
Genetics Matters: Voyaging from the Past into the Future of Humanity and Sustainability

 , , , and
, , , and
Abstract
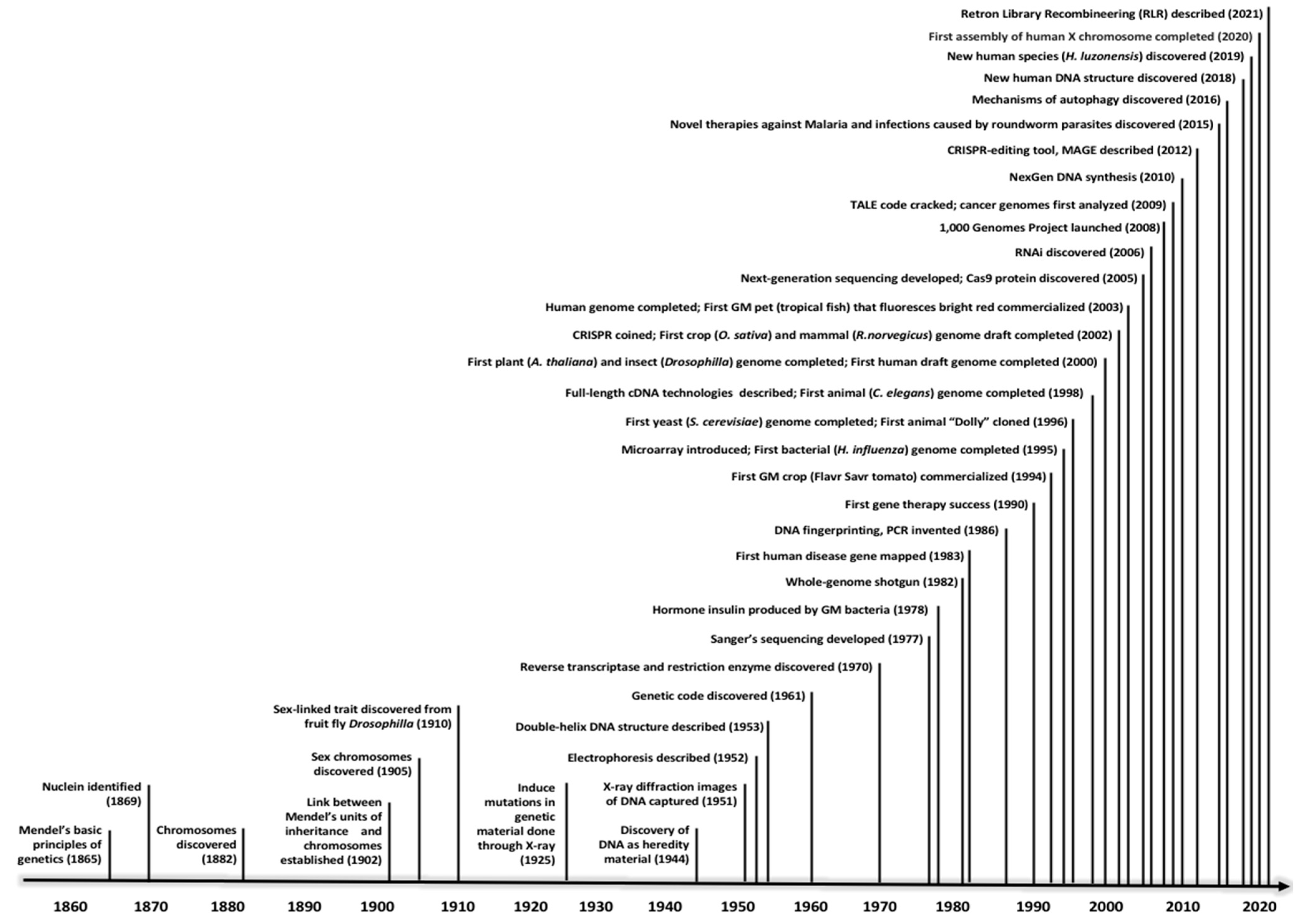
1. A Trip down Memory Lane: One and a Half Centuries into the Intriguing Study of Heredity
2. How Has the Cracking of Genetic Code Improved Life on Earth?
2.1. Medicine
2.2. Agriculture
2.3. Biodiversity
3. Genetic Revolution in the 21st Century: The Polemic of Gene Editing
4. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Griffiths, A.J.F.; Doebley, J.; Peichel, C.; Wassarman, D.A. Introduction to Genetic Analysis 2-36; W.H. Freeman and Company: New York, NY, USA, 2020. [Google Scholar]
- Fairbanks, D.J. Mendel and Darwin: Untangling a persistent enigma. Heredity 2019, 124, 263–273. [Google Scholar] [CrossRef]
- Mendel, G. Versuche uber Plflanzen-Hybriden. In Verhandlungen des Naturforschenden Ver-Eines in Brünn, Bd. IV Für das Jahr 1865 Abhand-Lungen; Electronic Scholarly Publishing Project; Verlage des Vereines: Brünn, Czechoslovakia, 1866; pp. 3–47. [Google Scholar]
- Howard, J.C. Why didn’t Darwin discover Mendel’s laws? J. Biol. 2009, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Paweletz, N. Walther Flemming: Pioneer of mitosis research. Nat. Rev. Mol. Cell Biol. 2001, 2, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. Sex Limited Inheritance in Drosophila. Science 1910, 32, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Miga, K.H.; Koren, S.; Rhie, A.; Vollger, M.R.; Gershman, A.; Bzikadze, A.; Brooks, S.; Howe, E.; Porubsky, D.; Logsdon, G.A.; et al. Telomere-to-telomere assembly of a complete human X chromosome. Nature 2020, 585, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.; Sankaran, N. DNA translated: Friedrich Miescher’s discovery of nuclein in its original context. Br. J. Hist. Sci. 2021, 54, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Hoppe-Seyler, F. Ueber die chemische Zusammensetzung des Eiters. Med.-Chem. Unters. 1871, 4, 486–501. [Google Scholar]
- Avery, O.T.; MacLeod, C.M.; Mccarty, M. Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type III. J. Exp. Med. 1944, 79, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.J. Who cares about the double helix? Nature 2003, 422, 803–804. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, M.W.; Jones, O.W.; Leder, P.; Clark, B.F.C.; Sly, W.S.; Pestka, S. On the Coding of Genetic Information. Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1963; pp. 549–557. [Google Scholar]
- Coffin, J.M. 50th anniversary of the discovery of reverse transcriptase. Mol. Biol. Cell 2021, 32, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Maxam, A.M.; Gilbert, W. A new method for sequencing DNA. Proc. Natl. Acad. Sci. USA 1977, 74, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, L.; Oliveira, D.; Høgdall, C.; Høgdall, E. Next Generation Sequencing Technology in the Clinic and Its Challenges. Cancers 2021, 13, 1751. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Heather, J.M.; Chain, B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mullis, K.; Faloona, F.; Scharf, S.; Saiki, R.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb. Symp. Quant. Biol. 1986, 51, 263–273. [Google Scholar] [CrossRef]
- Udugama, B.; Kadhiresan, P.; Kozlowski, H.N.; Malekjahani, A.; Osborne, M.; Li, V.Y.C.; Chen, H.; Mubareka, S.; Gubbay, J.B.; Chan, W.C.W. Diagnosing COVID-19: The Disease and Tools for Detection. ACS Nano 2020, 14, 3822–3835. [Google Scholar] [CrossRef]
- Slatko, B.E.; Gardner, A.F.; Ausubel, F.M. Overview of Next-Generation Sequencing Technologies. Curr. Protoc. Mol. Biol. 2018, 122, e59. [Google Scholar] [CrossRef]
- Wheeler, D.A.; Srinivasan, M.; Egholm, M.; Shen, Y.; Chen, L.; McGuire, A.; He, W.; Chen, Y.-J.; Makhijani, V.; Roth, G.T.; et al. The complete genome of an individual by massively parallel DNA sequencing. Nature 2008, 452, 872–876. [Google Scholar] [CrossRef]
- Bentley, D.R.; Balasubramanian, S.; Swerdlow, H.P.; Smith, G.P.; Milton, J.; Brown, C.G.; Hall, K.P.; Evers, D.J.; Barnes, C.L.; Bignell, H.R.; et al. Accurate whole human genome sequencing using reversible terminator chemistry. Nature 2008, 456, 53–59. [Google Scholar] [CrossRef]
- Glanzmann, B.; Jooste, T.; Ghoor, S.; Gordon, R.; Mia, R.; Mao, J.; Li, H.; Charls, P.; Douman, C.; Kotze, M.J.; et al. Human whole genome sequencing in South Africa. Sci. Rep. 2021, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR-Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef]
- Ishino, Y.; Krupovic, M.; Forterre, P. History of CRISPR-Cas from Encounter with a Mysterious Repeated Sequence to Genome Editing Technology. J. Bacteriol. 2018, 200, e00580-17. [Google Scholar] [CrossRef] [PubMed]
- McTague, A.; Rossignoli, G.; Ferrini, A.; Barral, S.; Kurian, M.A. Genome Editing in iPSC-Based Neural Systems: From Disease Models to Future Therapeutic Strategies. Front. Genome Ed. 2021, 3, 630600. [Google Scholar] [CrossRef] [PubMed]
- Yanofsky, C. Establishing the Triplet Nature of the Genetic Code. Cell 2007, 128, 815–818. [Google Scholar] [CrossRef]
- Kanzi, A.M.; San, J.E.; Chimukangara, B.; Wilkinson, E.; Fish, M.; Ramsuran, V.; De Oliveira, T. Next Generation Sequencing and Bioinformatics Analysis of Family Genetic Inheritance. Front. Genet. 2020, 11, 544162. [Google Scholar] [CrossRef] [PubMed]
- Dicenzo, G.C.; Finan, T.M. The Divided Bacterial Genome: Structure, Function, and Evolution. Microbiol. Mol. Biol. Rev. 2017, 81, e00019-17. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Petrone, M.E.; Holmes, E.C. We shouldn’t worry when a virus mutates during disease outbreaks. Nat. Microbiol. 2020, 5, 529–530. [Google Scholar] [CrossRef]
- Ganna, A. COVID-19 Host Genetics Initiative. Mapping the human genetic architecture of COVID-19. Nature 2021, 600, 472–477. [Google Scholar] [CrossRef]
- Rasmussen, S.A.; Abul-Husn, N.S.; Casanova, J.-L.; Daly, M.J.; Rehm, H.L.; Murray, M.F. The intersection of genetics and COVID-19 in 2021: Preview of the 2021 Rodney Howell Symposium. Genet. Med. 2021, 23, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Lewin, H.A.; Robinson, G.E.; Kress, W.J.; Baker, W.J.; Coddington, J.; Crandall, K.A.; Durbin, R.; Edwards, S.V.; Forest, F.; Gilbert, M.T.P.; et al. Earth BioGenome Project: Sequencing life for the future of life. Proc. Natl. Acad. Sci. USA 2018, 115, 4325–4333. [Google Scholar] [CrossRef]
- Exposito-Alonso, M.; Drost, H.; Burbano, H.A.; Weigel, D. The Earth BioGenome project: Opportunities and challenges for plant genomics and conservation. Plant J. 2019, 102, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome Sequence of Aedes aegypti, a Major Arbovirus Vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef]
- Elsik, C.G.; Tellam, R.L.; Worley, K.C. The Genome Sequence of Taurine Cattle: A Window to Ruminant Biology and Evolution. Science 2009, 324, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, C.T.; Alföldi, J.; Lee, A.P.; Fan, S.; Philippe, H.; MacCallum, I.; Braasch, I.; Manousaki, T.; Schneider, I.; Rohner, N.; et al. The African coelacanth genome provides insights into tetrapod evolution. Nature 2013, 496, 311–316. [Google Scholar] [CrossRef]
- The Chimpanzee Sequencing and Analysis Consortium. Initial sequence of the chimpanzee genome and comparison with the human genome. Nature 2005, 437, 69–87. [Google Scholar] [CrossRef] [PubMed]
- The Marmoset Genome Sequencing and Analysis Consortium. The common marmoset genome provides insight into primate biology and evolution. Nat. Genet. 2014, 46, 850–857. [Google Scholar] [CrossRef]
- Li, R.; Fan, W.; Tian, G.; Zhu, H.; He, L.; Cai, J.; Huang, Q.; Cai, Q.; Li, B.; Bai, Y.; et al. The sequence and de novo assembly of the giant panda genome. Nature 2009, 463, 311–317. [Google Scholar] [CrossRef]
- Weinstock, G.M.; Robinson, G.E.; Gibbs, R.A.; Weinstock, G.M.; Weinstock, G.M.; Robinson, G.E.; Worley, K.C.; Evans, J.D.; Maleszka, R.; Robertson, H.M.; et al. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar] [CrossRef]
- Kasahara, M.; Naruse, K.; Sasaki, S.; Nakatani, Y.; Qu, W.; Ahsan, B.; Yamada, T.; Nagayasu, Y.; Doi, K.; Kasai, Y.; et al. The medaka draft genome and insights into vertebrate genome evolution. Nature 2007, 447, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.C.; Hillier, L.W.; Graves, J.A.M.; Birney, E.; Ponting, C.P.; Grützner, F.; Belov, K.; Miller, W.; Clarke, L.; Chinwalla, A.T.; et al. Genome analysis of the platypus reveals unique signatures of evolution. Nature 2008, 453, 175–183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yue Tribolium Genome Sequencing Consortium; Richards, S.; Gibbs, R.A.; Weinstock, G.M.; Brown, S.J.; Denell, R. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar] [CrossRef]
- Sodergren, E.; Weinstock, G.M.; Davidson, E.H.; Cameron, R.A.; Gibbs, R.A.; Angerer, R.C.; Angerer, L.M.; Arnone, M.I.; Burgess, D.R.; Burke, R.D.; et al. The Genome of the Sea Urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.-E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Scally, A.; Dutheil, J.; Hillier, L.W.; Jordan, G.E.; Goodhead, I.; Herrero, J.; Hobolth, A.; Lappalainen, T.; Mailund, T.; Marques-Bonet, T.; et al. Insights into hominid evolution from the gorilla genome sequence. Nature 2012, 483, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Nowoshilow, S.; Schloissnig, S.; Fei, J.-F.; Dahl, A.; Pang, A.W.C.; Pippel, M.; Winkler, S.; Hastie, A.R.; Young, G.; Roscito, J.G.; et al. The axolotl genome and the evolution of key tissue formation regulators. Nature 2018, 554, 50–55. [Google Scholar] [CrossRef]
- Burga, A.; Wang, W.; Ben-David, E.; Wolf, P.C.; Ramey, A.M.; Verdugo, C.; Lyons, K.; Parker, P.G.; Kruglyak, L. A genetic signature of the evolution of loss of flight in the Galapagos cormorant. Science 2017, 356, eaal3345. [Google Scholar] [CrossRef]
- Babb, P.L.; Lahens, N.F.; Correa-Garhwal, S.; Nicholson, D.; Kim, E.J.; HogenEsch, J.B.; Kuntner, M.; Higgins, L.; Hayashi, C.Y.; Agnarsson, I.; et al. The Nephila clavipes genome highlights the diversity of spider silk genes and their complex expression. Nat. Genet. 2017, 49, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ong-Abdullah, M.; Low, E.-T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.-L.; Ooi, S.-E.; Chan, K.-L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in old and new worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Project, A.G.; Albert, V.A.; Barbazuk, W.B.; Depamphilis, C.W.; Der, J.P.; Leebens-Mack, J.; Ma, H.; Palmer, J.D.; Rounsley, S.; Sankoff, D.; et al. The Amborella Genome and the Evolution of Flowering Plants. Science 2013, 342, 1241089. [Google Scholar] [CrossRef]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.D.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.X.; Gouzy, J.; Schoof, H.; et al. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Raymond, O.; Gouzy, J.; Just, J.; Badouin, H.; Verdenaud, M.; Lemainque, A.; Vergne, P.; Moja, S.; Choisne, N.; Pont, C.; et al. The Rosa genome provides new insights into the domestication of modern roses. Nat. Genet. 2018, 50, 772–777. [Google Scholar] [CrossRef] [PubMed]
- D’Hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.-C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 Maize Genome: Complexity, Diversity, and Dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Hou, S.; Feng, Y.; Yu, Q.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M.; et al. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83–89. [Google Scholar] [CrossRef]
- Diambra, L.A.; The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Myburg, A.A.; Grattapaglia, D.; Tuskan, G.A.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Salojärvi, J.; Smolander, O.-P.; Nieminen, K.; Rajaraman, S.; Safronov, O.; Safdari, P.; Lamminmäki, A.; Immanen, J.; Lan, T.; Tanskanen, J.; et al. Genome sequencing and population genomic analyses provide insights into the adaptive landscape of silver birch. Nat. Genet. 2017, 49, 904–912. [Google Scholar] [CrossRef]
- Teh, B.T.; Lim, K.; Yong, C.H.; Ng, C.C.Y.; Rao, S.R.; Rajasegaran, V.; Lim, W.K.; Ong, C.K.; Chan, K.; Cheng, V.K.Y.; et al. The draft genome of tropical fruit durian (Durio zibethinus). Nat. Genet. 2017, 49, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Badouin, H.; Gouzy, J.; Grassa, C.J.; Murat, F.; Staton, S.E.; Cottret, L.; Lelandais-Briere, C.; Owens, G.L.; Carrere, S.; Mayjonade, B.; et al. The sunflower genome provides insights into oil metabolism, flowering and Asterid evolution. Nature 2017, 546, 148–152. [Google Scholar] [CrossRef]
- Luo, M.-C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef]
- Varshney, R.K.; Shi, C.; Thudi, M.; Mariac, C.; Wallace, J.; Qi, P.; Zhang, H.; Zhao, Y.; Wang, X.; Rathore, A.; et al. Pearl millet genome sequence provides a resource to improve agronomic traits in arid environments. Nat. Biotechnol. 2017, 35, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Pel, H.J.; de Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E.; Calvo, S.E.; Cuomo, C.; Ma, L.-J.; Wortman, J.R.; Batzoglou, S.; Lee, S.-I.; Baştürkmen, M.; Spevak, C.C.; Clutterbuck, J.; et al. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005, 438, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Wood, V.; Gwilliam, R.; Rajandream, M.-A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; et al. The genome sequence of Schizosaccharomyces pombe. Nature 2002, 415, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef]
- Ohm, R.A.; De Jong, J.F.; Lugones, L.G.; Aerts, A.; Kothe, E.; Stajich, J.E.; de Vries, R.P.; Record, E.; Levasseur, A.; Baker, S.E.; et al. Genome sequence of the model mushroom Schizophyllum commune. Nat. Biotechnol. 2010, 28, 957–963. [Google Scholar] [CrossRef]
- Butler, G.; Rasmussen, M.D.; Lin, M.F.; Santos, M.A.S.; Sakthikumar, S.; Munro, C.A.; Rheinbay, E.; Grabherr, M.; Forche, A.; Reedy, J.L.; et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature 2009, 459, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, M.A.; Albang, R.; Albermann, K.; Badger, J.H.; Daran, J.-M.; Driessen, A.J.M.; Garcia-Estrada, C.; Fedorova, N.D.; Harris, D.M.; Heijne, W.H.M.; et al. Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat. Biotechnol. 2008, 26, 1161–1168. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) [Internet]. U.S. National Library of Medicine, National Center for Biotechnology Information: Bethesda, MD, USA. 1988. Available online: http://www.ncbi.nlm.nih.gov (accessed on 23 November 2021).
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Brown, T.A. Genomes, 2nd ed.; Chapter 14, Mutation, Repair and Recombination; Wiley-Liss: Oxford, UK, 2002. [Google Scholar]
- Loewe, L.; Hill, W.G. The population genetics of mutations: Good, bad and indifferent. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1153–1167. [Google Scholar] [CrossRef]
- Dong, A.C.; Rivella, S. Gene Addition Strategies for beta-Thalassemia and Sickle Cell Anemia. Adv. Exp. Med. Biol. 2017, 1013, 155–176. [Google Scholar] [CrossRef]
- Zook, J.M.; Hansen, N.F.; Olson, N.D.; Chapman, L.; Mullikin, J.C.; Xiao, C.; Sherry, S.; Koren, S.; Phillippy, A.M.; Boutros, P.C.; et al. A robust benchmark for detection of germline large deletions and insertions. Nat. Biotechnol. 2020, 38, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.N.; Chang, A.; Boyer, H.W.; Helling, R.B. Potential biohazards of recombinant DNA molecules. Proc. Natl. Acad. Sci. USA 1974, 71, 2593–2594. [Google Scholar] [CrossRef]
- Seymour, M. Rapid progression and future of environmental DNA research. Commun. Biol. 2019, 2, 80. [Google Scholar] [CrossRef]
- Turnbull, C.; Lillemo, M.; Hvoslef-Eide, T.A.K. Global Regulation of Genetically Modified Crops Amid the Gene Edited Crop Boom—A Review. Front. Plant Sci. 2021, 12, 630396. [Google Scholar] [CrossRef]
- Chapman, P.B.; Hauschild, A.; Robert, C.; Haanen, J.B.; Ascierto, P.; Larkin, J.; Dummer, R.; Garbe, C.; Testori, A.; Maio, M.; et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 2011, 364, 2507–2516. [Google Scholar] [CrossRef]
- Kwak, E.L.; Bang, Y.-J.; Camidge, D.R.; Shaw, A.T.; Solomon, B.; Maki, R.G.; Ou, S.-H.I.; Dezube, B.J.; Jänne, P.A.; Costa, D.; et al. Anaplastic Lymphoma Kinase Inhibition in Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2010, 363, 1693–1703. [Google Scholar] [CrossRef]
- Druker, B.J.; Guilhot, F.; O’Brien, S.G.; Gathmann, I.; Kantarjian, H.M.; Gattermann, N.; Deininger, M.W.; Silver, R.T.; Goldman, J.M.; Stone, R.M.; et al. Five-Year Follow-up of Patients Receiving Imatinib for Chronic Myeloid Leukemia. N. Engl. J. Med. 2006, 355, 2408–2417. [Google Scholar] [CrossRef]
- Chamayou, S.; Sicali, M.; Lombardo, D.; Alecci, C.; Ragolia, C.; Maglia, E.; Liprino, A.; Cardea, C.; Storaci, G.; Romano, S.; et al. Universal strategy for preimplantation genetic testing for cystic fibrosis based on next generation sequencing. J. Assist. Reprod. Genet. 2019, 37, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Simón, C. Embryo Genetics. Genes 2021, 12, 118. [Google Scholar] [CrossRef] [PubMed]
- Aiyegbusi, O.L.; Macpherson, K.; Elston, L.; Myles, S.; Washington, J.; Sungum, N.; Briggs, M.; Newsome, P.N.; Calvert, M.J. Patient and public perspectives on cell and gene therapies: A systematic review. Nat. Commun. 2020, 11, 6265. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana, M.; Six, E.; Peyrou, C.L.; André, I.; Hacein-Bey-Abina, S. Gene Therapy for X-Linked Severe Combined Immunodeficiency: Where Do We Stand? Hum. Gene Ther. 2016, 27, 108–116. [Google Scholar] [CrossRef]
- Maguire, A.M.; Simonelli, F.; Pierce, E.A.; Pugh, E.N., Jr.; Mingozzi, F.; Bennicelli, J.; Banfi, S.; Marshall, K.A.; Francesco Testa, C.O.T.; Surace, E.M.; et al. Safety and efficacy of gene transfer for Leber’s congenital amaurosis. N. Engl. J. Med. 2008, 358, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- Hollon, T. Researchers and regulators reflect on first gene therapy death. Nat. Med. 2000, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Frank, K.M.; Hogarth, D.K.; Miller, J.L.; Mandal, S.; Mease, P.J.; Samulski, R.J.; Weisgerber, G.A.; Hart, J. Investigation of the Cause of Death in a Gene-Therapy Trial. N. Engl. J. Med. 2009, 361, 161–169. [Google Scholar] [CrossRef]
- Pepper, M.S.; Alessandrini, M.; Pope, A.; Van Staden, W.; Green, R.J. Cell and gene therapies at the forefront of innovative medical care: Implications for South Africa. S. Afr. Med. J. 2018, 109, 20–22. [Google Scholar] [CrossRef]
- Simon, J.; Arthur, C.; McGuire, V.A. Gene-Targeting Vectors. Adv. Struct. Saf. Stud. 2009, 561, 127–144. [Google Scholar] [CrossRef]
- von Ahsen, N.; Oellerich, M. Chip-Based Genotyping: Translation of Pharmacogenetic Research to Clinical Practice. Clin. Chem. 2007, 53, 1186–1187. [Google Scholar] [CrossRef]
- Drew, Y.; Ledermann, J.; Hall, G.; Rea, D.; Glasspool, R.; Highley, M.; Jayson, G.; Sludden, J.; Murray, J.; Jamieson, D.; et al. Phase 2 multicentre trial investigating intermittent and continuous dosing schedules of the poly(ADP-ribose) polymerase inhibitor rucaparib in germline BRCA mutation carriers with advanced ovarian and breast cancer. Br. J. Cancer 2016, 114, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Compagni, A.; Bartoli, S.; Buehrlen, B.; Fattore, G.; Ibarreta, D.; De Mesa, E.G. Avoiding adverse drug reactions by pharmacogenetic testing: A systematic review of the economic evidence in the case of TPMT and AZA-induced side effects. Int. J. Technol. Assess. Health Care 2008, 24, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Sabatello, M.; Eyal, G.; Lee, S.S.-J.; Rowe, J.W.; Stiles, D.F.; Swanson, A.; Appelbaum, P.S. Is precision medicine relevant in the age of COVID-19? Genet. Med. 2021, 23, 999–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-D.; Zhai, W.; Yang, H.-C.; Fan, R.-X.; Cao, X.; Zhong, L.; Wang, L.; Liu, F.; Wu, H.; Cheng, L.-G.; et al. The genomics of selection in dogs and the parallel evolution between dogs and humans. Nat. Commun. 2013, 4, 1860. [Google Scholar] [CrossRef]
- Lobato-Gómez, M.; Hewitt, S.; Capell, T.; Christou, P.; Dhingra, A.; Girón-Calva, P.S. Transgenic and genome-edited fruits: Background, constraints, benefits, and commercial opportunities. Hortic. Res. 2021, 8, 166. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef]
- Kranthi, K.R.; Stone, G.D. Long-term impacts of Bt cotton in India. Nat. Plants 2020, 6, 188–196. [Google Scholar] [CrossRef]
- Mollaee, M.; Mobli, A.; Chauhan, B.S. The response of glyphosate-resistant and glyphosate-susceptible biotypes of Echinochloa colona to carbon dioxide, soil moisture and glyphosate. Sci. Rep. 2020, 10, 329. [Google Scholar] [CrossRef]
- Van Der Straeten, D.; Bhullar, N.K.; De Steur, H.; Gruissem, W.; MacKenzie, D.; Pfeiffer, W.; Qaim, M.; Slamet-Loedin, I.; Strobbe, S.; Tohme, J.; et al. Multiplying the efficiency and impact of biofortification through metabolic engineering. Nat. Commun. 2020, 11, 5203. [Google Scholar] [CrossRef]
- Tang, G.; Qin, J.; Dolnikowski, G.G.; Russell, R.M.; Grusak, M.A. Golden Rice is an effective source of vitamin A. Am. J. Clin. Nutr. 2009, 89, 1776–1783. [Google Scholar] [CrossRef]
- Paine, J.A.; Shipton, C.A.; Chaggar, S.; Howells, R.; Kennedy, M.J.; Vernon, G.; Wright, S.Y.; Hinchliffe, E.; Adams, J.L.; Silverstone, A.L.; et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nat. Biotechnol. 2005, 23, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Jones, D. Genetic engineering of a mouse: Dr. Frank Ruddle and somatic cell genetics. Yale J. Biol. Med. 2011, 84, 117–124. [Google Scholar] [PubMed]
- Volobueva, A.; Orekhov, A.; Deykin, A. An update on the tools for creating transgenic animal models of human diseases—Focus on atherosclerosis. Braz. J. Med. Biol. Res. 2019, 52, e8108. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B. Pollution: Transgenic pigs to the rescue. eLife 2018, 7, e37641. [Google Scholar] [CrossRef]
- Gavin, W. ATryn(®): 1st GE (genetically engineered) animal success story for production of a human recombinant pharmaceutical. BMC Proc. 2014, 8, O4. [Google Scholar] [CrossRef]
- Eriksson, S.; Jonas, E.; Rydhmer, L.; Röcklinsberg, H. Invited review: Breeding and ethical perspectives on genetically modified and genome edited cattle. J. Dairy Sci. 2018, 101, 1–17. [Google Scholar] [CrossRef]
- Campbell, K.H.S.; McWhir, J.; Ritchie, W.A.; Wilmut, I. Sheep cloned by nuclear transfer from a cultured cell line. Nature 1996, 380, 64–66. [Google Scholar] [CrossRef]
- Giles, J.; Knight, J. Dolly’s death leaves researchers woolly on clone ageing issue. Nature 2003, 421, 776. [Google Scholar] [CrossRef][Green Version]
- Liu, Z.; Cai, Y.; Wang, Y.; Nie, Y.; Zhang, C.; Xu, Y.; Zhang, X.; Lu, Y.; Wang, Z.; Poo, M.; et al. Cloning of Macaque Monkeys by Somatic Cell Nuclear Transfer. Cell 2018, 172, 881–887.e7. [Google Scholar] [CrossRef]
- Qiu, P.; Jiang, J.; Liu, Z.; Cai, Y.; Huang, T.; Wang, Y.; Liu, Q.; Nie, Y.; Liu, F.; Cheng, J.; et al. BMAL1 knockout macaque monkeys display reduced sleep and psychiatric disorders. Natl. Sci. Rev. 2019, 6, 87–100. [Google Scholar] [CrossRef]
- Gurumurthy, C.B.; Quadros, R.M.; Richardson, G.P.; Poluektova, L.Y.; Mansour, S.L.; Ohtsuka, M. Genetically modified mouse models to help fight COVID-19. Nat. Protoc. 2020, 15, 3777–3787. [Google Scholar] [CrossRef]
- Brogard, J.M.; Blickle, J.F.; Paris-Bockel, D. Genetically engineered insulin: Five years of experience. Drugs Exp. Clin. Res. 1985, 11, 397–406. [Google Scholar] [PubMed]
- Chakrabarty, A.M. Bioengineered bugs, drugs and contentious issues in patenting. Bioeng. Bugs 2010, 1, 2–8. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lin, H.; Wang, Q.; Shen, Q.; Zhan, J.; Zhao, Y. Genetic engineering of microorganisms for biodiesel production. Bioengineered 2013, 4, 292–304. [Google Scholar] [CrossRef]
- Pandey, A.; Kamle, M.; Yadava, L.; Muthukumar, M.; Kumar, P.; Gupta, V.; Ashfaque, M.; Pandey, B. Genetically Modified Food: Its uses, Future Prospects and Safety Assessments. Biotechnology 2010, 9, 444–458. [Google Scholar] [CrossRef]
- Osanai, T.; Oikawa, A.; Numata, K.; Kuwahara, A.; Iijima, H.; Doi, Y.; Saito, K.; Hirai, M.Y. Pathway-Level Acceleration of Glycogen Catabolism by a Response Regulator in the Cyanobacterium Synechocystis Species PCC 6803. Plant Physiol. 2014, 164, 1831–1841. [Google Scholar] [CrossRef]
- Tournier, V.; Topham, C.M.; Gilles, A.; David, B.; Folgoas, C.; Moya-Leclair, E.; Kamionka, E.; Desrousseaux, M.-L.; Texier, H.; Gavalda, S.; et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 2020, 580, 216–219. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.; Carleton, W.C.; Groucutt, H.S. Climate change, not human population growth, correlates with Late Quaternary megafauna declines in North America. Nat. Commun. 2021, 12, 965. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.; Norder, S.; Ah-Peng, C.; Borges, P.A.; Burns, K.C.; de Nascimento, L.; Meyer, J.-Y.; Montes, E.; et al. Scientists’ warning—The outstanding biodiversity of islands is in peril. Glob. Ecol. Conserv. 2021, 31, e01847. [Google Scholar] [CrossRef]
- Jung, M.; Arnell, A.; de Lamo, X.; García-Rangel, S.; Lewis, M.; Mark, J.; Merow, C.; Miles, L.; Ondo, I.; Pironon, S.; et al. Areas of global importance for conserving terrestrial biodiversity, carbon and water. Nat. Ecol. Evol. 2021, 5, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.A.; Roemer, G.W.; Donlan, C.J.; Dickson, B.G.; Matocq, M.; Malaney, J.L. Ecology: Gene tweaking for conservation. Nature 2013, 501, 485–486. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibanez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418–10423. [Google Scholar] [CrossRef] [PubMed]
- Bay, R.A.; Harrigan, R.J.; Le Underwood, V.; Gibbs, H.L.; Smith, T.B.; Ruegg, K. Genomic signals of selection predict climate-driven population declines in a migratory bird. Science 2018, 359, 83–86. [Google Scholar] [CrossRef]
- Bay, R.A.; Rose, N.; Barrett, R.; Bernatchez, L.; Ghalambor, C.; Lasky, J.R.; Brem, R.B.; Palumbi, S.R.; Ralph, P. Predicting Responses to Contemporary Environmental Change Using Evolutionary Response Architectures. Am. Nat. 2017, 189, 463–473. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef]
- Coates, D.J.; Byrne, M.; Moritz, C. Genetic Diversity and Conservation Units: Dealing With the Species-Population Continuum in the Age of Genomics. Front. Ecol. Evol. 2018, 6, 165. [Google Scholar] [CrossRef]
- Wolf, J.; Ellegren, J.B.W.W.H. Making sense of genomic islands of differentiation in light of speciation. Nat. Rev. Genet. 2017, 18, 87–100. [Google Scholar] [CrossRef]
- McGee, M.D.; Borstein, S.R.; Meier, J.I.; Marques, D.A.; Mwaiko, S.; Taabu, A.; Kishe, M.A.; O’Meara, B.; Bruggmann, R.; Excoffier, L.; et al. The ecological and genomic basis of explosive adaptive radiation. Nature 2020, 586, 75–79. [Google Scholar] [CrossRef]
- Thomson, A.I.; Archer, F.I.; Coleman, M.A.; Gajardo, G.; Goodall-Copestake, W.P.; Hoban, S.; Laikre, L.; Miller, A.D.; O’Brien, D.; Pérez-Espona, S.; et al. Charting a course for genetic diversity in the UN Decade of Ocean Science. Evol. Appl. 2021, 14, 1497–1518. [Google Scholar] [CrossRef]
- Karger, B.L.; Guttman, A. DNA sequencing by CE. Electrophoresis 2009, 30, S196–S202. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Queralt-Rosinach, N.; Bravo, Àlex; Deu-Pons, J.; Bauer-Mehren, A.; Baron, M.; Sanz, F.; Furlong, L.I. DisGeNET: A discovery platform for the dynamical exploration of human diseases and their genes. Database 2015, 2015, bav028. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; Van Der Oost, J. Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR Interference Limits Horizontal Gene Transfer in Staphylococci by Targeting DNA. Science 2008, 322, 1843–1845. [Google Scholar] [CrossRef]
- Hale, C.R.; Zhao, P.; Olson, S.; Duff, M.O.; Graveley, B.R.; Wells, L.; Terns, R.M.; Terns, M.P. RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex. Cell 2009, 139, 945–956. [Google Scholar] [CrossRef]
- Garneau, J.E.; Dupuis, M.-È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef]
- Gao, X.; Tao, Y.; Lamas, V.; Huang, M.; Yeh, W.-H.; Pan, B.; Hu, Y.-J.; Hu, J.H.; Thompson, D.B.; Shu, Y.; et al. Treatment of autosomal dominant hearing loss by in vivo delivery of genome editing agents. Nature 2018, 553, 217–221. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, J.; Roscoe, B.P.; Liu, P.; Yao, Q.; Lazzarotto, C.R.; Clement, M.K.; Cole, M.; Luk, K.; Baricordi, C.; et al. Highly efficient therapeutic gene editing of human hematopoietic stem cells. Nat. Med. 2019, 25, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Giuliano, C.J.; Sayles, N.M.; Sheltzer, J.M. CRISPR/Cas9 mutagenesis invalidates a putative cancer dependency targeted in on-going clinical trials. eLife 2017, 6, e24179. [Google Scholar] [CrossRef] [PubMed]
- Raikwar, S.P.; Kikkeri, N.S.; Sakuru, R.; Saeed, D.; Zahoor, H.; Premkumar, K.; Mentor, S.; Thangavel, R.; Dubova, I.; Ahmed, M.E.; et al. Next Generation Precision Medicine: CRISPR-mediated Genome Editing for the Treatment of Neurodegenerative Disorders. J. Neuroimmune Pharmacol. 2019, 14, 608–641. [Google Scholar] [CrossRef] [PubMed]
- Pursey, E.; Sünderhauf, D.; Gaze, W.H.; Westra, E.R.; Van Houte, S. CRISPR-Cas antimicrobials: Challenges and future prospects. PLoS Pathog. 2018, 14, e1006990. [Google Scholar] [CrossRef] [PubMed]
- Knott, G.J.; Doudna, J.A. CRISPR-Cas guides the future of genetic engineering. Science 2018, 361, 866–869. [Google Scholar] [CrossRef]
- Loureiro, A.; Da Silva, G.J. CRISPR-Cas: Converting A Bacterial Defence Mechanism into A State-of-the-Art Genetic Manipulation Tool. Antibiotics 2019, 8, 18. [Google Scholar] [CrossRef]
- Kaminski, M.M.; Abudayyeh, O.O.; Gootenberg, J.S.; Zhang, F.; Collins, J.J. CRISPR-based diagnostics. Nat. Biomed. Eng. 2021, 5, 643–656. [Google Scholar] [CrossRef]
- Haque, E.; Taniguchi, H.; Hassan, M.; Bhowmik, P.; Karim, M.R.; Śmiech, M.; Zhao, K.; Rahman, M.; Islam, T. Application of CRISPR/Cas9 Genome Editing Technology for the Improvement of Crops Cultivated in Tropical Climates: Recent Progress, Prospects, and Challenges. Front. Plant Sci. 2018, 9, 617. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef]
- Lino, C.A.; Harper, J.C.; Carney, J.P.; Timlin, J.A. Delivering CRISPR: A review of the challenges and approaches. Drug Deliv. 2018, 25, 1234–1257. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. Ther. 2020, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, C.T.; Deshpande, P.S.; Dever, D.P.; Camarena, J.; Lemgart, V.T.; Cromer, M.K.; Vakulskas, C.A.; Collingwood, M.A.; Zhang, L.; Bode, N.M.; et al. Identification of preexisting adaptive immunity to Cas9 proteins in humans. Nat. Med. 2019, 25, 249–254. [Google Scholar] [CrossRef]
- Ho, B.X.; Loh, S.J.H.; Chan, W.-K.; Soh, B.S. In Vivo Genome Editing as a Therapeutic Approach. Int. J. Mol. Sci. 2018, 19, 2721. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.; Mahfouz, M.M. Genome editing: The road of CRISPR/Cas9 from bench to clinic. Exp. Mol. Med. 2016, 48, e265. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Platt, R.; Zhang, F. Therapeutic genome editing: Prospects and challenges. Nat. Med. 2015, 21, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, B.L. Homing Endonucleases: From Microbial Genetic Invaders to Reagents for Targeted DNA Modification. Structure 2011, 19, 7–15. [Google Scholar] [CrossRef]
- Zhou, F.; Zhu, H.; Fu, C. Editorial: Clinical Therapeutic Development Against Cancers Resistant to Targeted Therapies. Front. Pharmacol. 2022, 12, 816896. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine. ClinicalTrials.gov. 2021. Available online: https://clinicaltrials.gov/ct2/home (accessed on 9 September 2021).
- Reyes, A.P.; Lanner, F. Towards a CRISPR view of early human development: Applications, limitations and ethical concerns of genome editing in human embryos. Development 2017, 144, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Cyranoski, D. The CRISPR-baby scandal: What’s next for human gene-editing. Nature 2019, 566, 440–442. [Google Scholar] [CrossRef]
- Wang, H.; Yang, H. Gene-edited babies: What went wrong and what could go wrong. PLoS Biol. 2019, 17, e3000224. [Google Scholar] [CrossRef]
- Lanphier, E.; Urnov, F.; Haecker, S.E.; Werner, M.; Smolenski, J. Don’t edit the human germ line. Nature 2015, 519, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Howard, H.C.; on behalf of the Public and Professional Policy Committee of the European Society of Human Genetics; van El, C.G.; Forzano, F.; Radojkovic, D.; Rial-Sebbag, E.; de Wert, G.; Borry, P.; Cornel, M.C. One small edit for humans, one giant edit for humankind? Points and questions to consider for a responsible way forward for gene editing in humans. Eur. J. Hum. Genet. 2018, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- De Wert, G.; Heindryckx, B.; Pennings, G.; Clarke, A.; Eichenlaub-Ritter, U.; van El, C.G.; Forzano, F.; Goddijn, M.; Howard, H.C.; Radojkovic, D.; et al. Responsible innovation in human germline gene editing: Background document to the recommendations of ESHG and ESHRE. Eur. J. Hum. Genet. 2018, 26, 450–470. [Google Scholar] [CrossRef] [PubMed]
- Eissenberg, J.C. In Our Image: The Ethics of CRISPR Genome Editing. Biomol. Concepts 2021, 12, 1–7. [Google Scholar] [CrossRef]
- de Wert, G.; Pennings, G.; Clarke, A.; Eichenlaub-Ritter, U.; van El, C.G.; Forzano, F.; Goddijn, M.; Heindryckx, B.; Howard, H.C.; Radojkovic, D.; et al. Human germline gene editing: Recommendations of ESHG and ESHRE. Eur. J. Hum. Genet. 2018, 26, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Mizushima, N. Autophagy and human diseases. Cell Res. 2014, 24, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. CRISPR/Cas gene therapy. J. Cell. Physiol. 2021, 236, 2459–2481. [Google Scholar] [CrossRef]
- Brokowski, C.; Adli, M. CRISPR Ethics: Moral Considerations for Applications of a Powerful Tool. J. Mol. Biol. 2019, 431, 88–101. [Google Scholar] [CrossRef]
- Krekora-Zając, D. Civil liability for damages related to germline and embryo editing against the legal admissibility of gene editing. Palgrave Commun. 2020, 6, 30. [Google Scholar] [CrossRef]
- Pauli, R.M. Achondroplasia: A comprehensive clinical review. Orphanet J. Rare Dis. 2019, 14, 1. [Google Scholar] [CrossRef]
- Ledford, H. CRISPR babies: When will the world be ready? Nature 2019, 570, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Hoggatt, J. Gene Therapy for “Bubble Boy” Disease. Cell 2016, 166, 263. [Google Scholar] [CrossRef] [PubMed]
- Vassena, R.; Heindryckx, B.; Peco, R.; Pennings, G.; Raya, A.; Sermon, K.; Veiga, A. Genome engineering through CRISPR/Cas9 technology in the human germline and pluripotent stem cells. Hum. Reprod. Updat. 2016, 22, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Plomin, R. Genotype-Environment Correlation in the Era of DNA. Behav. Genet. 2014, 44, 629–638. [Google Scholar] [CrossRef]
- Georgiades, E.; Klissouras, V.; Baulch, J.; Wang, G.; Pitsiladis, Y. Why nature prevails over nurture in the making of the elite athlete. BMC Genom. 2017, 18, 835. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Gao, Y.; Dominguez, A.A.; Qi, L.S. CRISPR technologies for precise epigenome editing. Nat. Cell Biol. 2021, 23, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.H. Genetic Assimilation of the Bithorax Phenotype. Evolution 1956, 10, 1–13. [Google Scholar] [CrossRef]
- Holliday, R. Epigenetics: A Historical Overview. Epigenetics 2006, 1, 76–80. [Google Scholar] [CrossRef]
- Song, Z.; Liu, J.; Han, J. Chromatin remodeling factors regulate environmental stress responses in plants. J. Integr. Plant Biol. 2021, 63, 438–450. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.; Goeman, J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.; Stok, A.P.; Thijssen, P.E.; Müller, F.; et al. DNA methylation signatures link prenatal famine exposure to growth and metabolism. Nat. Commun. 2014, 5, 5592. [Google Scholar] [CrossRef] [PubMed]
- Lumey, L.; Stein, A.D.; Susser, E. Prenatal Famine and Adult Health. Annu. Rev. Public Health 2011, 32, 237–262. [Google Scholar] [CrossRef]
- Clair, D.S. Rates of Adult Schizophrenia Following Prenatal Exposure to the Chinese Famine of 1959–1961. JAMA J. Am. Med. Assoc. 2005, 294, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Shchukina, I.; Bagaitkar, J.; Shpynov, O.; Loginicheva, E.; Porter, S.; Mogilenko, D.A.; Wolin, E.; Collins, P.; Demidov, G.; Artomov, M.; et al. Enhanced epigenetic profiling of classical human monocytes reveals a specific signature of healthy aging in the DNA methylome. Nat. Aging 2021, 1, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.S.; Adams, D.M.; Haghani, A.; Lu, A.T.; Zoller, J.; Breeze, C.E.; Arnold, B.D.; Ball, H.C.; Carter, G.G.; Cooper, L.N.; et al. DNA methylation predicts age and provides insight into exceptional longevity of bats. Nat. Commun. 2021, 12, 1615. [Google Scholar] [CrossRef]
- Yang, G.; Huang, X. Methods and applications of CRISPR/Cas system for genome editing in stem cells. Cell Regen. 2019, 8, 33–41. [Google Scholar] [CrossRef]
- Collins, F.S. NIH Director’s Blog; Morgan, K., Ed.; National Institutes of Health, U.S. Department of Health and Human Services: Atlanta, GA, USA, 2020; Volume 2021.
- Haapaniemi, E.; Botla, S.; Persson, J.; Schmierer, B.; Taipale, J. CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response. Nat. Med. 2018, 24, 927–930. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR-Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef]
- Arnold, C. What’s new in clinical CRISPR? Nat. Med. 2021, 27, 184–185. [Google Scholar] [CrossRef]
- Callaway, E.; Ledford, H. Pioneers of revolutionary CRISPR gene editing win chemistry Nobel. Nature 2020, 586, 346–347. [Google Scholar] [CrossRef]
- Schubert, M.G.; Goodman, D.B.; Wannier, T.M.; Kaur, D.; Farzadfard, F.; Lu, T.K.; Shipman, S.L.; Church, G.M. High-throughput functional variant screens via in vivo production of single-stranded DNA. Proc. Natl. Acad. Sci. USA 2021, 118, e2018181118. [Google Scholar] [CrossRef] [PubMed]
- Sibbald, B. Death but one unintended consequence of gene-therapy trial. Can. Med. Assoc. J. 2001, 164, 1612. [Google Scholar]
- Savulescu, J. Harm, ethics committees and the gene therapy death. J. Med. Ethics 2001, 27, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Zilberman, D.; Holland, T.G.; Trilnick, I. Agricultural GMOs—What We Know and Where Scientists Disagree. Sustainability 2018, 10, 1514. [Google Scholar] [CrossRef]
- Klümper, W.; Qaim, M. A Meta-Analysis of the Impacts of Genetically Modified Crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef]
- Carlson, D.; Lancto, C.A.; Zang, B.; Kim, E.-S.; Walton, M.; Oldeschulte, D.; Seabury, C.; Sonstegard, T.S.; Fahrenkrug, S.C. Production of hornless dairy cattle from genome-edited cell lines. Nat. Biotechnol. 2016, 34, 479–481. [Google Scholar] [CrossRef] [PubMed]
- McConnachie, E.; Hötzel, M.J.; Robbins, J.A.; Shriver, A.; Weary, D.M.; Von Keyserlingk, M.A.G. Public attitudes towards genetically modified polled cattle. PLoS ONE 2019, 14, e0216542. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, J.F.; Rossello, F.J.; Tan, J.P.; Davidson, K.C.; Valdes, D.S.; Schröder, J.; Sun, Y.B.Y.; Chen, J.; Knaupp, A.S.; et al. Reprogramming roadmap reveals route to human induced trophoblast stem cells. Nature 2020, 586, 101–107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Kingdom | Species | Relevance | Estimated Genome Size (Mbp) | Reference |
| Animalia | Aedes mosquito (Aedes aegypti) | Primary vector for yellow and dengue fevers | 1380 | [37] |
| Cattle (Bos taurus) | Ruminant biology and evolution | 2870 | [38] | |
| Coelacanth (Latimeria chalumnae) | Tetrapod evolution | 2860 | [39] | |
| Common chimpanzee (Pan troglodytes) | Model organism (human population genetics and evolution) | 2400 | [40] | |
| Common marmoset (Callithrix jacchus) | Biomedical research application | 2260 | [41] | |
| Giant panda (Ailuropoda melanoleuca) | Foundation for promoting mammalian genetic research | 2250 | [42] | |
| Honeybee (Apis mellifera) | Model organism (social behaviour and global ecology) | 1800 | [43] | |
| Japanese medaka (Oryzias latipes) | Vertebrate evolution | 700 | [44] | |
| Pacific oyster (Crassostrea gigas) | Lophotrochozoa evolution | 559 | [45] | |
| Platypus (Ornithorhynchus anatinus) | Model organism (combination of reptilian and mammalian characters) | 1840 | [46] | |
| Red flour beetle (Tribolium castaneum) | Model organism (beetle and pest) | 160 | [47] | |
| Sea urchin (Strongylocentrotus purpuratus) | Model organism (developmental and system biology) | 814 | [48] | |
| Sponges (Amphimedon queenslandica) | Animal origins and early evolution | 167 | [49] | |
| Two-spotted spider mite (Tetranychus urticae) | Cosmopolitan agricultural pest | 90 | [50] | |
| Western gorilla (Gorilla gorilla) | Human origins and evolution | 5400 | [51] | |
| Mexican oxolotl (Ambystoma mexicanum) | Evolutionary changes in key tissue formation regulators | 32,000 | [52] | |
| Galapagos cormorant (Phalacrocorax harrisi) | Evolutionary changes in the size and proportion of limbs | 1200 | [53] | |
| Golden orb-weaver (Nephila clavipes) | Diversity of spider silk genes and their complex expression | 2440 | [54] | |
| Plantae | African oil palm (Elaeis guineensis) | Oil-bearing crop | 1800 | [55] |
| Amborella (Amborella trichopoda) | Angiosperm evolution | 870 | [56] | |
| Barrel medic (Medicago truncatula) | Model organism (legume) | 246 | [57] | |
| China rose (Rosa chinensis) | Model organism (ornamental plant) | 560 | [58] | |
| Dwarf banana (Musa acuminata) | A genome of modern cultivar | 523 | [59] | |
| Maize (Zea mays) | Major cereal crop | 2300 | [60] | |
| Papaya (Carica papaya) | Tropical fruit crop | 372 | [61] | |
| Peanut (A. duranensis, A. ipaensis, A. hypogaea) | Polyploid genetic mechanisms | 2540 | [62,63] | |
| Pigeon pea (Cajanus cajan) | Model organism (legume) | 833 | [64] | |
| Potato (Solanum tuberosum) | Major root crop | 844 | [65] | |
| Quinoa (Chenopodium quinoa) | Future crop | 1500 | [66] | |
| Rose gum (Eucalyptus grandis) | Fibre and timber crop | 640 | [67] | |
| Sorghum (Sorghum bicolor) | Major cereal crop | 730 | [68] | |
| Soybean (Glycine max) | Major protein and oil crop | 1115 | [69] | |
| Tomato (Solanum lycopersicum) | Major vegetable crop | 900 | [70] | |
| Silver birch (Betula pendula) | Model organism (forest biotechnology) | 440 | [71] | |
| Durian (Durio zibethinus) | Tropical fruit biology and agronomy | 738 | [72] | |
| Sunflower (Helianthus annuus) | Oil metabolism, flowering, and Asterid evolution | 3600 | [73] | |
| Tausch’s goatgrass (Aegilops tauschii) | Genetic resources for wheat | 4300 | [74] | |
| Barley (Hordeum vulgare) | Major cereal crop | 4800 | [75] | |
| Pearl millet (Pennisetum glaucum) | Future crop | 1790 | [76] | |
| Fungi | Black mold (Aspergillus niger) | Model fungal | 34 | [77] |
| Filamentous fungus (Aspergillus nidulans, A. fumigatus, A. oryzae) | Model fungal | 40 | [78] | |
| Fission yeast (Schizosaccharomyces pombe) | Model yeast | 14 | [79] | |
| Rice blast fungus (Magnaporthe grisea) | Model fungal | 40 | [80] | |
| Split gill (Schizophyllum commune) | Model mushroom | 39 | [81] | |
| Yeast (Candida albicans) | Human pathogen | 4 | [82] | |
| Filamentous fungus (Penicillium chrysogenum) | Industrial use | 32 | [83] |
| Property | CRISPR/Cas9 | TALEN | ZFN | Meganuclease (Homing Endonucleases) |
|---|---|---|---|---|
| Essential components | sgRNA and Cas9 | TALE and FokI | ZFP and FokI | Meganuclease (nuclease domain) |
| Backbone origin | Bacteria (Streptococcus pyogenes) | Bacteria (Xanthomonas spp.) | Mostly prevalent in eukaryotes | Microbial mobile genetic elements |
| Ease of engineering | Easy; facile design of gRNA, standard cloning methods and oligo synthesis | Moderate; complex cloning methods are required | Difficult; substantial protein engineering is required | Difficult; substantial protein engineering is required |
| Recognition site | 22 bp (20-bp guide sequence and 2-bp protospacer adjacent motif (PAM) for Cas9); 44 bp for double nicking | 28–40 bp per TALEN pair | 18–36 bp per ZFN pair; guanine-rich region | 14–40 bp |
| Specificity | Highly predictable (DNA–RNA interaction). Multiple mismatches tolerated | Less predictable (DNA–protein interaction). Small number of mismatches tolerated | Less predictable (DNA–protein interaction). Small number of mismatches tolerated | Less predictable (DNA–protein interaction). Small number of mismatches tolerated |
| Targeting constraints | Targeted sequence must precede PAM | T must be the 5′ targeted base for each TALEN monomer | Non-G-rich sequences are difficult to target | Low efficiency in targeting novel sequences |
| Ease of in vivo delivery | Moderate | Difficult | Relatively easy | Relatively easy |
| Multiplexing ability | Feasible | Challenging | Challenging | Challenging |
| Affordability | Highly affordable (1–3 days) | Affordable but time consuming (5–7 days) | Resource intensive and time consuming (7–15 days) | Resource intensive and time consuming (up to 100 days) |
| Methylation sensitivity | No | Sensitive | Sensitive | Sensitive |
| Clinical or pre-clinical stage | Clinical trial application for refractory non-small-cell lung cancer, sickle cell disease, and beta-thalassemia | Clinical trial application for relapse or refractory acute myeloid leukaemia | Clinical trial application for HIV and Hunter’s syndrome | Clinical trial application for non-Hodgkin Lymphoma and multiple myeloma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, A.; Harikrishna, J.A.; Redwood, C.S.; Lit, L.C.; Nath, S.K.; Chua, K.H. Genetics Matters: Voyaging from the Past into the Future of Humanity and Sustainability. Int. J. Mol. Sci. 2022, 23, 3976. https://doi.org/10.3390/ijms23073976
Cheng A, Harikrishna JA, Redwood CS, Lit LC, Nath SK, Chua KH. Genetics Matters: Voyaging from the Past into the Future of Humanity and Sustainability. International Journal of Molecular Sciences. 2022; 23(7):3976. https://doi.org/10.3390/ijms23073976
Chicago/Turabian StyleCheng, Acga, Jennifer Ann Harikrishna, Charles S. Redwood, Lei Cheng Lit, Swapan K. Nath, and Kek Heng Chua. 2022. "Genetics Matters: Voyaging from the Past into the Future of Humanity and Sustainability" International Journal of Molecular Sciences 23, no. 7: 3976. https://doi.org/10.3390/ijms23073976
APA StyleCheng, A., Harikrishna, J. A., Redwood, C. S., Lit, L. C., Nath, S. K., & Chua, K. H. (2022). Genetics Matters: Voyaging from the Past into the Future of Humanity and Sustainability. International Journal of Molecular Sciences, 23(7), 3976. https://doi.org/10.3390/ijms23073976