
Mechano Growth Factor Accelerates ACL Repair and Improves Cell Mobility of Mechanically Injured Human ACL Fibroblasts by Targeting Rac1-PAK1/2 and RhoA-ROCK1 Pathways


Abstract
:1. Introduction
2. Results
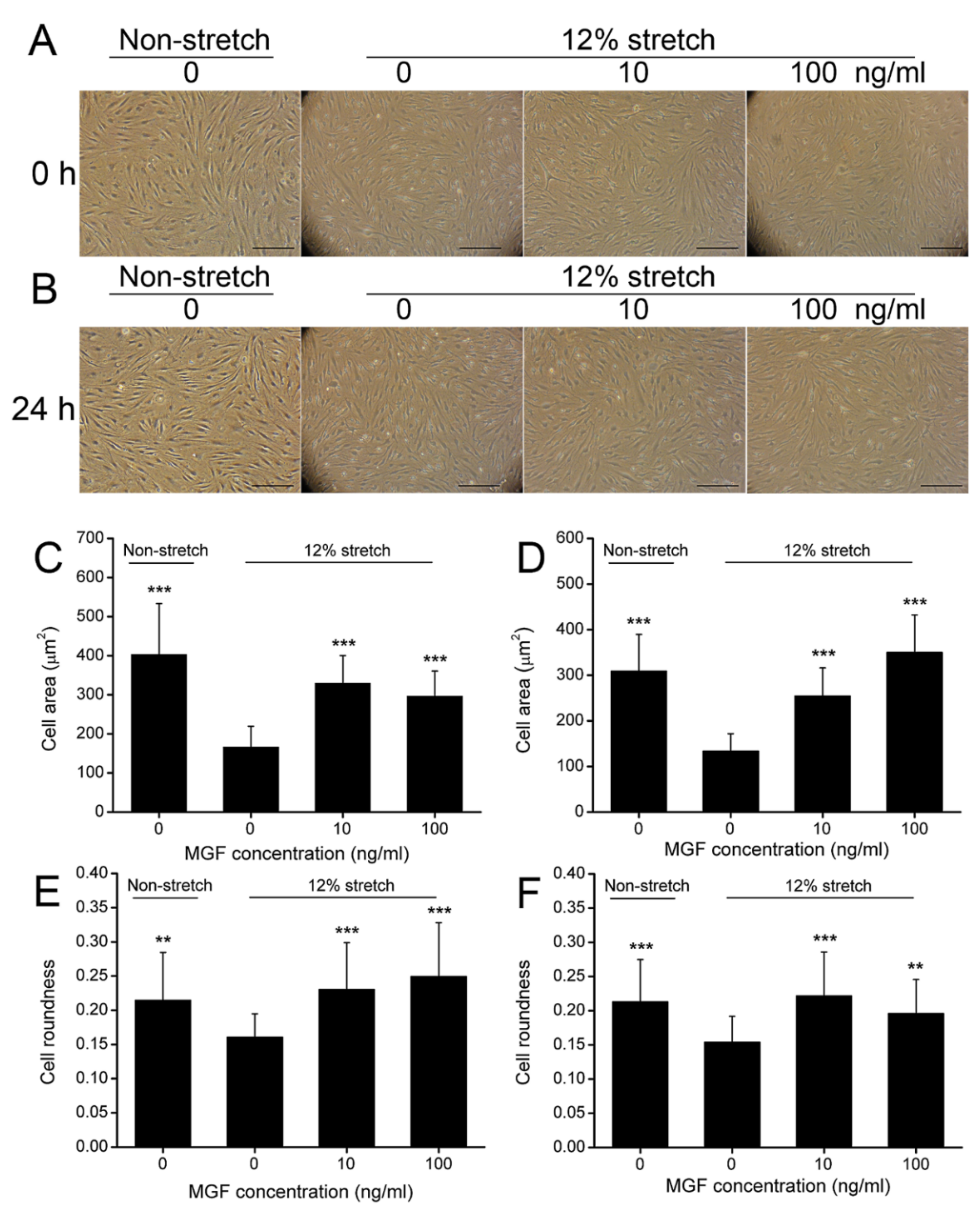
2.1. MGF Improved Cell Deformation of ACL Fibroblasts after Injurious Stretch
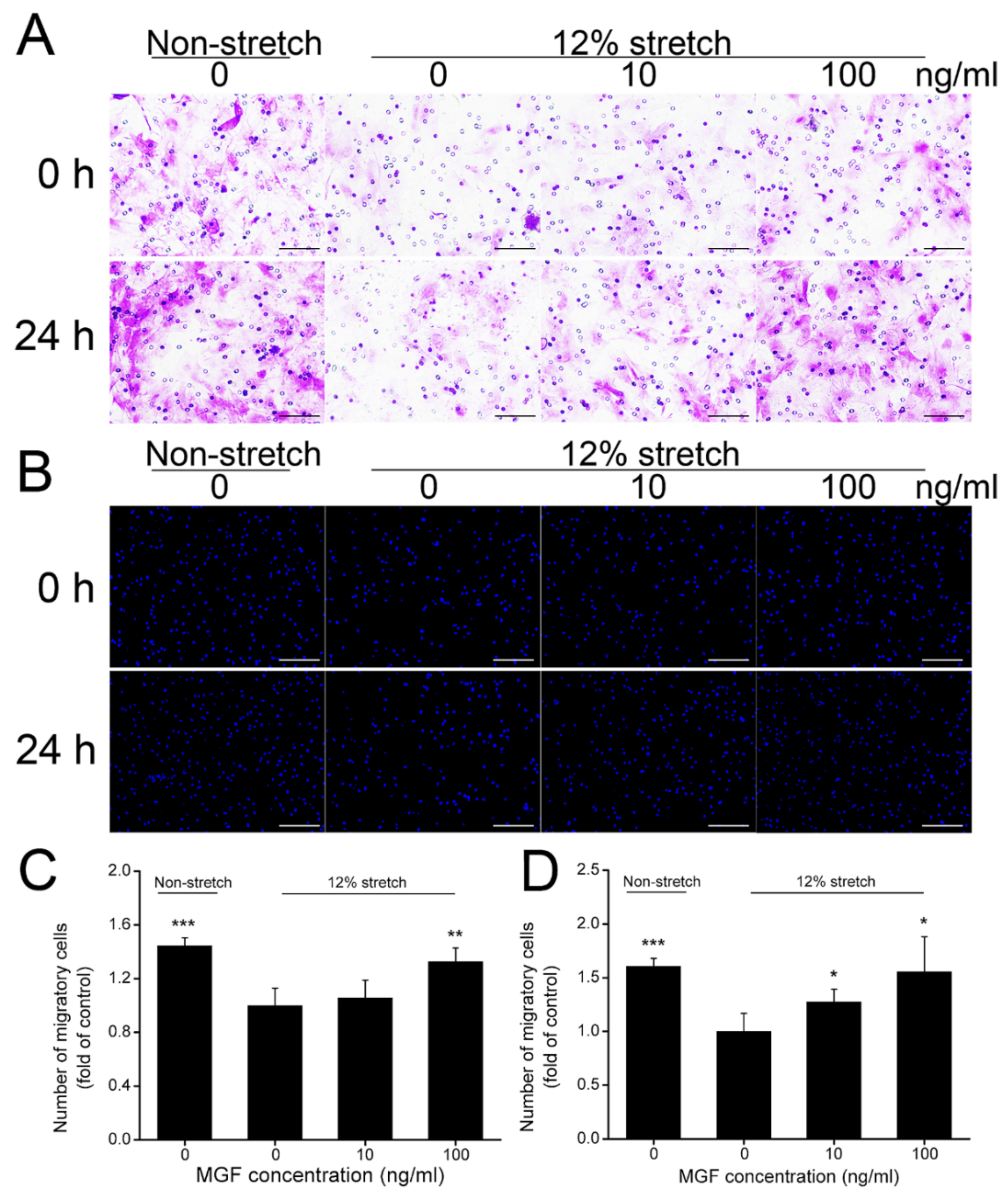
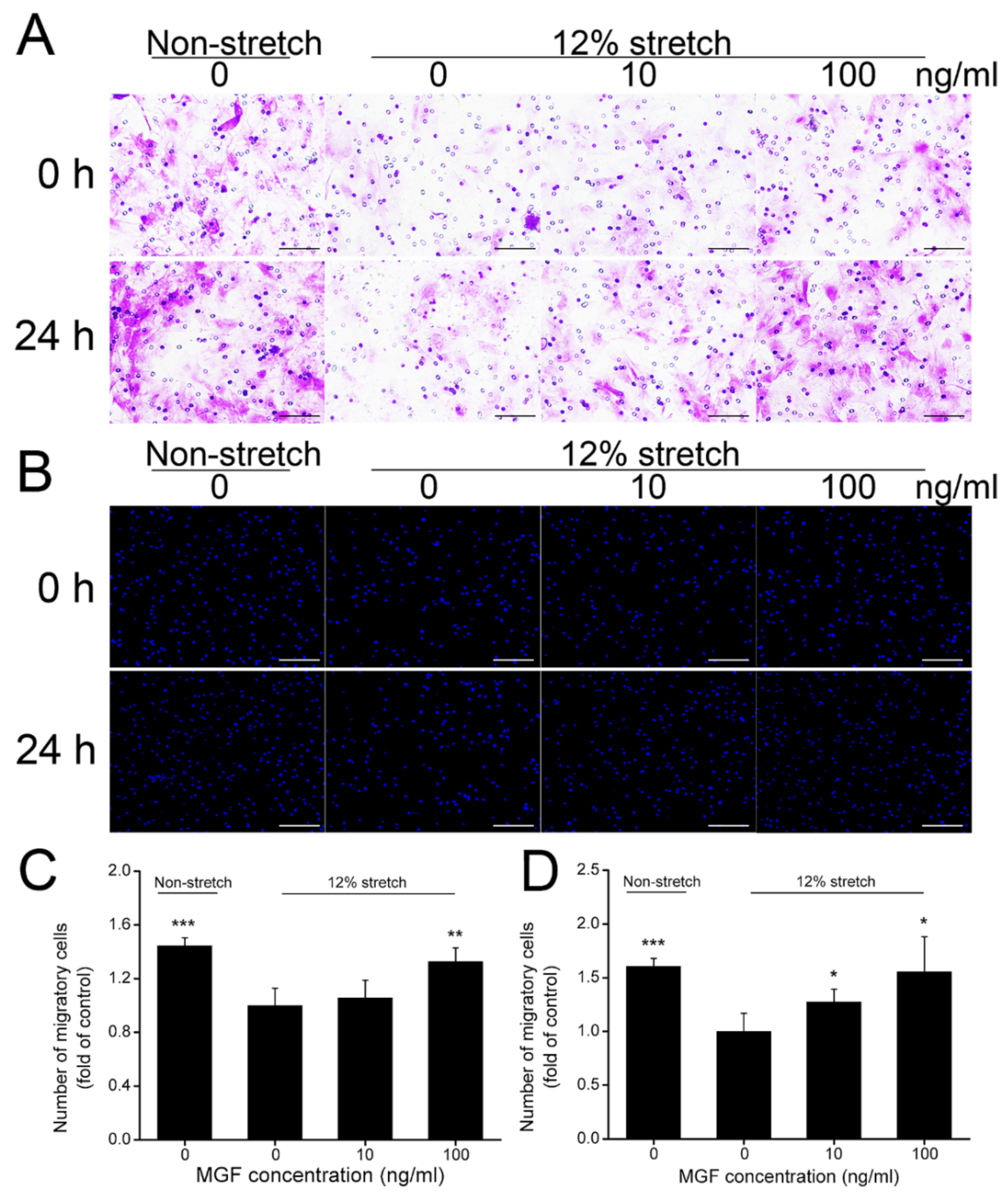
2.2. MGF Improved Cell Migration of ACL Fibroblasts after Injurious Stretch
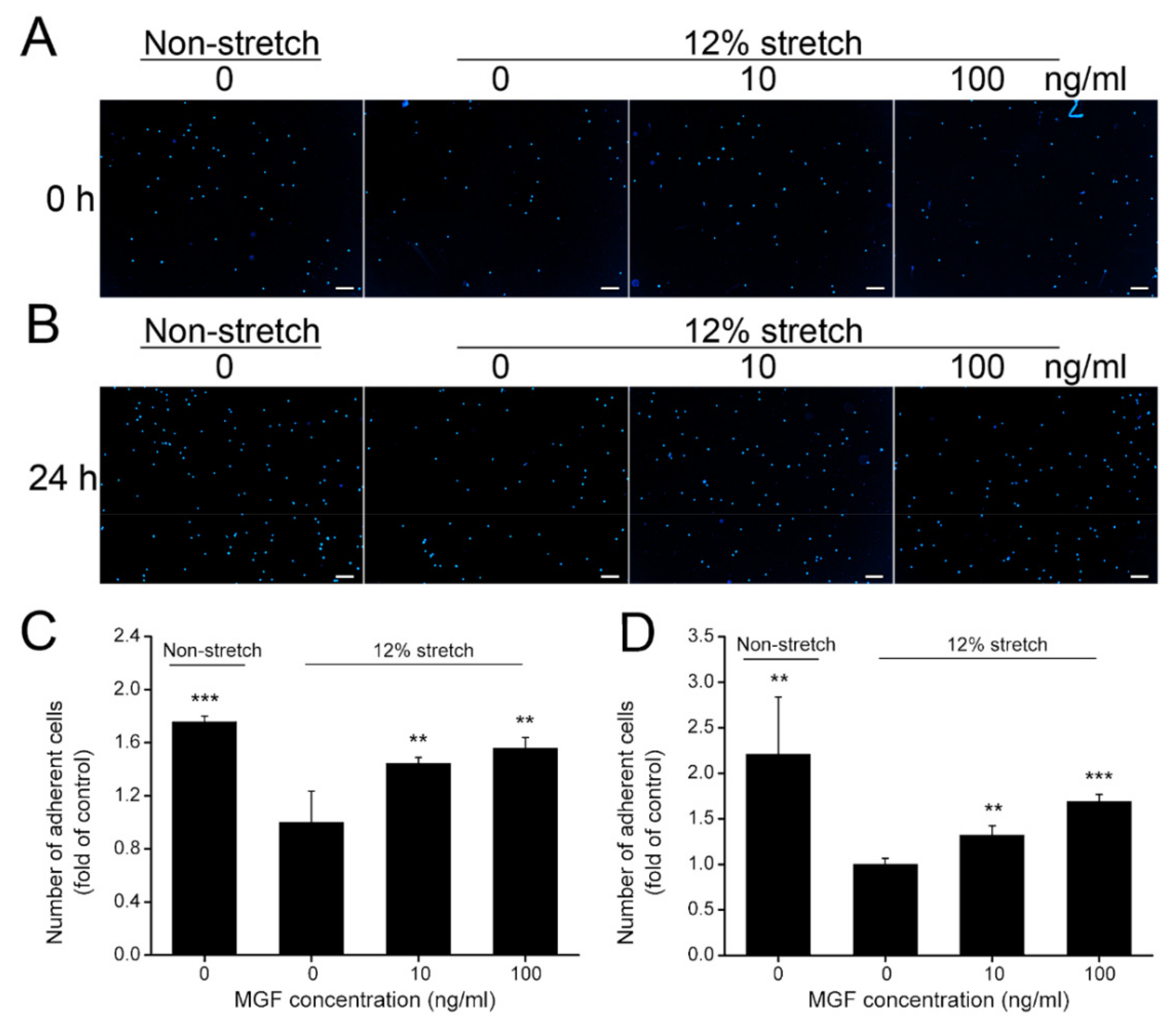
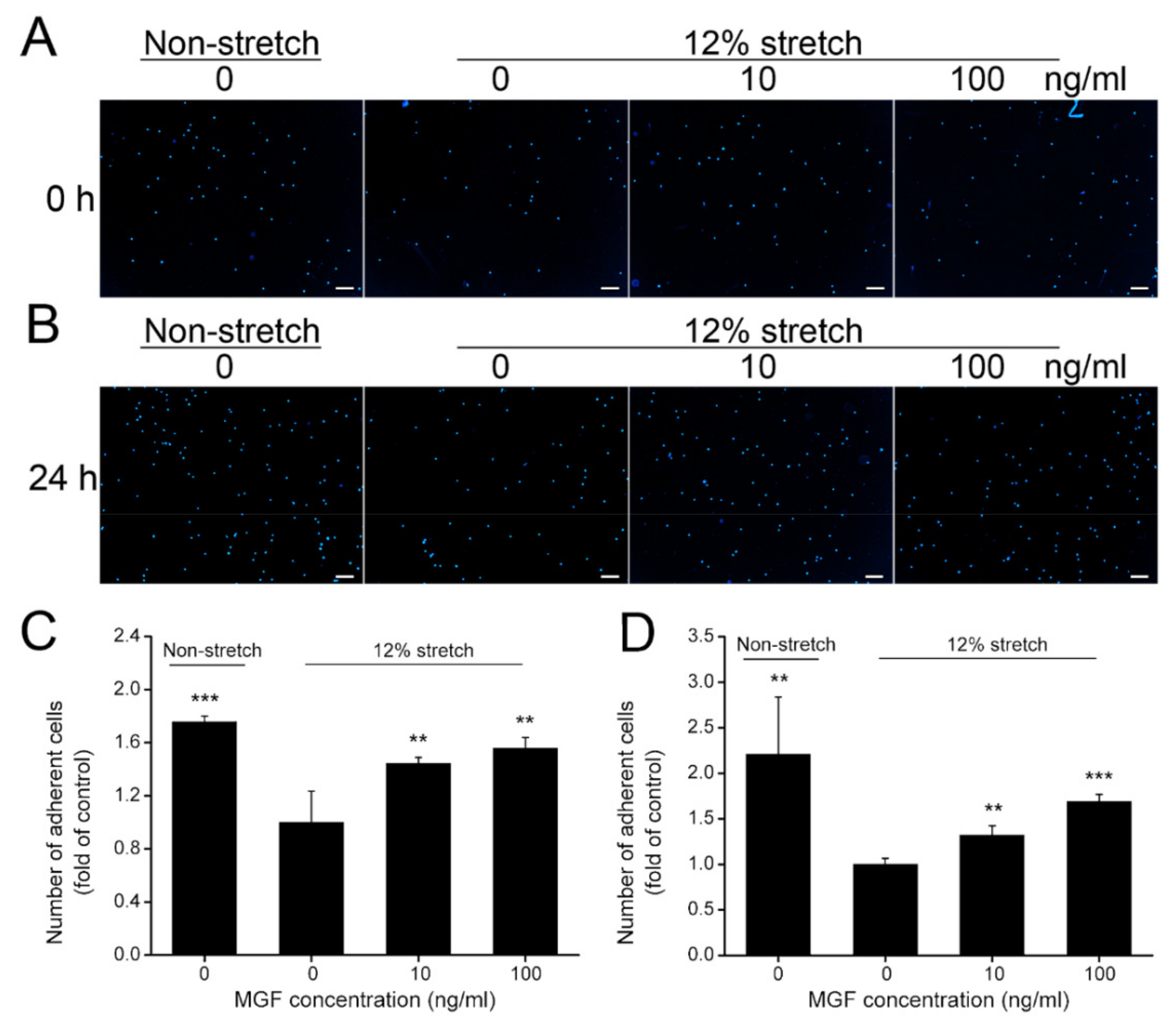
2.3. MGF Increased Cell Adhesion of ACL Fibroblasts after Injurious Stretch
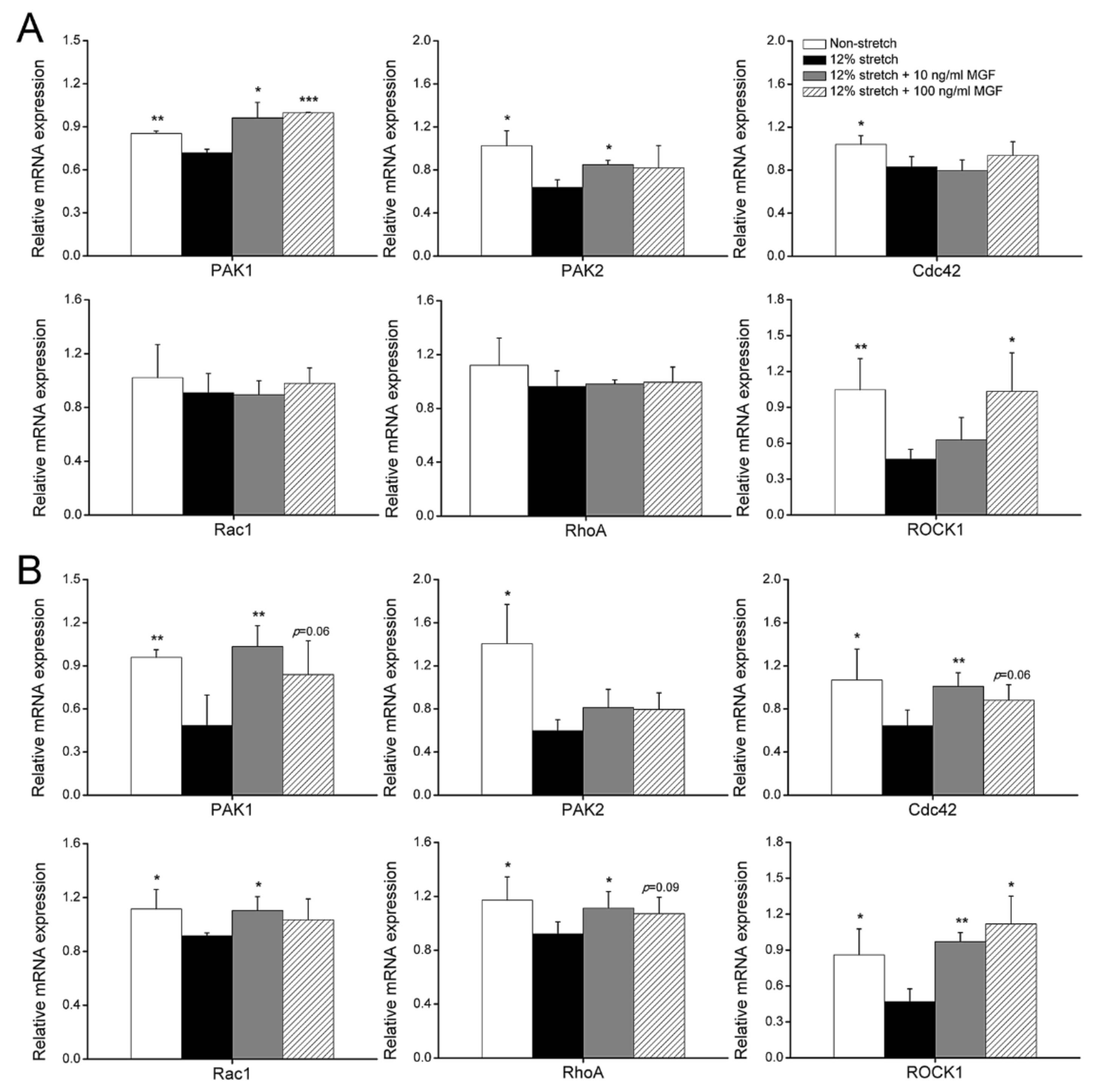
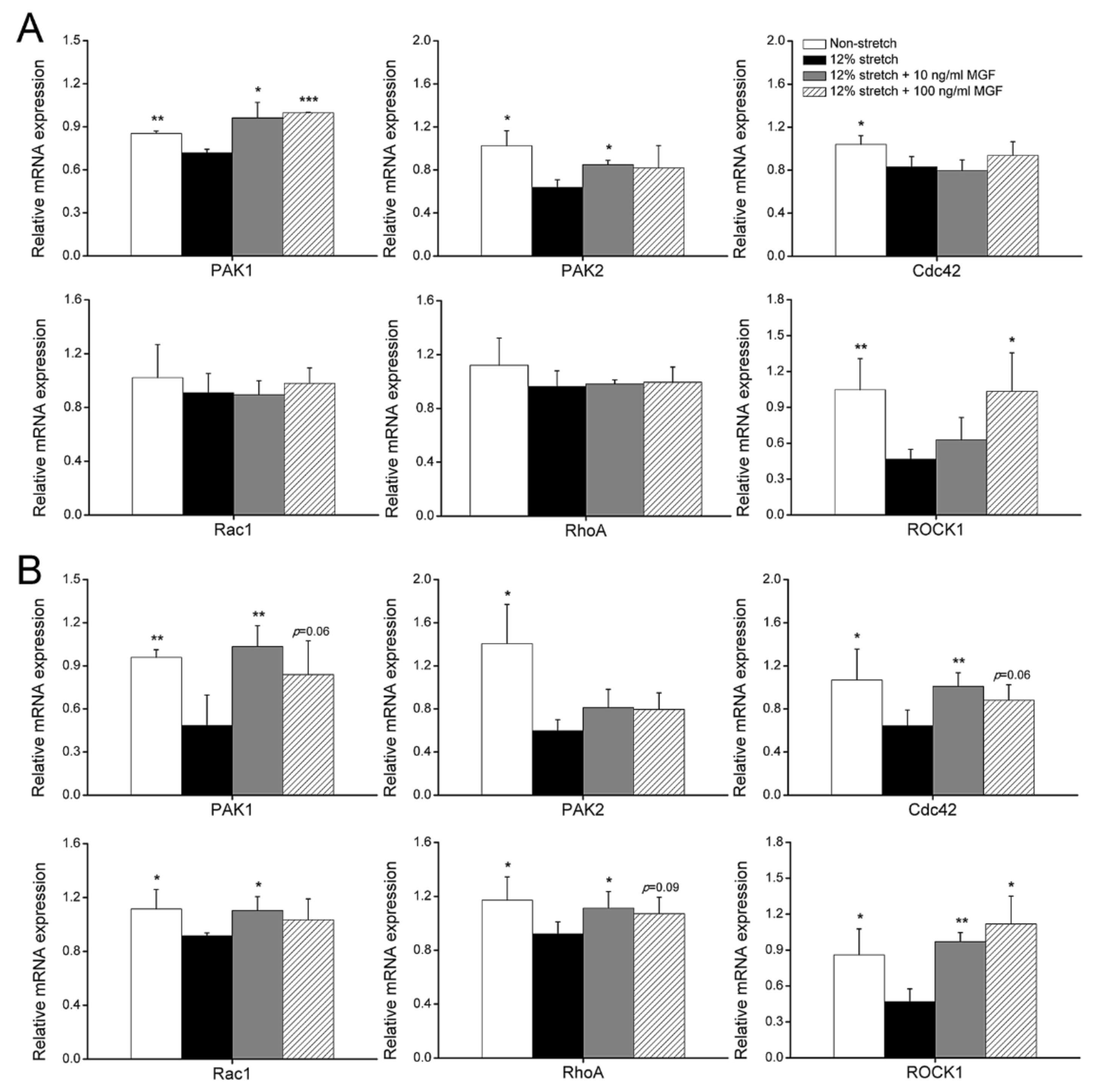
2.4. MGF Regulated Cell Mobility-Relevant Molecules Expression in ACL Fibroblasts after Injurious Stretch
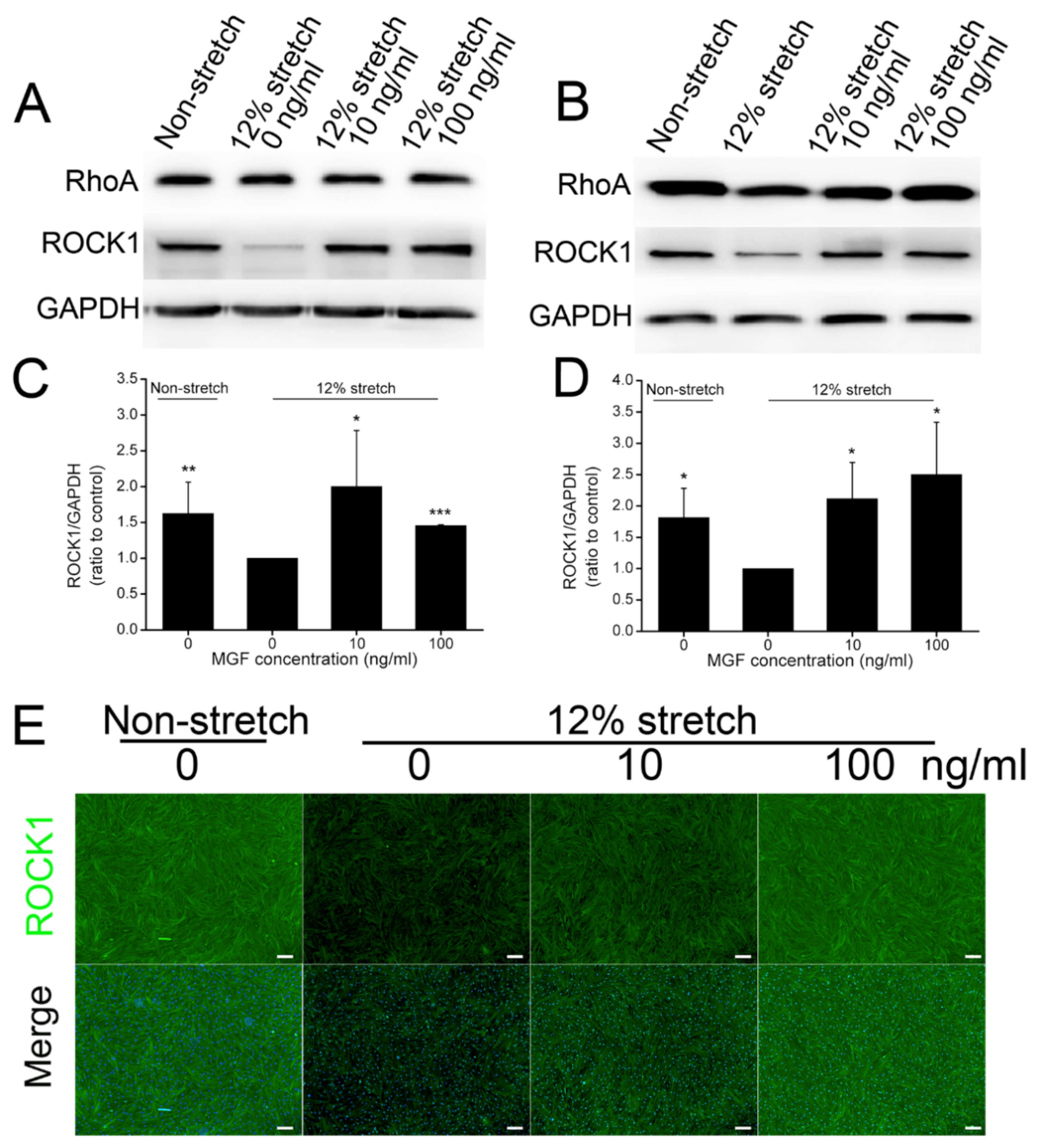
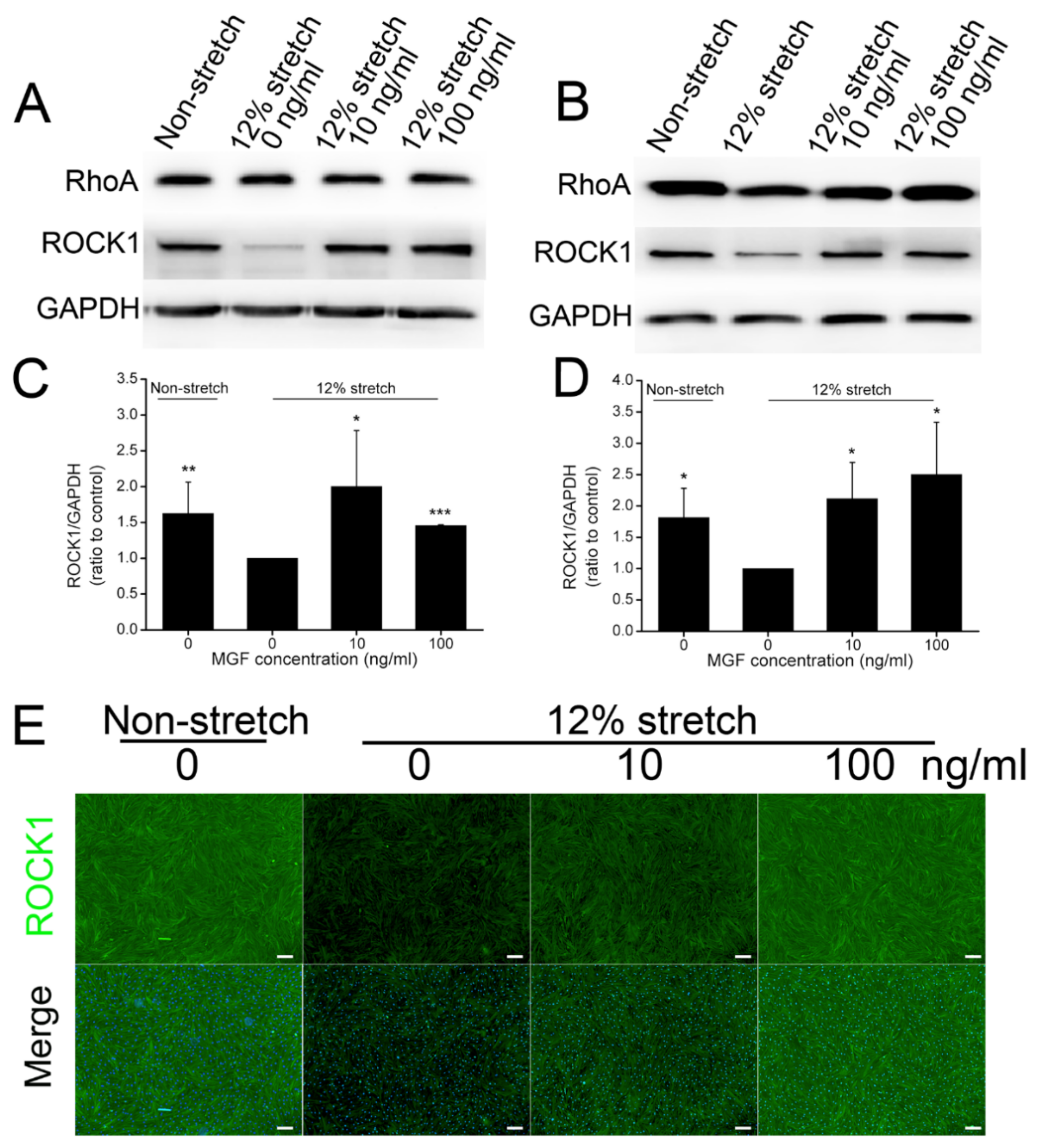
2.5. MGF Regulated ROCK1 Protein Expression in ACL Fibroblasts after Injurious Stretch
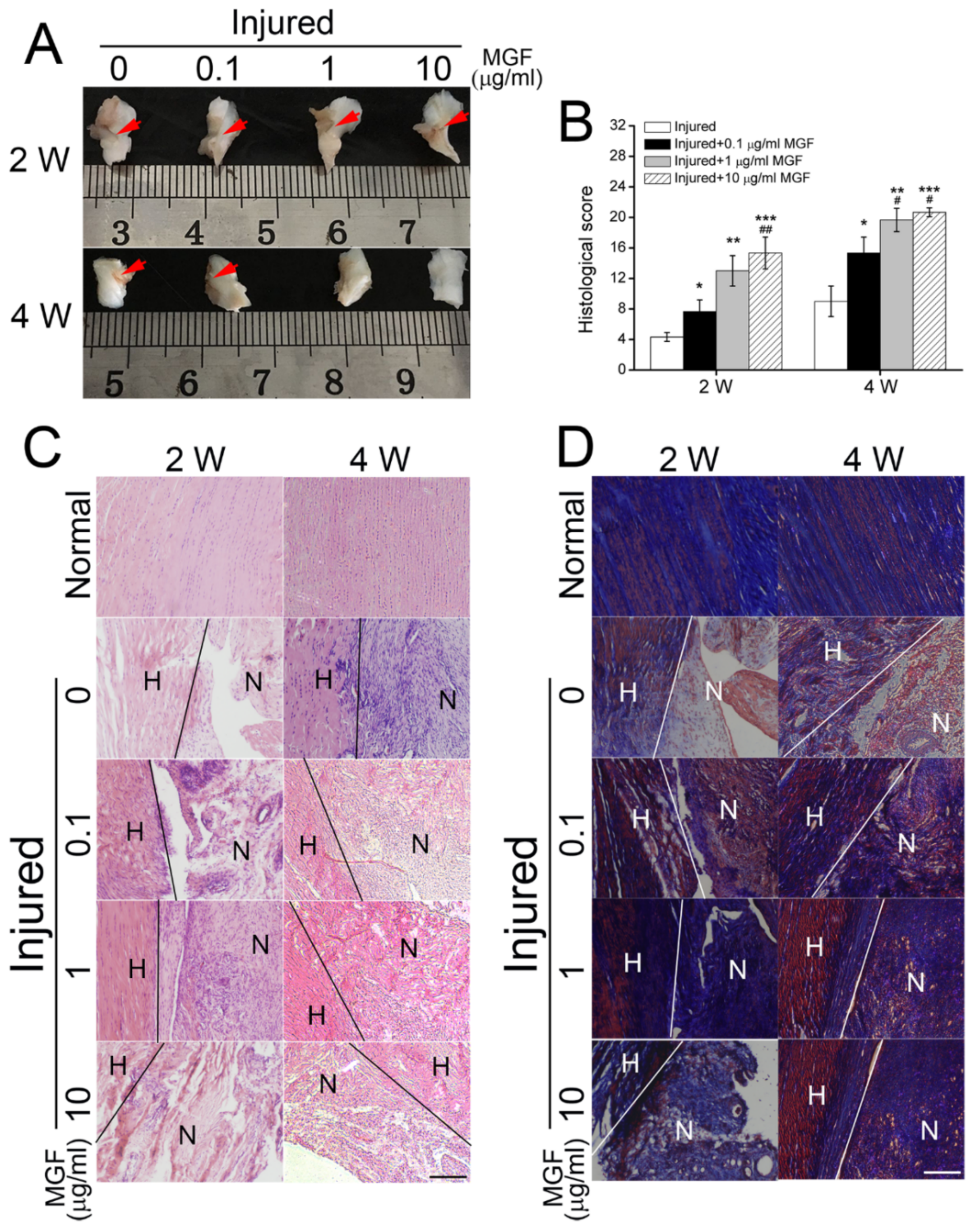
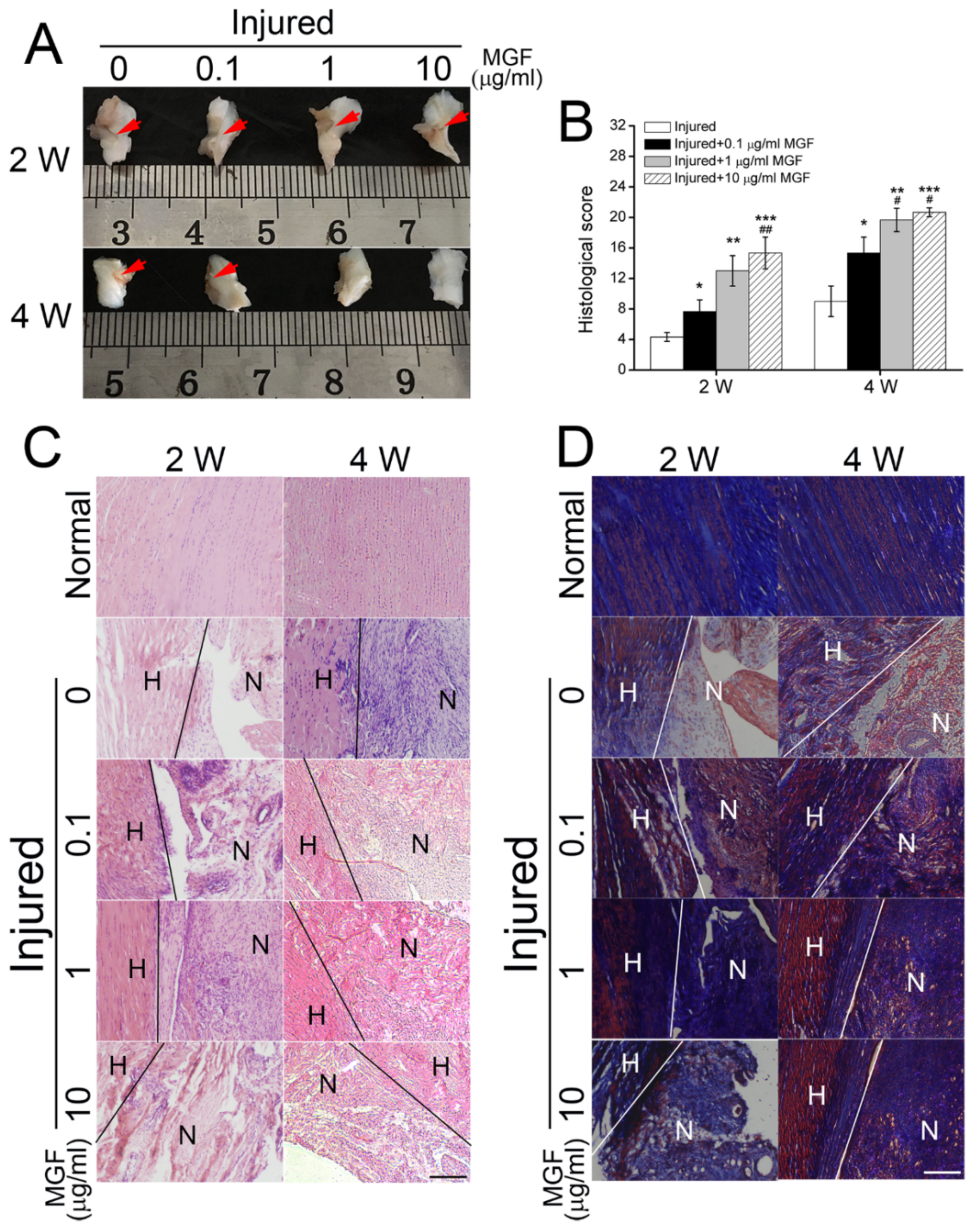
2.6. MGF Accelerated ACL Regeneration after Partial ACLT Surgery
2.7. MGF Improved ECM Synthesis of ACL after Partial ACLT Surgery
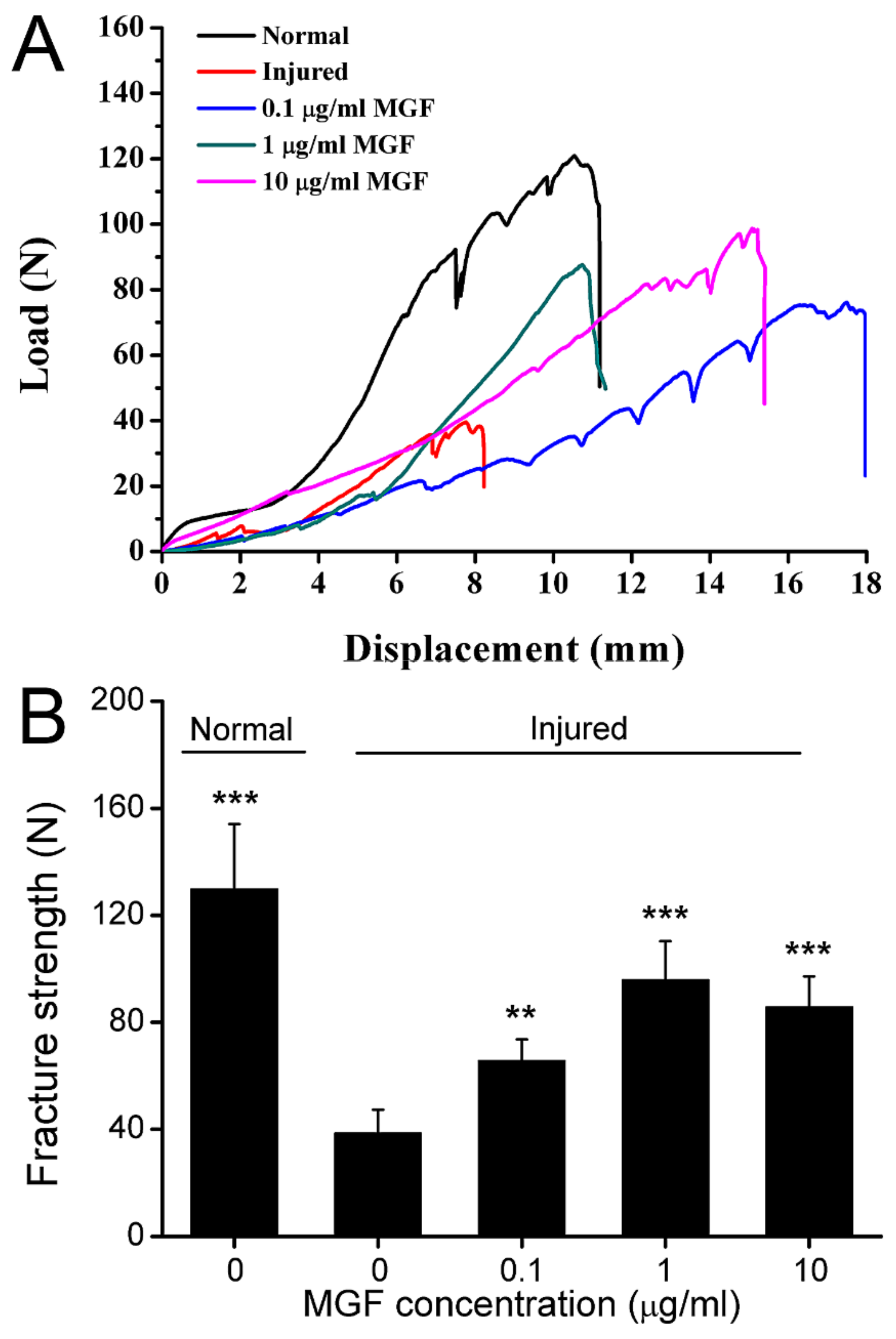
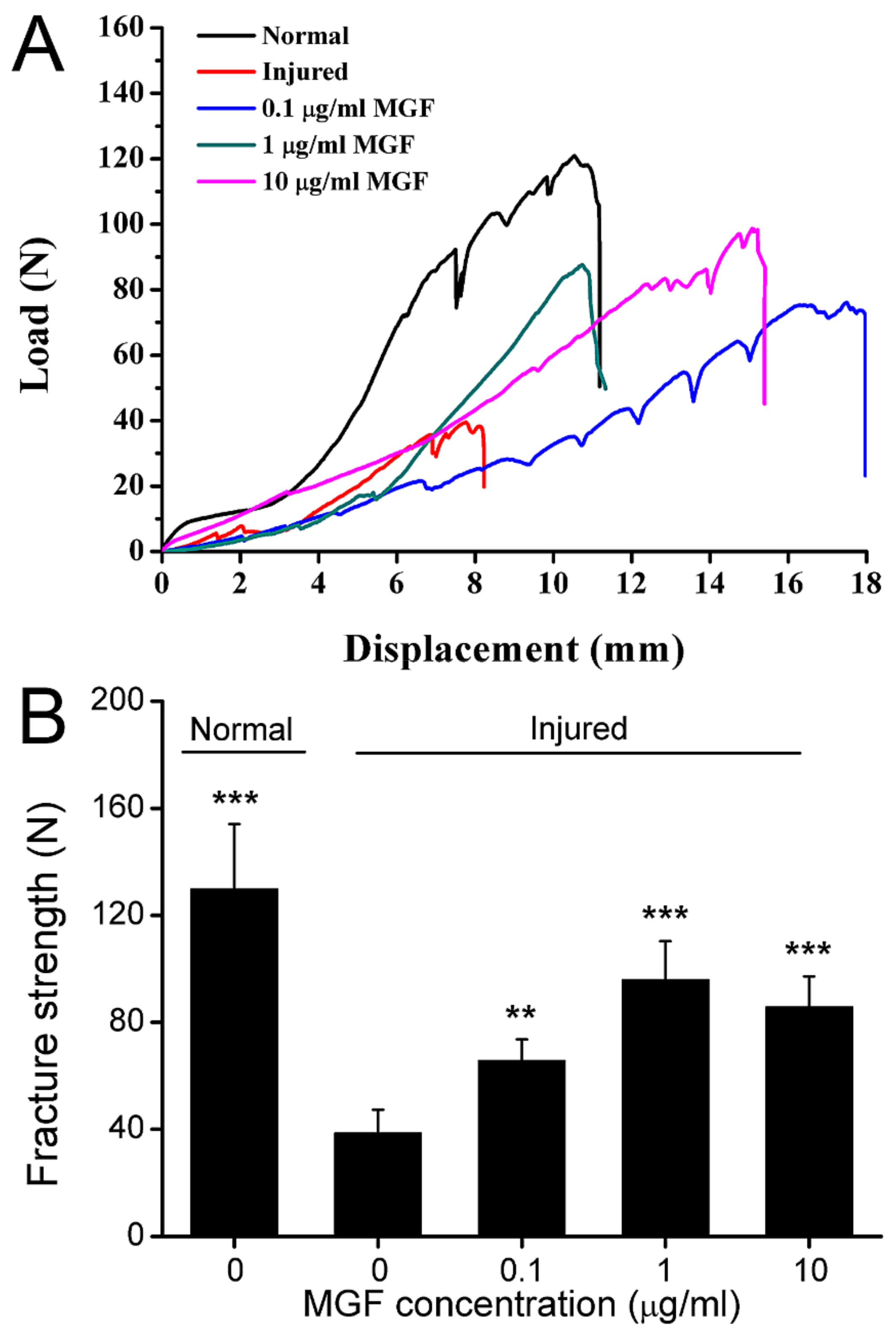
2.8. MGF Improved Mechanical Properties of ACL after Partial ACLT Surgery
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
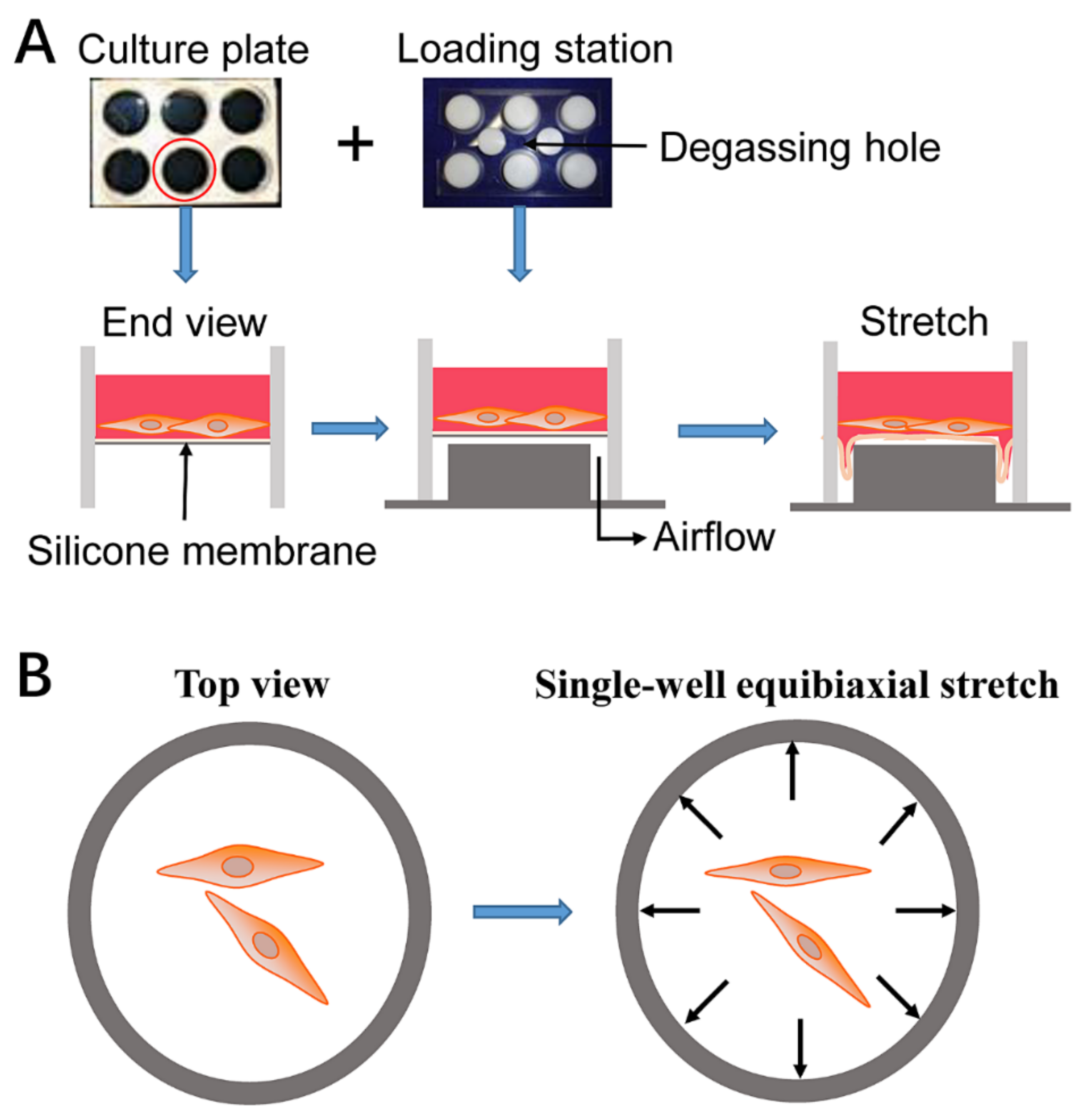
4.2. Cell Stretch
4.3. Cell Area and Roundness
4.4. Cell Migration
4.5. Cell Adhesion
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Western Blotting Assay
4.8. Immunofluorescence Assay
4.9. Rabbit Partial ACL Transection Model
4.10. Hematoxylin-Eosin and Masson Staining
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arianjam, A.; Inacio, M.C.S.; Funahashi, T.T.; Maletis, G.B. Analysis of 2019 patients undergoing anterior cruciate ligament reconstruction from a community-based registry. Am. J. Sports Med. 2017, 45, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pan, T.; Im, H.J.; Fu, F.H.; Wang, J.H. Differential properties of human ACL and MCL stem cells may be responsible for their differential healing capacity. BMC Med. 2011, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Date, H.; Furumatsu, T.; Sakoma, Y.; Yoshida, A.; Hayashi, Y.; Abe, N.; Ozaki, T. GDF-5/7 and bFGF activate intergrin alpha2-mediated cellular migration in rabbit ligament fibroblasts. J. Orthop. Res. 2010, 28, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Q.; Tang, Z.Y.; Xue, R.Y.; Singh, G.K.; Liu, W.Q.; Lv, Y.G.; Yang, L. Differential response to CoCl2-stimulated hypoxia on HIF-1a, VEGF, and MMP-2 expression in ligament cells. Mol. Cell. Biochem. 2012, 360, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Tang, Z.Y.; Xue, R.Y.; Singh, G.K.; Shi, K.N.; Lv, Y.G.; Yang, L. Combined effects of TNF-a, IL-1b, and HIF-1a on MMP-2 production in ACL fibroblasts under mechanical stretch: An in vitro study. J. Orthop. Res. 2011, 20, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Toy, B.J.; Yeasting, R.A.; Morse, D.E.; McCann, P. Arterial supply to the human anterior cruciate ligament. J. Athl. Train. 1995, 30, 149–152. [Google Scholar] [PubMed]
- Seneviratne, A.; Attia, E.; Williams, R.J.; Rodeo, S.A.; Hannafin, J.A. The effect of estrogen on ovine anterior cruciate ligament fibroblasts: Cell proliferation and collagen synthesis. Am. J. Sports Med. 2004, 32, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Chou, S.H.; Shen, P.C.; Chou, P.H.; Ho, M.L.; Tien, Y.C. Extracorporeal shock wave promotes activation of anterior cruciate ligament remnant cells and their paracrine regulation of bone marrow stromal cells’ proliferation, migration, collagen synthesis, and differentiation. Bone Jt. Res. 2020, 9, 458–468. [Google Scholar] [CrossRef]
- Beynnon, B.D.; Fleming, B.C.; Labovitch, R.; Parsons, B. Chronic anterior cruciate ligament deficiency is associated with increased anterior translation of the tibia during the transition from nonweightbearing to weightbearing. J. Orthop. Res. 2002, 20, 332–337. [Google Scholar] [CrossRef]
- Musahl, V.; Karlsson, J. Anterior cruciate ligament tear. N. Engl. J. Med. 2019, 380, 2341–2384. [Google Scholar] [CrossRef]
- Hirokawa, S.; Tsuruno, R. Three-dimensional deformation and stress distribution in an analytical/computational model of the anterior cruciate ligament. J. Biomech. 2000, 33, 1069–1077. [Google Scholar] [CrossRef]
- Xie, F.; Yang, L.; Guo, L.; Wang, Z.J.; Dai, G. A study on construction three-dimensional nonlinear finite element model and stress distribution analysis of anterior cruciate ligament. J. Biomech. Eng. 2009, 131, 120117. [Google Scholar] [CrossRef] [PubMed]
- Liederbach, M.; Dilgen, F.E.; Rose, D.J. Incidence of anterior cruciate ligament injuries among elite ballet and modern dancer: A 5-year prospective study. Am. J. Sports Med. 2008, 36, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Liderbach, M.; Kremenic, I.J.; Orishimo, K.F.; Pappas, E.; Hagins, M. Comparison of landing biomechanics between male and female dancers and athletes, part 2: Influence of fatigue and implications for anterior cruciate ligament injury. Am. J. Sports Med. 2014, 42, 1089–1095. [Google Scholar] [CrossRef]
- Hill, M.; Goldspink, G. Expression and splicing of the insulin-like growth factor gene in rodent muscle is associated with muscle satellite (stem) cell activation following local tissue damage. J. Physiol. 2003, 549 Pt 2, 409–418. [Google Scholar] [CrossRef]
- Matheny, R.W., Jr.; Nindl, B.C.; Adamo, M.L. Minireview: Mechano-growth factor: A putative product of IGF-1 gene expression involved in tissue repair and regeneration. Endocrinology 2010, 151, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Wang, Y.L.; Qiu, J.H.; Zhang, B.B.; Sun, J.; Lv, Y.G.; Yang, L. A novel mechanical loading model for studying the distributions of strain and mechano-growth factor expression. Arch. Biochem. Biophys. 2011, 511, 8–13. [Google Scholar] [CrossRef]
- Carpenter, V.; Matthews, K.; Devlin, G.; Stuart, S.; Jensen, J.; Conaglen, J.; Jeanplong, F.; Goldspink, P.; Yang, S.Y.; Goldspink, G.; et al. Mechano-growth factor reduces loss of cardiac function in acute myocardial infraction. Heart Lung Circ. 2008, 17, 33–39. [Google Scholar] [CrossRef]
- Beresewicz, M.; Majewska, M.; Makarewicz, D.; Vayro, S.; Zabłocka, B.; Górecki, D.C. Changes in the expression of insulin-like growth factor 1 variants in the postnatal brain development and in neonatal hypoxia-ischaemia. Int. J. Dev. Neurosci. 2010, 28, 91–97. [Google Scholar] [CrossRef]
- Sha, Y.Q.; Yang, L.; Lv, Y.G. MGF E peptide improves anterior cruciate ligament repair by inhibiting hypoxia-induced cell apoptosis and accelerating angiogenesis. J. Cell. Physiol. 2019, 234, 8846–8861. [Google Scholar] [CrossRef]
- Sha, Y.Q.; Lv, Y.G.; Xu, Z.L.; Yang, L.; Hao, X.Y.; Afandi, R. MGF E peptide pretreatment improves the proliferation and osteogenic differentiation of BMSCs via MEK-ERK1/2 and PI3K-Akt pathway under severe hypoxia. Life Sci. 2017, 189, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.Q.; Cai, W.J.; Khalid, A.M.; Chi, Q.J.; Wang, J.; Sun, T.; Wang, C.L. pretreatment with mechano growth factor E peptide attenuates osteoarthritis through improving cell proliferation and extracellular matrix synthesis in chondrocytes under severe hypoxia. Int. Immunopharmacol. 2021, 97, 107628. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wu, F.; Yeung, E.W.; Li, Y. IGF-IEc expression, regulation and biological function in different tissues. Growth Horm. IGF Res. 2010, 20, 275–281. [Google Scholar] [CrossRef]
- Yang, S.; Alnageeb, M.; Simpson, H.; Goldspink, G. Cloning and characterization of an IGF-1 isoform expressed in skeletal muscle subjected to stretch. J. Muscle Res. Cell Motil. 1996, 17, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Hao, X.; Sha, Y.; Yang, L. Pretreatment with mechano-growth factor E peptide protects bone marrow mesenchymal cells against damage by fluid shear stress. Biotechnol. Lett. 2014, 36, 2559–2569. [Google Scholar] [CrossRef] [PubMed]
- Causeret, F.; Hidalgo-Sanchez, M.; Fort, P.; Backer, S.; Popoff, M.R.; Gauthier-Rouvière, C.; Bloch-Gallego, E. Distincet roles of Rac1/Cdc42 and Rho/RocK for axon outgrowth and nucleokinesis of precerebellar neurons toward netrin 1. Development 2004, 131, 2481–2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saso, J.; Shields, S.K.; Zuo, Y.F.; Chakraborty, C. Role of Rho GTPases in human tropblast migration induced by IGFBP1. Biol. Reprod. 2012, 86, 1–9. [Google Scholar] [CrossRef]
- Yang, L.; Tsai, C.M.; Hsieh, A.H.; Lin, V.S.; Akeson, W.H.; Sung, K.L. Adhesion strength of differential of human ligament fibroblasts to collagen types I and III. J. Orthop. Res. 1999, 17, 755–762. [Google Scholar] [CrossRef]
- Feng, C.H.; Cheng, Y.C.; Chao, P.H.G. The influence and interactions of substrate thickness, organization and dimensionality on cell morphology and migration. Acta Biomater. 2013, 9, 5502–5510. [Google Scholar] [CrossRef]
- Tsai, C.H.; Lin, B.J.; Chao, P.H.G. α2β1 integrin and RhoA mediates electric field-induced ligament fibroblasts migration directionality. J. Orthop. Res. 2013, 31, 322–327. [Google Scholar] [CrossRef]
- Yu, X.H.; Mengsteab, P.Y.; Narayanan, G.; Nair, L.S.; Laurencin, C.T. Enhancing the surface properties of a bioengineered anterior cruciate ligament matrix for use with point-of-care stem cell therapy. Engineering 2021, 7, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Marmura, H.; Tremblay, P.F.; Getgood, A.M.J.; Bryant, D.M. The knee injury and osteoarthritis outcome score does not have adequate structural validity for use with young, active patients with ACL tears. Clin. Orthop. Relat. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, C.; Mouton, C.; Magosch, A.; Komnos, G.A.; Menetrey, J.; Ollivier, M.; Seil, R. The aspiration test reveals an instability of the posterior horn of the lateral meniscus in almost one-third of ACL-injured patients. Knee Surg. Sports Traumatol. Arthrosc. 2021. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Powers, M.J.F.; Narez, G.E.; Dejardin, L.M.; Donahue, T.H.; Haut, R.C. Post-traumatic osteoarthritis in rabbits following traumatic injury and surgical reconstruction of the knee. Ann. Biomed. Eng. 2022, 50, 169–182. [Google Scholar]
- Yan, Y.; Singh, G.K.; Zhang, F.F.; Wang, P.; Liu, W.Q.; Zhong, L.; Yang, L. Comparative study of normal and rheumatoid arthritis fibroblast-like synoviocytes proliferation under cyclic mechanical stretch: Role of prostaglandin E2. Connect. Tissue Res. 2012, 53, 246–254. [Google Scholar] [CrossRef]
- Du, V.; Luciani, N.; Richard, S.; Mary, G.; Gay, C.; Mazuel, F.; Reffay, M.; Menasché, P.; Agbulut, O.; Wilhelm, C. A 3D magnetic tissue stretcher for remote mechanical control of embryonic stem cell differentiation. Nat. Commun. 2017, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Lee, H.S.; Villarreal, F.; Teng, A.; Lu, E.; Reynolds, S.; Qin, C.; Smith, J.; Sung, K.L.P. Differential MMP-2 activity of ligament cells under mechanical stretch injury: An in vitro study on human ACL and MCL fibroblasts. J. Orthop. Res. 2005, 23, 949–957. [Google Scholar] [CrossRef]
- Xu, C.M.; Zhang, Y.J.; Sutrisno, L.; Yang, L.; Chen, R.F.; Sung, K.L.P. Bay11-7082 facilitates wound healing by antagonizing mechanical injury- and TNF-α-induced expression of MMPs in posterior cruciate ligament. Connect. Tissue Res. 2019, 60, 311–322. [Google Scholar] [CrossRef]
- Sha, Y.Q.; Afandi, R.; Zhang, B.B.; Yang, L.; Lv, Y.G. MGF E peptide pretreatment improves collagen synthesis and cell proliferation of injured human ACL fibroblasts via MEK-ERK1/2 signaling pathway. Growth Factors 2017, 35, 29–38. [Google Scholar] [CrossRef]
- Murray, M.M.; Martin, S.D.; Martin, T.L.; Spector, M. Histological changes in the human anterior cruciate ligament after rupture. J. Bone Jt. Surg. Am. 2000, 82, 1387–1397. [Google Scholar] [CrossRef]
- Castro, P.A.T.S.; Machanocker, D.H.; Luna, G.F.; Barbosa, G.M.; Cunha, J.E.; Cunha, T.M.; Cunha, F.Q.; Russo, T.L.; Salvini, T.F. Clinical-like cryotherapy in acute knee arthritis protects neuromuscular junctions of quadriceps and reduces joint inflammation in mice. Biomed. Res. Int. 2022, 2022, 7442289. [Google Scholar] [CrossRef] [PubMed]
- Kim-Wang, S.Y.; Holt, A.G.; McGowan, A.M.; Danyluk, S.T.; Goode, A.P.; Lau, B.C.; Toth, A.P.; Wittstein, J.R.; DeFrate, L.E.; Yi, J.S.; et al. Immune cell profiles in synovial fluid after anterior cruciate ligament and meniscus injuries. Arthritis Res. Ther. 2021, 23, 280. [Google Scholar] [CrossRef] [PubMed]
- Bellririchard, M.; Snider, C.; Kuroki, K.; Brockman, J.; Grant, D.A. The use of gold nanoparticles in improving ACL graft performance in an ovine model. J. Biomater. Appl. 2022, 36, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Kartus, J.; Movin, T.; Karlsson, J. Donor-site morbidity and anterior knee problems after anterior cruciate ligament reconstruction using autografts. Arthroscopy 2001, 17, 971–980. [Google Scholar] [CrossRef]
- Ge, Z.; Yang, F.; Goh, J.C.H.; Ramakrishna, S.; Lee, E.H. Biomaterials and scaffolds for ligament tissue engineering. J. Biomed. Mater. Res. A 2006, 77, 639–652. [Google Scholar] [CrossRef]
- Leong, N.L.; Petrigliano, F.A.; McAllister, D.R. Current tissue engineering strategies in anterior cruciate ligament reconstruction. J. Biomed. Mater. Res. A 2014, 102, 1614–1624. [Google Scholar] [CrossRef]
- Kondo, E.; Yasuda, K.; Onodera, J.; Kawaguchi, Y.; Kitamura, N. Effects of remnant tissue preservation on clinical and arthroscopic results after anatomic double-bundle anterior cruciate ligament reconstruction. Am. J. Sports Med. 2015, 43, 1882–1892. [Google Scholar] [CrossRef]
- Murray, M.M.; Spector, M. The migration of cells from the ruptured human anterior cruciate ligament into collagen-glycosaminoglycan regeneration templates in vitro. Biomaterials 2001, 22, 2393–2402. [Google Scholar] [CrossRef]
- Cai, L.; Brophy, R.H.; Tycksen, E.D.; Duan, X.; Nunley, R.M.; Rai, M.F. Distinct expression pattern of periostin splice variants in chondrocytes and ligament progenitor cells. FASEB J. 2019, 33, 8386–8405. [Google Scholar] [CrossRef]
- Asai, K.; Nakase, J.; Yoshimizu, R.; Kimura, M.; Tsuchiya, H. Does remnant tissue preservation in anterior cruciate ligament reconstruction influence the creation of the rectangular femoral tunnel? J. Orthop. Surg. 2021, 29, 23094990211061249. [Google Scholar] [CrossRef]
- Gögele, C.; Konrad, J.; Hahn, J.; Breier, A.; Schröpfer, M.; Meyer, M.; Merkel, R.; Hoffmann, B.; Schulze-Tanzil, G. Maintenance of ligament homeostasis of spheroid-colonized embroidered and functionalized scaffolds after 3D stretch. Int. J. Mol. Sci. 2021, 22, 8204. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.Y.; Liao, J.F.; Yang, W.B.; Zhou, X.D.; Sung, K.L.P.; Xie, J. Influences of tumor necrosis factor-α on lysyl oxidases and matrix metalloproteinases of injured anterior cruciate ligament and medial collateral ligament fibroblasts. J. Knee Surg. 2017, 30, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Chi, Q.J.; Sha, Y.Q.; Xu, K.; Xu, C.M.; Chen, C.; Huang, W.; Chen, P.X.; Chen, P.; Yang, L.; et al. Mechanical inury and IL-1β regulated LOXs and MMP-1, 2, 3 expression in ACL fibroblasts co-cultured with synoviocytes. Biotechnol. Lett. 2020, 42, 1567–1579. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Mei, J.; Qiang, L.; Fang, W.; Yijun, N. LncRNA uc.48+ regulates immune and inflammatory reactions mediated by the P2X7 receptor in type 2 diabetic mice. Exp. Ther. Med. 2020, 20, 230. [Google Scholar]
- Murray, M.M.; Spindler, K.P.; Ballard, P.; Welch, T.P.; Zurakowski, D.; Nanney, L.B. Enhanced histologic repair in a central wound in the anterior cruciate ligament with a collagen-platelet-rich plasma scaffold. J. Orthop. Res. 2007, 25, 1007–1017. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Amplicon Length | Annealing Temperature |
|---|---|---|---|
| Cdc42 | F: 5′-CCATCGGAATATGTACCGACTG-3′ | 128 bp | 60 °C |
| R: 5′-CTCAGCGGTCGTAATCTGTCA-3′ | |||
| Rac1 | F: 5′-ATGTCCGTGCAAAGTGGTATC-3′ | 249 bp | 60 °C |
| R: 5′-CTCGGATCGCTTCGTCAAACA-3′ | |||
| RhoA | F: 5′- GATTGGCGCTTTTGGGTACAT-3′ | 85 bp | 60 °C |
| R: 5′-AGCAGCTCTCGTAGCCATTTC-3′ | |||
| ROCK1 | F: 5′-AAGTGAGGTTAGGGCGAAATG-3′ | 219 bp | 60 °C |
| R: 5′-AAGGTAGTTGATTGCCAACGAA-3′ | |||
| PAK1 | F: 5′-CAACTCGGGACGTGGCTAC-3′ | 81 bp | 60 °C |
| R: 5′-CAGTATTCCGGGTCAAAGCAT-3′ | |||
| PAK2 | F: 5′-TGAGCACACCATCCATGTTGG-3′ | 86 bp | 60 °C |
| R: 5′-AGGTCTGTAGTAATCGAGCCC-3′ | |||
| GAPDH | F: 5′-GGATTTGGTCGTATTGGG-3′ | 218 bp | 60 °C |
| R: 5′-GCTCCTGGAAGATGGTGAT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sha, Y.; Zhang, B.; Chen, L.; Hong, H.; Chi, Q. Mechano Growth Factor Accelerates ACL Repair and Improves Cell Mobility of Mechanically Injured Human ACL Fibroblasts by Targeting Rac1-PAK1/2 and RhoA-ROCK1 Pathways. Int. J. Mol. Sci. 2022, 23, 4331. https://doi.org/10.3390/ijms23084331
Sha Y, Zhang B, Chen L, Hong H, Chi Q. Mechano Growth Factor Accelerates ACL Repair and Improves Cell Mobility of Mechanically Injured Human ACL Fibroblasts by Targeting Rac1-PAK1/2 and RhoA-ROCK1 Pathways. International Journal of Molecular Sciences. 2022; 23(8):4331. https://doi.org/10.3390/ijms23084331
Chicago/Turabian StyleSha, Yongqiang, Beibei Zhang, Liping Chen, Huhai Hong, and Qingjia Chi. 2022. "Mechano Growth Factor Accelerates ACL Repair and Improves Cell Mobility of Mechanically Injured Human ACL Fibroblasts by Targeting Rac1-PAK1/2 and RhoA-ROCK1 Pathways" International Journal of Molecular Sciences 23, no. 8: 4331. https://doi.org/10.3390/ijms23084331
APA StyleSha, Y., Zhang, B., Chen, L., Hong, H., & Chi, Q. (2022). Mechano Growth Factor Accelerates ACL Repair and Improves Cell Mobility of Mechanically Injured Human ACL Fibroblasts by Targeting Rac1-PAK1/2 and RhoA-ROCK1 Pathways. International Journal of Molecular Sciences, 23(8), 4331. https://doi.org/10.3390/ijms23084331