Mb | oxy-Mb | deoxy-Mb | oxy-Mb | deoxy-Mb | |
| Kd (µM) | 20.7 ± 2.7 a* | - | 6.9± 1.1 b1 | 13.7± 1.6 c1* | 1.9 ± 0.2 d2 | 6.4 ± 0.2 b1 |
| Ka (µM) | 0.05 ± 0.0 a* | - | 0.2 ± 0.03 b1 | 0.1± 0.0 c1* | 0.5 ± 0.04 d2 | 0.15 ± 0.0 b1 |
| c value | 22.4 ± 1.4 a | - | 67.5 ± 15.5 b1 | 30.4 ± 4.6 c1* | 267.9 ± 30.4 d2 | 61.2 ± 0.9 b1 |
| ΔH° (kcal mol−1) | −53.1 ± 0.3 a* | - | 151.5 ± 18.5 b1 | 149.5 ± 13.0 b1 | 34 ± 3.5 c2 | 12.6 ± 0.4 d3 |
| ΔS° (cal mol−1 K−1) | −0.15 ± 0.01 a* | - | 0.5 ± 0.06 b1 | 0.55 ± 0.03 b1 | 0.14 ± 0.01 c2 | 0.1 ± 0.0 d3 |
| No. of binding sites | 0.9 ± 0.7 b* | NDB | 0.8 ± 0.03 a1 | 0.8 ± 0.03 a1 | 1.0 ± 0.06 a2 | 0.8 ± 0.02 a2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adepu, K.K.; Bhandari, D.; Anishkin, A.; Adams, S.H.; Chintapalli, S.V. Myoglobin Interaction with Lactate Rapidly Releases Oxygen: Studies on Binding Thermodynamics, Spectroscopy, and Oxygen Kinetics. Int. J. Mol. Sci. 2022, 23, 4747. https://doi.org/10.3390/ijms23094747
Adepu KK, Bhandari D, Anishkin A, Adams SH, Chintapalli SV. Myoglobin Interaction with Lactate Rapidly Releases Oxygen: Studies on Binding Thermodynamics, Spectroscopy, and Oxygen Kinetics. International Journal of Molecular Sciences. 2022; 23(9):4747. https://doi.org/10.3390/ijms23094747
Chicago/Turabian StyleAdepu, Kiran Kumar, Dipendra Bhandari, Andriy Anishkin, Sean H. Adams, and Sree V. Chintapalli. 2022. "Myoglobin Interaction with Lactate Rapidly Releases Oxygen: Studies on Binding Thermodynamics, Spectroscopy, and Oxygen Kinetics" International Journal of Molecular Sciences 23, no. 9: 4747. https://doi.org/10.3390/ijms23094747
APA StyleAdepu, K. K., Bhandari, D., Anishkin, A., Adams, S. H., & Chintapalli, S. V. (2022). Myoglobin Interaction with Lactate Rapidly Releases Oxygen: Studies on Binding Thermodynamics, Spectroscopy, and Oxygen Kinetics. International Journal of Molecular Sciences, 23(9), 4747. https://doi.org/10.3390/ijms23094747