
Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients


 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
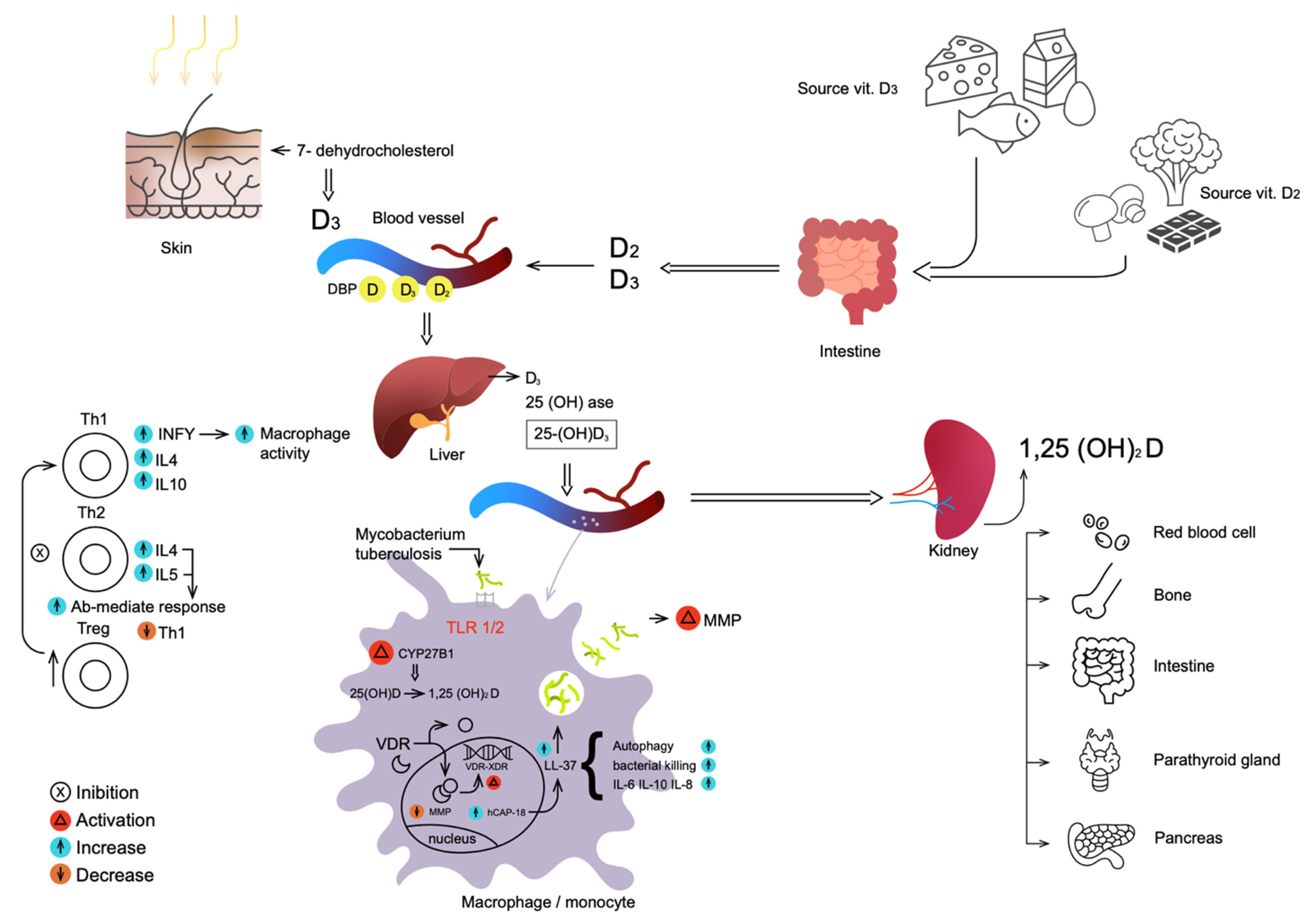
2. Metabolism of Vitamin D
3. Vitamin D Deficiency
4. Vitamin D and Tuberculosis
4.1. Innate Immunity
4.2. Acquired Immunity
4.3. Anti-Inflammatory Activity
5. Polymorphisms in VDR
6. The Role of Vitamin D in Prevention of Tuberculosis
7. The Role of Vitamin D in Treatment of Tuberculosis
8. Synergism with Vitamin A in Prevention and Treatment of TB
9. Side Effects of Vitamin D Supplementation
10. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, R.; Christakos, S. Mechanisms Underlying the Regulation of Innate and Adaptive Immunity by Vitamin D. Nutrients 2015, 7, 8251–8260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riek, A.E.; Rajagopal, R.; Bernal-Mizrachi, C. Vitamin D and the cardiovascular system. In Vitamin D; Academic Press: Cambridge, MA, USA, 2018; pp. 545–562. [Google Scholar]
- Gil, Á.; Plaza-Diaz, J.; Mesa, M.D. Vitamin D: Classic and Novel Actions. Ann. Nutr. Metab. 2018, 72, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.M.; Shin, E.A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, P.D.; Chen, W.Y.; Ajala, O.N.; Hazra, A.; Cook, N.; Bubes, V.; Lee, I.M.; Giovannucci, E.L.; Willett, W.; Buring, J.E.; et al. VITAL Research Group Effect of Vitamin D3 Supplements on Development of Advanced Cancer: A Secondary Analysis of the VITAL Randomized Clinical Trial. JAMA Netw. Open 2020, 3, e2025850. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Bhalla, A.K.; Amento, E.P.; Clemens, T.L.; Holick, M.F.; Krane, S.M. Specific high-affinity receptors for 1,25-dihydroxyvitamin D3 in human peripheral blood mononuclear cells: Presence in monocytes and induction in T lymphocytes following activation. J. Clin. Endocrinol. Metab. 1983, 57, 1308–1310. [Google Scholar] [CrossRef] [PubMed]
- Provvedini, D.M.; Tsoukas, C.D.; Deftos, L.J.; Manolagas, S.C. 1,25-Dihydroxyvitamin D 3 Receptors in Human Leukocytes. Science 1983, 221, 1181–1183. [Google Scholar] [CrossRef] [PubMed]
- Khajavi, A.; Amirhakimi, G.H. The rachitic lung. Pulmonary findings in 30 infants and children with malnutritional rickets. Clin. Pediatr. 1977, 16, 36–38. [Google Scholar] [CrossRef] [PubMed]
- Martineau, A.R.; Jolliffe, D.A.; Hooper, R.L.; Greenberg, L.; Aloia, J.F.; Bergman, P.; Dubnov-Raz, G.; Esposito, S.; Ganmaa, D.; Ginde, A.A.; et al. Vitamin D supplementation to prevent acute respiratory tract infections: Systematic review and meta-analysis of individual participant data. Br. J. Med. 2017, 356, i6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Tuberculosis Report. 2021. Available online: https://www.who.int/publications/i/item/9789240037021 (accessed on 20 February 2022).
- Minardi, M.L.; Fato, I.; Di Gennaro, F.; Mosti, S.; Mastrobattista, A.; Cerva, C.; Libertone, R.; Saracino, A.; Goletti, D.; Girardi, E.; et al. Common and Rare Hematological Manifestations and Adverse Drug Events during Treatment of Active TB: A State of Art. Microorganisms 2021, 9, 1477. [Google Scholar] [CrossRef]
- Turnbull, E.R.; Drobniewski, F. Vitamin D supplementation: A comprehensive review on supplementation for tuberculosis prophylaxis. Expert Rev. Respir. Med. 2015, 9, 269–275. [Google Scholar] [CrossRef]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M.; Endocrine Society. Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesareo, R.; Attanasio, R.; Caputo, M.; Castello, R.; Chiodini, I.; Falchetti, A.; Guglielmi, R.; Papini, E.; Santonati, A.; Scillitani, A.; et al. AME and Italian AACE Chapter Italian Association of Clinical Endocrinologists (AME) and Italian Chapter of the American Association of Clinical Endocrinologists (AACE) Position Statement: Clinical Management of Vitamin D Deficiency in Adults. Nutrients 2018, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Bischoff-Ferrari, H.A.; Giovannucci, E.; Willett, W.C.; Dietrich, T.; Dawson-Hughes, B. Estimation of optimal serum concentrations of 25-hydroxyvitamin D for multiple health outcomes. Am. J. Clin. Nutr. 2006, 84, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Junaid, K.; Rehman, A. Impact of vitamin D on infectious disease-tuberculosis—A review. Clin. Nutr. Exp. 2019, 25, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Roelandts, R. A new light on Niels Finsen, a century after his nobel prize. Photodermatol. Photoimmunol. Photomed. 2005, 21, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Sancho-Vaello, E.; Gil-Carton, D.; François, P.; Bonetti, E.-J.; Kreir, M.; Pothula, K.R.; Kleinekathöfer, U.; Zeth, K. The structure of the antimicrobial human cathelicidin LL-37 shows oligomerization and channel formation in the presence of membrane mimics. Sci. Rep. 2020, 10, 17356. [Google Scholar] [CrossRef] [PubMed]
- Luong, K.V.; Nguyen, L.T. Impact of Vitamin D in the Treatment of Tuberculosis. Am. J. Med Sci. 2011, 341, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuk, J.M.; Shin, D.M.; Lee, H.M.; Yang, C.S.; Jin, H.S.; Kim, K.K.; Lee, Z.W.; Lee, S.H.; Kim, J.M.; Jo, E.K. Vitamin D3 Induces Autophagy in Human Monocytes/Macrophages via Cathelicidin. Cell Host Microbe 2009, 6, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.M.; Yuk, J.M.; Lee, H.M.; Lee, S.H.; Son, J.W.; Harding, C.V.; Kim, J.M.; Modlin, R.L.; Jo, E.K. Mycobacterial lipoprotein activates autophagy via TLR2/1/CD14 and a functional vitamin D receptor signalling. Cell. Microbiol. 2010, 12, 1648–1665. [Google Scholar] [CrossRef] [Green Version]
- Rekha, R.S.; Muvva, S.J.R.; Wan, M.; Raqib, R.; Bergman, P.; Brighenti, S.; Gudmundsson, G.H.; Agerberth, B. Phenylbutyrate induces LL-37-dependent autophagy and intracellular killing of Mycobacterium tuberculosis in human macrophages. Autophagy 2015, 11, 1688–1699. [Google Scholar] [CrossRef] [Green Version]
- Mily, A.; Rekha, R.S.; Kamal, S.M.; Akhtar, E.; Sarker, P.; Rahim, Z.; Gudmundsson, G.H.; Agerberth, B.; Raqib, R. Oral intake of phenylbutyrate with or without vitamin D3upregulates the cathelicidin LL-37 in human macrophages: A dose finding study for treatment of tuberculosis. BMC Pulm. Med. 2013, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Rockett, K.A.; Brookes, R.; Udalova, I.; Vidal, V.; Hill, A.V.; Kwiatkowski, D. 1,25-Dihydroxyvitamin D3 induces nitric oxide synthase and suppresses growth of Mycobacterium tuberculosis in a human macrophage-like cell line. Infect. Immun. 1998, 66, 5314–5321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabri, M.; Stenger, S.; Shin, D.-M.; Yuk, J.-M.; Liu, P.T.; Realegeno, S.; Lee, H.-M.; Krutzik, S.R.; Schenk, M.; Sieling, P.A.; et al. Vitamin D Is Required for IFN-γ–Mediated Antimicrobial Activity of Human Macrophages. Sci. Transl. Med. 2011, 3, 104ra102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coussens, A.; Timms, P.M.; Boucher, B.J.; Venton, T.R.; Ashcroft, A.T.; Skolimowska, K.H.; Newton, S.M.; Wilkinson, K.A.; Davidson, R.N.; Griffiths, C.J.; et al. 1α,25-dihydroxyvitamin D3inhibits matrix metalloproteinases induced byMycobacterium tuberculosisinfection. Immunology 2009, 127, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, S.; Bergman, P.; Martineau, A.R. Vitamin D and tuberculosis: Where next? J. Intern. Med. 2018, 284, 145–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, C.C.; Davis, C.T.; Zhu, W.; Bowman-Kirigin, J.A.; Walker, A.E.; Tai, Z.; Thomas, K.R.; Donato, A.J.; Lesniewski, L.A.; Li, D.Y. Dietary Vitamin D and Its Metabolites Non-Genomically Stabilize the Endothelium. PLoS ONE 2015, 10, e0140370. [Google Scholar] [CrossRef] [Green Version]
- Torres-Juarez, F.; Cardenas-Vargas, A.; Montoya-Rosales, A.; González-Curiel, I.; Garcia-Hernandez, M.H.; Enciso-Moreno, J.A.; Hancock, R.E.; Rivas-Santiago, B. LL-37 Immunomodulatory Activity during Mycobacterium tuberculosis Infection in Macrophages. Infect. Immun. 2015, 83, 4495–4503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhang, C.Y.; Wu, X.M.; Tian, L.; Li, C.Z.; Zhao, Q.M.; Zhang, P.H.; Yang, S.M.; Yang, H.; Zhang, X.T.; et al. A case-control study on the vitamin D receptor gene polymorphisms and susceptibility to pulmonary tuberculosis. Zhonghua Liu Xing Bing Xue Za Zhi 2003, 24, 389–392. (In Chinese) [Google Scholar] [PubMed]
- Pizzol, D.; Di Gennaro, F.; Chhaganlal, K.D.; Fabrizio, C.; Monno, L.; Putoto, G.; Saracino, A. Prevalence of diabetes mellitus in newly diagnosed pulmonary tuberculosis in Beira, Mozambique. Afr. Health Sci. 2017, 17, 773–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, R.J.; Llewelyn, M.; Toossi, Z.; Patel, P.; Pasvol, G.; Lalvani, A.; Wright, D.; Latif, M.; Davidson, R.N. Influence of vitamin D deficiency and vitamin D receptor polymorphisms on tuberculosis among Gujarati Asians in west London: A case-control study. Lancet 2000, 355, 618–621. [Google Scholar] [CrossRef]
- Roth, D.E.; Soto, G.; Arenas, F.; Bautista, C.T.; Ortiz, J.; Rodriguez, R.; Cabrera, L.; Gilman, R.H. Association between Vitamin D Receptor Gene Polymorphisms and Response to Treatment of Pulmonary Tuberculosis. J. Infect. Dis. 2004, 190, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Valdivielso, J.M.; Fernandez, E. Vitamin D receptor polymorphisms and diseases. Clin. Chim. Acta 2006, 371, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Timms, P.M.; Mannan, N.; Hitman, G.A.; Noonan, K.; Mills, P.G.; Syndercombe-Court, D.; Aganna, E.; Price, C.P.; Boucher, B.J. Circulating MMP9, vitamin D and variation in the TIMP-1 response with VDR genotype: Mechanisms for inflammatory damage in chronic disorders? QJM Int. J. Med. 2002, 95, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.D.; Schwartz, Z.; Schmitz, J.; Muniz, O.E.; Lu, Y.; Calderón, F.; Howell, D.S.; Boyan, B.D. Vitamin D Regulation of Metal loproteinase Activity in Matrix Vesicles. Connect. Tissue Res. 1996, 35, 331–336. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.J. A meta-analysis on associations between vitamin D receptor genetic variants and tuberculosis. Microb. Pathog. 2019, 130, 59–64. [Google Scholar] [CrossRef]
- Gao, L.; Tao, Y.; Zhang, L.; Jin, Q. Vitamin D receptor genetic polymorphisms and tuberculosis: Updated systematic review and meta-analysis. Int. J. Tuberc. Lung Dis. 2010, 14, 15–23. [Google Scholar] [PubMed]
- Huang, L.; Liu, C.; Liao, G.; Yang, X.; Tang, X.; Chen, J. Vitamin D Receptor Gene FokI Polymorphism Contributes to Increasing the Risk of Tuberculosis: An Update Meta-Analysis. Medicine 2015, 94, e2256. [Google Scholar] [CrossRef] [PubMed]
- Elsafi, S.S.M.S.; Nour, B.M.; Abakar, A.D.; Omer, I.H.; Almugadam, B.S. Vitamin D level and it is association with the severity of pulmonary tuberculosis in patients attended to Kosti Teaching Hospital, Sudan. AIMS Microbiol. 2020, 6, 65–74. [Google Scholar] [CrossRef]
- Huang, S.J.; Wang, X.H.; Liu, Z.-D.; Cao, W.-L.; Han, Y.; Ma, A.-G.; Xu, S.-F. Vitamin D deficiency and the risk of tuberculosis: A meta-analysis. Drug Des. Dev. Ther. 2016, 11, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturini, E.; Facchini, L.; Martinez-Alier, N.; Novelli, V.; Galli, L.; De Martino, M.; Chiappini, E. Vitamin D and tuberculosis: A multicenter study in children. BMC Infect. Dis. 2014, 14, 652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowle, A.J.; Ross, E.J.; May, M.H. Inhibition by 1,25(OH)2-vitamin D3 of the multiplication of virulent tubercle bacilli in cultured human macrophages. Infect. Immun. 1987, 55, 2945–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganmaa, D.; Uyanga, B.; Zhou, X.; Gantsetseg, G.; Delgerekh, B.; Enkhmaa, D.; Khulan, D.; Ariunzaya, S.; Sumiya, E.; Bolortuya, B.; et al. Vitamin D Supplements for Prevention of Tuberculosis Infection and Disease. N. Engl. J. Med. 2020, 383, 359–368. [Google Scholar] [CrossRef]
- Aibana, O.; Huang, C.-C.; Aboud, S.; Arnedo-Pena, A.; Becerra, M.C.; Bellido-Blasco, J.B.; Bhosale, R.; Calderon, R.; Chiang, S.; Contreras, C.; et al. Vitamin D status and risk of incident tuberculosis disease: A nested case-control study, systematic review, and individual-participant data meta-analysis. PLoS Med. 2019, 16, e1002907. [Google Scholar] [CrossRef]
- Bouillon, R.; Manousaki, D.; Rosen, C.; Trajanoska, K.; Rivadeneira, F.; Richards, J.B. The health effects of vitamin D supplementation: Evidence from human studies. Nat. Rev. Endocrinol. 2022, 18, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, M.; Huang, Z.-X.; Bao, R.; Yu, Q.; Dai, M.; Wang, X.; Rao, Y. Calcitriol enhances pyrazinamide treatment of murine tuberculosis. Chin. Med. J. 2019, 132, 2089–2095. [Google Scholar] [CrossRef] [PubMed]
- Daley, P.; Jagannathan, V.; John, K.R.; Sarojini, J.; Latha, A.; Vieth, R.; Suzana, S.; Jeyaseelan, L.; Christopher, D.J.; Smieja, M.; et al. Adjunctive vitamin D for treatment of active tuberculosis in India: A randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2015, 15, 528–534. [Google Scholar] [CrossRef]
- Tukvadze, N.; Sanikidze, E.; Kipiani, M.; Hebbar, G.; Easley, K.; Shenvi, N.; Kempker, R.R.; Frediani, J.; Mirtskhulava, V.; Alvarez, J.A.; et al. High-dose vitamin D3 in adults with pulmonary tuberculosis: A double-blind randomized controlled trial. Am. J. Clin. Nutr. 2015, 102, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Grobler, L.; Nagpal, S.; Sudarsanam, T.D.; Sinclair, D. Nutritional supplements for people being treated for active tuberculosis. Cochrane Database Syst. Rev. 2016, 2016, CD006086. [Google Scholar] [CrossRef] [Green Version]
- Chesdachai, S.; Zughaier, S.M.; Hao, L.; Kempker, R.R.; Blumberg, H.M.; Ziegler, T.R.; Tangpricha, V. The effects of first-line anti-tuberculosis drugs on the actions of vitamin D in human macrophages. J. Clin. Transl. Endocrinol. 2016, 6, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keflie, T.S.; Nölle, N.; Lambert, C.; Nohr, D.; Biesalski, H.K. Vitamin D deficiencies among tuberculosis patients in Africa: A systematic review. Nutrition 2015, 31, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.S.; Tukvadze, N.; Frediani, J.K.; Kipiani, M.; Sanikidze, E.; Nichols, M.M.; Hebbar, G.; Kempker, R.R.; Mirtskhulava, V.; Kalandadze, I.; et al. Effects of sunlight and diet on vitamin D status of pulmonary tuberculosis patients in Tbilisi, Georgia. Nutrition 2012, 28, 362–366. [Google Scholar] [CrossRef] [Green Version]
- Sloan, D.J.; Mwandumba, H.C.; Kamdolozi, M.; Shani, D.; Chisale, B.; Dutton, J.; Khoo, S.H.; Allain, T.J.; Davies, G.R. Vitamin D deficiency in Malawian adults with pulmonary tuberculosis: Risk factors and treatment outcomes. Int. J. Tuberc. Lung Dis. 2015, 19, 904–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patti, G.; Pellegrino, C.; Ricciardi, A.; Novara, R.; Cotugno, S.; Papagni, R.; Guido, G.; Totaro, V.; De Iaco, G.; Romanelli, F.; et al. Potential Role of Vitamins A, B, C, D and E in TB Treatment and Prevention: A Narrative Review. Antibiotics 2021, 10, 1354. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.K.; Kaul, D.; Sharma, M. Synergistic action of vitamin D and retinoic acid restricts invasion of macrophages by pathogenic mycobacteria. J. Microbiol. Immunol. Infect. = Wei Mian Yu Gan Ran Za Zhi 2008, 41, 17–25. [Google Scholar]
- Malihi, Z.; Wu, Z.; Lawes, C.; Scragg, R. Adverse events from large dose vitamin D supplementation taken for one year or longer. J. Steroid Biochem. Mol. Biol. 2019, 188, 29–37. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Serum Value of 25(OH)D (ng/mL) | |
|---|---|
| 30–100 | Sufficiency |
| 20–30 | Insufficiency |
| <20 | Deficiency |
| Pro-Inflammatory/Antimicrobial Actions of Vit. D | Anti-Inflammatory Actions of Vit. D |
|---|---|
| Induces destruction of the bacterial cell by activating the cathelicidin/LL-37 system in infected macrophages | Induces the expansion of T-reg lymphocytes, which in turn limit the activity of Th1 |
| Induces autophagy in infected macrophages | Attenuates M. tuberculosis-induced expression of MMP |
| Inhibits the growth of MT in infected macrophages through the production of nitrogen and oxygen reactants | Stabilization of the endothelium and of the barrier function in the presence of inflammatory mediators |
| Stimulates production of INF-γ by Th1 lymphocytes, which enhances antibacterial activity of macrophages | Reduction of pro-inflammatory cytokines and increase of anti-inflammatory ones, without reducing the antimycobacterial activity |
| Enhances the antibody-mediated response by Th2 lymphocytes via the secretion of IL4 and IL5 |
| Polymorphism in VDR Gene | Allele’s Possible Combination | Effects |
|---|---|---|
| TaqI | TT | Higher cell turnover, reduction in TIMPs |
| Tt | Increase in culture conversion, quick negativization of sputum | |
| tt | Reduction of active TB | |
| Bsml | BB | Transcript stability |
| Bb | - | |
| bb | Significant association with active TB | |
| Fokl | FF | Increase in transcription, quick negativization of sputum |
| Ff | - | |
| ff | Extended form of pulmonary TB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papagni, R.; Pellegrino, C.; Di Gennaro, F.; Patti, G.; Ricciardi, A.; Novara, R.; Cotugno, S.; Musso, M.; Guido, G.; Ronga, L.; et al. Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients. Int. J. Mol. Sci. 2022, 23, 3860. https://doi.org/10.3390/ijms23073860
Papagni R, Pellegrino C, Di Gennaro F, Patti G, Ricciardi A, Novara R, Cotugno S, Musso M, Guido G, Ronga L, et al. Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients. International Journal of Molecular Sciences. 2022; 23(7):3860. https://doi.org/10.3390/ijms23073860
Chicago/Turabian StylePapagni, Roberta, Carmen Pellegrino, Francesco Di Gennaro, Giulia Patti, Aurelia Ricciardi, Roberta Novara, Sergio Cotugno, Maria Musso, Giacomo Guido, Luigi Ronga, and et al. 2022. "Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients" International Journal of Molecular Sciences 23, no. 7: 3860. https://doi.org/10.3390/ijms23073860
APA StylePapagni, R., Pellegrino, C., Di Gennaro, F., Patti, G., Ricciardi, A., Novara, R., Cotugno, S., Musso, M., Guido, G., Ronga, L., Stolfa, S., Bavaro, D. F., Romanelli, F., Totaro, V., Lattanzio, R., De Iaco, G., Palmieri, F., Saracino, A., & Gualano, G. (2022). Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients. International Journal of Molecular Sciences, 23(7), 3860. https://doi.org/10.3390/ijms23073860