
Definition of IgG Subclass-Specific Glycopatterns in Idiopathic Membranous Nephropathy: Aberrant IgG Glycoforms in Blood

 , , , , , , , and
, , , , , , , and
Abstract
: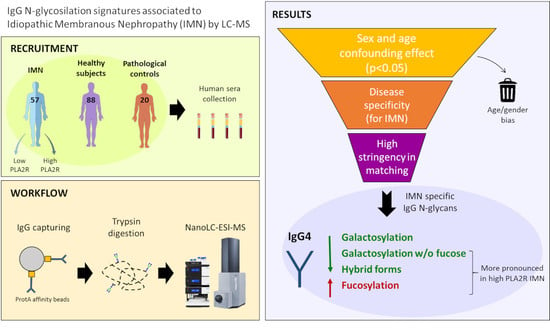
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
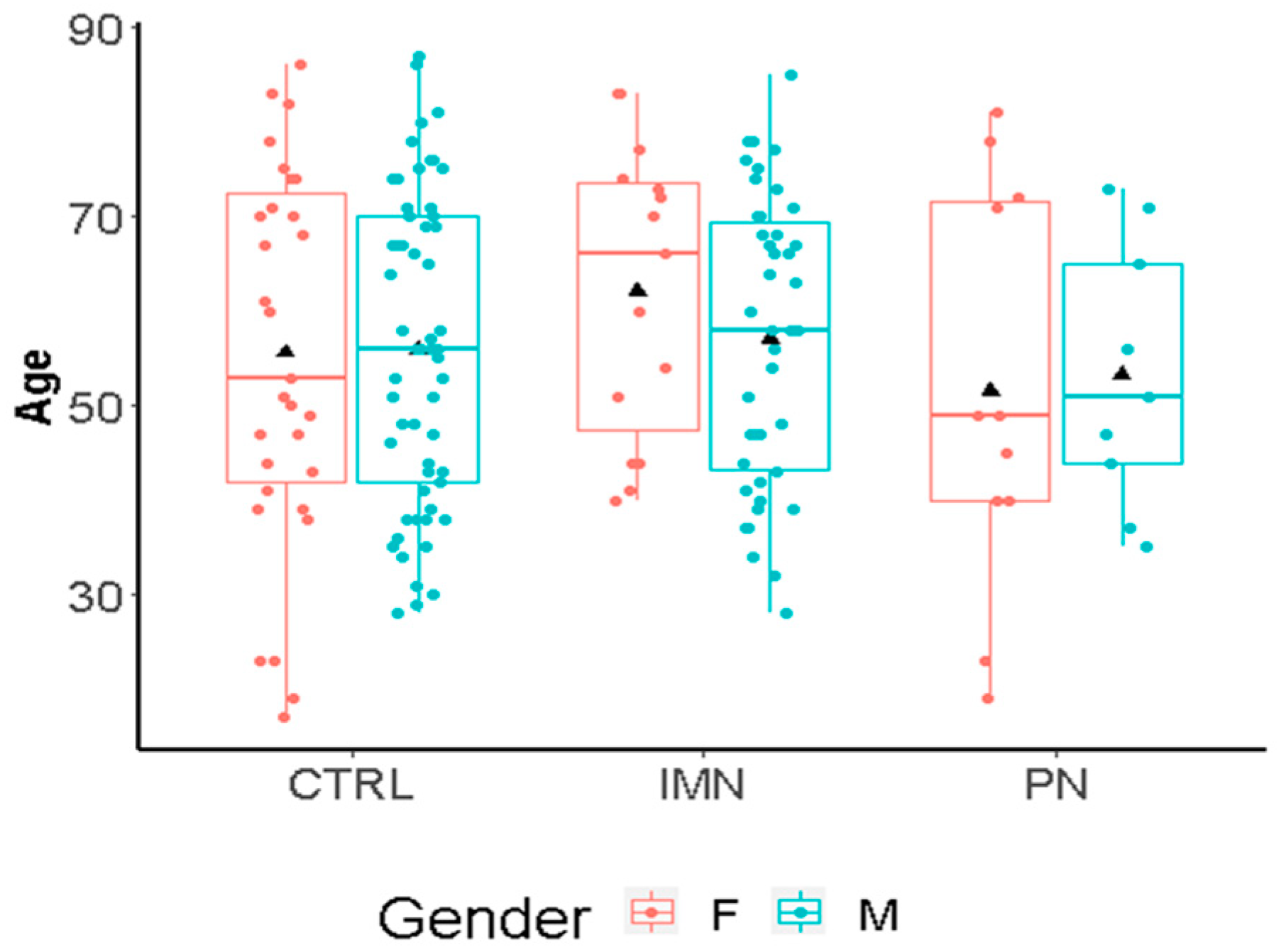
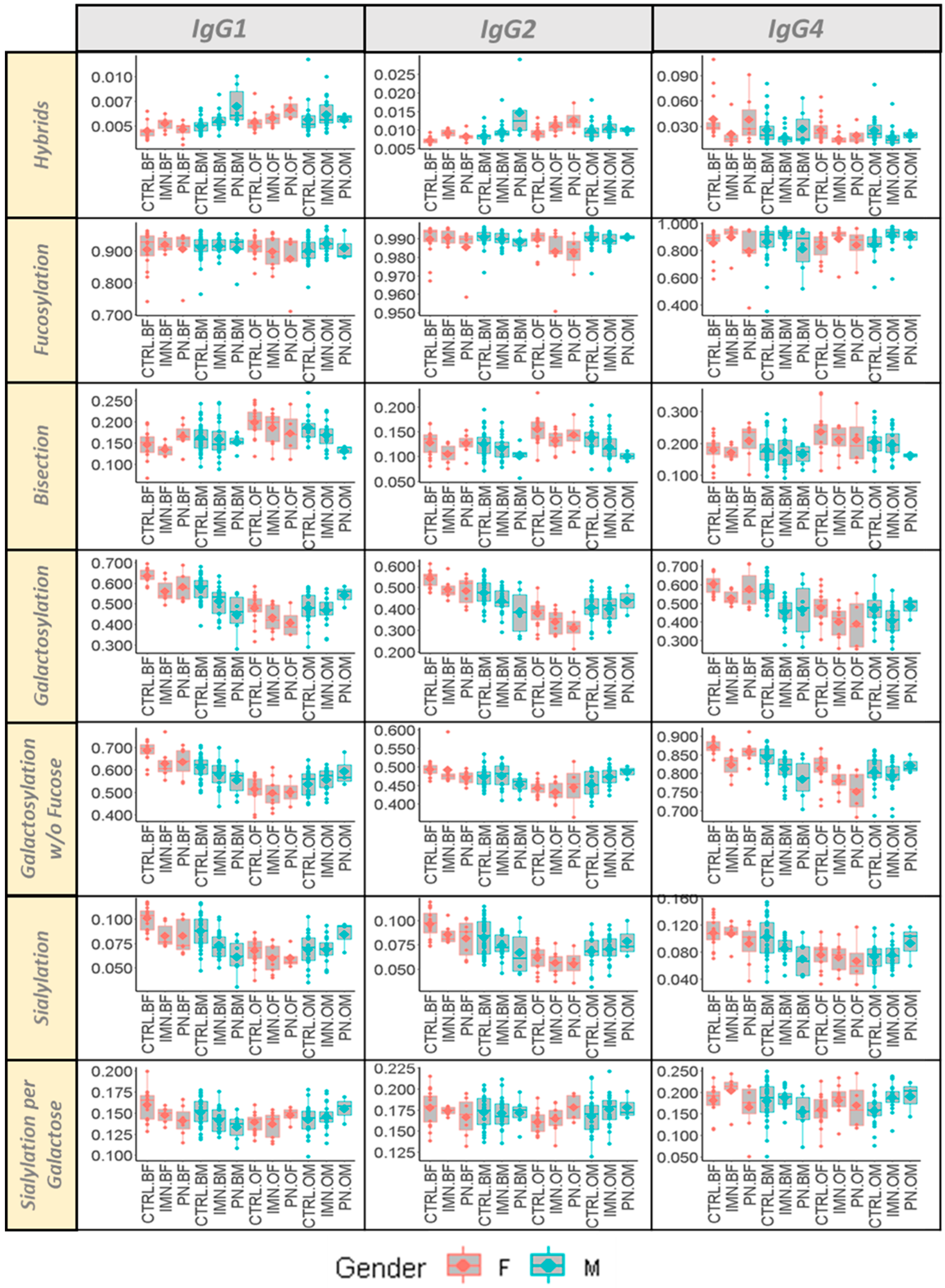
2.1. Age and Sex Effect on IgG Fc Glycosylation in the Cohort
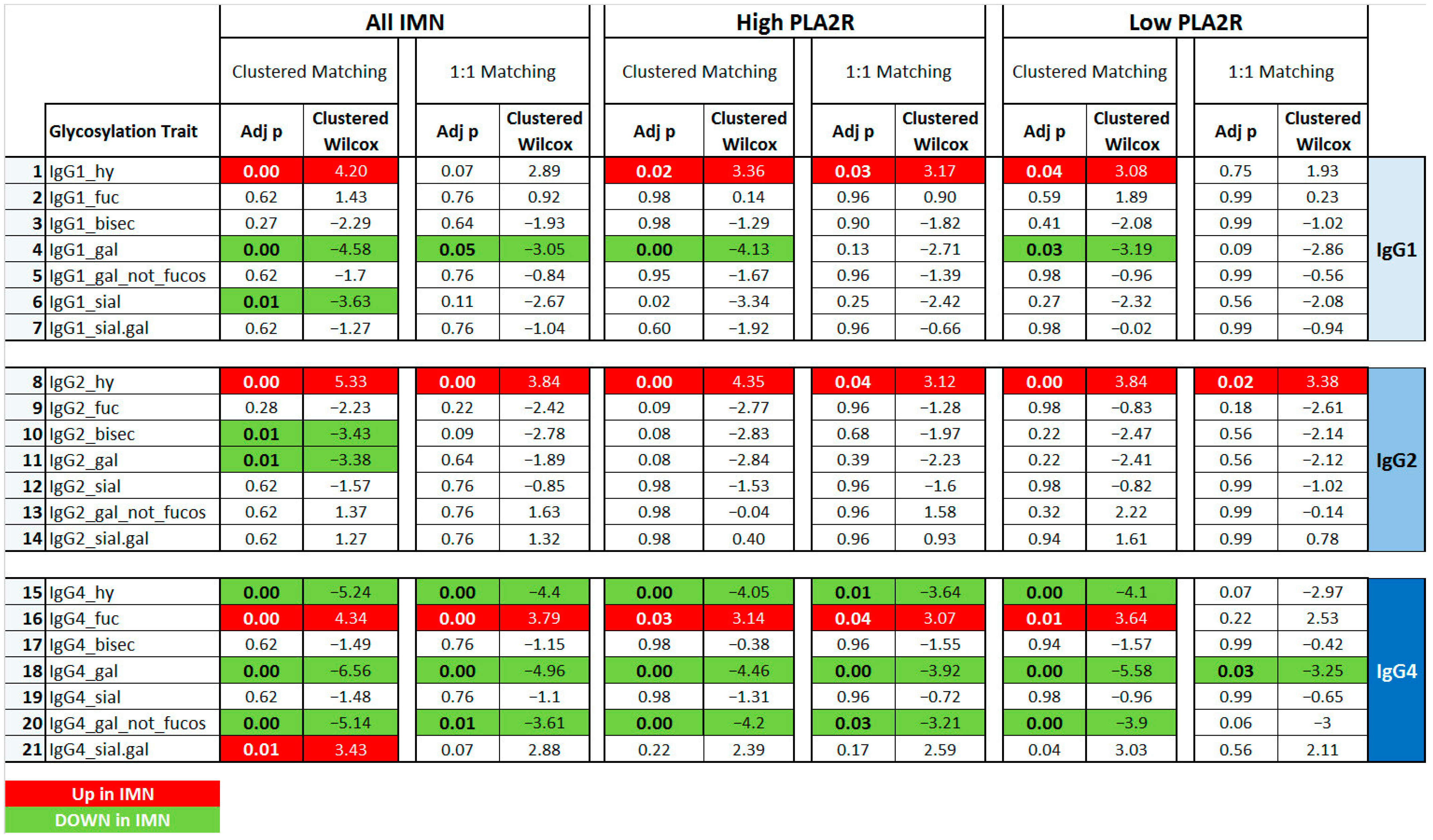
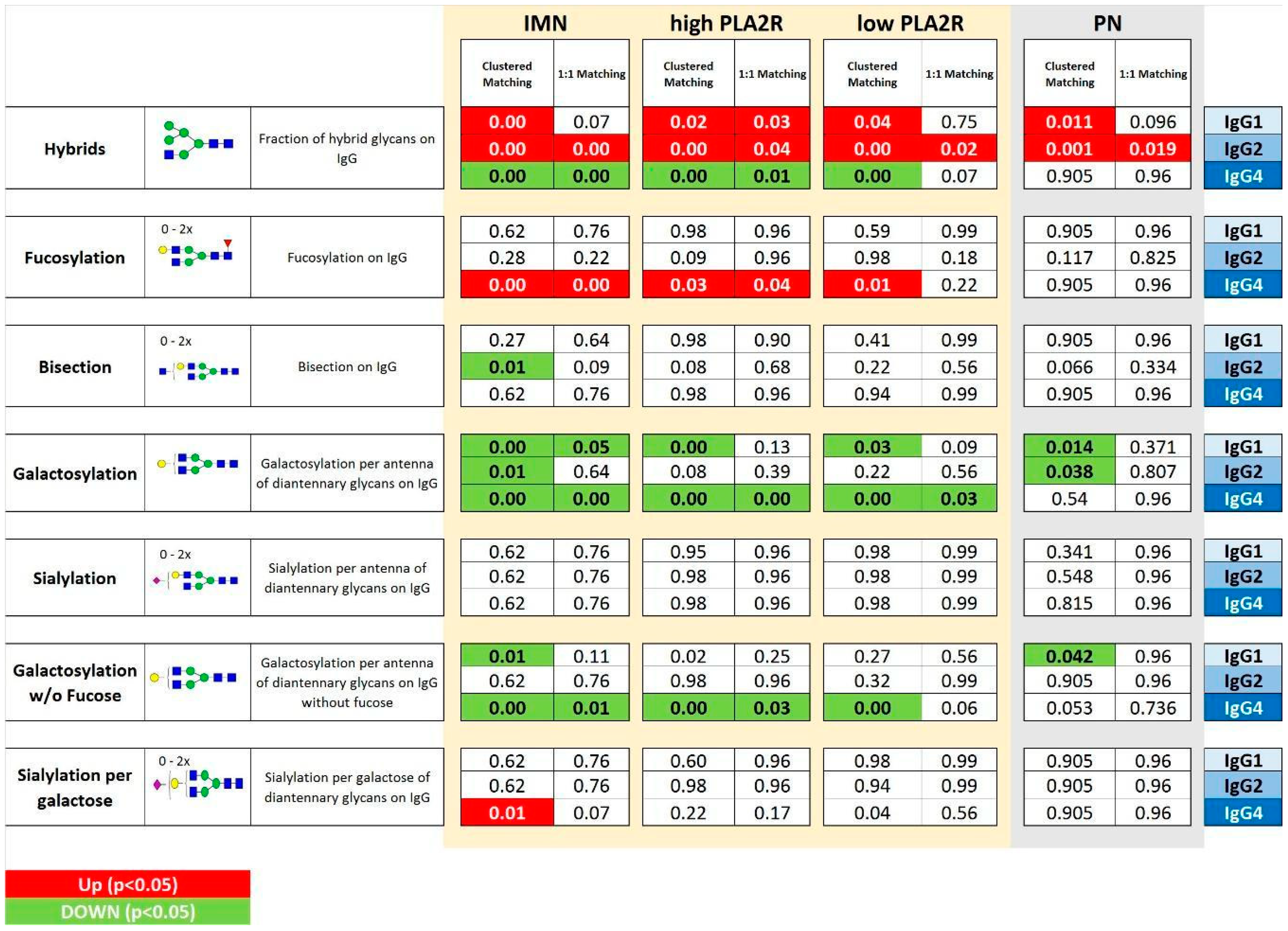
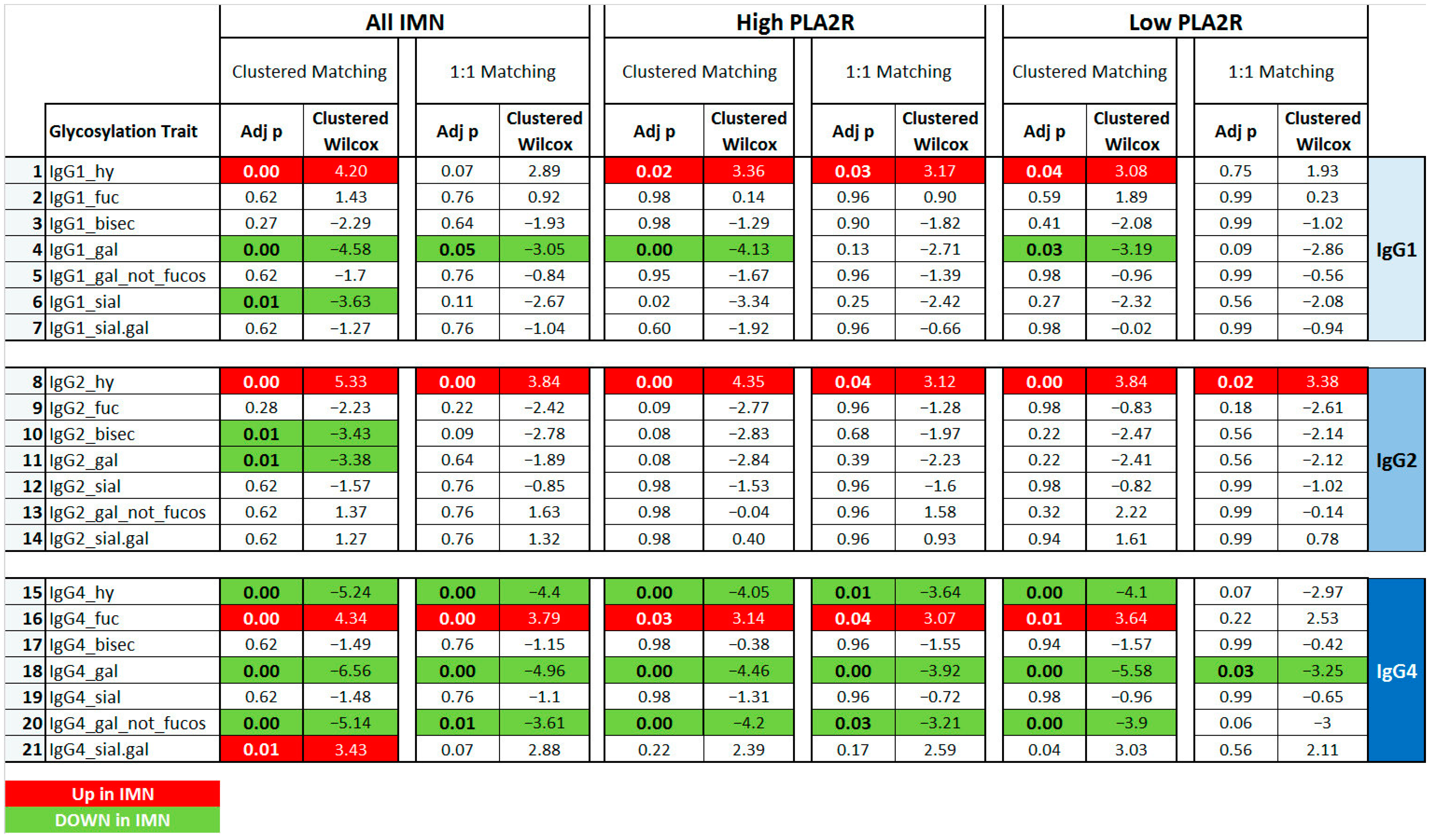
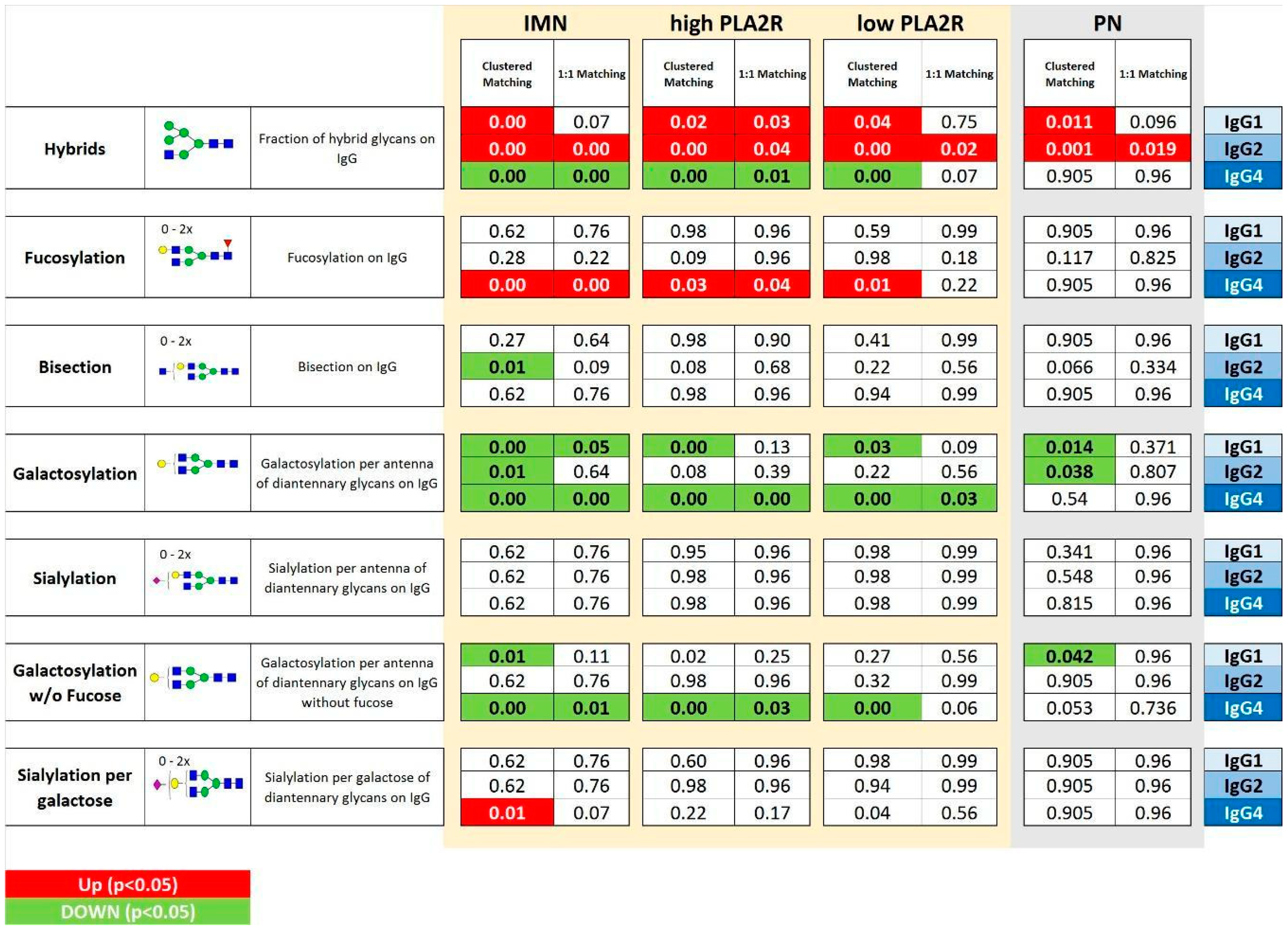
2.2. IgG Glycan Signatures of IMN Patients
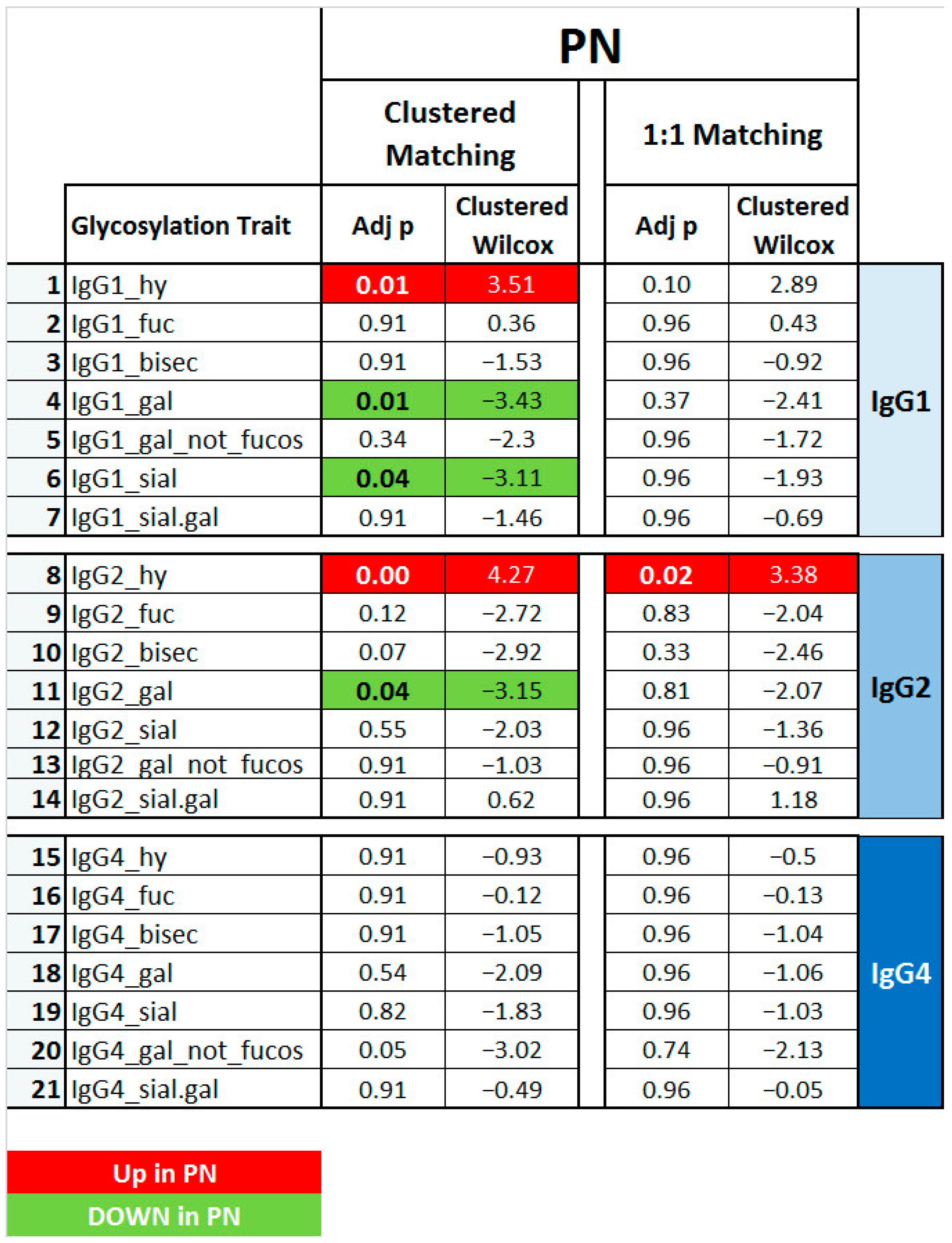
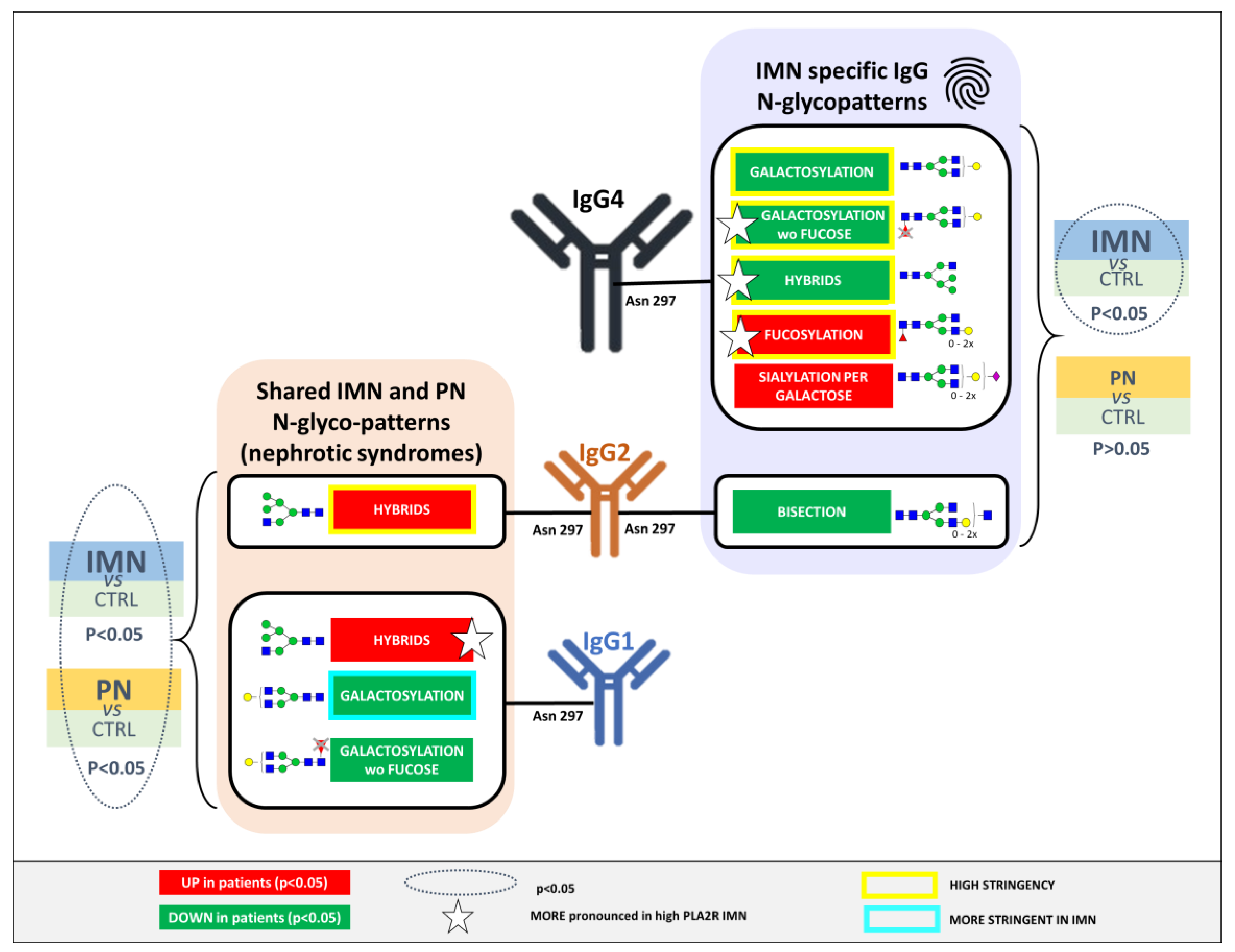
2.3. IMN-Specificity of IgG Glycan Signatures
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Human Sera Collection
4.3. IgG Capturing and Trypsin Digestion
4.4. LC-MS Analysis of IgG Glycopeptides
4.5. Data Pre-Processing
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wasserstein, A.G. Membranous glomerulonephritis. J. Am. Soc. Nephrol. 1997, 8, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Glassock, R.J. Diagnosis and natural course of membranous nephropathy. Semin. Nephrol. 2003, 23, 324–332. [Google Scholar] [CrossRef]
- Mallick, N.P.; Short, C.D.; Manos, J. Clinical membranous nephropathy. Nephron 1983, 34, 209–219. [Google Scholar] [CrossRef]
- Jennette, J.C.; Olson, J.L.; Schwartz, M.M.; Silva, F. Heptinstall’s Pathology of the Kidney; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; ISBN 9780781747509. [Google Scholar]
- Sinico, R.A.; Mezzina, N.; Trezzi, B.; Ghiggeri, G.M.; Radice, A. Immunology of membranous nephropathy: From animal models to humans. Clin. Exp. Immunol. 2016, 183, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, L.H.J.; Bonegio, R.G.B.; Lambeau, G.; Beck, D.M.; Powell, D.W.; Cummins, T.D.; Klein, J.B.; Salant, D.J. M-type phospholipase A2 receptor as target antigen in idiopathic membranous nephropathy. N. Engl. J. Med. 2009, 361, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhao, Y.; Fu, P. Diagnostic Test Accuracy of Serum Anti-PLA2R Autoantibodies and Glomerular PLA2R Antigen for Diagnosing Idiopathic Membranous Nephropathy: An Updated Meta-Analysis. Front. Med. 2018, 5, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, S. Membranous nephropathy: A single disease or a pattern of injury resulting from different diseases. Clin. Kidney J. 2021, 14, 2166–2169. [Google Scholar] [CrossRef]
- Rovin, B.H.; Adler, S.G.; Barratt, J.; Bridoux, F.; Burdge, K.A.; Chan, T.M.; Cook, H.T.; Fervenza, F.C.; Gibson, K.L.; Glassock, R.J.; et al. Executive summary of the KDIGO 2021 Guideline for the Management of Glomerular Diseases. Kidney Int. 2021, 100, 753–779. [Google Scholar] [CrossRef]
- Hu, S.-L.; Wang, D.; Gou, W.-J.; Lei, Q.-F.; Ma, T.-A.; Cheng, J.-Z. Diagnostic value of phospholipase A2 receptor in idiopathic membranous nephropathy: A systematic review and meta-analysis. J. Nephrol. 2014, 27, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Ronco, P.; Debiec, H. Pathogenesis of membranous nephropathy: Recent advances and future challenges. Nat. Rev. Nephrol. 2012, 8, 203–213. [Google Scholar] [CrossRef] [PubMed]
- van de Logt, A.-E.; Fresquet, M.; Wetzels, J.F.; Brenchley, P. The anti-PLA2R antibody in membranous nephropathy: What we know and what remains a decade after its discovery. Kidney Int. 2019, 96, 1292–1302. [Google Scholar] [CrossRef]
- Bally, S.; Debiec, H.; Ponard, D.; Dijoud, F.; Rendu, J.; Fauré, J.; Ronco, P.; Dumestre-Perard, C. Phospholipase A2 Receptor-Related Membranous Nephropathy and Mannan-Binding Lectin Deficiency. J. Am. Soc. Nephrol. 2016, 27, 3539–3544. [Google Scholar] [CrossRef]
- Salant, D.J. Genetic variants in membranous nephropathy: Perhaps a perfect storm rather than a straightforward conformeropathy? J. Am. Soc. Nephrol. 2013, 24, 525–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, G.; Lorenzen, J.M.; Ma, H.; de Haan, N.; Seeger, H.; Zaghrini, C.; Brandt, S.; Kölling, M.; Wegmann, U.; Kiss, B.; et al. Altered glycosylation of IgG4 promotes lectin complement pathway activation in anti-PLA2R1-associated membranous nephropathy. J. Clin. Investig. 2021, 131, e140453. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, G.; Treffers, L.; Plomp, R.; Bentlage, A.E.H.; de Boer, M.; Koeleman, C.A.M.; Lissenberg-Thunnissen, S.N.; Visser, R.; Brouwer, M.; Mok, J.Y.; et al. Decoding the Human Immunoglobulin G-Glycan Repertoire Reveals a Spectrum of Fc-Receptor- and Complement-Mediated-Effector Activities. Front. Immunol. 2017, 8, 877. [Google Scholar] [CrossRef]
- Peschke, B.; Keller, C.W.; Weber, P.; Quast, I.; Lünemann, J.D. Fc-Galactosylation of Human Immunoglobulin Gamma Isotypes Improves C1q Binding and Enhances Complement-Dependent Cytotoxicity. Front. Immunol. 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed]
- Subedi, G.P.; Barb, A.W. The immunoglobulin G1 N-glycan composition affects binding to each low affinity Fc γ receptor. MAbs 2016, 8, 1512–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippold, S.; Nicolardi, S.; Domínguez-Vega, E.; Heidenreich, A.-K.; Vidarsson, G.; Reusch, D.; Haberger, M.; Wuhrer, M.; Falck, D. Glycoform-resolved FcɣRIIIa affinity chromatography-mass spectrometry. MAbs 2019, 11, 1191–1196. [Google Scholar] [CrossRef] [Green Version]
- Goulabchand, R.; Vincent, T.; Batteux, F.; Eliaou, J.-F.; Guilpain, P. Impact of autoantibody glycosylation in autoimmune diseases. Autoimmun. Rev. 2014, 13, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Barrios, C.; Zierer, J.; Gudelj, I.; Štambuk, J.; Ugrina, I.; Rodríguez, E.; Soler, M.J.; Pavić, T.; Šimurina, M.; Keser, T.; et al. Glycosylation Profile of IgG in Moderate Kidney Dysfunction. J. Am. Soc. Nephrol. 2016, 27, 933–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renfrow, M.B.; Novak, J. What insights can proteomics give us into IgA nephropathy (Berger’s disease)? Expert Rev. Proteom. 2017, 14, 645–647. [Google Scholar] [CrossRef] [Green Version]
- van Timmeren, M.M.; van der Veen, B.S.; Stegeman, C.A.; Petersen, A.H.; Hellmark, T.; Collin, M.; Heeringa, P. IgG glycan hydrolysis attenuates ANCA-mediated glomerulonephritis. J. Am. Soc. Nephrol. 2010, 21, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, M. Antibody Glycosylation Predicts Relapse in Autoimmune Vasculitis. EBioMedicine 2017, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X. IgG N-glycans. Adv. Clin. Chem. 2021, 105, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Plomp, R.; Ruhaak, L.R.; Uh, H.-W.; Reiding, K.R.; Selman, M.; Houwing-Duistermaat, J.J.; Slagboom, P.E.; Beekman, M.; Wuhrer, M. Subclass-specific IgG glycosylation is associated with markers of inflammation and metabolic health. Sci. Rep. 2017, 7, 12325. [Google Scholar] [CrossRef] [Green Version]
- Štambuk, J.; Nakić, N.; Vučković, F.; Pučić-Baković, M.; Razdorov, G.; Trbojević-Akmačić, I.; Novokmet, M.; Keser, T.; Vilaj, M.; Štambuk, T.; et al. Global variability of the human IgG glycome. Aging 2020, 12, 15222–15259. [Google Scholar] [CrossRef]
- Baković, M.P.; Selman, M.H.J.; Hoffmann, M.; Rudan, I.; Campbell, H.; Deelder, A.M.; Lauc, G.; Wuhrer, M. High-throughput IgG Fc N-glycosylation profiling by mass spectrometry of glycopeptides. J. Proteome Res. 2013, 12, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wang, Y.; Qin, X.; Li, H.; Guo, Y.; Wang, Y.; Liu, H.; Wang, X.; Song, G.; Li, F.; et al. Change in IgG1 Fc N-linked glycosylation in human lung cancer: Age- and sex-related diagnostic potential. Electrophoresis 2013, 34, 2407–2416. [Google Scholar] [CrossRef]
- Ercan, A. Sex effect on the correlation of immunoglobulin G glycosylation with rheumatoid arthritis disease activity. Turkish J. Biol. Turk Biyol. Derg. 2020, 44, 406–416. [Google Scholar] [CrossRef]
- Costa, D.M.d.N.; Valente, L.M.; Vajgel Fernandes, G.; Sandrin-Garcia, P.; da Cruz, H.L.A.; Crovella, S.; Cavalcante, A.H.S.; Cavalcante, M.A.G.d.M.; de Oliveira, C.B.L.; de Vasconcelos, C.d.A.J.; et al. Mannose-Binding Lectin2 Gene Polymorphism and IgG4 in Membranous Nephropathy. Nephron 2018, 139, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Olio, F.; Malagolini, N. Immunoglobulin G Glycosylation Changes in Aging and Other Inflammatory Conditions. Exp. Suppl. 2021, 112, 303–340. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, F. Glycobiology of Aging. Subcell. Biochem. 2018, 90, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Pozdzik, A.; Brochériou, I.; David, C.; Touzani, F.; Goujon, J.M.; Wissing, K.M. Membranous Nephropathy and Anti-Podocytes Antibodies: Implications for the Diagnostic Workup and Disease Management. BioMed Res. Int. 2018, 2018, 6281054. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, F.; Vanhooren, V.; Chen, C.C.; Slagboom, P.E.; Wuhrer, M.; Franceschi, C. N-glycomic biomarkers of biological aging and longevity: A link with inflammaging. Ageing Res. Rev. 2013, 12, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Lagattuta, K.A.; Nigrovic, P.A. Estrogen-Driven Changes in Immunoglobulin G Fc Glycosylation. Exp. Suppl. 2021, 112, 341–361. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Im, W. Effects of N-Glycan Composition on Structure and Dynamics of IgG1 Fc and Their Implications for Antibody Engineering. Sci. Rep. 2017, 7, 12659. [Google Scholar] [CrossRef] [Green Version]
- Krištić, J.; Vučković, F.; Menni, C.; Klarić, L.; Keser, T.; Beceheli, I.; Pučić-Baković, M.; Novokmet, M.; Mangino, M.; Thaqi, K.; et al. Glycans are a novel biomarker of chronological and biological ages. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, Y.X.; Zhang, J.; Yu, X.W.; Ge, S.S.; Wang, H.; Dong, J.; Guo, X.H.; Wang, W. Comparison of gender specific structure profiles of immunoglobulin G N-glycans. Zhonghua Liu Xing Bing Xue Za Zhi 2016, 37, 1409–1412. [Google Scholar] [CrossRef] [PubMed]
- Sanada, F.; Taniyama, Y.; Muratsu, J.; Otsu, R.; Shimizu, H.; Rakugi, H.; Morishita, R. Source of Chronic Inflammation in Aging. Front. Cardiovasc. Med. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ząbczyńska, M.; Link-Lenczowski, P.; Pocheć, E. Glycosylation in Autoimmune Diseases. Adv. Exp. Med. Biol. 2021, 1325, 205–218. [Google Scholar] [CrossRef]
- Zaytseva, O.O.; Sharapov, S.Z.; Perola, M.; Esko, T.; Landini, A.; Hayward, C.; Wilson, J.F.; Lauc, G.; Aulchenko, Y.S.; Klarić, L.; et al. Investigation of the causal relationships between human IgG N-glycosylation and twelve common diseases associated with changes in the IgG N-glycome. Hum. Mol. Genet. 2021. [Google Scholar] [CrossRef]
- Kao, C.-C.; Wang, S.-Y.; Chuang, Y.-K.; Lee, W.-Y.; Chang, W.-C.; Wu, M.-S.; Kuo, T.-C.; Tsai, I.-L. Clinical Mass Spectrometry Discovered Human IgG Sialylation as a Potential Biosignature for Kidney Function. J. Pers. Med. 2021, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Na, W.; Yi, K.; Song, Y.S.; Park, M.H. Dissecting the relationships of IgG subclasses and complements in membranous lupus nephritis and idiopathic membranous nephropathy. PLoS ONE 2017, 12, e0174501. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.B.; Syed, B.A. From the analyst’s couch: Next-generation antibodies. Nat. Rev. Drug Discov. 2014, 13, 413–414. [Google Scholar] [CrossRef]
- Han, J.; Zhou, Z.; Zhang, R.; You, Y.; Guo, Z.; Huang, J.; Wang, F.; Sun, Y.; Liu, H.; Cheng, X.; et al. Fucosylation of anti-dsDNA IgG1 correlates with disease activity of treatment-naïve systemic lupus erythematosus patients. EBioMedicine 2022, 77, 103883. [Google Scholar] [CrossRef] [PubMed]
- Takei, D.; Harada, K.; Nouso, K.; Miyahara, K.; Dohi, C.; Matsushita, H.; Kinugasa, H.; Hiraoka, S.; Nishimura, S.-I.; Okada, H. Clinical utility of a serum glycome analysis in patients with colorectal cancer. J. Gastroenterol. Hepatol. 2022, 37, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, K.; Nouso, K.; Dohi, C.; Morimoto, Y.; Kinugasa, H.; Wada, N.; Takeuchi, Y.; Kuwaki, K.; Onishi, H.; Ikeda, F.; et al. Alteration of N-glycan profiles in patients with chronic hepatitis and hepatocellular carcinoma. Hepatol. Res. 2015, 45, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Nouso, K.; Miyahara, K.; Wada, N.; Dohi, C.; Morimoto, Y.; Kinugasa, H.; Takeuchi, Y.; Yasunaka, T.; Kuwaki, K.; et al. Use of non-invasive serum glycan markers to distinguish non-alcoholic steatohepatitis from simple steatosis. J. Gastroenterol. Hepatol. 2015, 30, 528–534. [Google Scholar] [CrossRef]
- Culver, E.L.; van de Bovenkamp, F.S.; Derksen, N.I.L.; Koers, J.; Cargill, T.; Barnes, E.; de Neef, L.A.; Koeleman, C.A.M.; Aalberse, R.C.; Wuhrer, M.; et al. Unique patterns of glycosylation in immunoglobulin subclass G4-related disease and primary sclerosing cholangitis. J. Gastroenterol. Hepatol. 2019, 34, 1878–1886. [Google Scholar] [CrossRef] [PubMed]
- Lippold, S.; Knaupp, A.; de Ru, A.H.; Tjokrodirijo, R.T.N.; van Veelen, P.A.; van Puijenbroek, E.; de Taeye, S.W.; Reusch, D.; Vidarsson, G.; Wuhrer, M.; et al. Fc gamma receptor IIIb binding of individual antibody proteoforms resolved by affinity chromatography-mass spectrometry. MAbs 2021, 13, 1982847. [Google Scholar] [CrossRef]
- Memarian, E.; M’t Hart, L.; Slieker, R.C.; Lemmers, R.F.L.; van der Heijden, A.A.; Rutters, F.; Nijpels, G.; Schoep, E.; Lieverse, A.G.; Sijbrands, E.J.G.; et al. Plasma protein N-glycosylation is associated with cardiovascular disease, nephropathy, and retinopathy in type 2 diabetes. BMJ Open Diabetes Res. Care 2021, 9, e002345. [Google Scholar] [CrossRef]
- Magorivska, I.; Döncző, B.; Dumych, T.; Karmash, A.; Boichuk, M.; Hychka, K.; Mihalj, M.; Szabó, M.; Csánky, E.; Rech, J.; et al. Glycosylation of random IgG distinguishes seropositive and seronegative rheumatoid arthritis. Autoimmunity 2018, 51, 111–117. [Google Scholar] [CrossRef]
- Washburn, N.; Schwab, I.; Ortiz, D.; Bhatnagar, N.; Lansing, J.C.; Medeiros, A.; Tyler, S.; Mekala, D.; Cochran, E.; Sarvaiya, H.; et al. Controlled tetra-Fc sialylation of IVIg results in a drug candidate with consistent enhanced anti-inflammatory activity. Proc. Natl. Acad. Sci. USA 2015, 112, E1297–E1306. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Huang, Q.; Wang, J.; Luo, K.; Liu, J.; Yan, W.; Jiang, F.; Xu, G. Efficacy and Safety of Different Immunosuppressive Therapies in Patients with Membranous Nephropathy and High PLA2R Antibody Titer. Front. Pharmacol. 2021, 12, 786334. [Google Scholar] [CrossRef]
- Jatem-Escalante, E.; Martín-Conde, M.L.; Gràcia-Lavedan, E.; Benítez, I.D.; Gonzalez, J.; Colás, L.; Garcia-Carrasco, A.; Martínez, C.; Segarra-Medrano, A. Monitoring anti-PLA2R antibody titres to predict the likelihood of spontaneous remission of membranous nephropathy. Clin. Kidney J. 2021, 14, 2556–2562. [Google Scholar] [CrossRef]
- Li, Y.Q.; Liu, Z.Z.; Lin, K.X.; Liu, R.H.; Ni, L.; Liu, G.X.; Lan, H.Y.; Shi, Y.J. Relationship between the status of phospholipase A2 receptor and prognosis of idiopathic membranous nephropathy. Nephrology 2020, 25, 144–149. [Google Scholar] [CrossRef]
- Radice, A.; Trezzi, B.; Maggiore, U.; Pregnolato, F.; Stellato, T.; Napodano, P.; Rolla, D.; Pesce, G.; D’Amico, M.; Santoro, D.; et al. Clinical usefulness of autoantibodies to M-type phospholipase A2 receptor (PLA2R) for monitoring disease activity in idiopathic membranous nephropathy (IMN). Autoimmun. Rev. 2016, 15, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Radice, A.; Pieruzzi, F.; Trezzi, B.; Ghiggeri, G.; Napodano, P.; D’Amico, M.; Stellato, T.; Brugnano, R.; Ravera, F.; Rolla, D.; et al. Diagnostic specificity of autoantibodies to M-type phospholipase A2 receptor (PLA2R) in differentiating idiopathic membranous nephropathy (IMN) from secondary forms and other glomerular diseases. J. Nephrol. 2018, 31, 271–278. [Google Scholar] [CrossRef]
- Falck, D.; Jansen, B.C.; de Haan, N.; Wuhrer, M. High-Throughput Analysis of IgG Fc Glycopeptides by LC-MS. Methods Mol. Biol. 2017, 1503, 31–47. [Google Scholar] [CrossRef]
- Wuhrer, M.; Stam, J.C.; van de Geijn, F.E.; Koeleman, C.A.M.; Verrips, C.T.; Dolhain, R.J.E.M.; Hokke, C.H.; Deelder, A.M. Glycosylation profiling of immunoglobulin G (IgG) subclasses from human serum. Proteomics 2007, 7, 4070–4081. [Google Scholar] [CrossRef] [PubMed]
- Jansen, B.C.; Falck, D.; de Haan, N.; Hipgrave Ederveen, A.L.; Razdorov, G.; Lauc, G.; Wuhrer, M. LaCyTools: A Targeted Liquid Chromatography-Mass Spectrometry Data Processing Package for Relative Quantitation of Glycopeptides. J. Proteome Res. 2016, 15, 2198–2210. [Google Scholar] [CrossRef] [PubMed]
- de Haan, N.; Boeddha, N.P.; Ekinci, E.; Reiding, K.R.; Emonts, M.; Hazelzet, J.A.; Wuhrer, M.; Driessen, G.J. Differences in IgG Fc Glycosylation Are Associated with Outcome of Pediatric Meningococcal Sepsis. MBio 2018, 9, e00546-18. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chinello, C.; de Haan, N.; Capitoli, G.; Trezzi, B.; Radice, A.; Pagani, L.; Criscuolo, L.; Signorini, S.; Galimberti, S.; Sinico, R.A.; et al. Definition of IgG Subclass-Specific Glycopatterns in Idiopathic Membranous Nephropathy: Aberrant IgG Glycoforms in Blood. Int. J. Mol. Sci. 2022, 23, 4664. https://doi.org/10.3390/ijms23094664
Chinello C, de Haan N, Capitoli G, Trezzi B, Radice A, Pagani L, Criscuolo L, Signorini S, Galimberti S, Sinico RA, et al. Definition of IgG Subclass-Specific Glycopatterns in Idiopathic Membranous Nephropathy: Aberrant IgG Glycoforms in Blood. International Journal of Molecular Sciences. 2022; 23(9):4664. https://doi.org/10.3390/ijms23094664
Chicago/Turabian StyleChinello, Clizia, Noortje de Haan, Giulia Capitoli, Barbara Trezzi, Antonella Radice, Lisa Pagani, Lucrezia Criscuolo, Stefano Signorini, Stefania Galimberti, Renato Alberto Sinico, and et al. 2022. "Definition of IgG Subclass-Specific Glycopatterns in Idiopathic Membranous Nephropathy: Aberrant IgG Glycoforms in Blood" International Journal of Molecular Sciences 23, no. 9: 4664. https://doi.org/10.3390/ijms23094664
APA StyleChinello, C., de Haan, N., Capitoli, G., Trezzi, B., Radice, A., Pagani, L., Criscuolo, L., Signorini, S., Galimberti, S., Sinico, R. A., Wuhrer, M., & Magni, F. (2022). Definition of IgG Subclass-Specific Glycopatterns in Idiopathic Membranous Nephropathy: Aberrant IgG Glycoforms in Blood. International Journal of Molecular Sciences, 23(9), 4664. https://doi.org/10.3390/ijms23094664