
Differential Response of Wheat Rhizosphere Bacterial Community to Plant Variety and Fertilization



 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
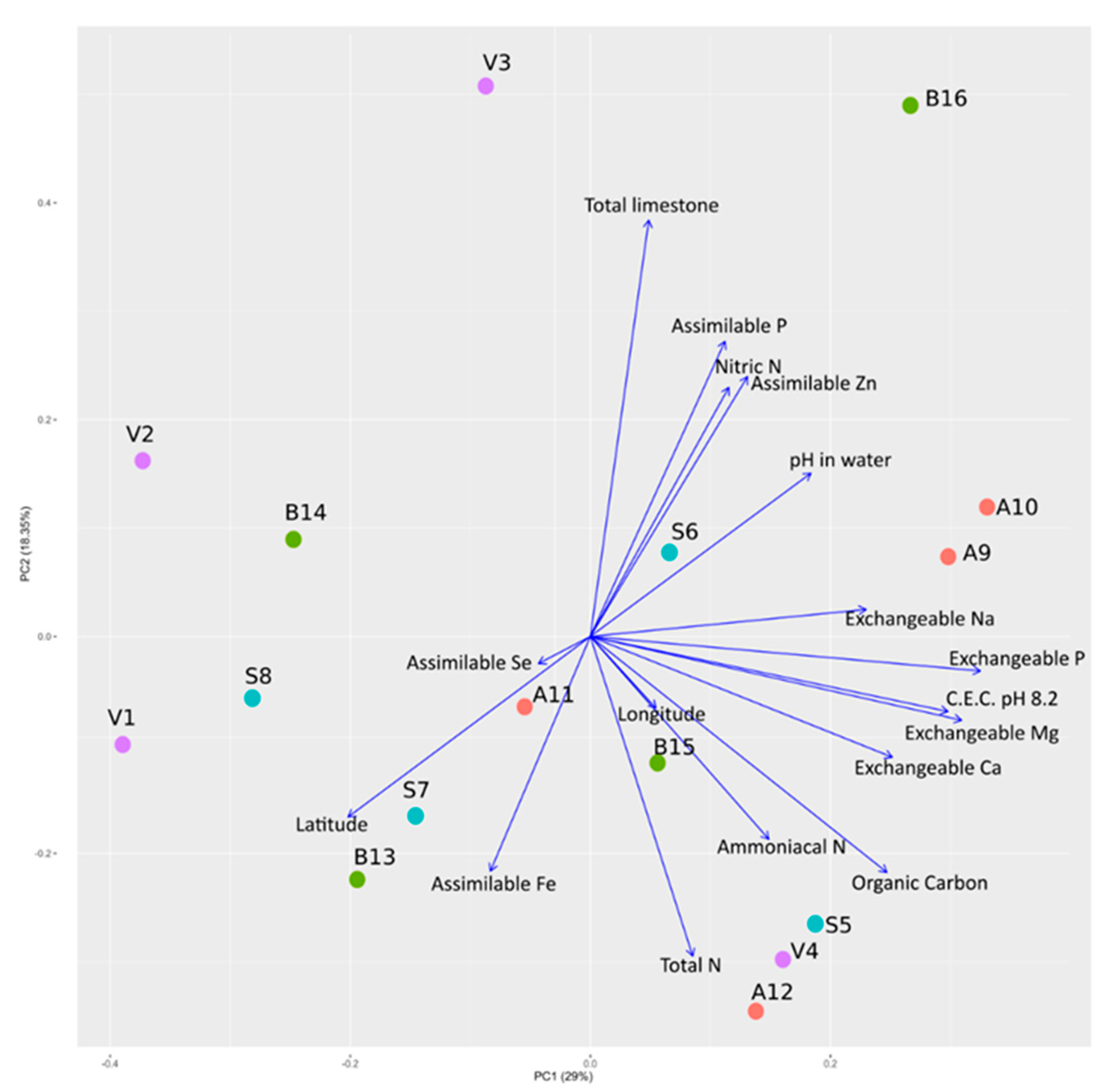
2.1. Physicochemical Characterization of the Soil
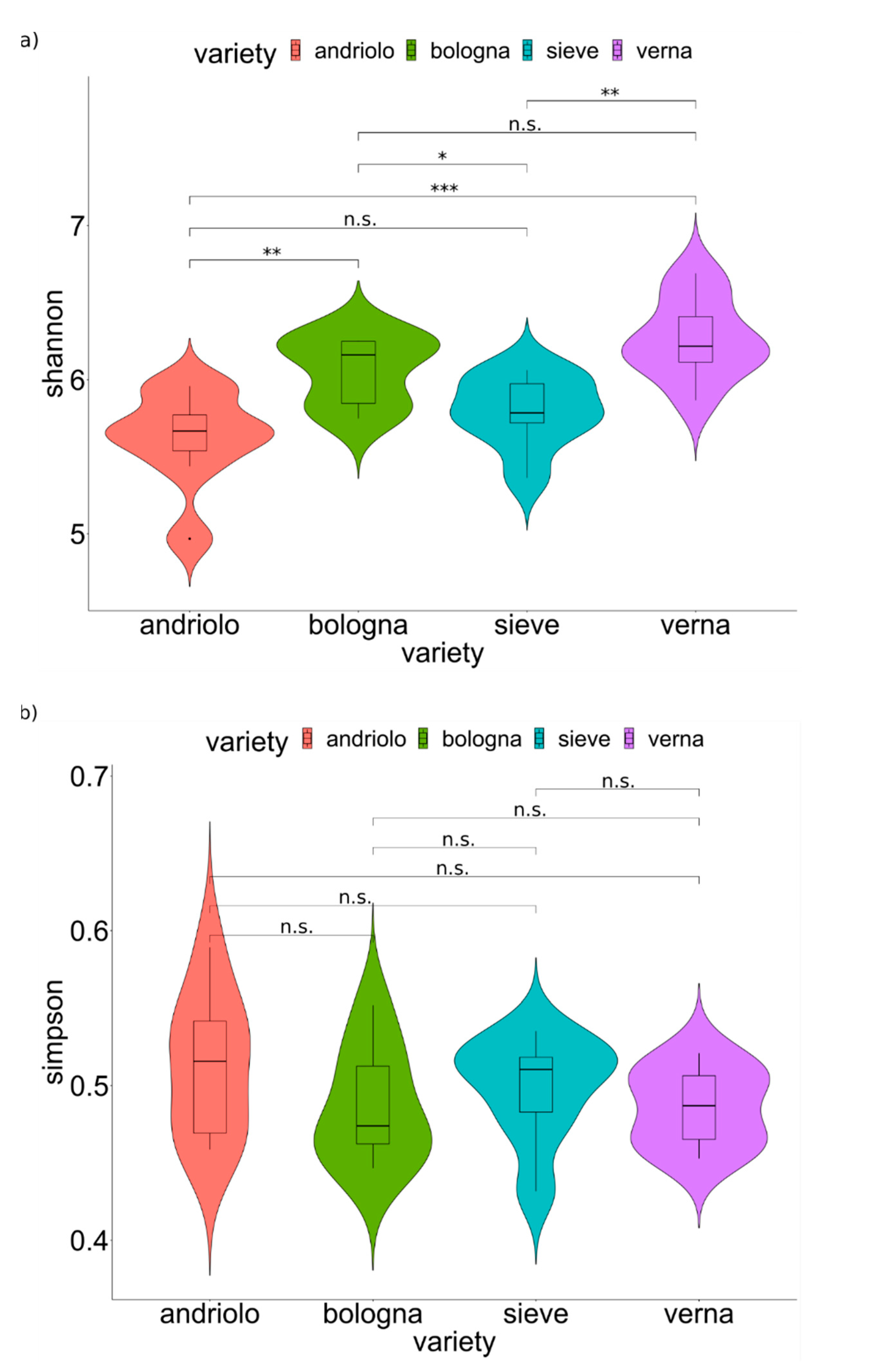
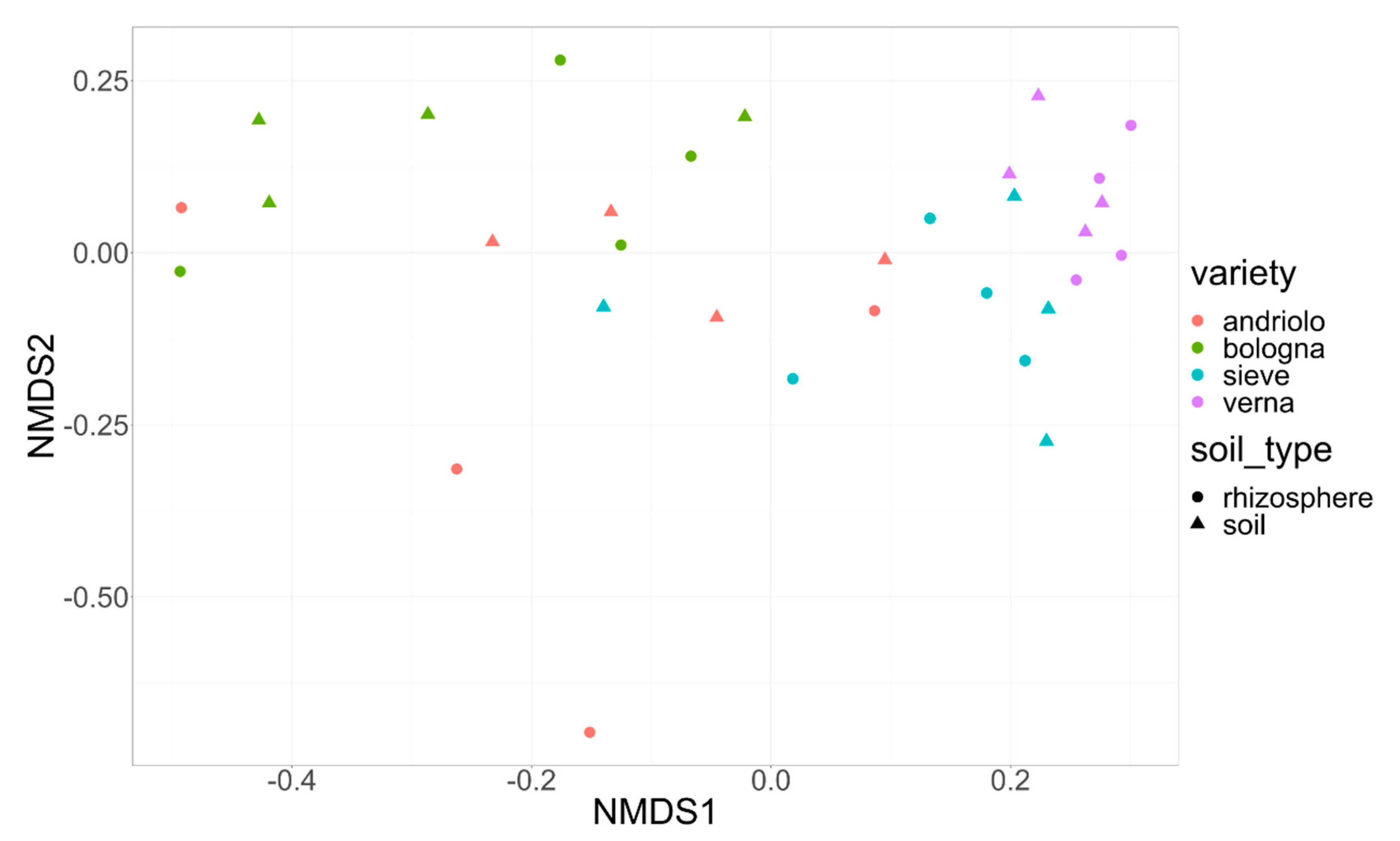
2.2. Diversity of the Soil and Rhizosphere Bacterial Community
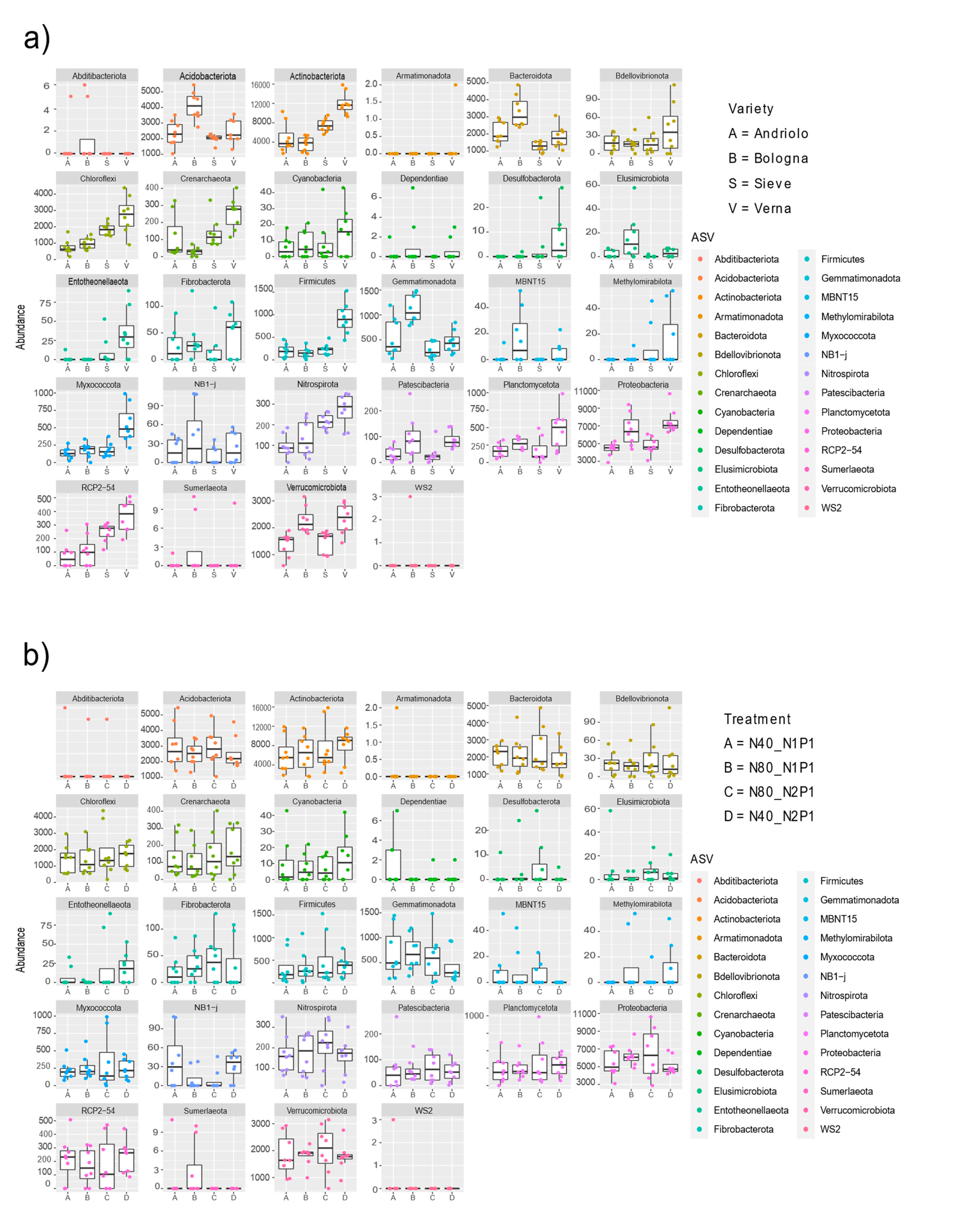
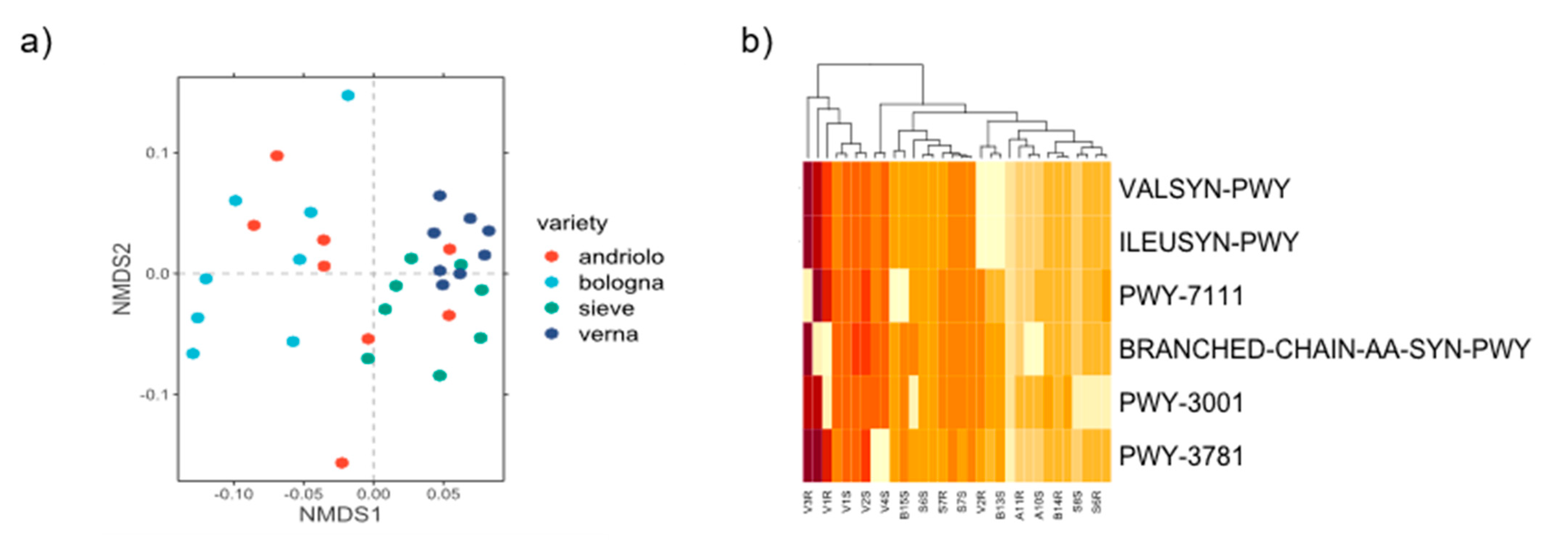
2.3. Variety Affects the Taxonomic Composition of Wheat Rhizobacterial Community
2.4. Prediction of Potential Functions and Metabolic Pathways
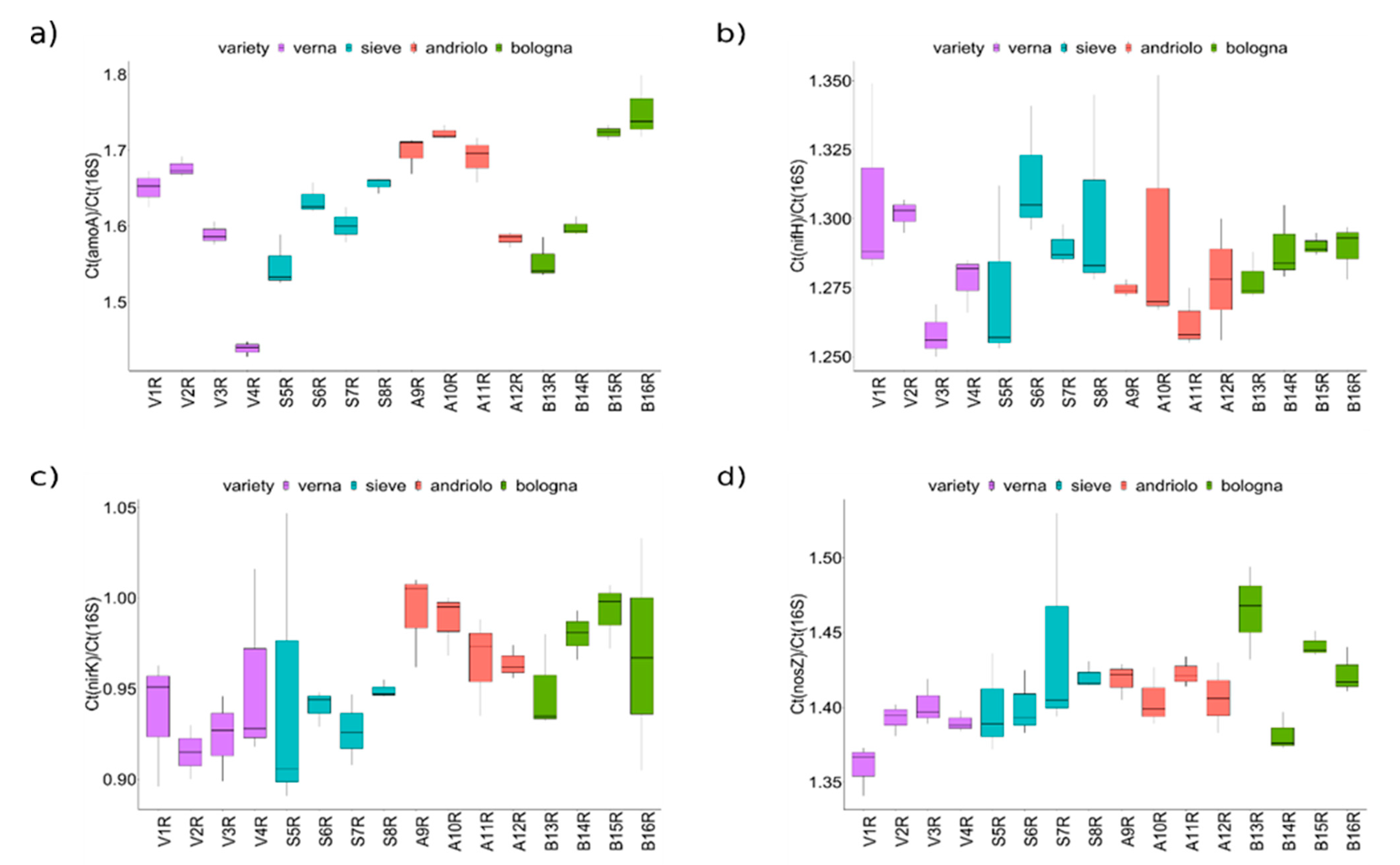
2.5. Quantitative PCR Analysis of the Nitrogen Cycle Genes
3. Discussion
4. Materials and Methods
4.1. Experimental Field and Plant Genotypes
4.2. Sampling and Samples Treatment
4.3. Soil Chemical and Physical Analyses
4.4. eDNA Extraction and 16S rRNA Gene Amplicon Sequencing
4.5. Quantitative PCR of Genes Involved in the Nitrogen Cycle
4.6. Bioinformatic and Statistical Analyses
4.7. Prediction of Functional Abundances
4.8. Links to Deposited Data
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaeppi, K.; Bulgarelli, D. The Plant Microbiome at Work. Mol. Plant-Microbe Interact. MPMI 2015, 212, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cañizares, C.; Jorrín, B.; Poole, P.S.; Tkacz, A. Understanding the holobiont: The interdependence of plants and their microbiome. Curr. Opin. Microbiol. 2017, 38, 188–196. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Romero, E.; Aguirre-Noyola, J.L.; Taco-Taype, N.; Martínez-Romero, J.; Zuñiga-Dávila, D. Plant microbiota modified by plant domestication. Syst. Appl. Microbiol. 2020, 43, 126106. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Escudero-Martinez, C.; Bulgarelli, D. Tracing the evolutionary routes of plant–microbiota interactions. Curr. Opin. Microbiol. 2019, 49, 34–40. [Google Scholar] [CrossRef]
- Checcucci, A.; Maida, I.; Bacci, G.; Ninno, C.; Bilia, A.R.; Biffi, S.; Firenzuoli, F.; Flamini, G.; Fani, R.; Mengoni, A. Is the plant-associated microbiota of Thymus spp. adapted to plant essential oil? Res. Microbiol. 2017, 168, 276–282. [Google Scholar] [CrossRef]
- Kristin, A.; Miranda, H. The root microbiota—A fingerprint in the soil? Plant Soil 2013, 671–686. [Google Scholar] [CrossRef]
- Fagorzi, C.; Bacci, G.; Huang, R.; Cangioli, L.; Checcucci, A.; Fini, M.; Perrin, E.; Natali, C.; diCenzo, G.C.; Mengoni, A. Nonadditive transcriptomic signatures of genotype-by-genotype interactions during the initiation of plant-rhizobium symbiosis. mSystems 2021, 6, 00974-20. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Stopnisek, N.; Shade, A. Prioritizing persistent microbiome members in the common bean rhizosphere: An integrated analysis of space, time, and plant genotype. ISME J. 2021, 15, 2708–2722. [Google Scholar] [CrossRef] [PubMed]
- Poole, N.; Donovan, J.; Erenstein, O. Viewpoint: Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef]
- Fróna, D.; Szenderák, J.; Harangi-Rákos, M. The Challenge of Feeding the World. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef] [Green Version]
- OECD-FAO. OECD-FAO Agricultural Outlook 2017-2026; OECD-FAO Agricultural Outlook; OECD: Paris, France, 2017. [Google Scholar]
- Herrera, J.M.; Levy Häner, L.; Mascher, F.; Hiltbrunner, J.; Fossati, D.; Brabant, C.; Charles, R.; Pellet, D. Lessons from 20 Years of Studies of Wheat Genotypes in Multiple Environments and under Contrasting Production Systems. Front. Plant Sci. 2020, 10, 1745. [Google Scholar] [CrossRef]
- Guerrini, L.; Napoli, M.; Mancini, M.; Masella, P.; Cappelli, A.; Parenti, A.; Orlandini, S. Wheat grain composition, dough rheology and bread quality as affected by nitrogen and sulfur fertilization and seeding density. Agronomy 2020, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Kavamura, V.N.; Robinson, R.J.; Hughes, D.; Clark, I.; Rossmann, M.; Soares de Melo, I.; Hirsch, P.R.; Mendes, R.; Mauchline, T.H. Wheat Dwarfing Influences Selection of the Rhizosphere Microbiome. Sci. Rep. 2020, 10, 1452. [Google Scholar] [CrossRef]
- Ellis, M.; Rebetzke, G.; Chandler, P.; Bonnett, D.; Spielmeyer, W.; Richards, R. The effect of different height reducing genes on the early growth of wheat. Funct. Plant Biol. 2004, 31, 583–589. [Google Scholar] [CrossRef]
- Arshad, M.A. A Review on Wheat Management, Strategies, Current Problems and Future Perspectives. Haya Saudi J. Life Sci. 2021, 6, 14–18. [Google Scholar] [CrossRef]
- Sanchez-Garcia, M.; Álvaro, F.; Peremarti, A.; Martín-Sánchez, J.A.; Royo, C. Changes in bread-making quality attributes of bread wheat varieties cultivated in Spain during the 20th century. Eur. J. Agron. 2015, 63, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Migliorini, P.; Spagnolo, S.; Torri, L.; Arnoulet, M.; Lazzerini, G.; Ceccarelli, S. Agronomic and quality characteristics of old, modern and mixture wheat varieties and landraces for organic bread chain in diverse environments of northern Italy. Eur. J. Agron. 2016, 79, 131–141. [Google Scholar] [CrossRef]
- Fan, M.S.; Zhao, F.J.; Fairweather-Tait, S.J.; Poulton, P.R.; Dunham, S.J.; McGrath, S.P. Evidence of decreasing mineral density in wheat grain over the last 160 years. J. Trace Elem. Med. Biol. 2008, 22, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Arzani, A.; Ashraf, M. Cultivated Ancient Wheats (Triticum spp.): A Potential Source of Health-Beneficial Food Products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Ghiselli, L.; Rossi, E.; Whittaker, A.; Dinelli, G.; Baglio, A.P.; Andrenelli, L.; Benedettelli, S. Nutritional characteristics of ancient Tuscan varieties of Triticum aestivum L. Ital. J. Agron. 2016, 11, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Donn, S.; Kirkegaard, J.A.; Perera, G.; Richardson, A.E.; Watt, M. Evolution of bacterial communities in the wheat crop rhizosphere. Environ. Microbiol. 2015, 17, 610–621. [Google Scholar] [CrossRef]
- Wolinska, A.; Kuzniar, A.; Galazka, A. Biodiversity in the Rhizosphere of selected winter wheat (Triticum aestivum L.) cultivars-genetic and catabolic fingerprinting. Agronomy 2020, 10, 953. [Google Scholar] [CrossRef]
- Simonin, M.; Dasilva, C.; Terzi, V.; Ngonkeu, E.L.M.; DIouf, D.; Kane, A.; Béna, G.; Moulin, L. Influence of plant genotype and soil on the wheat rhizosphere microbiome: Evidences for a core microbiome across eight African and European soils. FEMS Microbiol. Ecol. 2020, 96, fiaa067. [Google Scholar] [CrossRef]
- Chen, S.; Waghmode, T.R.; Sun, R.; Kuramae, E.E.; Hu, C.; Liu, B. Root-associated microbiomes of wheat under the combined effect of plant development and nitrogen fertilization. Microbiome 2019, 7, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkacz, A.; Pini, F.; Turner, T.R.; Bestion, E.; Simmonds, J.; Howell, P.; Greenland, A.; Cheema, J.; Emms, D.M.; Uauy, C.; et al. Agricultural Selection of Wheat Has Been Shaped by Plant-Microbe Interactions. Front. Microbiol. 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Spor, A.; Roucou, A.; Mounier, A.; Bru, D.; Breuil, M.C.; Fort, F.; Vile, D.; Roumet, P.; Philippot, L.; Violle, C. Domestication-driven changes in plant traits associated with changes in the assembly of the rhizosphere microbiota in tetraploid wheat. Sci. Rep. 2020, 10, 12234. [Google Scholar] [CrossRef]
- Germida, J.; Siciliano, S. Taxonomic diversity of bacteria associated with the roots of modern, recent and ancient wheat cultivars. Biol. Fertil. Soils 2001, 33, 410–415. [Google Scholar] [CrossRef]
- Evenson, R.E.; Gollin, D. Assessing the impact of the Green Revolution, 1960 to 2000. Science 2003, 300, 758–762. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Wu, Y.; Zhu, M.; Yu, W.; Yao, H.; Zhu, Y.G.; Chu, H. Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome 2019, 7, 1–10. [Google Scholar] [CrossRef]
- Iannucci, A.; Canfora, L.; Nigro, F.; De Vita, P.; Beleggia, R. Relationships between root morphology, root exudate compounds and rhizosphere microbial community in durum wheat. Appl. Soil Ecol. 2021, 158, 103781. [Google Scholar] [CrossRef]
- Mamine, F.; Farès, M.H. Barriers and levers to developing wheat–pea intercropping in Europe: A review. Sustainability 2020, 12, 6962. [Google Scholar] [CrossRef]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2008, 285, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Caddell, D.F.; Xu, G.; Dahlen, L.; Washington, L.; Yang, J.; Coleman-Derr, D. Genome wide association study reveals plant loci controlling heritability of the rhizosphere microbiome. ISME J. 2021, 15, 3181–3194. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Zeeshan, M.; Farooq, S.; Ali, I.; Khan, A.; Zhou, X.B. Irrigation and nitrogen fertilization alter soil bacterial communities, soil enzyme activities, and nutrient availability in maize crop. Front. Microbiol. 2022, 105, 833758. [Google Scholar] [CrossRef] [PubMed]
- Ossowicki, A.; Tracanna, V.; Petrus, M.L.C.; van Wezel, G.; Raaijmakers, J.M.; Medema, M.H.; Garbeva, P. Microbial and volatile profiling of soils suppressive to Fusarium culmorum of wheat. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Shen, J.P.; Di, H.J.; Zhang, L.M.; Zhang, C.; He, J.Z. Variation of soil nitrate and bacterial diversity along soil profiles in manure disposal maize field and adjacent woodland. J. Soils Sediments 2020, 20, 3557–3568. [Google Scholar] [CrossRef]
- Wang, X.; Wang, S.; Jiang, Y.; Zhou, J.; Han, C.; Zhu, G. Comammox bacterial abundance, activity, and contribution in agricultural rhizosphere soils. Sci. Total Environ. 2020, 727, 138563. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmi, K.D.; Kuhr, S.L.; Johnson, V.A.; Mattern, P.J.; Schmidt, J.W. Results of the Seventh International Winter Wheat Performance Nursery Grown in 1975; ral Resources, Agricultural Experiment Station: Lincoln, NB, USA, 1977. [Google Scholar]
- Verrillo, F.; Badeck, F.-W.; Terzi, V.; Rizza, F.; Bernardo, L.; Di Maro, A.; Fares, C.; Zaldei, A.; Miglietta, F.; Moschella, A.; et al. Elevated field atmospheric CO2 concentrations affect the characteristics of winter wheat (cv. Bologna) grains. Crop Pasture Sci. 2017, 68, 713–725. [Google Scholar] [CrossRef]
- Jimenez, R.R.; Ladha, J.K. Automated elemental analysis: A rapid and reliable but expensive measurement of total carbon and nitrogen in plant and soil samples. Commun. Soil Sci. Plant Anal. 2008, 24, 1897–1924. [Google Scholar] [CrossRef]
- Matejovic, I. Determination of carbon and nitrogen in samples of various soils by the dry combustion. Commun. Soil Sci. Plant Anal. 1997, 28, 1499–1511. [Google Scholar] [CrossRef]
- Vitti, C.; Stellacci, A.M.; Leogrande, R.; Mastrangelo, M.; Cazzato, E.; Ventrella, D. Assessment of organic carbon in soils: A comparison between the Springer–Klee wet digestion and the dry combustion methods in Mediterranean soils (Southern Italy). CATENA 2016, 137, 113–119. [Google Scholar] [CrossRef]
- Verardo, D.J.; Froelich, P.N.; McIntyre, A. Determination of organic carbon and nitrogen in marine sediments using the Carlo Erba NA-1500 analyzer. Deep Sea Res. Part A. Oceanogr. Res. Pap. 1990, 37, 157–165. [Google Scholar] [CrossRef]
- Houba, V.J.G.; Huijbregts, A.W.M.; Wilting, P.; Novozamsky, I.; Gort, G. Sugar yield, nitrogen uptake by sugar beet and optimal nitrogen fertilization in relation to nitrogen soil analyses and several additional factors. Biol. Fertil. Soils 1995, 19, 55–59. [Google Scholar] [CrossRef]
- APHA; APHA Standard Methods, Method 4500-NO3-E—Cadmium Reduction Method. 22nd ed. America Public Health Association: Washington, DC, USA, 2000.
- EPA EPA Method 353.2; Determination of Nitrate-Nitrite Nitrogen by Automated Colorimetry. Revision 2.0. August 1993. United States Environmental Protection Agency: Cincinnati, OH, USA, 19 August 1993.
- ASTM D1426-15; Standard Test Methods for Ammonia Nitrogen in Water. ASTM International: West Conshohocken, PA, USA, 2015.
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part 2 Chemical and Microbiological Properties; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Mehlich, A. Use of Triethanolamine Acetate-Barium Hydroxide Buffer for the Determination of Some Base Exchange Properties and Lime Requirement of Soil. Soil Sci. Soc. Am. J. 1939, 3, 162–166. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Bacci, G.; Chiellini, C.; Fagorzi, C.; Niccolai, E.; Taddei, A.; Ricci, F.; Ringressi, M.N.; Borrelli, R.; Melli, F.; et al. Preliminary comparison of oral and intestinal human microbiota in patients with colorectal cancer: A pilot study. Front. Microbiol. 2018, 8, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelrhman, K.F.A.; Bacci, G.; Nistri, A.; Mengoni, A.; Ugolini, A. Diet and gut microbiota of two supralittoral amphipods Orchestia montagui and Talitrus saltator living in different microhabitats. Estuar. Coast. Shelf Sci. 2017, 197, 119–125. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wright, E.S.; Yilmaz, L.S.; Noguera, D.R. DECIPHER, a search-based approach to chimera identification for 16S rRNA sequences. Appl. Environ. Microbiol. 2012, 78, 717–725. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.A.; Lahti, L. Microbiome data science. J. Biosci. 2019, 44, 115. [Google Scholar] [CrossRef]
- Wickham Hadley ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6.
- Kandlikar, G.S.; Gold, Z.J.; Cowen, M.C.; Meyer, R.S.; Freise, A.C.; Kraft, N.J.B.; Moberg-Parker, J.; Sprague, J.; Kushner, D.J.; Curd, E.E. ranacapa: An R package and Shiny web app to explore environmental DNA data with exploratory statistics and interactive visualizations. F1000Research 2018, 7, 1734. [Google Scholar] [CrossRef]
- Jelihovschi, E.; Faria, J.C.; Allaman, I.B.; Allaman, M.I.B. ScottKnott: A package for performing the Scott-Knott clustering algorithm in R. TEMA 2014, 15, 3–17. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: For prediction of metagenome functions. Nat Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Codes | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S5S | V2S | B14S | A11S | A12S | B13S | V1S | S6S | S7S | S8S | V4S | V3S | B16S | B15S | A9S | A10S | |
| Organic C (g/kg) | 0.9 | 0.7 | 0.8 | 1 | 1.1 | 1 | 0.8 | 0.9 | 0.8 | 0.8 | 1 | 0.7 | 0.9 | 1 | 1 | 1.1 |
| Organic matter (g/kg) | 1.6 | 1.3 | 1.4 | 1.7 | 1.8 | 1.7 | 1.4 | 1.6 | 1.4 | 1.4 | 1.7 | 1.2 | 1.5 | 1.7 | 1.8 | 1.9 |
| Total N (g/kg) | 1.1 | 1 | 1.1 | 1.1 | 1.2 | 1.2 | 1 | 1.1 | 1.3 | 1.1 | 1.2 | 1 | 1 | 1.2 | 1.1 | 1.2 |
| Nitric N (mg/kg) | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 6 | 4 | 4 | 4 |
| Ammoniacal N (mg/kg) | 6 | 2 | 4 | 3 | 6.4 | 5 | 5 | 6.5 | 4 | 4 | 5 | 3 | 5.6 | 6 | 4 | 4 |
| Assimilable P (mg/kg) | 5 | 4 | 4 | 5 | 6 | 4 | 4 | 9 | 4 | 4 | 4 | 11 | 7 | 4 | 5 | 6 |
| Exc. P (as K2O) (mg/kg) | 569 | 316 | 321 | 381 | 407 | 378 | 313 | 487 | 378 | 376 | 607 | 362 | 559 | 436 | 591 | 607 |
| Exc. P (mg/kg) | 236 | 131 | 133 | 158 | 169 | 157 | 130 | 202 | 157 | 156 | 252 | 150 | 232 | 181 | 245 | 252 |
| Total limestone (g/kg) | 12 | 15 | 15 | 12 | 9 | 13 | 12 | 13 | 12 | 11 | 11 | 15 | 18 | 13 | 14 | 15 |
| ExcCa (mg/kg) | 3497 | 2768 | 2746 | 3279 | 3538 | 2798 | 2640 | 2962 | 2778 | 2594 | 3140 | 3111 | 2995 | 2918 | 3095 | 3116 |
| Exc. Ca (as CaO) (mg/kg) | 4895.8 | 3875.2 | 3844.4 | 4590.6 | 4953.2 | 3917.2 | 3696 | 4146.8 | 3889.2 | 3631.6 | 4396 | 4355.4 | 4193 | 4085.2 | 4333 | 4362.4 |
| Exc. Mg (mg/kg) | 153 | 96 | 84 | 119 | 114 | 106 | 88 | 112 | 104 | 94 | 147 | 75 | 155 | 134 | 163 | 149 |
| Exc. Mg (as MgO) (mg/kg) | 255 | 160 | 140 | 198 | 190 | 177 | 147 | 187 | 173 | 157 | 245 | 125 | 258 | 223 | 272 | 248 |
| Exc. Na (mg/kg) | 18 | 13 | 16 | 13 | 17 | 15 | 15 | 19 | 19 | 19 | 18 | 16 | 20 | 17 | 21 | 19 |
| Assimilable Fe (mg/kg) | 12.5 | 11 | 11 | 11 | 9.1 | 12 | 12 | 9.3 | 11 | 10 | 12 | 10 | 8.5 | 10 | 11 | 11 |
| Assimilable Zn (mg/kg) | 0.4 | 0.3 | 0.4 | 0.4 | 0.3 | 0.4 | 0.3 | 0.6 | 0.5 | 0.4 | 0.5 | 1 | 0.5 | 0.6 | 0.5 | 0.8 |
| Assimilable Se (mg/kg) | 0.3 | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.3 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 |
| pH in water | 8.2 | 8.2 | 8.2 | 8.2 | 8.2 | 8.1 | 8 | 8.2 | 8.2 | 8.2 | 8.1 | 8.2 | 8.2 | 8.1 | 8.3 | 8.3 |
| C.E.C. pH 8.2 (meq/100 g) | 19 | 15 | 15 | 18 | 19 | 15 | 14 | 16 | 15 | 14 | 18 | 17 | 17 | 16 | 18 | 18 |
| Variety | Code | Nitrogen Fertilization (N kg ha−1 yr−1) | Phosphorous Fertilization (P2O5 kg ha−1 yr−1) | Latitude (N) | Longitude (E) |
|---|---|---|---|---|---|
| Verna | V1S | 40 | 80 | 43.051129° | 11.690833° |
| V1R | 40 | 80 | 43.051129° | 11.690833° | |
| V2S | 80 | 160 | 43.050959° | 11.691126° | |
| V2R | 80 | 160 | 43.050959° | 11.691126° | |
| V3S | 80 | 80 | 43.050755° | 11.691438° | |
| V3R | 80 | 80 | 43.050755° | 11.691438° | |
| V4S | 40 | 40 | 43.050547° | 11.691890° | |
| V4R | 40 | 40 | 43.050547° | 11.691890° | |
| Sieve | S5S | 40 | 40 | 43.050953° | 11.692320° |
| S5R | 40 | 40 | 43.050953° | 11.692320° | |
| S6S | 80 | 80 | 43.051126° | 11.691964° | |
| S6R | 80 | 80 | 43.051126° | 11.691964° | |
| S7S | 80 | 160 | 43.051343° | 11.691626° | |
| S7R | 80 | 160 | 43.051343° | 11.691626° | |
| S8S | 40 | 80 | 43.051525° | 11.691219° | |
| S8R | 40 | 80 | 43.051525° | 11.691219° | |
| Andriolo | A9S | 40 | 40 | 43.049740° | 11.690994° |
| A9R | 40 | 40 | 43.049740° | 11.690994° | |
| A10S | 80 | 80 | 43.049901° | 11.690666° | |
| A10R | 80 | 80 | 43.049901° | 11.690666° | |
| A11S | 80 | 160 | 43.050036° | 11.690376° | |
| A11R | 80 | 160 | 43.050036° | 11.690376° | |
| A12S | 40 | 80 | 43.050180° | 11.690020° | |
| A12R | 40 | 80 | 43.050180° | 11.690020° | |
| Bologna | B13S | 40 | 80 | 43.050763° | 11.690400° |
| B13R | 40 | 80 | 43.050763° | 11.690400° | |
| B14S | 80 | 160 | 43.050531° | 11.690761° | |
| B14R | 80 | 160 | 43.050531° | 11.690761° | |
| B15S | 80 | 80 | 43.050347° | 11.691104° | |
| B15R | 80 | 80 | 43.050347° | 11.691104° | |
| B16S | 40 | 40 | 43.050193° | 11.691440° | |
| B16R | 40 | 40 | 43.050193° | 11.691440° |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cangioli, L.; Mancini, M.; Napoli, M.; Fagorzi, C.; Orlandini, S.; Vaccaro, F.; Mengoni, A. Differential Response of Wheat Rhizosphere Bacterial Community to Plant Variety and Fertilization. Int. J. Mol. Sci. 2022, 23, 3616. https://doi.org/10.3390/ijms23073616
Cangioli L, Mancini M, Napoli M, Fagorzi C, Orlandini S, Vaccaro F, Mengoni A. Differential Response of Wheat Rhizosphere Bacterial Community to Plant Variety and Fertilization. International Journal of Molecular Sciences. 2022; 23(7):3616. https://doi.org/10.3390/ijms23073616
Chicago/Turabian StyleCangioli, Lisa, Marco Mancini, Marco Napoli, Camilla Fagorzi, Simone Orlandini, Francesca Vaccaro, and Alessio Mengoni. 2022. "Differential Response of Wheat Rhizosphere Bacterial Community to Plant Variety and Fertilization" International Journal of Molecular Sciences 23, no. 7: 3616. https://doi.org/10.3390/ijms23073616
APA StyleCangioli, L., Mancini, M., Napoli, M., Fagorzi, C., Orlandini, S., Vaccaro, F., & Mengoni, A. (2022). Differential Response of Wheat Rhizosphere Bacterial Community to Plant Variety and Fertilization. International Journal of Molecular Sciences, 23(7), 3616. https://doi.org/10.3390/ijms23073616