
Metabolic Alteration Analysis of Steroid Hormones in Niemann–Pick Disease Type C Model Cell Using Liquid Chromatography/Tandem Mass Spectrometry

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results and Discussion
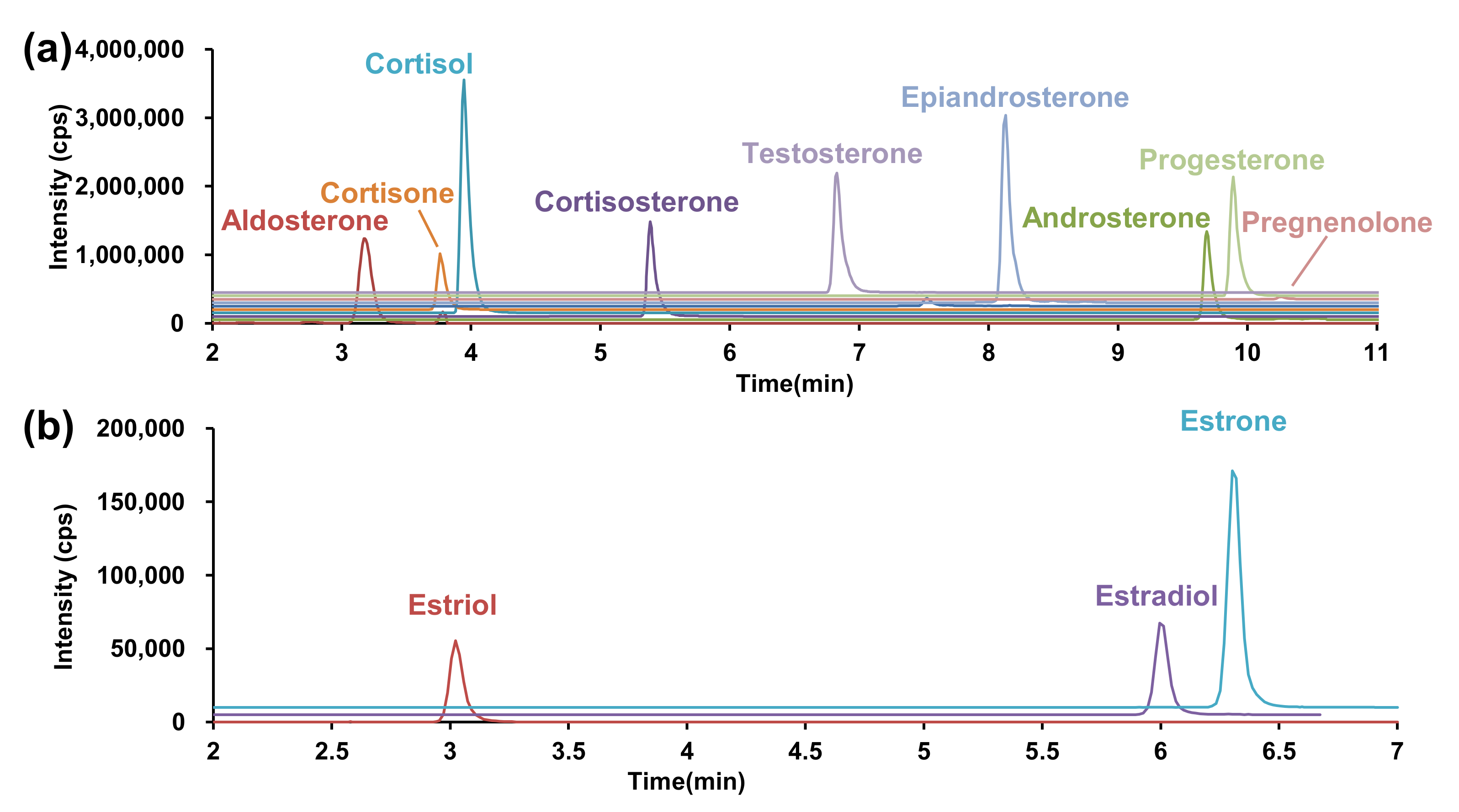
2.1. MS/MS Optimization for Steroid Hormone Analysis
2.2. LC optimization for Steroid Hormone Analysis
2.3. Extract Recovery
2.4. Analytical Method Validation
2.4.1. Matrix Factor
2.4.2. Calibration Curves
2.4.3. Reproducibility Test
2.5. Cells
2.5.1. Confirmation of Cholesterol Accumulation Using Filipin Staining
2.5.2. Organelle Observation
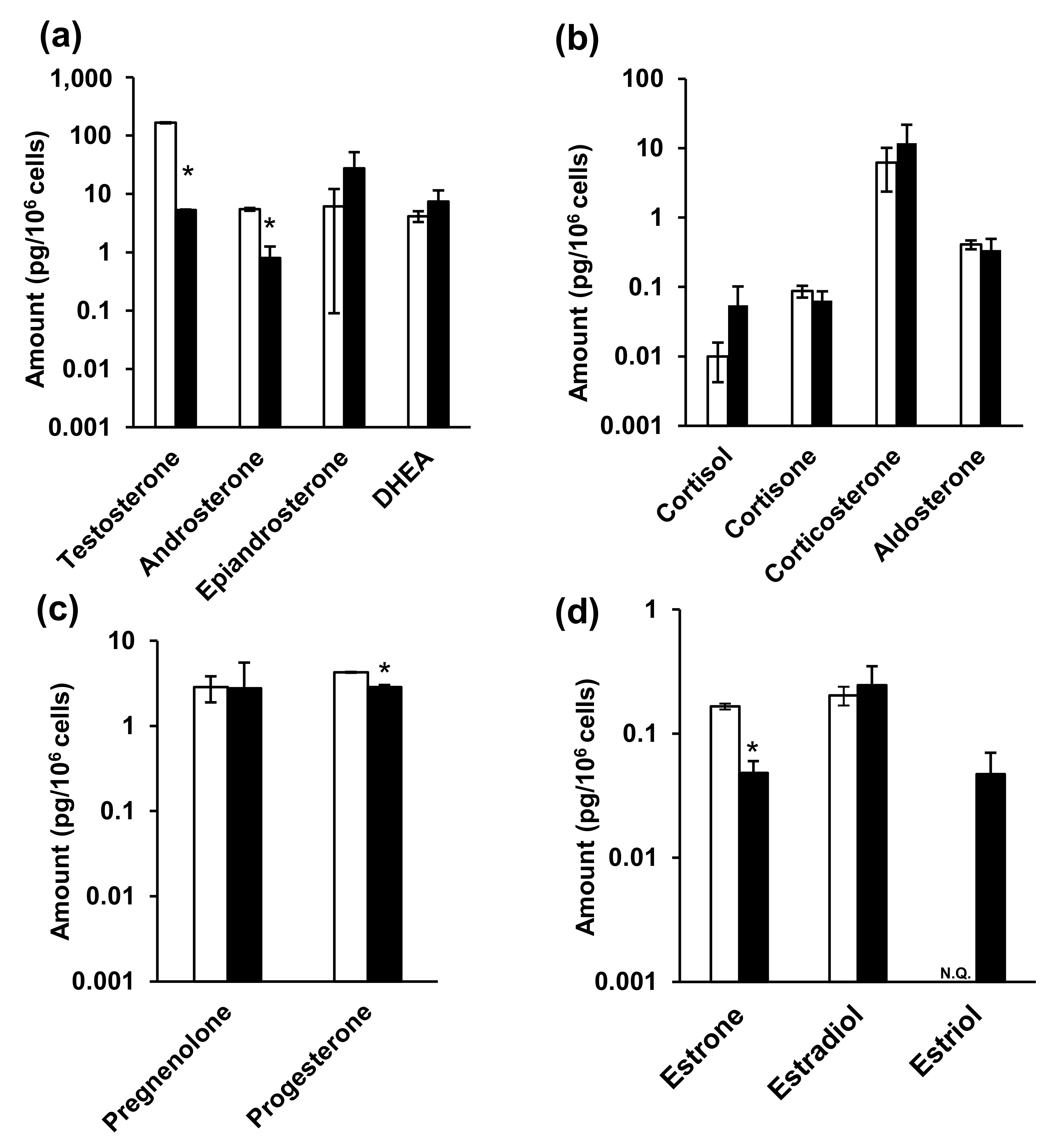
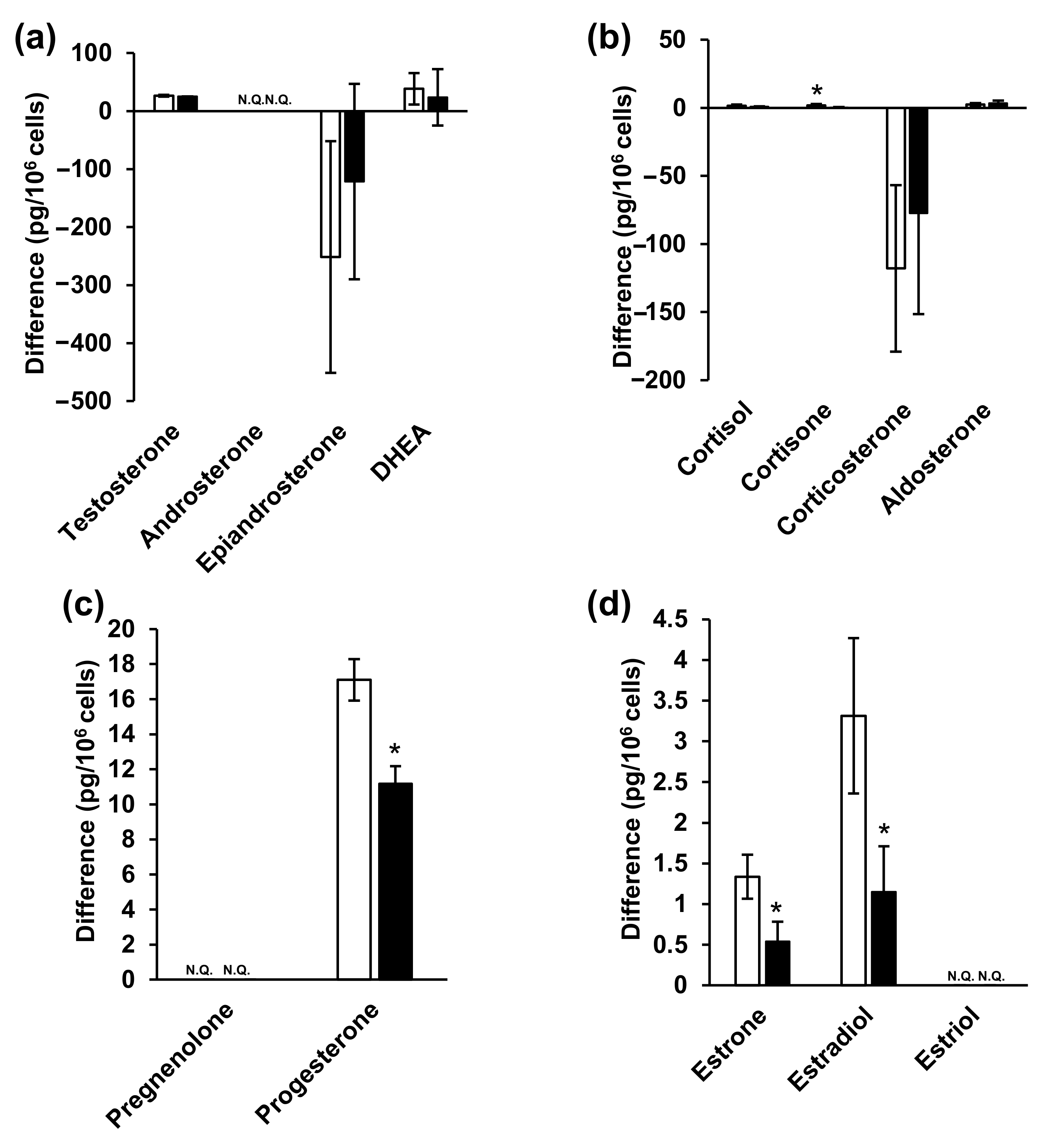
2.6. Steroid Hormone Analysis in NPC Model Cell
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. LC–MS/MS Equipment
3.3. ESI–MS/MS Conditions
3.4. LC Conditions
3.4.1. Positive Ion Detection Mode
3.4.2. Negative Ion Detection Mode
3.5. Preparation of Standard Steroid Hormone Solutions
3.6. Preparation of IS Solution
3.7. Liquid–Liquid Extraction
3.8. Matrix Effect
3.9. Calibration Curve Preparation
3.10. Reproducibility Test
3.11. NPC Model Cell and Medium
3.12. Cholesterol Staining
3.13. Organelle Observation
3.14. Cells for LC–MS/MS Analysis of Steroid Hormones
3.15. Sample Preparation Procedure
3.16. Quantification of Steroid Hormones in the Sample
3.17. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanier, M.T. Niemann-Pick Disease Type C. Orphanet J. Rare Dis. 2010, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Carstea, E.D.; Morris, J.A.; Coleman, K.G.; Loftus, S.K.; Zhang, D.; Cummings, C.; Gu, J.; Rosenfeld, M.A.; Pavan, W.J.; Krizman, D.B.; et al. Niemann-Pick C1 Disease Gene: Homology to Mediators of Cholesterol Homeostasis. Science 1997, 277, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Carstea, E.D.; Polymeropoulos, M.H.; Parker, C.C.; Detera-Wadleigh, S.D.; O’Neill, R.R.; Patterson, M.C.; Goldin, E.; Xiao, H.; Straub, R.E.; Vanier, M.T. Linkage of Niemann-Pick Disease Type C to Human Chromosome 18. Proc. Natl. Acad. Sci. USA 1993, 90, 2002–2004. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, S.J.; Ward, C.P.; Fensom, A.H. Complementation Studies in Niemann-Pick Disease Type C Indicate the Existence of a Second Group. J. Med. Genet. 1994, 31, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Naureckiene, S.; Sleat, D.E.; Lackland, H.; Fensom, A.; Vanier, M.T.; Wattiaux, R.; Jadot, M.; Lobel, P. Identification of HE1 as the Second Gene of Niemann-Pick C Disease. Science 2000, 290, 2298–2301. [Google Scholar] [CrossRef]
- Kwon, H.J.; Abi-Mosleh, L.; Wang, M.L.; Deisenhofer, J.; Goldstein, J.L.; Brown, M.S.; Infante, R.E. Structure of N-Terminal Domain of NPC1 Reveals Distinct Subdomains for Binding and Transfer of Cholesterol. Cell 2009, 137, 1213–1224. [Google Scholar] [CrossRef]
- Patterson, M.C.; Hendriksz, C.J.; Walterfang, M.; Sedel, F.; Vanier, M.T.; Wijburg, F. Recommendations for the Diagnosis and Management of Niemann-Pick Disease Type C: An Update. Mol. Genet. Metab. 2012, 106, 330–344. [Google Scholar] [CrossRef]
- Patterson, M.C.; Clayton, P.; Gissen, P.; Anheim, M.; Bauer, P.; Bonnot, O.; Dardis, A.; Dionisi-Vici, C.; Klünemann, H.H.; Latour, P.; et al. Recommendations for the Detection and Diagnosis of Niemann-Pick Disease Type C: An Update. Neurol. Clin. Pract. 2017, 7, 499–511. [Google Scholar] [CrossRef]
- Geberhiwot, T.; Moro, A.; Dardis, A.; Ramaswami, U.; Sirrs, S.; Marfa, M.P.; Vanier, M.T.; Walterfang, M.; Bolton, S.; Dawson, C.; et al. Consensus Clinical Management Guidelines for Niemann-Pick Disease Type C. Orphanet J. Rare Dis. 2018, 13, 50. [Google Scholar] [CrossRef]
- Patterson, M. Niemann-Pick Disease Type C; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Ohno, K. Niemann-Pick Type C Disease: Pathophysiology, Diagnosis and Treatment. No Hattatsu 2016, 48, 81–88. [Google Scholar] [CrossRef]
- Patterson, M.C.; Mengel, E.; Vanier, M.T.; Schwierin, B.; Muller, A.; Cornelisse, P.; Pineda, M.; Amado-Fondo, A.; Amraoui, Y.; Andria, G.; et al. Stable or Improved Neurological Manifestations during Miglustat Therapy in Patients from the International Disease Registry for Niemann-Pick Disease Type C: An Observational Cohort Study. Orphanet J. Rare Dis. 2015, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Ory, D.S.; Ottinger, E.A.; Farhat, N.Y.; King, K.A.; Jiang, X.; Weissfeld, L.; Berry-Kravis, E.; Davidson, C.D.; Bianconi, S.; Keener, L.A.; et al. Intrathecal 2-Hydroxypropyl-β-Cyclodextrin Decreases Neurological Disease Progression in Niemann-Pick Disease, Type C1: A Non-Randomised, Open-Label, Phase 1–2 Trial. Lancet 2017, 390, 1758–1768. [Google Scholar] [CrossRef]
- Shioi, R.; Karaki, F.; Yoshioka, H.; Noguchi-Yachide, T.; Ishikawa, M.; Dodo, K.; Hashimoto, Y.; Sodeoka, M.; Ohgane, K. Image-Based Screen Capturing Misfolding Status of Niemann-Pick Type C1 Identifies Potential Candidates for Chaperone Drugs. PLoS ONE 2020, 15, e0243746. [Google Scholar] [CrossRef] [PubMed]
- Pipalia, N.H.; Saad, S.Z.; Subramanian, K.; Cross, A.; al-Motawa, A.; Garg, K.; Blagg, B.S.J.; Neckers, L.; Helquist, P.; Wiest, O.; et al. HSP90 Inhibitors Reduce Cholesterol Storage in Niemann-Pick Type C1 Mutant Fibroblasts. J. Lipid Res. 2021, 63, 100114. [Google Scholar] [CrossRef]
- Kirkegaard, T.; Roth, A.G.; Petersen, N.H.T.; Mahalka, A.K.; Olsen, O.D.; Moilanen, I.; Zylicz, A.; Knudsen, J.; Sandhoff, K.; Arenz, C.; et al. Hsp70 Stabilizes Lysosomes and Reverts Niemann-Pick Disease-Associated Lysosomal Pathology. Nature 2010, 463, 549–553. [Google Scholar] [CrossRef]
- Fukuda, H.; Karaki, F.; Dodo, K.; Noguchi-Yachide, T.; Ishikawa, M.; Hashimoto, Y.; Ohgane, K. Phenanthridin-6-One Derivatives as the First Class of Non-Steroidal Pharmacological Chaperones for Niemann-Pick Disease Type C1 Protein. Bioorg. Med. Chem. Lett. 2017, 27, 2781–2787. [Google Scholar] [CrossRef]
- Paulina Ordonez, M.; Roberts, E.A.; Kidwell, C.U.; Yuan, S.H.; Plaisted, W.C.; Goldstein, L.S.B. Disruption and Therapeutic Rescue of Autophagy in a Human Neuronal Model of Niemann Pick Type C1. Hum. Mol. Genet. 2012, 21, 2651–2662. [Google Scholar] [CrossRef]
- Elrick, M.J.; Yu, T.; Chung, C.; Lieberman, A.P. Impaired Proteolysis Underlies Autophagic Dysfunction in Niemann-Pick Type C Disease. Hum. Mol. Genet. 2012, 21, 4876–4887. [Google Scholar] [CrossRef]
- Ordonez, M.P. Defective Mitophagy in Human Niemann-Pick Type C1 Neurons Is Due to Abnormal Autophagy Activation. Autophagy 2012, 8, 1157–1158. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, M.; Qiu, X.; Deis, J.A.; Huang, H.; Tang, Q.Q.; Chen, X. Niemann-Pick Type C2 Deficiency Impairs Autophagylysosomal Activity, Mitochondrial Function, and TLR Signaling in Adipocytes. J. Lipid Res. 2016, 57, 1644–1658. [Google Scholar] [CrossRef]
- Torres, S.; Balboa, E.; Zanlungo, S.; Enrich, C.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Lysosomal and Mitochondrial Liaisons in Niemann-Pick Disease. Front. Physiol. 2017, 8, 982. [Google Scholar] [CrossRef]
- Yu, W.; Gong, J.S.; Ko, M.; Garver, W.S.; Yanagisawa, K.; Michikawa, M. Altered Cholesterol Metabolism in Niemann-Pick Type C1 Mouse Brains Affects Mitochondrial Function. J. Biol. Chem. 2005, 280, 11731–11739. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; Matías, N.; Baulies, A.; Nuñez, S.; Alarcon-Vila, C.; Martinez, L.; Nuño, N.; Fernandez, A.; Caballeria, J.; Levade, T.; et al. Mitochondrial GSH Replenishment as a Potential Therapeutic Approach for Niemann Pick Type C Disease. Redox Biol. 2017, 11, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hales, D.B. Overview of Steroidogenic Enzymes in the Pathway from Cholesterol to Active Steroid Hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Steroid Hormone Synthesis in Mitochondria. Mol. Cell. Endocrinol. 2013, 379, 62–73. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Cole, T.J.; Short, K.L.; Hooper, S.B. The Science of Steroids. Semin. Fetal Neonatal Med. 2019, 24, 170–175. [Google Scholar] [CrossRef]
- Holst, J.P.; Soldin, O.P.; Guo, T.; Soldin, S.J. Steroid Hormones: Relevance and Measurement in the Clinical Laboratory. Clin. Lab. Med. 2004, 24, 105–118. [Google Scholar] [CrossRef]
- Gévry, N.Y.; Lopes, F.L.; Ledoux, S.; Murphy, B.D. Aberrant Intracellular Cholesterol Transport Disrupts Pituitary and Ovarian Function. Mol. Endocrinol. 2004, 18, 1778–1786. [Google Scholar] [CrossRef][Green Version]
- Chen, G.; Li, H.M.; Chen, Y.R.; Gu, X.S.; Duan, S. Decreased Estradiol Release from Astrocytes Contributes to the Neurodegeneration in a Mouse Model of Niemann-Pick Disease Type C. GLIA 2007, 55, 1509–1518. [Google Scholar] [CrossRef]
- Xie, C.; Richardson, J.A.; Turley, S.D.; Dietschy, J.M. Cholesterol Substrate Pools and Steroid Hormone Levels Are Normal in the Face of Mutational Inactivation of NPC1 Protein. J. Lipid Res. 2006, 47, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.D.; Gong, W.; Verot, L.; Mellon, S.H. Niemann-Pick Type C Disease Involves Disrupted Neurosteroidogenesis and Responds to Allopregnanolone. Nat. Med. 2004, 10, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Akpovi, C.D.; Murphy, B.D.; Erickson, R.P.; Pelletier, R.-M. Dysregulation of Testicular Cholesterol Metabolism Following Spontaneous Mutation of the Niemann-Pick C1 Gene in Mice1. Biol. Reprod. 2014, 91, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Mellon, S.H.; Gong, W.; Schonemann, M.D. Endogenous and Synthetic Neurosteroids in Treatment of Niemann-Pick Type C Disease. Brain Res. Rev. 2008, 57, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Disorders in the Initial Steps of Steroid Hormone Synthesis. J. Steroid Biochem. Mol. Biol. 2017, 165, 18–37. [Google Scholar] [CrossRef]
- Lamri, A.; Pigeyre, M.; Garver, W.S.; Meyre, D. The Extending Spectrum of NPC1-Related Human Disorders: From Niemann–Pick C1 Disease to Obesity. Endocr. Rev. 2018, 39, 192–220. [Google Scholar] [CrossRef]
- Barnard, G.; Kohen, F. Monitoring Ovarian Function by the Simultaneous Time-Resolved Fluorescence Immunoassay of Two Urinary Steroid Metabolites. Clin. Chem. 1998, 44, 1520–1528. [Google Scholar] [CrossRef]
- Keevil, B.G. LC–MS/MS Analysis of Steroids in the Clinical Laboratory. Clin. Biochem. 2016, 49, 989–997. [Google Scholar] [CrossRef]
- Higashi, T.; Shimada, K. Derivatization of Neutral Steroids to Enhance Their Detection Characteristics in Liquid Chromatography-Mass Spectrometry. Anal. Bioanal. Chem. 2004, 378, 875–882. [Google Scholar] [CrossRef]
- Higashi, T.; Ogawa, S. Chemical Derivatization for Enhancing Sensitivity during LC/ESI–MS/MS Quantification of Steroids in Biological Samples: A Review. J. Steroid Biochem. Mol. Biol. 2016, 162, 57–69. [Google Scholar] [CrossRef]
- Higaki, K.; Ninomiya, H.; Sugimoto, Y.; Suzuki, T.; Taniguchi, M.; Niwa, H.; Pentchev, P.G.; Vanier, M.T.; Ohno, K. Isolation of NPC1-Deficient Chinese Hamster Ovary Cell Mutants by Gene Trap Mutagenesis. J. Biochem. 2001, 129, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Keski-Rahkonen, P.; Desai, R.; Jimenez, M.; Harwood, D.T.; Handelsman, D.J. Measurement of Estradiol in Human Serum by LC-MS/MS Using a Novel Estrogen-Specific Derivatization Reagent. Anal. Chem. 2015, 87, 7180–7186. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Shimada, K. Application of Cookson-Type Reagents for Biomedical HPLC and LC/MS Analyses: A Brief Overview. Biomed. Chromatogr. 2017, 31, e3808. [Google Scholar] [CrossRef] [PubMed]
- Lavrynenko, O.; Nedielkov, R.; Möller, H.M.; Shevchenko, A. Girard Derivatization for LC-MS/MS Profiling of Endogenous Ecdysteroids in Drosophila. J. Lipid Res. 2013, 54, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Ogawa, S. Isotope-Coded ESI-Enhancing Derivatization Reagents for Differential Analysis, Quantification and Profiling of Metabolites in Biological Samples by LC/MS: A Review. J. Pharm. Biomed. Anal. 2016, 130, 181–193. [Google Scholar] [CrossRef]
- Yuan, T.F.; Le, J.; Cui, Y.; Peng, R.; Wang, S.T.; Li, Y. An LC-MS/MS Analysis for Seven Sex Hormones in Serum. J. Pharm. Biomed. Anal. 2019, 162, 34–40. [Google Scholar] [CrossRef]
- Yuan, T.F.; Le, J.; Wang, S.T.; Li, Y. An LC/MS/MS Method for Analyzing the Steroid Metabolome with High Accuracy and from Small Serum Samples. J. Lipid Res. 2020, 61, 580–586. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Kellermann, G. Simultaneous Determination of Three Estrogens in Human Saliva without Derivatization or Liquid-Liquid Extraction for Routine Testing via Miniaturized Solid Phase Extraction with LC-MS/MS Detection. Talanta 2018, 178, 464–472. [Google Scholar] [CrossRef]
- Márta, Z.; Bobály, B.; Fekete, J.; Magda, B.; Imre, T.; Mészáros, K.V.; Bálint, M.; Szabó, P.T. Simultaneous Determination of Thirteen Different Steroid Hormones Using Micro UHPLC-MS/MS with on-Line SPE System. J. Pharm. Biomed. Anal. 2018, 150, 258–267. [Google Scholar] [CrossRef]
- Gaudl, A.; Kratzsch, J.; Bae, Y.J.; Kiess, W.; Thiery, J.; Ceglarek, U. Liquid Chromatography Quadrupole Linear Ion Trap Mass Spectrometry for Quantitative Steroid Hormone Analysis in Plasma, Urine, Saliva and Hair. J. Chromatogr. A 2016, 1464, 64–71. [Google Scholar] [CrossRef]
- Tan, A.; Wu, Y.; Wong, M.; Licollari, A.; Bolger, G.; Fanaras, J.C.; Shopp, G.; Helson, L. Use of Basic Mobile Phase to Improve Chromatography and Boost Sensitivity for Quantifying Tetrahydrocurcumin in Human Plasma by LC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1028, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Gu, G.; Gui, X.; Wu, Y.; Bolger, G.; Licollari, A.; Fanaras, J.C. Improving the Selectivity and Sensitivity for Quantifying 8-α-Hydroxy-Mutilin in Rabbit Tissues by Using Basic Mobile Phases and Negative Ionization. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1072, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Nagasawa, E.; Uchida, T.; Sakamaki, H. Highly Sensitive Detection of Pesticide Residues by LC-MS/MS Using Ammonia Solution as an Eluent. Bunseki Kagaku 2020, 69, 141–150. [Google Scholar] [CrossRef]
- Lewis, K.M.; Archer, R.D. PKa Values of Estrone, 17β-Estradiol and 2-Methoxyestrone. Steroids 1979, 34, 485–499. [Google Scholar] [CrossRef]
- Nouri, M.Z.; Kroll, K.J.; Webb, M.; Denslow, N.D. Quantification of Steroid Hormones in Low Volume Plasma and Tissue Homogenates of Fish Using LC-MS/MS. Gen. Comp. Endocrinol. 2020, 296, 113543. [Google Scholar] [CrossRef]
- Maekawa, M.; Jinnoh, I.; Narita, A.; Iida, T.; Saigusa, D.; Iwahori, A.; Nittono, H.; Okuyama, T.; Eto, Y.; Ohno, K.; et al. Investigation of Diagnostic Performance of Five Urinary Cholesterol Metabolites for Niemann-Pick Disease Type C. J. Lipid Res. 2019, 60, 2074–2081. [Google Scholar] [CrossRef]
- Iwahori, A.; Maekawa, M.; Narita, A.; Kato, A.; Sato, T.; Ogura, J.; Sato, Y.; Kikuchi, M.; Noguchi, A.; Higaki, K.; et al. Development of a Diagnostic Screening Strategy for Niemann–Pick Diseases Based on Simultaneous Liquid Chromatography-Tandem Mass Spectrometry Analyses of N-Palmitoyl-O-Phosphocholine-Serine and Sphingosylphosphorylcholine. Biol. Pharm. Bull. 2020, 43, 1398–1406. [Google Scholar] [CrossRef]
- Maekawa, M.; Misawa, Y.; Sotoura, A.; Yamaguchi, H.; Togawa, M.; Ohno, K.; Nittono, H.; Kakiyama, G.; Iida, T.; Hofmann, A.F.; et al. LC/ESI-MS/MS Analysis of Urinary 3β-Sulfooxy-7β-N-Acetylglucosaminyl-5-Cholen-24-Oic Acid and Its Amides: New Biomarkers for the Detection of Niemann–Pick Type C Disease. Steroids 2013, 78, 967–972. [Google Scholar] [CrossRef]
- Maekawa, M.; Narita, A.; Jinnoh, I.; Iida, T.; Marquardt, T.; Mengel, E.; Eto, Y.; Clayton, P.T.; Yamaguchi, H.; Mano, N. Diagnostic Performance Evaluation of Sulfate-Conjugated Cholesterol Metabolites as Urinary Biomarkers of Niemann–Pick Disease Type C. Clin. Chim. Acta 2019, 494, 58–63. [Google Scholar] [CrossRef]
- Shoji, S.; Maekawa, M.; Ogura, J.; Sato, T.; Mano, N. Identification Cholesterol Metabolites Altered before the Onset of Nonalcoholic Steatohepatitis by Targeted Metabolomics. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2022, 1867, 159135. [Google Scholar] [CrossRef]
- Welford, R.W.D.; Garzotti, M.; Marques Lourenço, C.; Mengel, E.; Marquardt, T.; Reunert, J.; Amraoui, Y.; Kolb, S.A.; Morand, O.; Groenen, P. Plasma Lysosphingomyelin Demonstrates Great Potential as a Diagnostic Biomarker for Niemann-Pick Disease Type C in a Retrospective Study. PLoS ONE 2014, 9, e114669. [Google Scholar] [CrossRef] [PubMed]
- Sobrido, M.J.; Bauer, P.; de Koning, T.; Klopstock, T.; Nadjar, Y.; Patterson, M.C.; Synofzik, M.; Hendriksz, C.J. Recommendations for Patient Screening in Ultra-Rare Inherited Metabolic Diseases: What Have We Learned from Niemann-Pick Disease Type C? Orphanet J. Rare Dis. 2019, 14, 20. [Google Scholar] [CrossRef]
- Feng, X.; Cozma, C.; Pantoom, S.; Hund, C.; Iwanov, K.; Petters, J.; Völkner, C.; Bauer, C.; Vogel, F.; Bauer, P.; et al. Determination of the Pathological Features of NPC1 Variants in a Cellular Complementation Test. Int. J. Mol. Sci. 2019, 20, 5185. [Google Scholar] [CrossRef] [PubMed]
- Pentchev, P.G.; Boothe, A.D.; Kurth, H.S. A Genetic Storage Disorder in BALB/C Mice with a Metabolic Block in Esterification of Exogenous Cholesterol. J. Biol. Chem. 1984, 259, 5784–5791. [Google Scholar] [CrossRef]
- Tint, G.S.; Pentchev, P.; Xu, G.; Batta, A.K.; Shefer, S.; Salen, G.; Honda, A. Cholesterol and Oxygenated Cholesterol Concentrations Are Markedly Elevated in Peripheral Tissue but Not in Brain from Mice with the Niemann-Pick Type C Phenotype. J. Inherit. Metab. Dis. 1998, 21, 853–863. [Google Scholar] [CrossRef]
- Pentchev, P.G.; Comly, M.E.; Kruth, H.S.; Vanier, M.T.; Wenger, D.A.; Patel, S.; Brady, R.O. A Defect in Cholesterol Esterification in Niemann-Pick Disease (Type C) Patients. Proc. Natl. Acad. Sci. USA 1985, 82, 8247–8251. [Google Scholar] [CrossRef]
- Vanier, M.T.; Latour, P. Laboratory Diagnosis of Niemann-Pick Disease Type C: The Filipin Staining Test. Methods Cell Biol. 2015, 126, 357–375. [Google Scholar] [CrossRef]
- Liedtke, M.; Völkner, C.; Hermann, A.; Frech, M.J. Impact of Organelle Transport Deficits on Mitophagy and Autophagy in Niemann–Pick Disease Type C. Cells 2022, 11, 507. [Google Scholar] [CrossRef]
- Davis, O.B.; Shin, H.R.; Lim, C.Y.; Wu, E.Y.; Kukurugya, M.; Maher, C.F.; Perera, R.M.; Ordonez, M.P.; Zoncu, R. NPC1-MTORC1 Signaling Couples Cholesterol Sensing to Organelle Homeostasis and Is a Targetable Pathway in Niemann-Pick Type C. Dev. Cell 2021, 56, 260–276.e7. [Google Scholar] [CrossRef]
- Hecker, M.; Hollert, H.; Cooper, R.; Vinggaard, A.M.; Akahori, Y.; Murphy, M.; Nellemann, C.; Higley, E.; Newsted, J.; Laskey, J.; et al. The OECD Validation Program of the H295R Steroidogenesis Assay: Phase 3. Final Inter-Laboratory Validation Study. Environ. Sci. Pollut. Res. 2011, 18, 503–515. [Google Scholar] [CrossRef]
- Weisser, J.J.; Hansen, C.H.; Poulsen, R.; Larsen, L.W.; Cornett, C.; Styrishave, B. Two Simple Cleanup Methods Combined with LC-MS/MS for Quantification of Steroid Hormones in In Vivo and In Vitro Assays. Anal. Bioanal. Chem. 2016, 408, 4883–4895. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Srivastava, S.; Burek, M.; Förster, C.Y.; Roy, P. Assessment of Estradiol-Induced Gene Regulation and Proliferation in an Immortalized Mouse Immature Sertoli Cell Line. Life Sci. 2016, 148, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Berthois, Y.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Phenol Red in Tissue Culture Media Is a Weak Estrogen: Implications Concerning the Study of Estrogen-Responsive Cells in Culture. Proc. Natl. Acad. Sci. USA 1986, 83, 2496–2500. [Google Scholar] [CrossRef] [PubMed]
- Strajhar, P.; Tonoli, D.; Jeanneret, F.; Imhof, R.M.; Malagnino, V.; Patt, M.; Kratschmar, D.v.; Boccard, J.; Rudaz, S.; Odermatt, A. Steroid Profiling in H295R Cells to Identify Chemicals Potentially Disrupting the Production of Adrenal Steroids. Toxicology 2017, 381, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Jee, J.P.; Jin, S.E.; Ban, E.; Lee, H.J.; Park, Y.; Park, Y.; Maeng, H.J.; Kim, H.T.; Kim, C.K. Isolation and Identification of Steroidogenic Peptides from Calf Spleen. Arch. Pharm. Res. 2012, 35, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.E.M.; Frøysa, H.G.; Karlsen, O.A.; Sagen, J.v.; Mellgren, G.; Verhaegen, S.; Ropstad, E.; Goksøyr, A.; Kellmann, R. LC-MS/MS Based Profiling and Dynamic Modelling of the Steroidogenesis Pathway in Adrenocarcinoma H295R Cells. Toxicol. Vitr. 2018, 52, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Kurlbaum, M.; Sbiera, S.; Kendl, S.; Martin Fassnacht, M.; Kroiss, M. Steroidogenesis in the NCI-H295 Cell Line Model Is Strongly Affected by Culture Conditions and Substrain. Exp. Clin. Endocrinol. Diabetes 2020, 128, 672–680. [Google Scholar] [CrossRef]
- Winther, C.S.; Nielsen, F.K.; Hansen, M.; Styrishave, B. Corticosteroid Production in H295R Cells during Exposure to 3 Endocrine Disrupters Analyzed with LC-MS/MS. Int. J. Toxicol. 2013, 32, 219–227. [Google Scholar] [CrossRef]
- Munkboel, C.H.; Baake, M.L.K.; Styrishave, B. Atorvastatin Decreases Steroid Production in H295R Cells and in Major Endocrine Tissues of Male Rats. Arch. Toxicol. 2018, 92, 1703–1715. [Google Scholar] [CrossRef]
- Machioka, K.; Mizokami, A.; Yamaguchi, Y.; Izumi, K.; Hayashi, S.; Namiki, M. Active Estrogen Synthesis and Its Function in Prostate Cancer-Derived Stromal Cells. Anticancer Res. 2015, 35, 221–228. [Google Scholar]
- Hansen, C.H.; Larsen, L.W.; Sørensen, A.M.; Halling-Sørensen, B.; Styrishave, B. The Six Most Widely Used Selective Serotonin Reuptake Inhibitors Decrease Androgens and Increase Estrogens in the H295R Cell Line. Toxicol. In Vitro 2017, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Blaschke, M.; Koepp, R.; Streit, F.; Beismann, J.; Manthey, G.; Seitz, M.T.; Kragl, A.; Siggelkow, H. The Rise in Expression and Activity of 11β-HSD1 in Human Mesenchymal Progenitor Cells Induces Adipogenesis through Increased Local Cortisol Synthesis. J. Steroid Biochem. Mol. Biol. 2021, 210, 105850. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, J.; Maeda, N.; Sato, M.; Ieko, T.; Inoue, H.; Iwasaki, T.; Iwano, H.; Yokota, H. Corticosterone Biosynthesis in Mouse Clonal Myoblastic C2C12 Cells. Steroids 2018, 138, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Koal, T.; Schmiederer, D.; Pham-Tuan, H.; Röhring, C.; Rauh, M. Standardized LC-MS/MS Based Steroid Hormone Profile-Analysis. J. Steroid Biochem. Mol. Biol. 2012, 129, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Cheng, Z.; Peng, Y.; Wang, Z.; Huang, C.; Liu, D.; Wang, B.; Pan, B.; Guo, W. A Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)-Based Assay for Simultaneous Quantification of Aldosterone, Renin Activity, and Angiotensin II in Human Plasma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2021, 1179, 122740. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Geller, D.H.; Rosen, M. Ovarian and Adrenal Androgen Biosynthesis and Metabolism. In Androgen Excess Disorders in Women; Humana Press: Totowa, NJ, USA, 2007; pp. 19–33. [Google Scholar]
- Illingworth, D.R.; Corbin, D.K.; Kemp, E.D.; Keenan, E.J. Hormone Changes during the Menstrual Cycle in Abetalipoproteinemia: Reduced Luteal Phase Progesterone in a Patient with Homozygous Hypobetalipoproteinemia. Proc. Natl. Acad. Sci. USA 1982, 79, 6685–6689. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenesis: Unanswered Questions. Trends Endocrinol. Metab. 2017, 28, 771–793. [Google Scholar] [CrossRef]
- Kallen, C.B.; Arakane, F.; Christenson, L.K.; Watari, H.; Devoto, L.; Strauss, J.F. Unveiling the Mechanism of Action and Regulation of the Steroidogenic Acute Regulatory Protein. Mol. Cell. Endocrinol. 1998, 145, 39–45. [Google Scholar] [CrossRef]
- Chien, Y.; Rosal, K.; Chung, B. chu Function of CYP11A1 in the Mitochondria. Mol. Cell. Endocrinol. 2017, 441, 55–61. [Google Scholar] [CrossRef]
- Simard, J.; Ricketts, M.-L.; Gingras, S.; Soucy, P.; Feltus, F.A.; Melner, M.H. Molecular Biology of the 3beta-Hydroxysteroid Dehydrogenase/Delta5-Delta4 Isomerase Gene Family. Endocr. Rev. 2005, 26, 525–582. [Google Scholar] [CrossRef]
- Akhtar, M.; Wright, J.N.; Lee-Robichaud, P. A Review of Mechanistic Studies on Aromatase (CYP19) and 17α-Hydroxylase-17,20-Lyase (CYP17). J. Steroid Biochem. Mol. Biol. 2011, 125, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M.; Clark, B.J.; Prough, R.A. Dehydroepiandrosterone Research: Past, Current, and Future, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 108, ISBN 9780128143612. [Google Scholar]
- Meng, L.; Yu, H.; Qu, J.; Niu, J.; Ni, F.; Han, P.; Wang, X. Two Cyp17 Genes Perform Different Functions in the Sex Hormone Biosynthesis and Gonadal Differentiation in Japanese Flounder (Paralichthys Olivaceus). Gene 2019, 702, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Carmina, E. Ovarian and Adrenal Hyperandrogenism. Ann. N. Y. Acad. Sci. 2006, 1092, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Chatuphonprasert, W.; Jarukamjorn, K.; Ellinger, I. Physiology and Pathophysiology of Steroid Biosynthesis, Transport and Metabolism in the Human Placenta. Front. Pharmacol. 2018, 9, 1027. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yang, S.; Fang, Z.; Gurates, B.; Tamura, M.; Zhou, J.; Sebastian, S. Role of Aromatase in Endometrial Disease. J. Steroid Biochem. Mol. Biol. 2001, 79, 19–25. [Google Scholar] [CrossRef]
- Thomas, M.P.; Potter, B.V.L. The Structural Biology of Oestrogen Metabolism. J. Steroid Biochem. Mol. Biol. 2013, 137, 27–49. [Google Scholar] [CrossRef]
- Walker, M.A.; Tian, R. NAD(H) in Mitochondrial Energy Transduction: Implications for Health and Disease. Curr. Opin. Physiol. 2018, 3, 101–109. [Google Scholar] [CrossRef]
- Allen, J.A.; Shankara, T.; Janus, P.; Buck, S.; Diemer, T.; Hales, K.H.; Hales, D.B. Energized, Polarized, and Actively Respiring Mitochondria Are Required for Acute Leydig Cell Steroidogenesis. Endocrinology 2006, 147, 3924–3935. [Google Scholar] [CrossRef]
- Midzak, A.S.; Chen, H.; Aon, M.A.; Papadopoulos, V.; Zirkin, B.R. ATP Synthesis, Mitochondrial Function, and Steroid Biosynthesis in Rodent Primary and Tumor Leydig Cells 1. Biol. Reprod. 2011, 84, 976–985. [Google Scholar] [CrossRef]
- Romano, S.; Mitro, N.; Giatti, S.; Diviccaro, S.; Pesaresi, M.; Spezzano, R.; Audano, M.; Garcia-Segura, L.M.; Caruso, D.; Melcangi, R.C. Diabetes Induces Mitochondrial Dysfunction and Alters Cholesterol Homeostasis and Neurosteroidogenesis in the Rat Cerebral Cortex. J. Steroid Biochem. Mol. Biol. 2018, 178, 108–116. [Google Scholar] [CrossRef]
- Pesaresi, M.; Maschi, O.; Giatti, S.; Miguel Garcia-Segura, L.; Caruso, D.; Cosimo Melcangi, R. Sex Differences in Neuroactive Steroid Levels in the Nervous System of Diabetic and Non-Diabetic Rats. Horm. Behav. 2009, 57, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Mitro, N.; Diviccaro, S.; Spezzano, R.; Audano, M.; Garcia-Segura, L.M.; Caruso, D.; Melcangi, R.C. Short-Term Effects of Diabetes on Neurosteroidogenesis in the Rat Hippocampus. J. Steroid Biochem. Mol. Biol. 2017, 167, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Meimaridou, E.; Goldsworthy, M.; Chortis, V.; Fragouli, E.; Foster, P.A.; Arlt, W.; Cox, R.; Metherell, L.A. NNT Is a Key Regulator of Adrenal Redox Homeostasis and Steroidogenesis in Male Mice. J. Endocrinol. 2018, 236, 13–28. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The Nuclear Receptor Superfamily: The Second Decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- DeMayo, F.J.; Zhao, B.; Takamoto, N.; Tsai, S.Y. Mechanisms of Action of Estrogen and Progesterone. Ann. N. Y. Acad. Sci. 2002, 955, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Kicman, A.T. Pharmacology of Anabolic Steroids. Br. J. Pharmacol. 2008, 154, 502–521. [Google Scholar] [CrossRef]
- Diaz Brinton, R. The Neurosteroid 3α-Hydroxy-5α-Pregnan-20-One Induces Cytoarchitectural Regression in Cultured Fetal Hippocampal Neurons. J. Neurosci. 1994, 14, 2763–2774. [Google Scholar] [CrossRef]
- Concas, A.; Mostallino, M.C.; Porcu, P.; Follesa, P.; Barbaccia, M.L.; Trabucchi, M.; Purdy, R.H.; Grisenti, P.; Biggio, G. Role of Brain Allopregnanolone in the Plasticity of γ-Aminobutyric Acid Type A Receptor in Rat Brain during Pregnancy and after Delivery. Proc. Natl. Acad. Sci. USA 1998, 95, 13284–13289. [Google Scholar] [CrossRef]
- Brussaard, A.B.; Wossink, J.; Lodder, J.C.; Kits, K.S. Progesterone-Metabolite Prevents Protein Kinase c-Dependent Modulation of γ-Aminobutyric Acid Type A Receptors in Oxytocin Neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 3625–3630. [Google Scholar] [CrossRef]
- Reddy, D.S. Neurosteroids. Endogenous Role in the Human Brain and Therapeutic Potentials. In Progress in Brain Research; Elsevier B.V.: Amsterdam, The Netherlands, 2010; Volume 186, pp. 113–137. [Google Scholar]
- Tsutsui, K.; Ukena, K.; Sakamoto, H.; Okuyama, S.I.; Haraguchi, S. Biosynthesis, Mode of Action, and Functional Significance of Neurosteroids in the Purkinje Cell. Front. Endocrinol. 2011, 2, 61. [Google Scholar] [CrossRef]
- Ko, D.C.; Milenkovic, L.; Beier, S.M.; Manuel, H.; Buchanan, J.A.; Scott, M.P. Cell-Autonomous Death of Cerebellar Purkinje Neurons with Autophagy in Niemann-Pick Type C Disease. PLoS Genet. 2005, 1, e7. [Google Scholar] [CrossRef]
- Kawazoe, T.; Yamamoto, T.; Narita, A.; Ohno, K.; Adachi, K.; Nanba, E.; Noguchi, A.; Takahashi, T.; Maekawa, M.; Eto, Y.; et al. Phenotypic Variability of Niemann-Pick Disease Type C Including a Case with Clinically Pure Schizophrenia: A Case Report. BMC Neurol. 2018, 18, 117. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, M.; Jinnoh, I.; Matsumoto, Y.; Narita, A.; Mashima, R.; Takahashi, H.; Iwahori, A.; Saigusa, D.; Fujii, K.; Abe, A.; et al. Structural Determination of Lysosphingomyelin-509 and Discovery of Novel Class Lipids from Patients with Niemann-Pick Disease Type C. Int. J. Mol. Sci. 2019, 20, 5018. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | Q1 (m/z) | Precursor Ion Form | Q3 (m/z) | DP (V) | CE (V) | CXP (V) | EP (V) | Polarity | Retention Time (min) |
|---|---|---|---|---|---|---|---|---|---|
| Testosterone | 289.0 | [M + H]+ | 97.1 | 100 | 8 | 29 | 6 | Positive | 7.1 |
| Androsterone | 273.1 | [M − H2O + H]+ | 255.1 | 146 | 8 | 19 | 14 | Positive | 10.1 |
| Epiandrosterone | 273.2 | [M − H2O + H]+ | 255.2 | 101 | 8 | 19 | 14 | Positive | 8.4 |
| DHEA | 271.0 | [M − H2O + H]+ | 253.1 | 81 | 12 | 19 | 4 | Positive | 7.8 |
| Cortisol | 363.0 | [M + H]+ | 120.9 | 100 | 8 | 31 | 8 | Positive | 4.1 |
| Cortisone | 361.1 | [M + H]+ | 163.0 | 100 | 8 | 33 | 10 | Positive | 3.9 |
| Corticosterone | 347.2 | [M + H]+ | 329.0 | 101 | 6 | 21 | 16 | Positive | 5.6 |
| Aldosterone | 361.1 | [M + H]+ | 343.1 | 61 | 8 | 25 | 6 | Positive | 3.3 |
| Pregnenolone | 299.2 | [M − H2O + H]+ | 91.1 | 161 | 8 | 71 | 8 | Positive | 10.6 |
| Progesterone | 315.0 | [M + H]+ | 97.0 | 100 | 8 | 27 | 6 | Positive | 10.3 |
| Estrone | 269.0 | [M − H]− | 144.8 | −80 | −6 | −48 | −9 | Negative | 6.4 |
| Estradiol | 271.0 | [M − H]− | 182.9 | −100 | −15 | −58 | −11 | Negative | 6.1 |
| Estriol | 287.1 | [M − H]− | 170.8 | −140 | −15 | −50 | −11 | Negative | 3.0 |
| Testosterone-2H3 | 292.1 | [M + H]+ | 96.9 | 100 | 8 | 27 | 6 | Positive | 7.1 |
| Androsterone-2H4 | 277.2 | [M − H2O + H]+ | 259.1 | 121 | 8 | 19 | 14 | Positive | 10.0 |
| DHEA-2H6 | 276.9 | [M − H2O + H]+ | 259.0 | 211 | 12 | 25 | 11 | Positive | 7.8 |
| Cortisol-13C3 | 366.2 | [M + H]+ | 123.9 | 101 | 8 | 31 | 8 | Positive | 4.1 |
| Aldosterone-2H7 | 368.3 | [M + H]+ | 350.0 | 91 | 8 | 25 | 16 | Positive | 3.3 |
| Pregnenolone-13C2,2H2 | 303.2 | [M − H2O + H]+ | 91.1 | 161 | 8 | 71 | 8 | Positive | 10.6 |
| Progesterone-2H9 | 324.2 | [M + H]+ | 100.1 | 100 | 8 | 29 | 8 | Positive | 10.2 |
| Estrone-2H4 | 273.0 | [M − H]− | 146.9 | −80 | −6 | −52 | −11 | Negative | 6.4 |
| Estradiol-13C3 | 274.2 | [M − H]− | 186.0 | −100 | −15 | −56 | −9 | Negative | 6.1 |
| Estriol-13C3 | 290.0 | [M − H]− | 173.9 | −140 | −15 | −50 | −11 | Negative | 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abe, A.; Maekawa, M.; Sato, T.; Sato, Y.; Kumondai, M.; Takahashi, H.; Kikuchi, M.; Higaki, K.; Ogura, J.; Mano, N. Metabolic Alteration Analysis of Steroid Hormones in Niemann–Pick Disease Type C Model Cell Using Liquid Chromatography/Tandem Mass Spectrometry. Int. J. Mol. Sci. 2022, 23, 4459. https://doi.org/10.3390/ijms23084459
Abe A, Maekawa M, Sato T, Sato Y, Kumondai M, Takahashi H, Kikuchi M, Higaki K, Ogura J, Mano N. Metabolic Alteration Analysis of Steroid Hormones in Niemann–Pick Disease Type C Model Cell Using Liquid Chromatography/Tandem Mass Spectrometry. International Journal of Molecular Sciences. 2022; 23(8):4459. https://doi.org/10.3390/ijms23084459
Chicago/Turabian StyleAbe, Ai, Masamitsu Maekawa, Toshihiro Sato, Yu Sato, Masaki Kumondai, Hayato Takahashi, Masafumi Kikuchi, Katsumi Higaki, Jiro Ogura, and Nariyasu Mano. 2022. "Metabolic Alteration Analysis of Steroid Hormones in Niemann–Pick Disease Type C Model Cell Using Liquid Chromatography/Tandem Mass Spectrometry" International Journal of Molecular Sciences 23, no. 8: 4459. https://doi.org/10.3390/ijms23084459
APA StyleAbe, A., Maekawa, M., Sato, T., Sato, Y., Kumondai, M., Takahashi, H., Kikuchi, M., Higaki, K., Ogura, J., & Mano, N. (2022). Metabolic Alteration Analysis of Steroid Hormones in Niemann–Pick Disease Type C Model Cell Using Liquid Chromatography/Tandem Mass Spectrometry. International Journal of Molecular Sciences, 23(8), 4459. https://doi.org/10.3390/ijms23084459