
Blocking the PCNA/NKp44 Checkpoint to Stimulate NK Cell Responses to Multiple Myeloma

 , , ,
, , ,  and
and 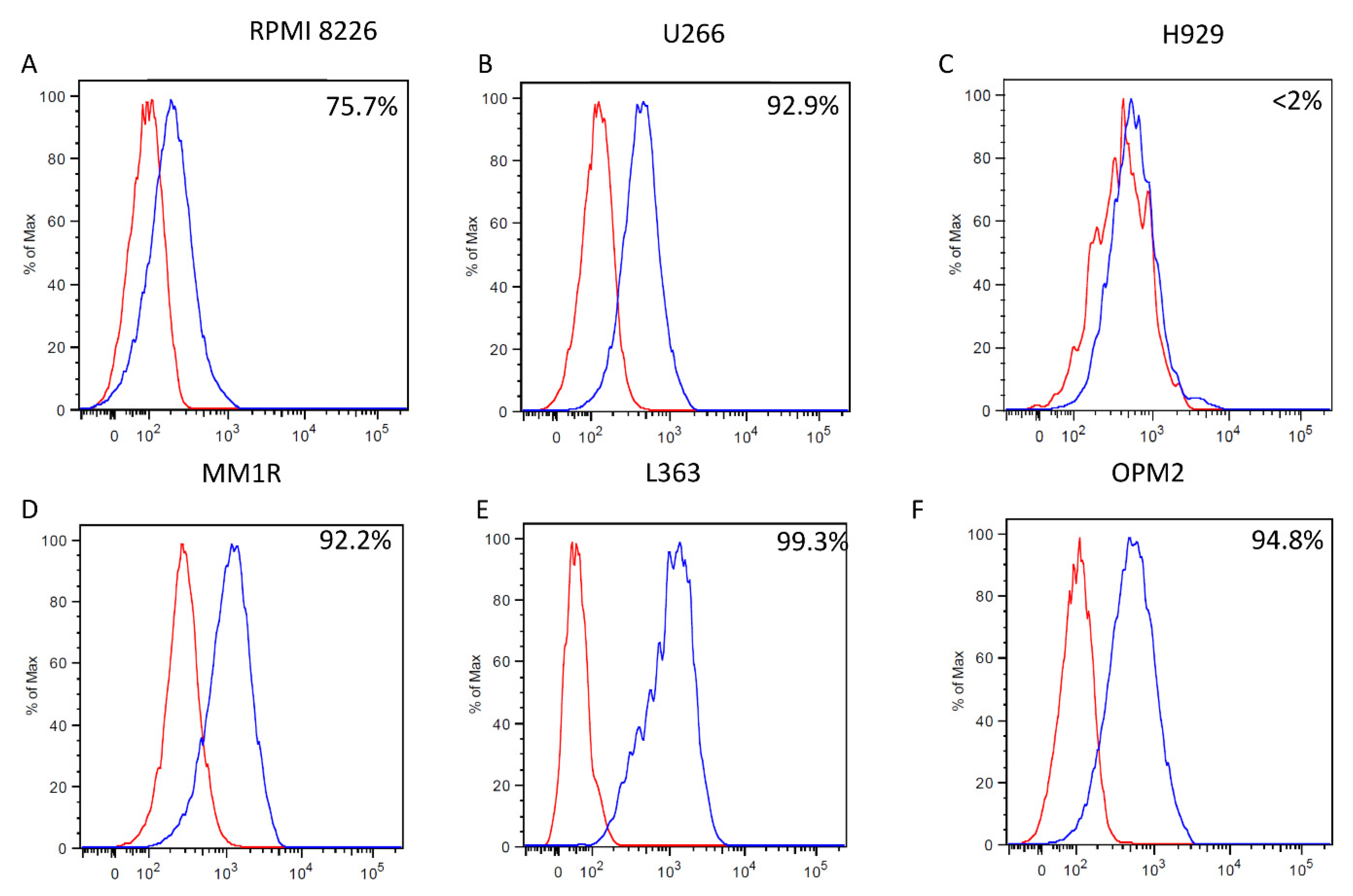{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Results
2.1. mAb 14-25-9 Detects Membrane PCNA on the Surface of Multiple Myeloma Cell Lines, and on Primary Myeloma Cells
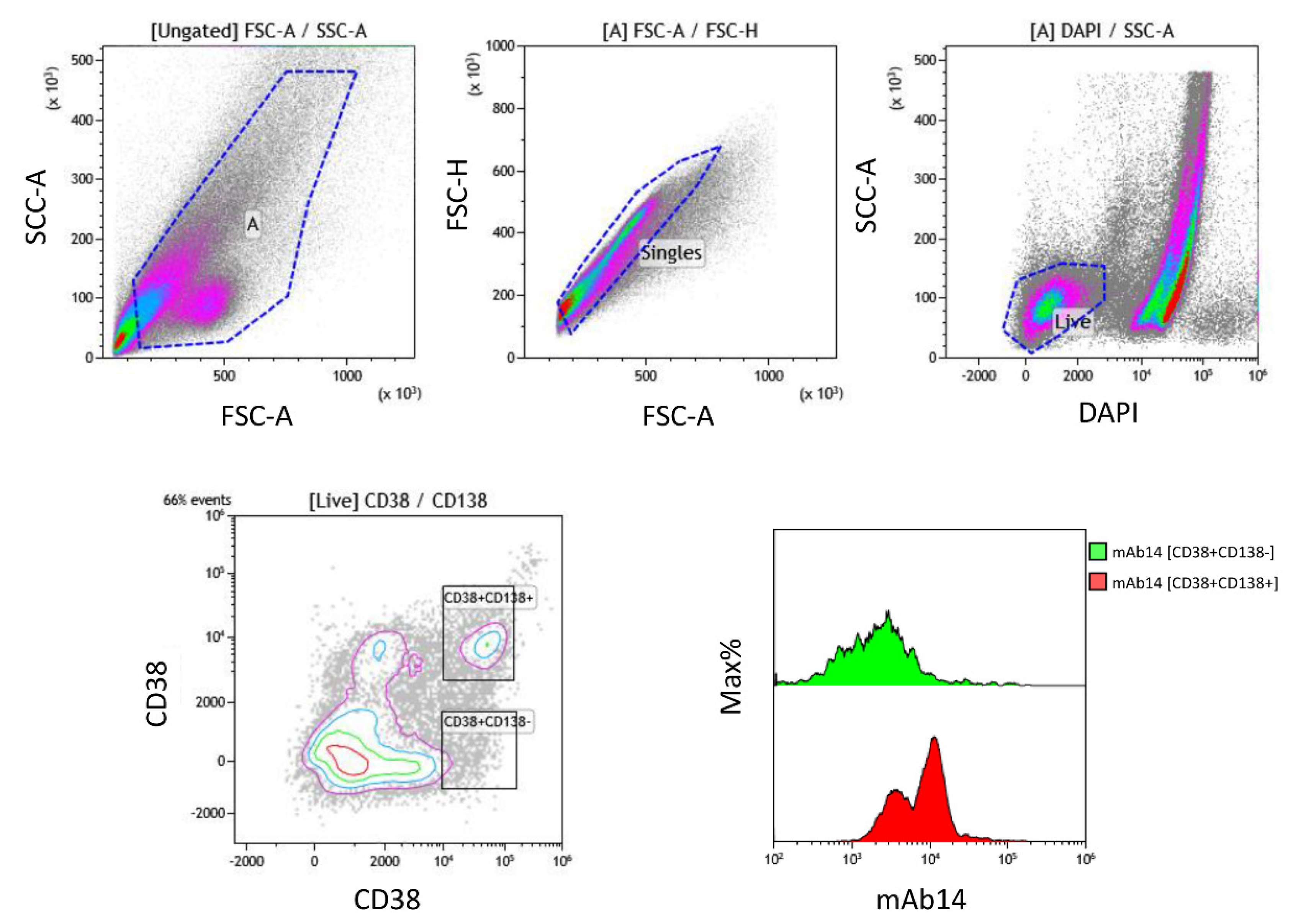
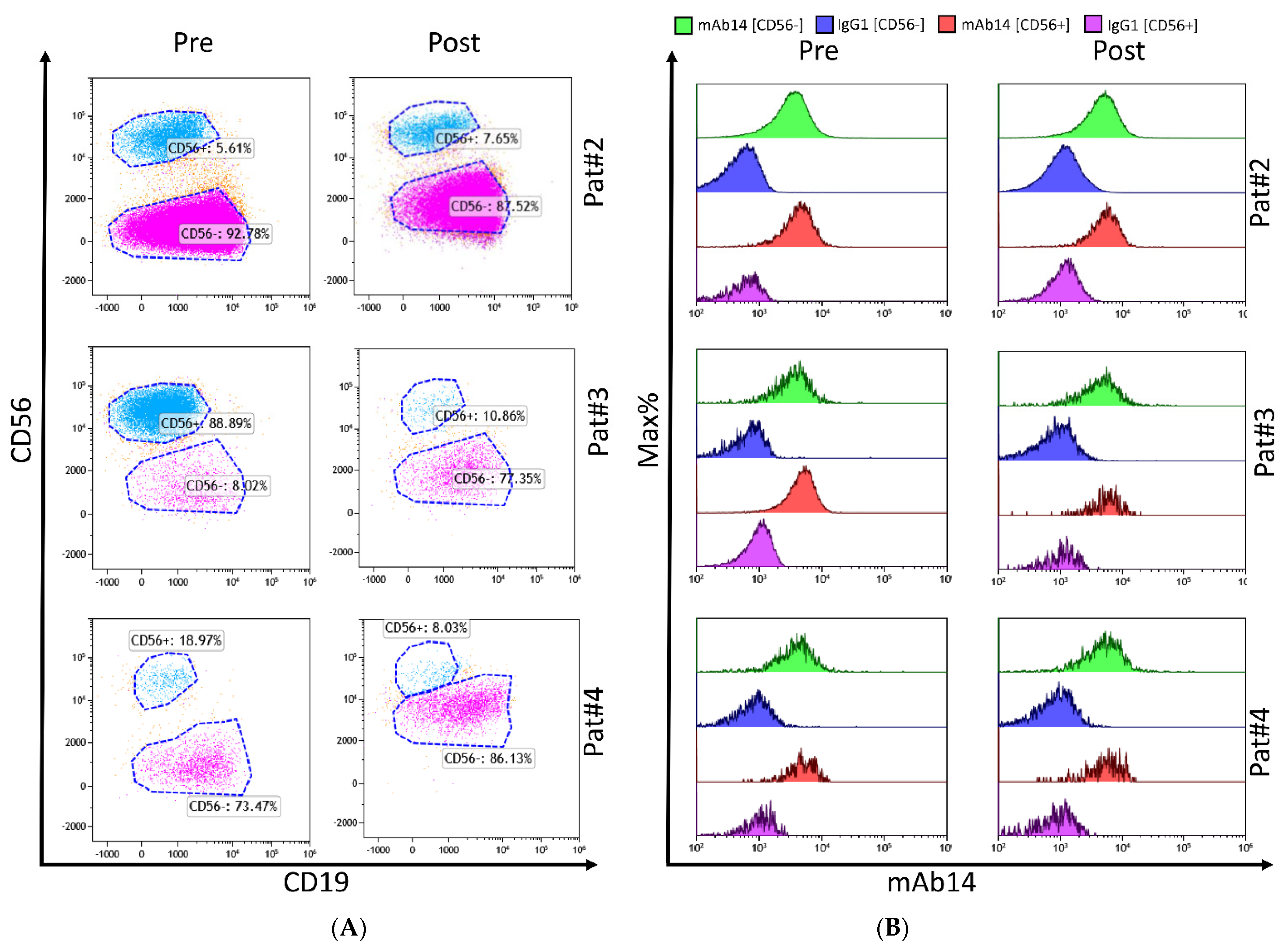
2.2. PCNA Is Expressed by All Various Plasma Cells from MM Bone Marrow
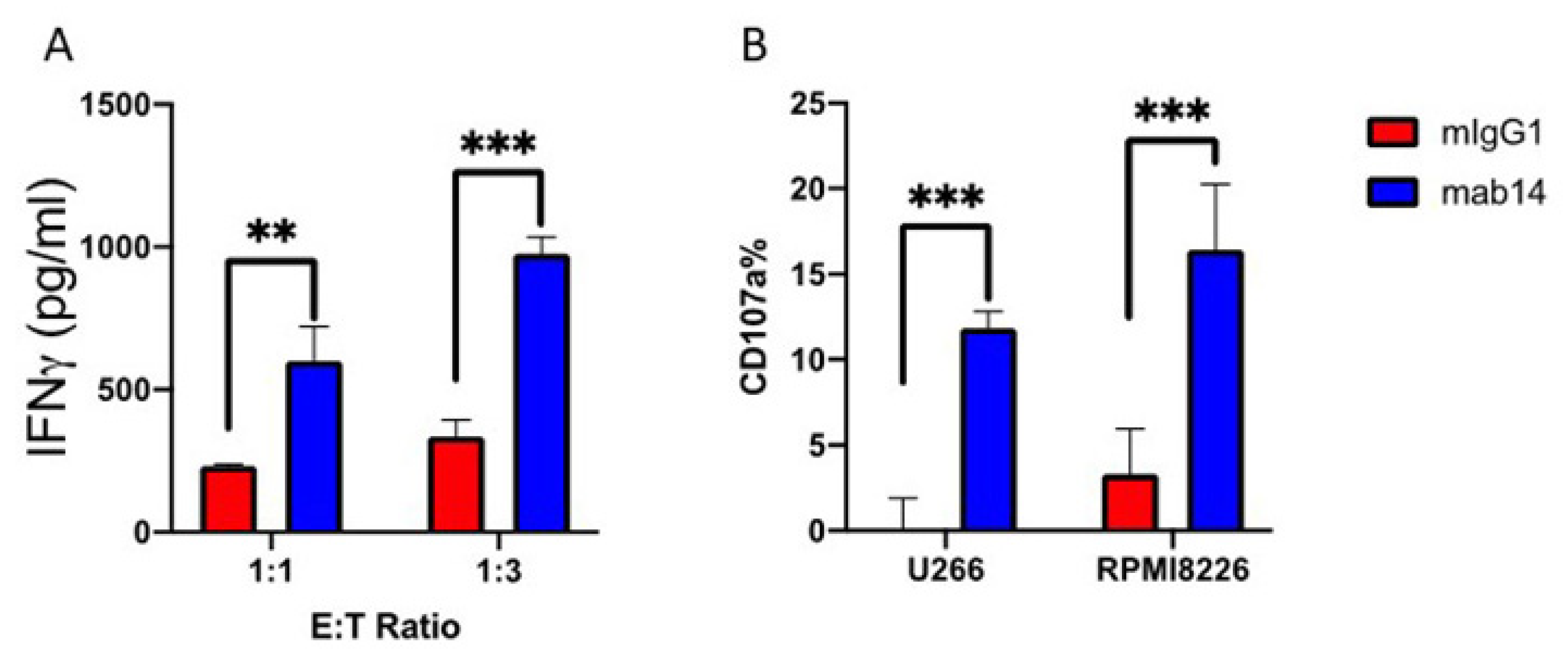
2.3. mAb14-25-9 Blocking of Membrane PCNA Can Enhance NK Cells Activities against Multiple Myeloma Cells
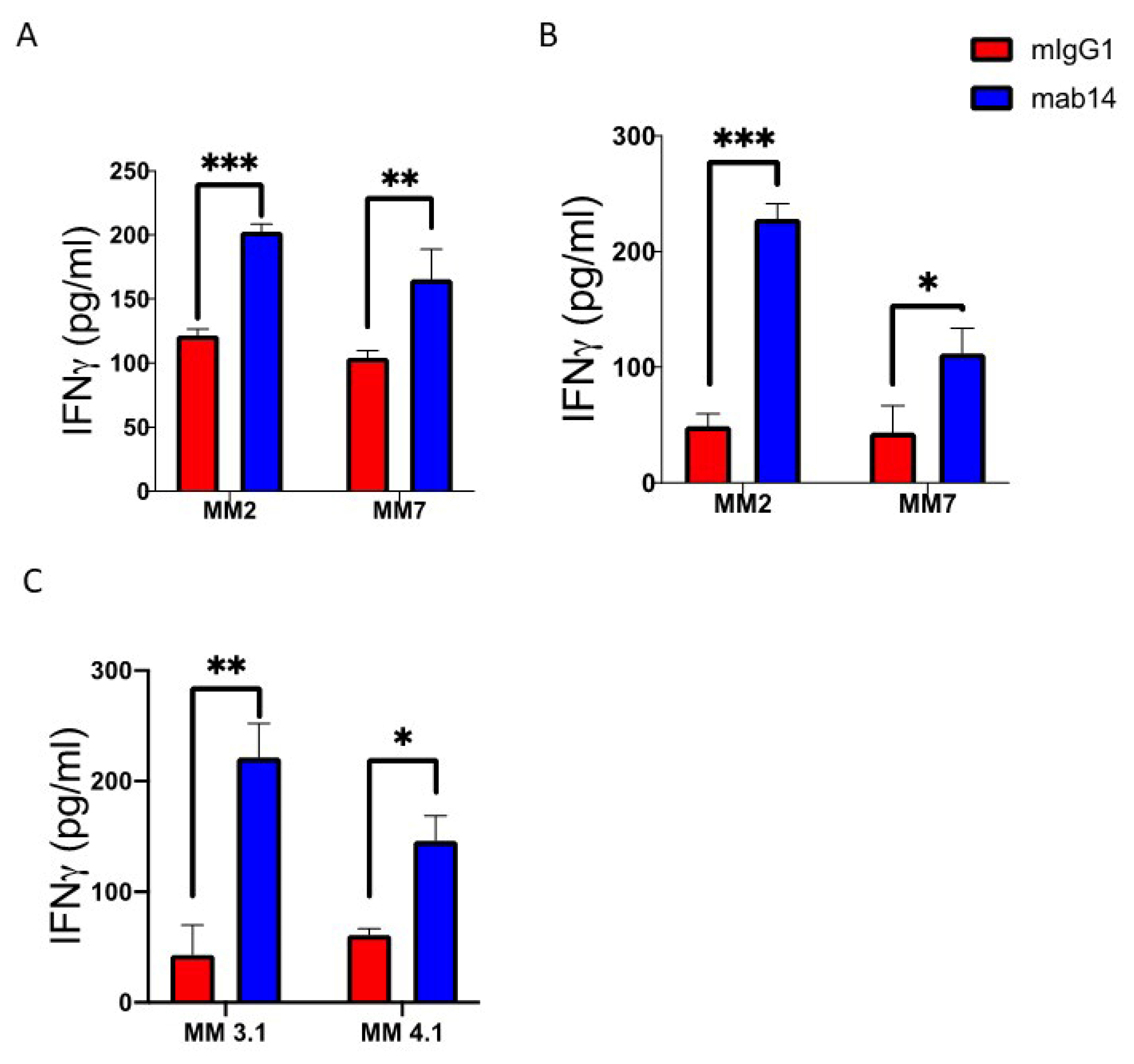
2.4. mAb 14-25-9 Enhances NK Cells Activities against Primary Multiple Myeloma Cells
3. Materials and Methods
3.1. Tissue Culture and Cells
3.2. Primary NK Cell Purification
3.3. Stimulation Assay and Measure of IFN-γ
3.4. NK Cell Degranulation CD107a Assays
3.5. Flow Cytometry
3.6. Primary Myeloma Samples
3.7. Statistical Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perussia, B.; Loza, M.J. Linear ‘2–0–1’Lymphocyte Development: Hypotheses on Cellular Bases for Immunity. Trends Immunol. 2003, 24, 235–241. [Google Scholar] [CrossRef]
- Moretta, A.; Marcenaro, E.; Parolini, S.; Ferlazzo, G.; Moretta, L. NK Cells at the Interface between Innate and Adaptive Immunity. Cell Death Differ. 2008, 15, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Martin, C.J.; Colonna, M. The Natural Cytotoxicity Receptors in Health and Disease. Front. Immunol. 2019, 10, 909. [Google Scholar] [CrossRef] [PubMed]
- Kruse, P.H.; Matta, J.; Ugolini, S.; Vivier, E. Natural Cytotoxicity Receptors and Their Ligands. Immunol. Cell Biol. 2014, 92, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a Novel Triggering Surface Molecule Specifically Expressed by Activated Natural Killer Cells, Is Involved in Non–Major Histocompatibility Complex–Restricted Tumor Cell Lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Sivori, S.; Vitale, M.; Morelli, L.; Sanseverino, L.; Augugliaro, R.; Bottino, C.; Moretta, L.; Moretta, A. P46, a Novel Natural Killer Cell–Specific Surface Molecule That Mediates Cell Activation. J. Exp. Med. 1997, 186, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Bottino, C.; Vitale, M.; Pende, D.; Cantoni, C.; Mingari, M.C.; Biassoni, R.; Moretta, L. Activating Receptors and Coreceptors Involved in Human Natural Killer Cell-Mediated Cytolysis. Annu. Rev. Immunol. 2001, 19, 197–223. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.W.T.; Campbell, K.S. Signal Transduction in Natural Killer Cells. Immunobiol. Nat. Kill. Cell Recept. 2006, 298, 23–57. [Google Scholar]
- Bottino, C.; Castriconi, R.; Moretta, L.; Moretta, A. Cellular Ligands of Activating NK Receptors. Trends Immunol. 2005, 26, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Arnon, T.I.; Achdout, H.; Lieberman, N.; Gazit, R.; Gonen-Gross, T.; Katz, G.; Bar-Ilan, A.; Bloushtain, N.; Lev, M.; Joseph, A. The Mechanisms Controlling the Recognition of Tumor-and Virus-Infected Cells by NKp46. Blood 2004, 103, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.; Hoffmann, S.C.; Jarahian, M.; Momburg, F.; Watzl, C. Expression Analysis of the Ligands for the Natural Killer Cell Receptors NKp30 and NKp44. PLoS ONE 2007, 2, e1339. [Google Scholar] [CrossRef] [PubMed]
- Jarahian, M.; Watzl, C.; Fournier, P.; Arnold, A.; Djandji, D.; Zahedi, S.; Cerwenka, A.; Paschen, A.; Schirrmacher, V.; Momburg, F. Activation of Natural Killer Cells by Newcastle Disease Virus Hemagglutinin-Neuraminidase. J. Virol. 2009, 83, 8108–8121. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Hershkovitz, O.; Peiris, M.; Zilka, A.; Bar-Ilan, A.; Nal, B.; Chu, K.; Kudelko, M.; Kam, Y.W.; Achdout, H. H5-Type Influenza Virus Hemagglutinin Is Functionally Recognized by the Natural Killer-Activating Receptor NKp44. J. Virol. 2008, 82, 2028–2032. [Google Scholar] [CrossRef] [PubMed]
- Mandelboim, O.; Lieberman, N.; Lev, M.; Paul, L.; Arnon, T.I.; Bushkin, Y.; Davis, D.M.; Strominger, J.L.; Yewdell, J.W.; Porgador, A. Recognition of Haemagglutinins on Virus-Infected Cells by NKp46 Activates Lysis by Human NK Cells. Nature 2001, 409, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Arnon, T.I.; Lev, M.; Katz, G.; Chernobrov, Y.; Porgador, A.; Mandelboim, O. Recognition of Viral Hemagglutinins by NKp44 but Not by NKp30. Eur. J. Immunol. 2001, 31, 2680–2689. [Google Scholar] [CrossRef]
- Ito, K.; Higai, K.; Sakurai, M.; Shinoda, C.; Yanai, K.; Azuma, Y.; Matsumoto, K. Binding of Natural Cytotoxicity Receptor NKp46 to Sulfate-and A2, 3-NeuAc-Containing Glycans and Its Mutagenesis. Biochem. Biophys. Res. Commun. 2011, 406, 377–382. [Google Scholar] [CrossRef]
- Bloushtain, N.; Qimron, U.; Bar-Ilan, A.; Hershkovitz, O.; Gazit, R.; Fima, E.; Korc, M.; Vlodavsky, I.; Bovin, N.; Porgador, A. Membrane-Associated Heparan Sulfate Proteoglycans Are Involved in the Recognition of Cellular Targets by NKp30 and NKp46. J. Immunol. 2004, 173, 2392–2401. [Google Scholar] [CrossRef] [PubMed]
- Hecht, M.-L.; Rosental, B.; Horlacher, T.; Hershkovitz, O.; de Paz, J.L.; Noti, C.; Schauer, S.; Porgador, A.; Seeberger, P.H. Natural Cytotoxicity Receptors NKp30, NKp44 and NKp46 Bind to Different Heparan Sulfate/Heparin Sequences. J. Proteome Res. 2009, 8, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, O.; Jarahian, M.; Zilka, A.; Bar-Ilan, A.; Landau, G.; Jivov, S.; Tekoah, Y.; Glicklis, R.; Gallagher, J.T.; Hoffmann, S.C. Altered Glycosylation of Recombinant NKp30 Hampers Binding to Heparan Sulfate: A Lesson for the Use of Recombinant Immunoreceptors as an Immunological Tool. Glycobiology 2008, 18, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, O.; Jivov, S.; Bloushtain, N.; Zilka, A.; Landau, G.; Bar-Ilan, A.; Lichtenstein, R.G.; Campbell, K.S.; van Kuppevelt, T.H.; Porgador, A. Characterization of the Recognition of Tumor Cells by the Natural Cytotoxicity Receptor, NKp44. Biochemistry 2007, 46, 7426–7436. [Google Scholar] [CrossRef] [PubMed]
- Jarahian, M.; Fiedler, M.; Cohnen, A.; Djandji, D.; Hämmerling, G.J.; Gati, C.; Cerwenka, A.; Turner, P.C.; Moyer, R.W.; Watzl, C. Modulation of NKp30-and NKp46-Mediated Natural Killer Cell Responses by Poxviral Hemagglutinin. PLoS Pathog. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; Shemesh, A.; Brusilovsky, M.; Porgador, A.; Campbell, K.S. Regulation of the Functions of Natural Cytotoxicity Receptors by Interactions with Diverse Ligands and Alterations in Splice Variant Expression. Front. Immunol. 2017, 8, 369. [Google Scholar] [CrossRef] [PubMed]
- Brandt, C.S.; Baratin, M.; Yi, E.C.; Kennedy, J.; Gao, Z.; Fox, B.; Haldeman, B.; Ostrander, C.D.; Kaifu, T.; Chabannon, C. The B7 Family Member B7-H6 Is a Tumor Cell Ligand for the Activating Natural Killer Cell Receptor NKp30 in Humans. J. Exp. Med. 2009, 206, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Arnon, T.I.; Achdout, H.; Levi, O.; Markel, G.; Saleh, N.; Katz, G.; Gazit, R.; Gonen-Gross, T.; Hanna, J.; Nahari, E. Inhibition of the NKp30 Activating Receptor by Pp65 of Human Cytomegalovirus. Nat. Immunol. 2005, 6, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Simhadri, V.R.; Reiners, K.S.; Hansen, H.P.; Topolar, D.; Simhadri, V.L.; Nohroudi, K.; Kufer, T.A.; Engert, A.; Pogge von Strandmann, E. Dendritic Cells Release HLA-B-Associated Transcript-3 Positive Exosomes to Regulate Natural Killer Function. PLoS ONE 2008, 3, e3377. [Google Scholar] [CrossRef] [PubMed]
- Rosental, B.; Brusilovsky, M.; Hadad, U.; Oz, D.; Appel, M.Y.; Afergan, F.; Yossef, R.; Rosenberg, L.A.; Aharoni, A.; Cerwenka, A. Proliferating Cell Nuclear Antigen Is a Novel Inhibitory Ligand for the Natural Cytotoxicity Receptor NKp44. J. Immunol. 2011, 187, 5693–5702. [Google Scholar] [CrossRef] [PubMed]
- Stoimenov, I.; Helleday, T. PCNA on the Crossroad of Cancer. Biochem. Soc. Trans. 2009, 37, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Stuart-Harris, R.; Caldas, C.; Pinder, S.E.; Pharoah, P. Proliferation Markers and Survival in Early Breast Cancer: A Systematic Review and Meta-Analysis of 85 Studies in 32,825 Patients. Breast 2008, 17, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, A.; Brusilovsky, M.; Hadad, U.; Teltsh, O.; Edri, A.; Rubin, E.; Campbell, K.S.; Rosental, B.; Porgador, A. Survival in Acute Myeloid Leukemia Is Associated with NKp44 Splice Variants. Oncotarget 2016, 7, 32933. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, A.; Brusilovsky, M.; Kundu, K.; Ottolenghi, A.; Campbell, K.S.; Porgador, A. Splice Variants of Human Natural Cytotoxicity Receptors: Novel Innate Immune Checkpoints. Cancer Immunol. Immunother. 2018, 67, 1871–1883. [Google Scholar] [CrossRef] [PubMed]
- Kundu, K.; Ghosh, S.; Sarkar, R.; Edri, A.; Brusilovsky, M.; Gershoni-Yahalom, O.; Yossef, R.; Shemesh, A.; Soria, J.-C.; Lazar, V. Inhibition of the NKp44-PCNA Immune Checkpoint Using a MAb to PCNA. Cancer Immunol. Res. 2019, 7, 1120–1134. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Prasad, M.; Kundu, K.; Cohen, L.; Yegodayev, K.M.; Zorea, J.; Joshua, B.-Z.; Lasry, B.; Dimitstein, O.; Bahat-Dinur, A. Tumor Tissue Explant Culture of Patient-Derived Xenograft as Potential Prioritization Tool for Targeted Therapy. Front. Oncol. 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, O.; Rosental, B.; Rosenberg, L.A.; Navarro-Sanchez, M.E.; Jivov, S.; Zilka, A.; Gershoni-Yahalom, O.; Brient-Litzler, E.; Bedouelle, H.; Ho, J.W. NKp44 Receptor Mediates Interaction of the Envelope Glycoproteins from the West Nile and Dengue Viruses with NK Cells. J. Immunol. 2009, 183, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Vieillard, V.; Strominger, J.L.; Debré, P. NK Cytotoxicity against CD4+ T Cells during HIV-1 Infection: A Gp41 Peptide Induces the Expression of an NKp44 Ligand. Proc. Natl. Acad. Sci. USA 2005, 102, 10981–10986. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Edeling, M.A.; Trifonov, V.; Luo, J.; Goyal, P.; Bohl, B.; Bando, J.K.; Kim, A.H.; Walker, J.; Andahazy, M. Natural Killer Cells Control Tumor Growth by Sensing a Growth Factor. Cell 2018, 172, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Anderson, K. Multiple Myeloma. N. Engl. J. Med. 2011, 364, 1046–1060. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Leleu, X.; Attal, M.; Arnulf, B.; Moreau, P.; Traulle, C.; Marit, G.; Mathiot, C.; Petillon, M.O.; Macro, M.; Roussel, M. Pomalidomide plus Low-Dose Dexamethasone Is Active and Well Tolerated in Bortezomib and Lenalidomide–Refractory Multiple Myeloma: Intergroupe Francophone Du Myélome 2009-02. Blood J. Am. Soc. Hematol. 2013, 121, 1968–1975. [Google Scholar] [CrossRef] [PubMed]
- Facon, T.; Lee, J.H.; Moreau, P.; Niesvizky, R.; Dimopoulos, M.; Hajek, R.; Pour, L.; Jurczyszyn, A.; Qiu, L.; Klippel, Z. Carfilzomib or Bortezomib with Melphalan-Prednisone for Transplant-Ineligible Patients with Newly Diagnosed Multiple Myeloma. Blood J. Am. Soc. Hematol. 2019, 133, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Afifi, S.; Michael, A.; Lesokhin, A. Immunotherapy: A New Approach to Treating Multiple Myeloma with Daratumumab and Elotuzumab. Ann. Pharmacother. 2016, 50, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Mahindra, A.; Laubach, J.; Raje, N.; Munshi, N.; Richardson, P.G.; Anderson, K. Latest Advances and Current Challenges in the Treatment of Multiple Myeloma. Nat. Rev. Clin. Oncol. 2012, 9, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, I.H.; Sergeant, R.; Szydlo, R.; Apperley, J.F.; DeLavallade, H.; Alsuliman, A.; Khoder, A.; Marin, D.; Kanfer, E.; Cooper, N. Interaction between KIR3DS1 and HLA-Bw4 Predicts for Progression-Free Survival after Autologous Stem Cell Transplantation in Patients with Multiple Myeloma. Blood J. Am. Soc. Hematol. 2010, 116, 2033–2039. [Google Scholar] [CrossRef] [PubMed]
- Kröger, N.; Shaw, B.; Iacobelli, S.; Zabelina, T.; Peggs, K.; Shimoni, A.; Nagler, A.; Binder, T.; Eiermann, T.; Madrigal, A. Comparison between Antithymocyte Globulin and Alemtuzumab and the Possible Impact of KIR-ligand Mismatch after Dose-reduced Conditioning and Unrelated Stem Cell Transplantation in Patients with Multiple Myeloma. Br. J. Haematol. 2005, 129, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.-T.; Dillon, M.; Song, W.; Leiba, M.; Li, X.-F.; Burger, P.; Lee, A.I.; Podar, K.; Hideshima, T.; Rice, A.G. Anti-CS1 Humanized Monoclonal Antibody HuLuc63 Inhibits Myeloma Cell Adhesion and Induces Antibody-Dependent Cellular Cytotoxicity in the Bone Marrow Milieu. Blood J. Am. Soc. Hematol. 2008, 112, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Hsi, E.D.; Steinle, R.; Balasa, B.; Szmania, S.; Draksharapu, A.; Shum, B.P.; Huseni, M.; Powers, D.; Nanisetti, A.; Zhang, Y. CS1, a Potential New Therapeutic Antibody Target for the Treatment of Multiple Myeloma. Clin. Cancer Res. 2008, 14, 2775–2784. [Google Scholar] [CrossRef]
- El-Sherbiny, Y.M.; Meade, J.L.; Holmes, T.D.; McGonagle, D.; Mackie, S.L.; Morgan, A.W.; Cook, G.; Feyler, S.; Richards, S.J.; Davies, F.E. The Requirement for DNAM-1, NKG2D, and NKp46 in the Natural Killer Cell-Mediated Killing of Myeloma Cells. Cancer Res. 2007, 67, 8444–8449. [Google Scholar] [CrossRef]
- Carbone, E.; Neri, P.; Mesuraca, M.; Fulciniti, M.T.; Otsuki, T.; Pende, D.; Groh, V.; Spies, T.; Pollio, G.; Cosman, D. HLA Class I, NKG2D, and Natural Cytotoxicity Receptors Regulate Multiple Myeloma Cell Recognition by Natural Killer Cells. Blood 2005, 105, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Soriani, A.; Zingoni, A.; Cerboni, C.; Iannitto, M.L.; Ricciardi, M.R.; di Gialleonardo, V.; Cippitelli, M.; Fionda, C.; Petrucci, M.T.; Guarini, A. ATM-ATR–Dependent up-Regulation of DNAM-1 and NKG2D Ligands on Multiple Myeloma Cells by Therapeutic Agents Results in Enhanced NK-Cell Susceptibility and Is Associated with a Senescent Phenotype. Blood J. Am. Soc. Hematol. 2009, 113, 3503–3511. [Google Scholar] [CrossRef]
- Bernal, M.; Garrido, P.; Jiménez, P.; Carretero, R.; Almagro, M.; López, P.; Navarro, P.; Garrido, F.; Ruiz-Cabello, F. Changes in Activatory and Inhibitory Natural Killer (NK) Receptors May Induce Progression to Multiple Myeloma: Implications for Tumor Evasion of T and NK Cells. Hum. Immunol. 2009, 70, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Fauriat, C.; Mallet, F.; Olive, D. Impaired Activating Receptor Expression Pattern in Natural Killer Cells from Patients with Multiple Myeloma. Leukemia 2006, 20, 732–733. [Google Scholar] [CrossRef] [PubMed]
- Benson Jr, D.M.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K. The PD-1/PD-L1 Axis Modulates the Natural Killer Cell versus Multiple Myeloma Effect: A Therapeutic Target for CT-011, a Novel Monoclonal Anti–PD-1 Antibody. Blood J. Am. Soc. Hematol. 2010, 116, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Bezman, N.A.; Jhatakia, A.; Kearney, A.Y.; Brender, T.; Maurer, M.; Henning, K.; Jenkins, M.R.; Rogers, A.J.; Neeson, P.J.; Korman, A.J. PD-1 Blockade Enhances Elotuzumab Efficacy in Mouse Tumor Models. Blood Adv. 2017, 1, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; Nakamura, K.; Vuckovic, S.; Hill, G.R.; Smyth, M.J. Immune Responses in Multiple Myeloma: Role of the Natural Immune Surveillance and Potential of Immunotherapies. Cell. Mol. Life Sci. 2016, 73, 1569–1589. [Google Scholar] [CrossRef] [PubMed]
- Pittari, G.; Vago, L.; Festuccia, M.; Bonini, C.; Mudawi, D.; Giaccone, L.; Bruno, B. Restoring Natural Killer Cell Immunity against Multiple Myeloma in the Era of New Drugs. Front. Immunol. 2017, 8, 1444. [Google Scholar] [CrossRef]
- Fionda, C.; Abruzzese, M.P.; Zingoni, A.; Cecere, F.; Vulpis, E.; Peruzzi, G.; Soriani, A.; Molfetta, R.; Paolini, R.; Ricciardi, M.R. The IMiDs Targets IKZF-1/3 and IRF4 as Novel Negative Regulators of NK Cell-Activating Ligands Expression in Multiple Myeloma. Oncotarget 2015, 6, 23609. [Google Scholar] [CrossRef] [PubMed]
- James, A.M.; Cohen, A.D.; Campbell, K.S. Combination Immune Therapies to Enhance Anti-Tumor Responses by NK Cells. Front. Immunol. 2013, 4, 481. [Google Scholar]
- Hideshima, T.; Chauhan, D.; Shima, Y.; Raje, N.; Davies, F.E.; Tai, Y.-T.; Treon, S.P.; Lin, B.; Schlossman, R.L.; Richardson, P. Thalidomide and Its Analogs Overcome Drug Resistance of Human Multiple Myeloma Cells to Conventional Therapy. Blood J. Am. Soc. Hematol. 2000, 96, 2943–2950. [Google Scholar]
- Fionda, C.; Soriani, A.; Malgarini, G.; Iannitto, M.L.; Santoni, A.; Cippitelli, M. Heat Shock Protein-90 Inhibitors Increase MHC Class I-Related Chain A and B Ligand Expression on Multiple Myeloma Cells and Their Ability to Trigger NK Cell Degranulation. J. Immunol. 2009, 183, 4385–4394. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Tricot, G.J.; Garg, T.K.; Malaviarachchi, P.A.; Szmania, S.M.; Kellum, R.E.; Storrie, B.; Mulder, A.; Shaughnessy, J.D., Jr.; Barlogie, B. Bortezomib Down-Regulates the Cell-Surface Expression of HLA Class I and Enhances Natural Killer Cell–Mediated Lysis of Myeloma. Blood J. Am. Soc. Hematol. 2008, 111, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Antonangeli, F.; Soriani, A.; Ricci, B.; Ponzetta, A.; Benigni, G.; Morrone, S.; Bernardini, G.; Santoni, A. Natural Killer Cell Recognition of in Vivo Drug-Induced Senescent Multiple Myeloma Cells. Oncoimmunology 2016, 5, e1218105. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, C.; Ricci, B.; Vulpis, E.; Fionda, C.; Ricciardi, M.R.; Petrucci, M.T.; Masuelli, L.; Peri, A.; Cippitelli, M.; Zingoni, A. Drug-Induced Senescent Multiple Myeloma Cells Elicit NK Cell Proliferation by Direct or Exosome-Mediated IL15 Trans-Presentation. Cancer Immunol. Res. 2018, 6, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Goding Sauer, A.; Ortiz, A.P.; Fedewa, S.A.; Pinheiro, P.S.; Tortolero-Luna, G.; Martinez-Tyson, D.; Jemal, A.; Siegel, R.L. Cancer Statistics for Hispanics/Latinos, 2018. CA Cancer J. Clin. 2018, 68, 425–445. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.G.; Lonial, S.; Jakubowiak, A.J.; Harousseau, J.; Anderson, K.C. Monoclonal Antibodies in the Treatment of Multiple Myeloma. Br. J. Haematol. 2011, 154, 745–754. [Google Scholar] [CrossRef]
- Bonello, F.; Mina, R.; Boccadoro, M.; Gay, F. Therapeutic Monoclonal Antibodies and Antibody Products: Current Practices and Development in Multiple Myeloma. Cancers 2020, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Weiner, L.M.; Surana, R.; Wang, S. Monoclonal Antibodies: Versatile Platforms for Cancer Immunotherapy. Nat. Rev. Immunol. 2010, 10, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.; Gupta, P.; Chen, X.; Cardillo, T.M.; Furman, R.R.; Chen, S.; Chang, C.-H.; Goldenberg, D.M. Therapy of B-Cell Malignancies by Anti–HLA-DR Humanized Monoclonal Antibody, IMMU-114, Is Mediated through Hyperactivation of ERK and JNK MAP Kinase Signaling Pathways. Blood J. Am. Soc. Hematol. 2010, 115, 5180–5190. [Google Scholar] [CrossRef] [PubMed]
- Laubach, J.P.; Richardson, P.G. CD38-Targeted Immunochemotherapy in Refractory Multiple Myeloma: A New Horizon. Clin. Cancer Res. 2015, 21, 2660–2662. [Google Scholar] [CrossRef]
- Palumbo, A.; Sonneveld, P. Preclinical and Clinical Evaluation of Elotuzumab, a SLAMF7-Targeted Humanized Monoclonal Antibody in Development for Multiple Myeloma. Expert Rev. Hematol. 2015, 8, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Tassone, P.; Goldmacher, V.S.; Neri, P.; Gozzini, A.; Shammas, M.A.; Whiteman, K.R.; Hylander-Gans, L.L.; Carrasco, D.R.; Hideshima, T.; Shringarpure, R. Cytotoxic Activity of the Maytansinoid Immunoconjugate B-B4–DM1 against CD138+ Multiple Myeloma Cells. Blood 2004, 104, 3688–3696. [Google Scholar] [CrossRef] [PubMed]
- Ocio, E.M.; Richardson, P.G.; Rajkumar, S.V.; Palumbo, A.; Mateos, M.V.; Orlowski, R.; Kumar, S.; Usmani, S.; Roodman, D.; Niesvizky, R. New Drugs and Novel Mechanisms of Action in Multiple Myeloma in 2013: A Report from the International Myeloma Working Group (IMWG). Leukemia 2014, 28, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Kretz-Rommel, A.; Qin, F.; Dakappagari, N.; Cofiell, R.; Faas, S.J.; Bowdish, K.S. Blockade of CD200 in the Presence or Absence of Antibody Effector Function: Implications for Anti-CD200 Therapy. J. Immunol. 2008, 180, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Bensinger, W.; Maziarz, R.T.; Jagannath, S.; Spencer, A.; Durrant, S.; Becker, P.S.; Ewald, B.; Bilic, S.; Rediske, J.; Baeck, J. A Phase 1 Study of Lucatumumab, a Fully Human Anti-CD 40 Antagonist Monoclonal Antibody Administered Intravenously to Patients with Relapsed or Refractory Multiple Myeloma. Br. J. Haematol. 2012, 159, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.-T.; Mayes, P.A.; Acharya, C.; Zhong, M.Y.; Cea, M.; Cagnetta, A.; Craigen, J.; Yates, J.; Gliddon, L.; Fieles, W. Novel Anti–B-Cell Maturation Antigen Antibody-Drug Conjugate (GSK2857916) Selectively Induces Killing of Multiple Myeloma. Blood J. Am. Soc. Hematol. 2014, 123, 3128–3138. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Kumar, S.K.; Orlowski, R.Z.; Cook, G.; Richardson, P.G.; Gertz, M.A.; Giralt, S.; Mateos, M.; Leleu, X.; Anderson, K.C. Management of Relapsed and Refractory Multiple Myeloma: Novel Agents, Antibodies, Immunotherapies and Beyond. Leukemia 2018, 32, 252–262. [Google Scholar] [CrossRef]
- Pazina, T.; James, A.M.; Colby, K.B.; Yang, Y.; Gale, A.; Jhatakia, A.; Kearney, A.Y.; Graziano, R.F.; Bezman, N.A.; Robbins, M.D. Enhanced SLAMF7 Homotypic Interactions by Elotuzumab Improves NK Cell Killing of Multiple Myeloma. Cancer Immunol. Res. 2019, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.S.; Cohen, A.D.; Pazina, T. Mechanisms of NK Cell Activation and Clinical Activity of the Therapeutic SLAMF7 Antibody, Elotuzumab in Multiple Myeloma. Front. Immunol. 2018, 9, 2551. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; James, A.M.; Jhatakia, A.; Graziano, R.F.; Bezman, N.A.; Robbins, M.D.; Cohen, A.D.; Campbell, K.S. Elotuzumab Promotes Self-Engagement of Signaling Lymphocytic Activation Molecule Family Member 7 (SLAMF7) Between Natural Killer (NK) and Multiple Myeloma (MM) Cells to Enhance Cytotoxicity. Clin. Lymphoma Myeloma Leuk. 2018, 18, S253–S254. [Google Scholar] [CrossRef]
- Horton, N.C.; Mathew, S.O.; Mathew, P.A. Novel Interaction between Proliferating Cell Nuclear Antigen and HLA I on the Surface of Tumor Cells Inhibits NK Cell Function through NKp44. PLoS ONE 2013, 8, e59552. [Google Scholar] [CrossRef] [PubMed]
- Edri, A.; Shemesh, A.; Iraqi, M.; Matalon, O.; Brusilovsky, M.; Hadad, U.; Radinsky, O.; Gershoni-Yahalom, O.; Dye, J.M.; Mandelboim, O. The Ebola-Glycoprotein Modulates the Function of Natural Killer Cells. Front. Immunol. 2018, 9, 1428. [Google Scholar] [CrossRef] [PubMed]
- Pratt, G.; Goodyear, O.; Moss, P. Immunodeficiency and Immunotherapy in Multiple Myeloma. Br. J. Haematol. 2007, 138, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Smyth, M.J.; Martinet, L. Cancer Immunoediting and Immune Dysregulation in Multiple Myeloma. Blood 2020, 136, 2731–2740. [Google Scholar] [CrossRef] [PubMed]
- Agata, Y.; Kawasaki, A.; Nishimura, H.; Ishida, Y.; Tsubat, T.; Yagita, H.; Honjo, T. Expression of the PD-1 Antigen on the Surface of Stimulated Mouse T and B Lymphocytes. Int. Immunol. 1996, 8, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Dosani, T.; Carlsten, M.; Maric, I.; Landgren, O. The Cellular Immune System in Myelomagenesis: NK Cells and T Cells in the Development of MM and Their Uses in Immunotherapies. Blood Cancer J. 2015, 5, e306. [Google Scholar] [CrossRef] [PubMed]
- Mahaweni, N.M.; Ehlers, F.A.I.; Bos, G.M.J.; Wieten, L. Tuning Natural Killer Cell Anti-Multiple Myeloma Reactivity by Targeting Inhibitory Signaling via KIR and NKG2A. Front. Immunol. 2018, 9, 2848. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M. NK Cells: New Issues and Challenges. Eur. J. Immunol. 2008, 38, 2927–2929. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; MacFarlane, A.W.; Bernabei, L.; Dulaimi, E.; Kotcher, R.; Yam, C.; Bezman, N.A.; Robbins, M.D.; Ross, E.A.; Campbell, K.S. Alterations of NK Cell Phenotype in the Disease Course of Multiple Myeloma. Cancers 2021, 13, 226. [Google Scholar] [CrossRef] [PubMed]
- Keats, J.J.; Chesi, M.; Egan, J.B.; Garbitt, V.M.; Palmer, S.E.; Braggio, E.; van Wier, S.; Blackburn, P.R.; Baker, A.S.; Dispenzieri, A. Clonal Competition with Alternating Dominance in Multiple Myeloma. Blood J. Am. Soc. Hematol. 2012, 120, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Leth-Larsen, R.; Lund, R.; Hansen, H.; Laenkholm, A.-V.; Tarin, D.; Jensen, O.N.; Ditzel, H.J. Metastasis-Related Plasma Membrane Proteins of Human Breast Cancer Cells Identified by Comparative Quantitative Mass Spectrometry. Mol. Cell. Proteom. 2009, 8, 1436–1449. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Pietsch, J.; Infanger, M.; Eucker, J.; Eilles, C.; Schoenberger, J. Diagnostic and Therapeutic Use of Membrane Proteins in Cancer Cells. Curr. Med. Chem. 2011, 18, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Kampen, K.R. Membrane Proteins: The Key Players of a Cancer Cell. J. Membr. Biol. 2011, 242, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zheng, H.; Liu, H.; Li, M.; Ren, W.; Liao, W.; Duan, Z.; Li, L.; Cao, Y. Immunoglobulin Expression and Its Biological Significance in Cancer Cells. Cell Mol. Immunol. 2008, 5, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Malenka, D.J.; Rogan, K.M.; Howe, R.C.; Nelson, R.S.; Wrobel, C.J.; Ahmed, A.; Humphreys, R.E. Variable Expression of Membrane Proteins among Murine Lymphoblastoid Tumors as Seen with Sodium Dodecyl Sulfate-Polyacrylamide Gradient Gel Electrophoresis of [35S] Methionine-Labeled Cell Membranes. Cancer Res. 1979, 39, 4782–4790. [Google Scholar]
- Zoellner, H.; Chami, B.; Kelly, E.; Moore, M.A.S. Increased Cell Size, Structural Complexity and Migration of Cancer Cells Acquiring Fibroblast Organelles by Cell-Projection Pumping. PLoS ONE 2019, 14, e0224800. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iraqi, M.; Edri, A.; Greenshpan, Y.; Goldstein, O.; Ofir, N.; Bolel, P.; Abu Ahmad, M.; Zektser, M.; Campbell, K.S.; Rouvio, O.; et al. Blocking the PCNA/NKp44 Checkpoint to Stimulate NK Cell Responses to Multiple Myeloma. Int. J. Mol. Sci. 2022, 23, 4717. https://doi.org/10.3390/ijms23094717
Iraqi M, Edri A, Greenshpan Y, Goldstein O, Ofir N, Bolel P, Abu Ahmad M, Zektser M, Campbell KS, Rouvio O, et al. Blocking the PCNA/NKp44 Checkpoint to Stimulate NK Cell Responses to Multiple Myeloma. International Journal of Molecular Sciences. 2022; 23(9):4717. https://doi.org/10.3390/ijms23094717
Chicago/Turabian StyleIraqi, Muhammed, Avishay Edri, Yariv Greenshpan, Oron Goldstein, Noa Ofir, Priyanka Bolel, Muhammad Abu Ahmad, Miri Zektser, Kerry S. Campbell, Ory Rouvio, and et al. 2022. "Blocking the PCNA/NKp44 Checkpoint to Stimulate NK Cell Responses to Multiple Myeloma" International Journal of Molecular Sciences 23, no. 9: 4717. https://doi.org/10.3390/ijms23094717
APA StyleIraqi, M., Edri, A., Greenshpan, Y., Goldstein, O., Ofir, N., Bolel, P., Abu Ahmad, M., Zektser, M., Campbell, K. S., Rouvio, O., Gazit, R., & Porgador, A. (2022). Blocking the PCNA/NKp44 Checkpoint to Stimulate NK Cell Responses to Multiple Myeloma. International Journal of Molecular Sciences, 23(9), 4717. https://doi.org/10.3390/ijms23094717