
Analysis of microRNAs in Exosomes of Breast Cancer Patients in Search of Molecular Prognostic Factors in Brain Metastases

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Patient Characteristics
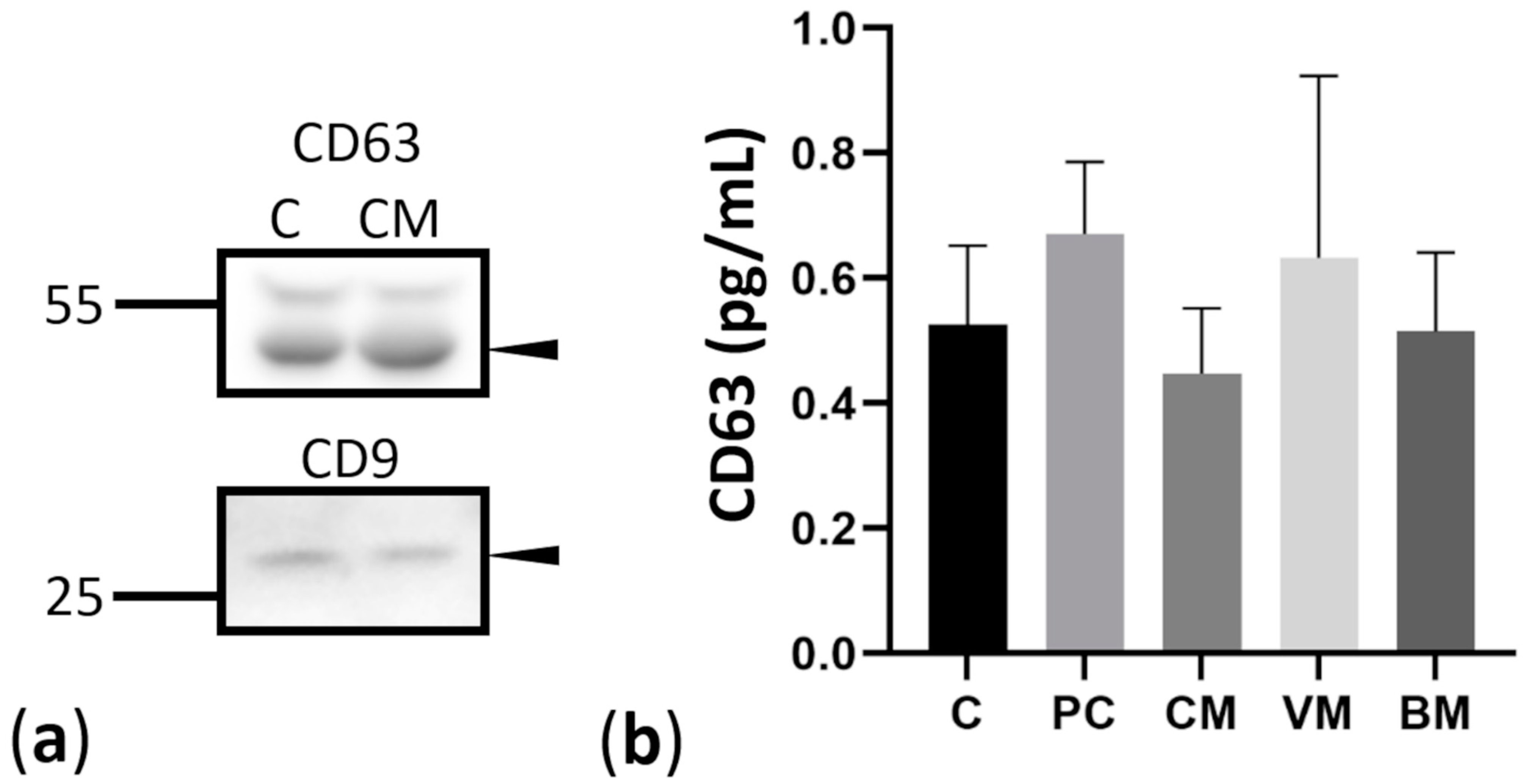
2.2. Isolation and Characterization of Exosomes Derived from Controls and Breast Cancer Patients with Brain Metastases
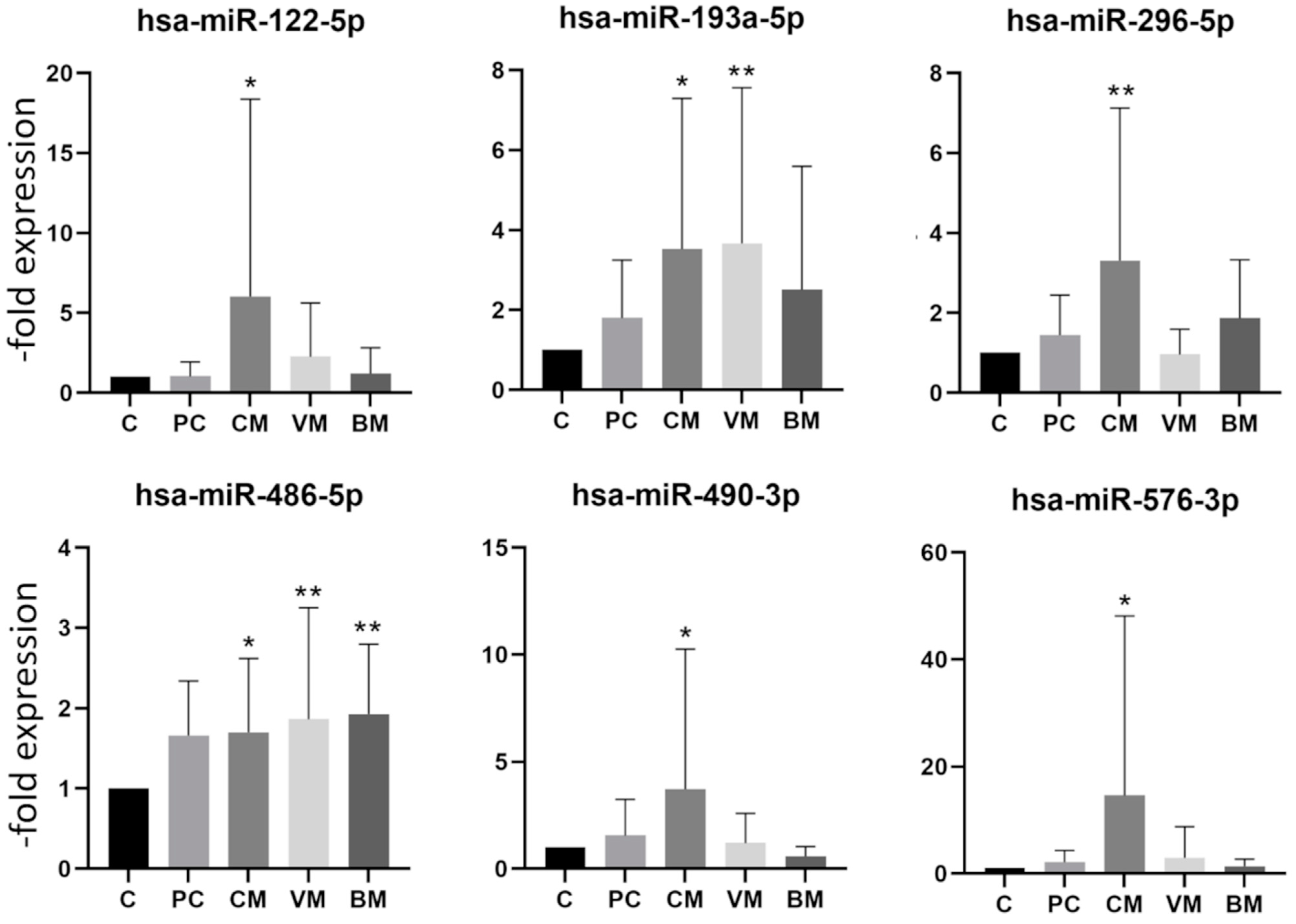
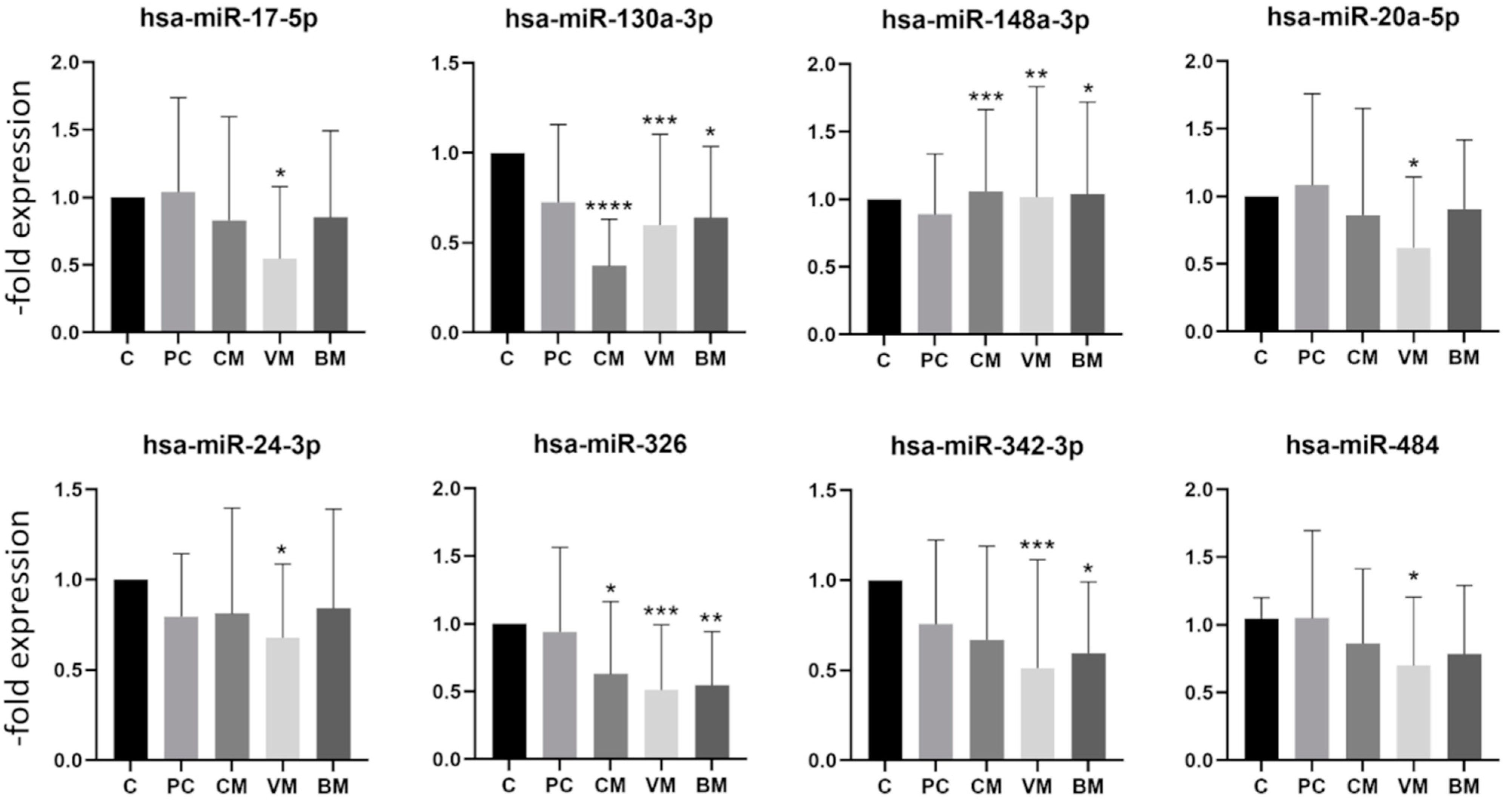
2.3. miRNA Expression in Exosomes Isolated from Patient Serum by Site of Metastasis
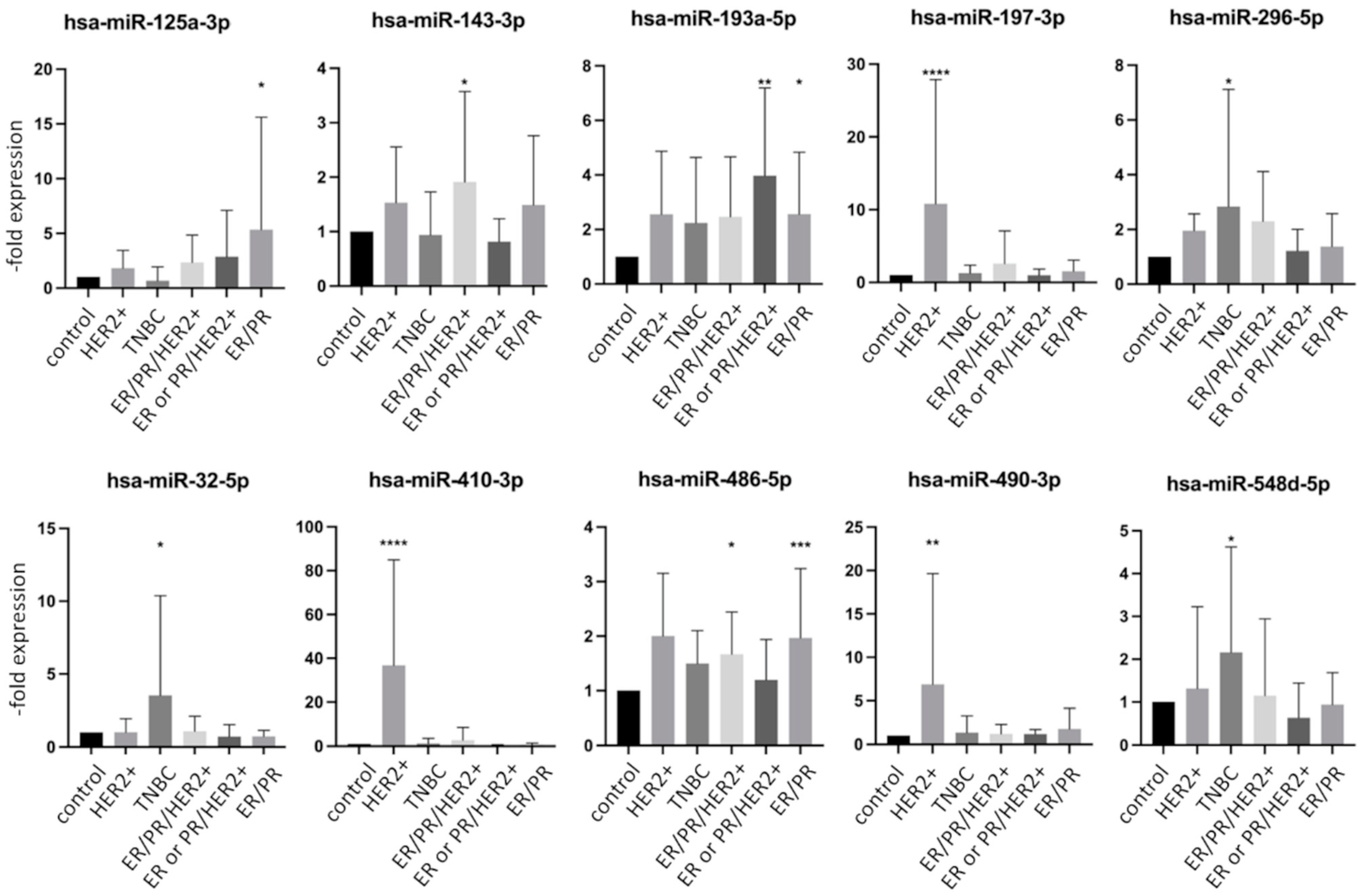
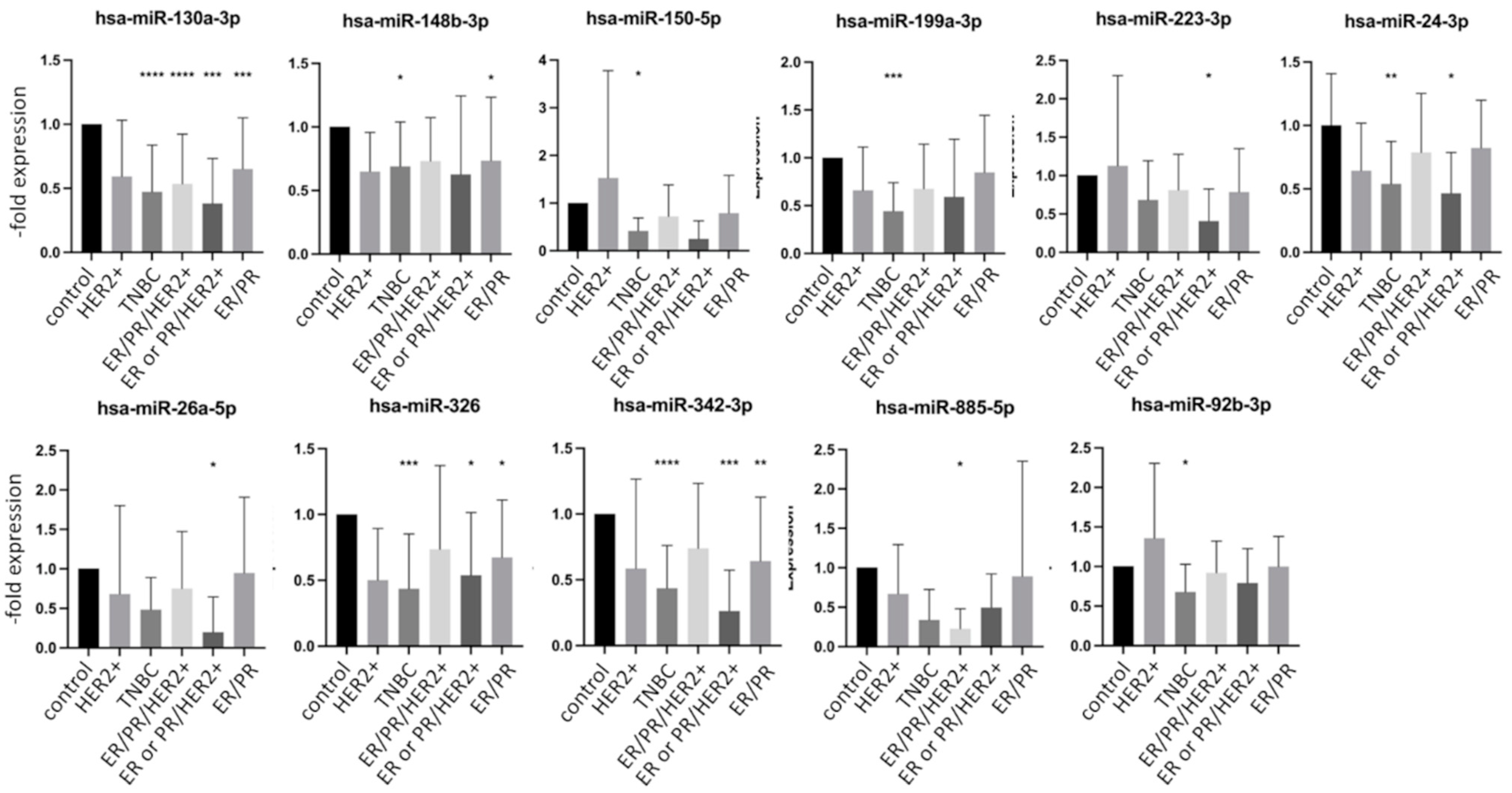
2.4. miRNA Expression in Exosomes Isolated from Patient Serum Classified by Tumor Characteristics
2.5. Screen of Databases for miR Targets and Signaling Pathways Involved
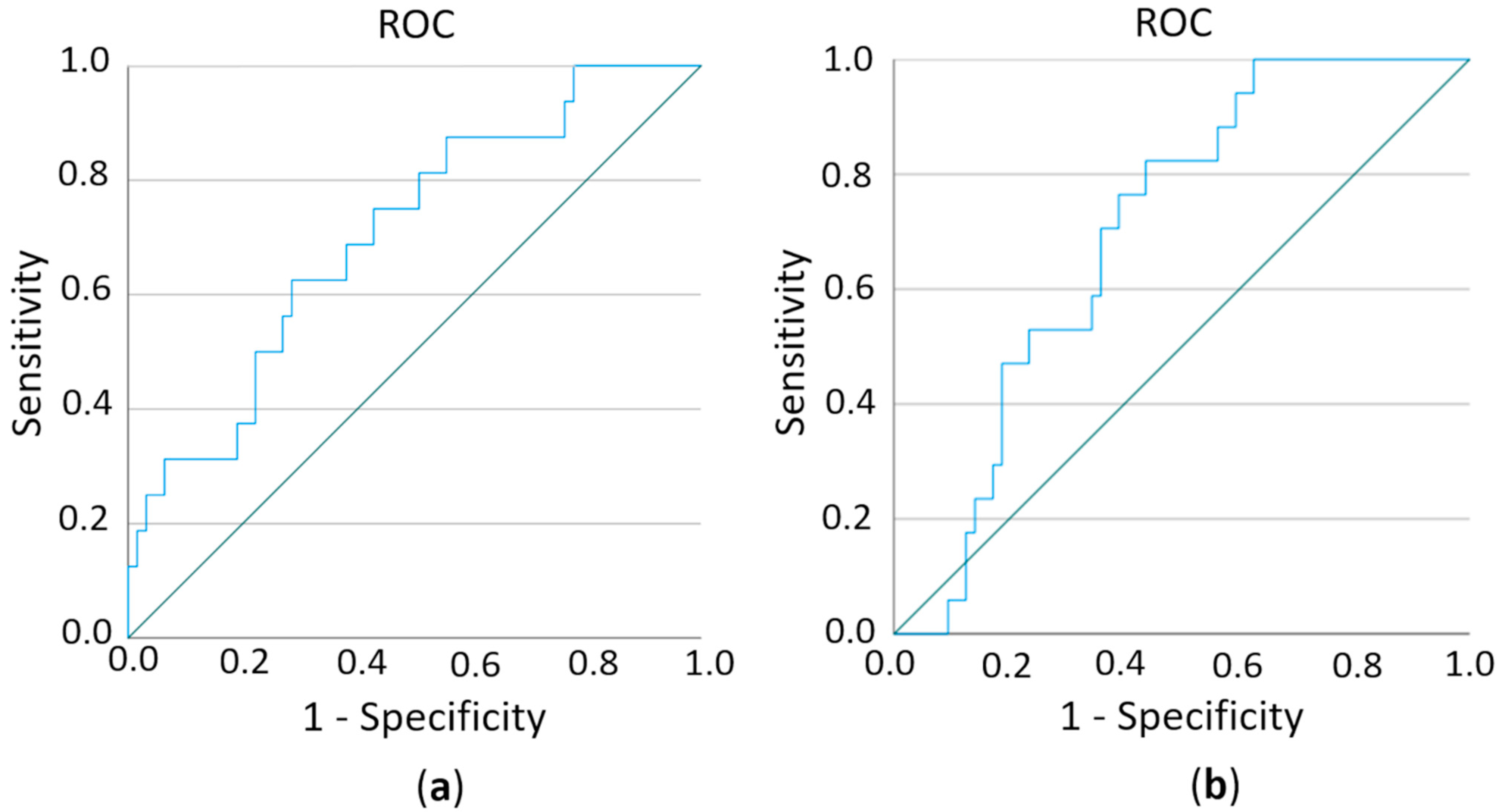
2.6. Estimation of Prognostic Potential of Differentially Expressed miRs
2.7. Correlation between Exosomal miRs and Tumor Characteristics
3. Discussion
4. Materials and Methods
4.1. Patients and Samples
4.2. Exosome Isolation from Patient Serum
4.3. Protein Extraction and Western Blot Analysis
4.4. CD63 Enzyme-Linked Immunosorbent Assay
4.5. RNA Extraction
4.6. cDNA Synthesis and miRNA Expression
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rick, J.W.; Shahin, M.; Chandra, A.; Dalle Ore, C.; Yue, J.K.; Nguyen, A.; Yagnik, G.; Sagar, S.; Arfaie, S.; Aghi, M.K. Systemic therapy for brain metastases. Crit. Rev. Oncol. Hematol. 2019, 142, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Curtaz, C.J.; Schmitt, C.; Blecharz-Lang, K.G.; Roewer, N.; Wockel, A.; Burek, M. Circulating MicroRNAs and Blood-Brain-Barrier Function in Breast Cancer Metastasis. Curr. Pharm. Des. 2020, 26, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Figueira, I.; Godinho-Pereira, J.; Galego, S.; Maia, J.; Hasko, J.; Molnar, K.; Malho, R.; Costa-Silva, B.; Wilhelm, I.; Krizbai, I.A.; et al. MicroRNAs and Extracellular Vesicles as Distinctive Biomarkers of Precocious and Advanced Stages of Breast Cancer Brain Metastases Development. Int. J. Mol. Sci. 2021, 22, 5214. [Google Scholar] [CrossRef]
- Sereno, M.; Videira, M.; Wilhelm, I.; Krizbai, I.A.; Brito, M.A. miRNAs in Health and Disease: A Focus on the Breast Cancer Metastatic Cascade towards the Brain. Cells 2020, 9, 1790. [Google Scholar] [CrossRef]
- Lee, R.; Feinbaum, R.; Ambros, V. A short history of a short RNA. Cell 2004, S116, S89–S92. [Google Scholar] [CrossRef]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Lowery, A.J.; Miller, N.; Devaney, A.; McNeill, R.E.; Davoren, P.A.; Lemetre, C.; Benes, V.; Schmidt, S.; Blake, J.; Ball, G.; et al. MicroRNA signatures predict oestrogen receptor, progesterone receptor and HER2/neu receptor status in breast cancer. Breast Cancer Res. 2009, 11, R27. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, D.; Ni, S.; Peng, Z.; Sheng, W.; Du, X. Plasma microRNAs are promising novel biomarkers for early detection of colorectal cancer. Int. J. Cancer 2010, 127, 118–126. [Google Scholar] [CrossRef]
- Maierthaler, M.; Benner, A.; Hoffmeister, M.; Surowy, H.; Jansen, L.; Knebel, P.; Chang-Claude, J.; Brenner, H.; Burwinkel, B. Plasma miR-122 and miR-200 family are prognostic markers in colorectal cancer. Int. J. Cancer 2017, 140, 176–187. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, C.; Chen, X.; Yang, C.; Li, K.; Wang, J.; Dai, J.; Hu, Z.; Zhou, X.; Chen, L.; et al. Expression profile of microRNAs in serum: A fingerprint for esophageal squamous cell carcinoma. Clin. Chem. 2010, 56, 1871–1879. [Google Scholar] [CrossRef]
- Andras, I.E.; Toborek, M. Extracellular vesicles of the blood-brain barrier. Tissue Barriers 2016, 4, e1131804. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef] [PubMed]
- St-Denis-Bissonnette, F.; Khoury, R.; Mediratta, K.; El-Sahli, S.; Wang, L.; Lavoie, J.R. Applications of Extracellular Vesicles in Triple-Negative Breast Cancer. Cancers 2022, 14, 451. [Google Scholar] [CrossRef]
- Teles, R.H.G.; Yano, R.S.; Villarinho, N.J.; Yamagata, A.S.; Jaeger, R.G.; Meybohm, P.; Burek, M.; Freitas, V.M. Advances in Breast Cancer Management and Extracellular Vesicle Research, a Bibliometric Analysis. Curr. Oncol. 2021, 28, 4504–4520. [Google Scholar] [CrossRef] [PubMed]
- Alexandra, T.; Maria, G.; Charalampos, T.; Eleni, Z.; George, Z.C.; Nikolaos, M.V. Exosomes in breast cancer management: Where do we stand? A literature review. Biol. Cell 2022. [Google Scholar] [CrossRef]
- Wang, M.; Yu, F.; Ding, H.; Wang, Y.; Li, P.; Wang, K. Emerging Function and Clinical Values of Exosomal MicroRNAs in Cancer. Mol. Ther. Nucleic Acids 2019, 16, 791–804. [Google Scholar] [CrossRef]
- Sempere, L.F.; Keto, J.; Fabbri, M. Exosomal MicroRNAs in Breast Cancer towards Diagnostic and Therapeutic Applications. Cancers 2017, 9, 71. [Google Scholar] [CrossRef]
- Nayak, L.; Lee, E.Q.; Wen, P.Y. Epidemiology of brain metastases. Curr. Oncol. Rep. 2012, 14, 48–54. [Google Scholar] [CrossRef]
- Piombino, C.; Mastrolia, I.; Omarini, C.; Candini, O.; Dominici, M.; Piacentini, F.; Toss, A. The Role of Exosomes in Breast Cancer Diagnosis. Biomedicines 2021, 9, 312. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thurlimann, B.; Senn, H.J.; Panel, M. Strategies for subtypes—Dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- Yi, Y.W.; Lee, J.H.; Kim, S.Y.; Pack, C.G.; Ha, D.H.; Park, S.R.; Youn, J.; Cho, B.S. Advances in Analysis of Biodistribution of Exosomes by Molecular Imaging. Int. J. Mol. Sci. 2020, 21, 665. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.; Brown, J.A.; Kerin, M.J. Metastatic breast cancer: The potential of miRNA for diagnosis and treatment monitoring. Cancer Metastasis Rev. 2015, 34, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Conley, A.; Minciacchi, V.R.; Lee, D.H.; Knudsen, B.S.; Karlan, B.Y.; Citrigno, L.; Viglietto, G.; Tewari, M.; Freeman, M.R.; Demichelis, F.; et al. High-throughput sequencing of two populations of extracellular vesicles provides an mRNA signature that can be detected in the circulation of breast cancer patients. RNA Biol. 2017, 14, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Renz, A.; Burek, C.; Mier, W.; Mozoluk, M.; Schulze-Osthoff, K.; Los, M. Cytochrome c is rapidly extruded from apoptotic cells and detectable in serum of anticancer-drug treated tumor patients. Adv. Exp. Med. Biol. 2001, 495, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Malla, B.; Aebersold, D.M.; Dal Pra, A. Protocol for serum exosomal miRNAs analysis in prostate cancer patients treated with radiotherapy. J. Transl. Med. 2018, 16, 223. [Google Scholar] [CrossRef]
- Chen, S.D.; Pan, H.Y.; Huang, J.B.; Liu, X.P.; Li, J.H.; Ho, C.J.; Tsai, M.H.; Yang, J.L.; Chen, S.F.; Chen, N.C.; et al. Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia. Cells 2020, 9, 1867. [Google Scholar] [CrossRef]
- Wang, L.; Cao, D.; Wang, L.; Zhao, J.; Nguyen, L.N.; Dang, X.; Ji, Y.; Wu, X.Y.; Morrison, Z.D.; Xie, Q.; et al. HCV-associated exosomes promote myeloid-derived suppressor cell expansion via inhibiting miR-124 to regulate T follicular cell differentiation and function. Cell Discov. 2018, 4, 51. [Google Scholar] [CrossRef]
- Palviainen, M.; Saraswat, M.; Varga, Z.; Kitka, D.; Neuvonen, M.; Puhka, M.; Joenvaara, S.; Renkonen, R.; Nieuwland, R.; Takatalo, M.; et al. Extracellular vesicles from human plasma and serum are carriers of extravesicular cargo-Implications for biomarker discovery. PLoS ONE 2020, 15, e0236439. [Google Scholar] [CrossRef]
- Porichis, F.; Hart, M.G.; Griesbeck, M.; Everett, H.L.; Hassan, M.; Baxter, A.E.; Lindqvist, M.; Miller, S.M.; Soghoian, D.Z.; Kavanagh, D.G.; et al. High-throughput detection of miRNAs and gene-specific mRNA at the single-cell level by flow cytometry. Nat. Commun. 2014, 5, 5641. [Google Scholar] [CrossRef]
- Precazzini, F.; Detassis, S.; Imperatori, A.S.; Denti, M.A.; Campomenosi, P. Measurements Methods for the Development of MicroRNA-Based Tests for Cancer Diagnosis. Int. J. Mol. Sci. 2021, 22, 1176. [Google Scholar] [CrossRef]
- Zuo, Y.; Zheng, W.; Tang, Q.; Liu, J.; Wang, S.; Xin, C. miR5763p overexpression enhances cisplatin sensitivity of ovarian cancer cells by dysregulating PDL1 and cyclin D1. Mol. Med. Rep. 2021, 23, 81. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Zhang, J.; Bu, X.; Wang, H.; Zhu, Y.; Geng, Y.; Nihira, N.T.; Tan, Y.; Ci, Y.; Wu, F.; Dai, X.; et al. Cyclin D-CDK4 kinase destabilizes PD-L1 via cullin 3-SPOP to control cancer immune surveillance. Nature 2018, 553, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Balci, S.; Ayaz, L.; Gorur, A.; Yildirim Yaroglu, H.; Akbayir, S.; Dogruer Unal, N.; Bulut, B.; Tursen, U.; Tamer, L. microRNA profiling for early detection of nonmelanoma skin cancer. Clin. Exp. Dermatol. 2016, 41, 346–351. [Google Scholar] [CrossRef]
- Greenawalt, E.J.; Edmonds, M.D.; Jain, N.; Adams, C.M.; Mitra, R.; Eischen, C.M. Targeting of SGK1 by miR-576-3p Inhibits Lung Adenocarcinoma Migration and Invasion. Mol. Cancer Res. 2019, 17, 289–298. [Google Scholar] [CrossRef]
- Meng, F.M.; Meng, F.M.; Song, X.L. MiR-576-3p is a novel marker correlated with poor clinical outcome in bladder cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 973–977. [Google Scholar]
- Liang, Z.; Li, S.; Xu, X.; Xu, X.; Wang, X.; Wu, J.; Zhu, Y.; Hu, Z.; Lin, Y.; Mao, Y.; et al. MicroRNA-576-3p inhibits proliferation in bladder cancer cells by targeting cyclin D1. Mol. Cells 2015, 38, 130–137. [Google Scholar] [CrossRef]
- Port, M.; Glaesener, S.; Ruf, C.; Riecke, A.; Bokemeyer, C.; Meineke, V.; Honecker, F.; Abend, M. Micro-RNA expression in cisplatin resistant germ cell tumor cell lines. Mol. Cancer 2011, 10, 52. [Google Scholar] [CrossRef]
- Lv, J.; Xia, K.; Xu, P.; Sun, E.; Ma, J.; Gao, S.; Zhou, Q.; Zhang, M.; Wang, F.; Chen, F.; et al. miRNA expression patterns in chemoresistant breast cancer tissues. Biomed. Pharm. 2014, 68, 935–942. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, J.; Li, J.; Shao, J.; Fang, L. MiR-130a-3p inhibits migration and invasion by regulating RAB5B in human breast cancer stem cell-like cells. Biochem. Biophys. Res. Commun. 2018, 501, 486–493. [Google Scholar] [CrossRef]
- Wilson, D.B.; Wilson, M.P. Identification and subcellular localization of human rab5b, a new member of the ras-related superfamily of GTPases. J. Clin. Investig. 1992, 89, 996–1005. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gopal Krishnan, P.D.; Golden, E.; Woodward, E.A.; Pavlos, N.J.; Blancafort, P. Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer. Cancers 2020, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, R.; Zhang, F.; Chen, Y.; Lv, Q.; Long, G.; Yang, K. MicroRNA-130a inhibits cell proliferation, invasion and migration in human breast cancer by targeting the RAB5A. Int. J. Clin. Exp. Pathol. 2015, 8, 384–393. [Google Scholar] [PubMed]
- Liu, L.; Wang, P.; Wang, Y.S.; Zhang, Y.N.; Li, C.; Yang, Z.Y.; Liu, Z.H.; Zhan, T.Z.; Xu, J.; Xia, C.M. MiR-130a-3p Alleviates Liver Fibrosis by Suppressing HSCs Activation and Skewing Macrophage to Ly6C(lo) Phenotype. Front. Immunol. 2021, 12, 696069. [Google Scholar] [CrossRef]
- Kjersem, J.B.; Ikdahl, T.; Lingjaerde, O.C.; Guren, T.; Tveit, K.M.; Kure, E.H. Plasma microRNAs predicting clinical outcome in metastatic colorectal cancer patients receiving first-line oxaliplatin-based treatment. Mol. Oncol. 2014, 8, 59–67. [Google Scholar] [CrossRef]
- Su, S.; Zhao, Q.; He, C.; Huang, D.; Liu, J.; Chen, F.; Chen, J.; Liao, J.Y.; Cui, X.; Zeng, Y.; et al. miR-142-5p and miR-130a-3p are regulated by IL-4 and IL-13 and control profibrogenic macrophage program. Nat. Commun. 2015, 6, 8523. [Google Scholar] [CrossRef]
- Lam, S.; Lodder, K.; Teunisse, A.F.; Rabelink, M.J.; Schutte, M.; Jochemsen, A.G. Role of Mdm4 in drug sensitivity of breast cancer cells. Oncogene 2010, 29, 2415–2426. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, H.; Lin, Z.; Zhang, G. Functional studies of miR-130a on the inhibitory pathways of apoptosis in patients with chronic myeloid leukemia. Cancer Gene Ther. 2015, 22, 573–580. [Google Scholar] [CrossRef]
- Rivlin, N.; Brosh, R.; Oren, M.; Rotter, V. Mutations in the p53 Tumor Suppressor Gene: Important Milestones at the Various Steps of Tumorigenesis. Genes Cancer 2011, 2, 466–474. [Google Scholar] [CrossRef]
- Hadjipanteli, A.; Doolan, P.; Kyriacou, E.; Constantinidou, A. Breast Cancer Brain Metastasis: The Potential Role of MRI Beyond Current Clinical Applications. Cancer Manag. Res. 2020, 12, 9953–9964. [Google Scholar] [CrossRef]
- Martin, A.M.; Cagney, D.N.; Catalano, P.J.; Warren, L.E.; Bellon, J.R.; Punglia, R.S.; Claus, E.B.; Lee, E.Q.; Wen, P.Y.; Haas-Kogan, D.A.; et al. Brain Metastases in Newly Diagnosed Breast Cancer: A Population-Based Study. JAMA Oncol. 2017, 3, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Adhami, M.; Haghdoost, A.A.; Sadeghi, B.; Malekpour Afshar, R. Candidate miRNAs in human breast cancer biomarkers: A systematic review. Breast Cancer 2018, 25, 198–205. [Google Scholar] [CrossRef]
- Gonzalez-Villasana, V.; Rashed, M.H.; Gonzalez-Cantu, Y.; Bayraktar, R.; Menchaca-Arredondo, J.L.; Vazquez-Guillen, J.M.; Rodriguez-Padilla, C.; Lopez-Berestein, G.; Resendez-Perez, D. Presence of Circulating miR-145, miR-155, and miR-382 in Exosomes Isolated from Serum of Breast Cancer Patients and Healthy Donors. Dis. Markers 2019, 2019, 6852917. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Mao, J.H.; Wang, B.Y.; Wang, L.X.; Wen, H.Y.; Xu, L.J.; Fu, J.X.; Yang, H. Exosomal miR-1910-3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-kappaB signaling pathway. Cancer Lett. 2020, 489, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Curtaz, C.J.; Schmitt, C.; Herbert, S.L.; Feldheim, J.; Schlegel, N.; Gosselet, F.; Hagemann, C.; Roewer, N.; Meybohm, P.; Wockel, A.; et al. Serum-derived factors of breast cancer patients with brain metastases alter permeability of a human blood-brain barrier model. Fluids Barriers CNS 2020, 17, 31. [Google Scholar] [CrossRef]
- Burek, M.; Konig, A.; Lang, M.; Fiedler, J.; Oerter, S.; Roewer, N.; Bohnert, M.; Thal, S.C.; Blecharz-Lang, K.G.; Woitzik, J.; et al. Hypoxia-Induced MicroRNA-212/132 Alter Blood-Brain Barrier Integrity Through Inhibition of Tight Junction-Associated Proteins in Human and Mouse Brain Microvascular Endothelial Cells. Transl. Stroke Res. 2019, 10, 672–683. [Google Scholar] [CrossRef]
- Kaiser, M.; Burek, M.; Britz, S.; Lankamp, F.; Ketelhut, S.; Kemper, B.; Forster, C.; Gorzelanny, C.; Goycoolea, F.M. The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers. Int. J. Mol. Sci. 2018, 20, 122. [Google Scholar] [CrossRef]
- Salvador, E.; Burek, M.; Lohr, M.; Nagai, M.; Hagemann, C.; Forster, C.Y. Senescence and associated blood-brain barrier alterations in vitro. Histochem. Cell Biol. 2021, 156, 283–292. [Google Scholar] [CrossRef]
- Burek, M.; Burmester, S.; Salvador, E.; Moller-Ehrlich, K.; Schneider, R.; Roewer, N.; Nagai, M.; Forster, C.Y. Kidney Ischemia/Reperfusion Injury Induces Changes in the Drug Transporter Expression at the Blood-Brain Barrier in vivo and in vitro. Front. Physiol. 2020, 11, 569881. [Google Scholar] [CrossRef]
- Blecharz-Lang, K.G.; Prinz, V.; Burek, M.; Frey, D.; Schenkel, T.; Krug, S.M.; Fromm, M.; Vajkoczy, P. Gelatinolytic activity of autocrine matrix metalloproteinase-9 leads to endothelial de-arrangement in Moyamoya disease. J. Cereb. Blood Flow Metab. 2018, 38, 1940–1953. [Google Scholar] [CrossRef]
- Gabbert, L.; Dilling, C.; Meybohm, P.; Burek, M. Deletion of Protocadherin Gamma C3 Induces Phenotypic and Functional Changes in Brain Microvascular Endothelial Cells In Vitro. Front. Pharmacol. 2020, 11, 590144. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | PC | CM | VM | BM | |
|---|---|---|---|---|---|
| Patients characteristics | |||||
| Total number | 18 | 15 | 16 | 18 | 16 |
| Median age | 60.3 | 61.6 | 62.9 | 61.1 | 61.8 |
| Deceased | 12 | 9 | 2 | ||
| Pre-/postmenopausal | 4/14 | 4/11 | 2/14 | 3/15 | 2/14 |
| Tumor characteristics | |||||
| ER/PR-positive | 8 | 4 | 8 | 10 | |
| HER2/neu-positive | 5 | 8 | 9 | 5 | |
| Triple-negative | 2 | 4 | 1 | 1 | |
| Grading | |||||
| Well differentiated (G1) | 1 | ||||
| Moderately differentiated (G2) | 10 | 9 | 9 | 11 | |
| Poorly differentiated (G3) | 5 | 7 | 8 | 5 | |
| % of Ki67-positive cells (median) | 22% | 43% | 34.7% | 33.9% | |
| Other | 1 |
| Correlates With: | Grading | % of Ki67-Positive Cells |
|---|---|---|
| % of Ki67-positive cells | r 0.513 ** p < 0.001 | |
| hsa-miR-132-3p | r −0.300 * p 0.043 | |
| hsa-miR-148a-3p | r −0.334 * p 0.022 | |
| hsa-miR-150-5p | r −0.343 * p 0.019 | |
| hsa-miR-197-3p | r −0.368 * p 0.012 | |
| hsa-miR-199a-3p | r −0.339 * p 0.021 | |
| hsa-miR-27a-3p | r −0.295 * p 0.044 | |
| hsa-miR-340-5p | r 0.334 * p 0.023 | |
| hsa-miR-342-3p | r −0.435 ** p 0.003 | |
| hsa-miR-425-5p | r −0.337 * p 0.021 | |
| hsa-miR-576-3p | r −0.328 * p 0.030 | |
| hsa-miR-885-5p | r −0.312 * p 0.042 | |
| hsa-miR-92b-3p | r −0.297 * p 0.043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtaz, C.J.; Reifschläger, L.; Strähle, L.; Feldheim, J.; Feldheim, J.J.; Schmitt, C.; Kiesel, M.; Herbert, S.-L.; Wöckel, A.; Meybohm, P.; et al. Analysis of microRNAs in Exosomes of Breast Cancer Patients in Search of Molecular Prognostic Factors in Brain Metastases. Int. J. Mol. Sci. 2022, 23, 3683. https://doi.org/10.3390/ijms23073683
Curtaz CJ, Reifschläger L, Strähle L, Feldheim J, Feldheim JJ, Schmitt C, Kiesel M, Herbert S-L, Wöckel A, Meybohm P, et al. Analysis of microRNAs in Exosomes of Breast Cancer Patients in Search of Molecular Prognostic Factors in Brain Metastases. International Journal of Molecular Sciences. 2022; 23(7):3683. https://doi.org/10.3390/ijms23073683
Chicago/Turabian StyleCurtaz, Carolin J., Leonie Reifschläger, Linus Strähle, Jonas Feldheim, Julia J. Feldheim, Constanze Schmitt, Matthias Kiesel, Saskia-Laureen Herbert, Achim Wöckel, Patrick Meybohm, and et al. 2022. "Analysis of microRNAs in Exosomes of Breast Cancer Patients in Search of Molecular Prognostic Factors in Brain Metastases" International Journal of Molecular Sciences 23, no. 7: 3683. https://doi.org/10.3390/ijms23073683
APA StyleCurtaz, C. J., Reifschläger, L., Strähle, L., Feldheim, J., Feldheim, J. J., Schmitt, C., Kiesel, M., Herbert, S.-L., Wöckel, A., Meybohm, P., & Burek, M. (2022). Analysis of microRNAs in Exosomes of Breast Cancer Patients in Search of Molecular Prognostic Factors in Brain Metastases. International Journal of Molecular Sciences, 23(7), 3683. https://doi.org/10.3390/ijms23073683